

Abstract
Plasminogen is the zymogen of plasmin, the major enzyme that degrades fibrin clots. In addition to its binding and activation on fibrin clots, plasminogen also specifically interacts with cell surfaces where it is more efficiently activated by plasminogen activators, compared with the reaction in solution. This results in association of the broad spectrum proteolytic activity of plasmin with cell surfaces that functions to promote cell migration. Here we review emerging data establishing a role for plasminogen, plasminogen receptors and the newly discovered plasminogen receptor, Plg-RKT, in macrophage recruitment in the inflammatory response and we address mechanisms by which the interplay between plasminogen and its receptors regulates inflammation.
Keywords: Macrophages, monocytes, plasminogen, plasminogen receptors, Plg-RKT, thioglycollate, tissue plasminogen activator, urokinase
1. Introduction
Plasminogen is the circulating zymogen of the enzyme, plasmin, the major enzyme responsible for degradation of fibrin clots (Bugge et al., 1995; Ploplis et al., 1995b). Over twenty-five years ago it was first recognized that plasminogen also specifically interacts with the surfaces of cells. This interaction is analogous to the interaction of plasminogen with fibrin in that plasminogen bound to cells is more efficiently activated by plasminogen activators compared with plasminogen in solution (Miles & Plow, 1985; Stricker et al., 1986; Hajjar et al., 1986; Loscalzo & Vaughan, 1987; Ellis et al., 1991; Gonias et al., 1989; Duval-Jobe & Parmely, 1994; Félez et al., 1996; Longstaff et al., 1999; Sinniger et al., 1999). Furthermore, plasmin remains associated with the cell surface where it is relatively protected from inactivation by α2-antiplasmin (Plow et al., 1986; Hall et al., 1991). This results in association of the broad spectrum proteolytic activity of plasmin with the cell surface leading to a wide array of physiological and pathological sequelae. Physiologic and pathophysiologic processes with plasmin-dependent cell migration as a central feature include inflammation (Ploplis et al., 1998; Plow et al., 1999; Busuttil et al., 2004), wound healing (Romer et al., 1996; Creemers et al., 2000), oncogenesis (Madureira et al., 2011a; Madureira et al., 2012), metastasis (Ranson et al., 1998; Palumbo et al., 2003), myogenesis, and muscle regeneration (Suelves et al., 2002; Lopez-Alemany et al., 2003; Lopez-Alemany et al., 2005). Cell surface plasmin also participates in neurite outgrowth (Jacovina et al., 2001; Gutierrez-Fernandez et al., 2009), and prohormone processing (Parmer et al., 2000; Jiang et al., 2001; Jiang et al., 2002; Bai et al., 2012). Furthermore, plasmin-induced cell signaling also affects the functions of inflammatory cells, via production of cytokines, reactive oxygen species, and other mediators (Syrovets et al., 2012; Li et al., 2012). Finally, plasminogen receptors regulate fibrinolysis (Ling et al., 2004; Dassah et al., 2009; Surette et al., 2011; Madureira et al., 2011b; Pluskota et al., 2004).
Plasminogen receptors are very broadly distributed on both prokaryotic and eukaryotic cells, including monocytes (Miles & Plow, 1987), monocytoid cells (Plow, Freaney, Plescia & Miles, 1986), macrophages (Silverstein et al., 1988), endothelial cells (Hajjar, Harpel, Jaffe & Nachman, 1986; Miles et al., 1988b), fibroblasts (Gonzalez-Gronow et al., 1994), platelets (Miles & Plow, 1985), adrenal medullary cells (Parmer, Mahata, Gong, Mahata, Jiang, O’Connor, Xi & Miles, 2000; Jiang, Yasothornsrikul, Taupenot, Miles & Parmer, 2002) and carcinoma cells (Durliat et al., 1992; Hembrough et al., 1995) as well as on cell-derived microvesicles (Lacroix et al., 2007; Lacroix et al., 2012). The structure and function of plasminogen receptors on prokaryotic cells have been comprehensively reviewed recently (Bhattacharya et al., 2012; Sanderson-Smith et al., 2012). Here, we focus on the structure and function of eukaryotic plasminogen receptors, with special emphasis on the novel plasminogen receptor, Plg-RKT (Andronicos et al., 2010) and the role of plasminogen receptors in macrophage migration.
2. The Plasminogen Activation System
Initiation of plasminogen activation results in generation of the broad spectrum serine protease, plasmin, from the circulating zymogen, plasminogen (Figure 1). Plasminogen is activated to plasmin by plasminogen activators (PA’s), via specific proteolytic cleavage (Castellino & Ploplis, 2005). Plasmin is the major enzyme responsible for degradation of fibrin clots (fibrinolysis) to maintain normal blood homeostasis (Ploplis, Carmeliet, Vazirzadeh, Van, I, Moons, Plow & Collen, 1995b; Bugge, Flick, Daugherty & Degen, 1995). Dysregulation of the plasminogen activation system can result in hemorrhage (excess fibrinolysis) or thrombosis (insufficient fibrinolysis). The plasminogen activation system is regulated by direct inhibition of plasmin (by the circulating serpin, α2-antiplasmin) and by synthesis and secretion of plasminogen activators and the serpin, plasminogen activator inhibitor 1 (PAI-1) (Collen, 1999). In a key regulatory step, plasminogen activation is promoted when plasminogen and its activator, tPA, bind concomitantly to lysine residues on the surface of fibrin clots, resulting in a marked reduction in the Km for activation of plasminogen compared with the reaction in solution (Hoylaerts et al., 1982).
Figure 1. The Plasminogen Activation System.

Initiation of plasminogen activation results in generation of the broad spectrum serine protease, plasmin, from the circulating zymogen, plasminogen. Plasminogen is activated to plasmin by plasminogen activators (PA’s), via specific proteolytic cleavage. The plasminogen activation system is regulated by direct inhibition of plasmin (by the circulating serpin, a2-antiplasmin) and by synthesis and secretion of plasminogen activators and the serpin, plasminogen activator inhibitor 1 (PAI-1).
2.1 Plasminogen
2.1.1 Initial Studies on Plasminogen
In 1933, Tillet and Garner made the first observation that cultures of β-hemolytic streptococci had the ability to lyse human fibrin clots and named the process fibrinolysis (Tillett & Garner, 1933). The active agent in the cultures was termed “fibrinolysin”. (The cultures were derived from patients suffering from various manifestations of acute streptococcal disease, including septicemia, erysipelas, scarlet fever, acute tonsillitis, and cellulitis.) Subsequently, in 1938, Milstone observed that fibrinolysis by β-hemolytic streptococci cultures requires a factor found in normal human blood, termed “lytic factor” (Milstone, 1941). Initially, the inactive form of this proteolytic enzyme was named serum tryptogen (Ferguson, 1943) and several reports had considered that serum tryptogen and trypsin might be identical. It was subsequently demonstrated that serum tryptogen and trypsin were not identical based on separate specificities of their activators, fibrinolysin and enterokinase, respectively, as well antibody neutralization studies that distinguished these enzymes (Kaplan, 1946). The terms that emphasize the plasma source of serum tryptogen proteolytic activity were subsequently adopted, plasmin and plasminogen (Christensen, 1945). Plasminogen was purified from human serum in 1949 (REMMERT & COHEN, 1949). Subsequently, the term streptokinase (SK) was substituted for fibrinolysin.
2.1.2 Plasminogen Structure
Human plasminogen is synthesized as an 810-amino acid protein, from which a 19 amino acid leader peptide is cleaved during secretion to yield the mature protein (Forsgren et al., 1987) of 791 amino acids with N-terminal glutamic acid (Figure 2). The only identified posttranslational modifications are N and O-linked glycosylations and phosphorylations (Wang et al., 1997a). The glycosylation sites are at Asn289 in approximately half of plasminogen molecules and an O-linked glycan at Thr346 in all plasminogen molecules (Hayes & Castellino, 1979a; Hayes & Castellino, 1979b). The two types, Plasminogen I and II, respectively, are named based on which type elutes first on lysine sepharose. Thus, Type II is glycosylated at one site and Type I is glycosylated at two sites. The differential glycoslation may affect the properties of plasminogen (Gonzalez-Gronow et al., 1990). The phosphorylation site is at Ser578 (Wang et al., 1997b).
Figure 2. Primary Structure of Human Plasminogen.

Human plasminogen is synthesized as an 810-amino acid protein, from which a 19 amino acid leader peptide is cleaved during secretion to yield the mature protein. Glycosylation sites are at Asn289 in approximately half of plasminogen molecules and an O-linked glycan at Thr346 in all plasminogen molecules. Cleavage by plasminogen activators occurs at Arg561–Val562 and results in the conversion of single chain plasminogen to the two-chain disulfide-linked serine protease, plasmin. The active site of plasmin is the catalytic triad His603/Asp646/Ser741(*). Plasmin-mediated cleavage of Glu-plasminogen at position Lys77–Lys78 results in conversion to Lys-Plasminogen and the release of the pan-Apple domain. Reprinted from (Ploplis, 2001) PMID:11229877, according to Frontiers in Bioscience policies.
The mature form of plasminogen is composed of seven structural modules which include, an N-terminal Pan-apple domain (a hair-pin loop structure with a characteristic pattern of 4 cysteines (Tordai et al., 1999) and 5-triple disulfide-linked domains termed kringles and named for their resemblance to a Danish pastry (Sottrup-Jensen et al., 1978). Four of the five kringles are able to bind lysine and its analogs. The kringles mediate the interaction of plasminogen with a number of key biological molecules that regulate plasmino(ogen) function (Castellino & Ploplis, 2005). Consequently, lysine and lysine analogs interfere with these interactions and are used clinically to inhibit the functional consequences of such interactions when necessary (McCormack, 2012). The seventh domain in plasminogen consists of the latent active site (Castellino & Ploplis, 2005).
2.1.3 Sites of Plasminogen Synthesis
Plasminogen circulates at high concentration (2 μM) (Collen & Verstraete, 1975) and liver is well known to be the major site of synthesis (Bohmfalk & Fuller, 1980; Saito et al., 1980). Plasminogen is also expressed at low levels extrahepatically in a number of tissues including brain (Tsirka et al., 1997; Basham & Seeds, 2001; Sappino et al., 1993; Zhang et al., 2002a), adrenal, kidney, testis, heart, lung, uterus, spleen, thymus, gut (Zhang et al., 2002b) and cornea (Twining et al., 1999). In addition, monocytes synthesize low levels of plasminogen (Zhang et al., 2003). Endogenous monocyte plasmin(ogen) may be particularly important when monocytoid cells migrate through tissues in plasminogen-poor, avascular environments.
2.1.4 Plasminogen Deficiency
Inherited severe hypoplasminogenemia (type I plasminogen deficiency), with markedly decreased plasminogen antigen (<5%–57% activity), is a rare systemic disease characterized by markedly impaired fibrinolysis that was first recognized in 1997 (Mingers et al., 1997). The impaired fibrinolysis leads to the formation of ligneous (wood-like) fibrin-rich pseudomembranes on mucosae during wound healing (Schuster et al., 2007). The most common clinical manifestation is an inflammation of the conjunctivae (ligneous conjunctivitis). The second most common manefestation of mucosal involvement in these patients is ligneous periodontitis and gingivitis. Ligneous lesions in the oral cavity appear as massive, painless ulcerated gingival enlargements covered with yellowish pseudomembranes without any tendency to bleed (Kurtulus et al., 2011). Other sites of mucosal involvement include the upper and lower gastrointestinal tracts, respiratory tract and these can lead to life threatening pseudomembraneous obstructions. In addition, membranes within the female genital tract are affected. Of a total of 102 patients (including 62 women) analyzed by 2011, two of the women had given birth to healthy children (by Caesarean section) (Klammt et al., 2011). In all other cases reported to date females with hypoplasminogenemia are infertile (Klammt, Kobelt, Aktas, Durak, Gokbuget, Hughes, Irkec, Kurtulus, Lapi, Mechoulam, Mendoza-Londono, Palumbo, Steitzer, Tabbara, Ozbek, Pucci, Sotomayor, Sturm, Drogies, Ziegler & Schuster, 2011). The central nervous system may also be affected in hypoplasminogenemia and occlusive hydrocephalus is observed in some patients.
Within the group of 102 hypoplasminogenemic patients studied, both homozygous and compound heterozygous plasminogen mutations with plasminogen functional activity from <5%–57% were identified within this group. Missense, splice site (premature stop codons) and frame shift mutations were present. It is interesting that patients that were more severely affected did not always present with the lowest plasminogen activity levels (Klammt, Kobelt, Aktas, Durak, Gokbuget, Hughes, Irkec, Kurtulus, Lapi, Mechoulam, Mendoza-Londono, Palumbo, Steitzer, Tabbara, Ozbek, Pucci, Sotomayor, Sturm, Drogies, Ziegler & Schuster, 2011). Even intra-familial variability was noted. Thus, Klammt and co-authors suggested that the penetrance of hypoplasminogenemia may be due to modifier genes or epigenetic factors that have not yet been identified (Klammt, Kobelt, Aktas, Durak, Gokbuget, Hughes, Irkec, Kurtulus, Lapi, Mechoulam, Mendoza-Londono, Palumbo, Steitzer, Tabbara, Ozbek, Pucci, Sotomayor, Sturm, Drogies, Ziegler & Schuster, 2011). Application of plasminogen eye drops or topical/systemic fresh frozen plasma as well as treatment with sex hormones may be effective in treating ligneous conjunctivitis. With severe disease with multi-organ involvement, treatment with plasminogen, corticosteroids and sex hormones may be partly efficient as well as thrombin inhibitors (Klammt, Kobelt, Aktas, Durak, Gokbuget, Hughes, Irkec, Kurtulus, Lapi, Mechoulam, Mendoza-Londono, Palumbo, Steitzer, Tabbara, Ozbek, Pucci, Sotomayor, Sturm, Drogies, Ziegler & Schuster, 2011).
Patients with (type II plasminogen deficiency) with reduced plasminogen activity, but normal or only slightly reduced plasminogen antigen levels have never been reported to develop pseudomembranous lesions (Schuster & Seregard, 2003).
Plasminogen deficient mice were independently generated by two groups in 1995 (Bugge, Flick, Daugherty & Degen, 1995; Ploplis, Carmeliet, Vazirzadeh, Van, I, Moons, Plow & Collen, 1995b) and the spontaneous phenotypes appear more severe than in hypoplasminogenemic humans. This difference may relate to the complete absence of plasminogen in the plasminogen knockout mice (whereas hypoplasminogenemic humans do have some residual plasminogen) as well as to differences between species. In addition, some phenotypes observed in mice are not able to be evaluated in living humans.
The phenotype of the plasminogen knockout mice recapitulates several of the spontaneous phenotypes observed in plasminogen deficient humans including ligneous conjunctivitis (Drew et al., 1998), chronic otitis media (Eriksson et al., 2006), periodontitis (Sulniute et al., 2011), and reduced wound healing (Romer, Bugge, Pyke, Lund, Flick, Degen & Dano, 1996; Bugge et al., 1996) while hydrocephalus is observed only rarely (Drew, Kaufman, Kombrinck, Danton, Daugherty, Degen & Bugge, 1998). Plasminogen deficient female mice exhibit reduced fertility (Ploplis et al., 1995a). Additional clinical manifestations in the knockout mice include fibrin-rich lesions in the liver, the urogenital tract, the respiratory tract, trachea, larynx, and gastrointestinal tract, ulcers of the colon and rectum, and the female genital tract and rectal and vaginal prolapse (Bugge, Flick, Daugherty & Degen, 1995; Ploplis, Carmeliet, Vazirzadeh, Van, I, Moons, Plow & Collen, 1995b; Bugge, Kombrinck, Flick, Daugherty, Danton & Degen, 1996), as well as compromised mammary development, involution and defects in lactation (Lund et al., 2000). Plasminogen deficient mice exhibit retarded postnatal growth after weaning (Ploplis, Carmeliet, Vazirzadeh, Van, I, Moons, Plow & Collen, 1995b; Hoover-Plow et al., 1999) and develop a severe wasting syndrome (Bugge, Flick, Daugherty & Degen, 1995). Mortality is high (Bugge, Flick, Daugherty & Degen, 1995).
The preponderance of fibrin deposition in organs, combined with the presence of fibrin-rich pseudomembranous lesions in the plasminogen deficient mice raised the issue of whether the spontaneous phenotypes can be ascribed to the lack of ability of these mice to resolve fibrin clots. In an elegant paper, Thomas Bugge and collaborators showed that removal of fibrinogen alleviated all of the spontaneous pathologies associated with plasminogen deficiency (Bugge, Kombrinck, Flick, Daugherty, Danton & Degen, 1996). Current studies are addressing the role of fibrin degradation when plasminogen deficient mice are challenged in various models.
2.2 Plasminogen Activation
Plasminogen is activated by several plasminogen activators (PA’s), including tissue plasminogen activator (tPA) (Wilson et al., 1980) and urokinase plasminogen activator (uPA) (Astrup & STERNDORFF, 1952), activated Factor XII, activated Factor XI and kallikrein of the con intrinsic coagulation system (contact activation system): FXIa, FXIIa and kallikrein (Colman, 1969; Miles et al., 1983; Schousboe et al., 1999), as well as bacterial plasminogen activators, represented by SK. The major physiologic plasminogen activators are tPA and uPA (Carmeliet et al., 1994). All of the plasminogen activators cleave plasminogen at the same Arg 561-Val 562 bond within the cleavage loop of plasminogen (Robbins et al., 1967). Cleavage results in the formation of plasmin consisting of the N-terminal heavy chain and a carboxyl-terminal-derived light chain, linked by two disulfide bonds. The serine protease catalytic triad is composed of His 603, asp 646 and Ser 741.
Different plasminogen activators exhibit different mechanisms of plasminogen activation. uPA is secreted as a single chain zymogen, single chain uPA (scuPA) from a variety of cell types including monocytes/macrophages (Unkeless et al., 1974; Vassalli et al., 1984), trophoblasts (Strickland et al., 1976; Sappino et al., 1989)and epithelial cells, particularly in the kidney (Sappino et al., 1991). The general mechanism of scuPA mediated activation of Glu-plasminogen consists of an initial low rate of conversion of Glu-plasminogen to Glu-plasmin by low levels of endogenous activity in scuPA, followed by feedback activation of scuPA to two chain uPA (uPA) (Lijnen et al., 1990; Fleury et al., 1993). On certain cell types activation of scuPA to uPA may be accelerated initially by matriptase (Kilpatrick et al., 2006). uPA is a more efficient plasminogen activator than scuPA, so that the overall activation of plasminogen becomes accelerated. Plasminogen activation by uPA is markedly enhanced when urokinase is bound to its cell surface receptor, uPAR (Ellis, Behrendt & Dano, 1991), and when plasminogen is concomitantly bound to the cell surface (Félez, Miles, Fábregas, Jardi, Plow & Lijnen, 1996).
Plasminogen activation by tPA does not take place efficiently in solution, but occurs efficiently on the fibrin surface. tPA has a high affinity for fibrin and fibrin-mediated enhanced plasminogen activation occurs when both tPA and plasminogen bind to fibrin, resulting in a large decrease in the Km for the reaction (Hoylaerts, Rijken, Lijnen & Collen, 1982; Rijken et al., 1982). tPA is secreted from endothelial cells (Todd, 1959) and neuroendocrine cells [reviewed in (Parmer & Miles, 1998)] as a single chain molecule and cleaved to a two chain form by plasmin. Single chain and two chain tPA have similar activities and it is thought that the mechanism involves single chain tPA adopting a two chain tPA conformation, once bound to fibrin (Loscalzo, 1988). Partially digested fibrin is more effective than undigested fibrin, most likely due to exposure of C-terminal lysines on fibrin that has been partially digested by plasmin (Higgins & Vehar, 1987).
The first plasminogen activator identified, SK, exhibits unusual properties with regard to plasminogen activation because it functions in a nonhydrolytic manner. The activation of human plasminogen by SK, involves the formation of a stoichiometric complex of SK with human plasminogen (SK-HPg), which induces the formation of an active site within human plasminogen within the complex to yield SK-HPg* (Shick and Castellino, 1974). SK-HPg* and SK complexed with plasmin (SK-HPm) are all effective activators of human plasminogen with SK-HPg′ being the most efficient (Davidson et al., 1990). After this, activation of plasminogen by SK proceeds as for activation by tPA and uPA. Activation of plasminogen by another bacterial plasminogen activator, staphylokinase (STAR) is similar to that of SK, but a major difference is that activation is enhanced in the presence of fibrin (Sakai et al., 1989) and endothelial cells (Okada et al., 2000).
Plasminogen may also be activated by factors of the intrinsic coagulation system: FXIa, FXIIa and kallikrein (Colman, 1969; Miles, Greengard & Griffin, 1983; Schousboe, Feddersen & Rojkjaer, 1999).
2.2.1 Conformational Regulation of Plasminogen Activation
Circulating plasminogen has an N-terminal glutamic acid (Glu-plasminogen). Glu-plasminogen circulates in a closed activation resistant form in the presence of physiologic concentrations of chloride ions (Violand et al., 1975; Violand et al., 1978; Urano et al., 1987a; Urano et al., 1987b; Urano et al., 1988), as a mechanism to prevent undesired activation. In the presence of lysine and lysine analogs such as ε-aminocaproic acid (EACA), the conformation of Glu-plasminogen is switched to a more readily activatable relaxed state. The effect of lysine analogs is consistent with the concept that the plasminogen kringles regulate this conformational change, since 4 of the 5 plasminogen kringles bind plasminogen (Sehl & Castellino, 1990; Mehart et al., 1991; Menhart et al., 1993). A number of peptide sequences within the Pan-apple domain that interact with kringle 1, kringle 4 and kringle 5 to stabilize the tight conformation have been identified (Horrevoets et al., 1995). In a major recent breakthrough the X-ray crystal structure of full length human Type II Glu-plasminogen was solved independently by two groups (Law et al., 2012; Xue et al., 2012) and their results have provided major insights into the interactions governing maintenance of the closed conformation of Glu-plasminogen as well as other aspects of plasminogen function. The seven domains of plasminogen are loosely clustered in a diamond-shaped zig zag assembly and the Pan-apple domain makes extensive contacts with kringle 4 and kringle 5 (Figure 3). Furthermore, a single chloride ion is bound at the interface of the kringle 4 and Pan-apple domains and the latent serine protease domain/kringle 2 interface, consistent with the key role of chloride ion in stabilizing closed plasminogen ions (Violand, Sodetz & Castellino, 1975; Violand, Byrne & Castellino, 1978; Urano, Chibber & Castellino, 1987a; Urano, de Serrano, Chibber & Castellino, 1987b; Urano, de Serrano, Gaffney & Castellino, 1988).
Figure 3. The Structure of Human Type II Glu-Plasminogen.

Plasminogen domains are labeled and colored as follows: Pan-apple domain, blue; kringle 1, pink; kringle 2, yellow; kringle 3, orange, kringle 4, green; kringle 5, purple; latent serine protease domain, cyan. Chloride ions Cl(1) and Cl(2) are at the kringle 4/Pan apple domain and kringle 2/serine protease domain interfaces, respectively, and are shown as spheres. Two other chloride ions, Cl(3) and Cl(4), bind to kringle 2 and the latent serine protease domain, respectively. The position of the activation loop (R561) is marked with a red sphere. The lysine binding site of kringle 1 is marked with an asterisk. Reprinted from (Law et al., 2012) with permission from Elsevier.
Another mechanism by which Glu-plasminogen can be converted to a relaxed conformation is by proteolytic cleavage. Proteolytic cleavage by plasmin (at several sites within the amino terminus of Glu-plasminogen) results in loss of a 77–79 amino acid peptide that includes the Pan-apple domain. The resulting plasminogen forms are collectively termed Lys-plasminogen as this is the predominant N-terminal amino acid present following cleavage by plasmin. Thus, the interactions of kringle 4 and kringle 5 with the Pan-apple domain are disrupted, allowing Lys plasminogen to adopt a more open relaxed form that is more activatable. Thus, Lys-plasminogen is insensitive to the effects of chloride ion or lysine analogs that regulate the closed conformation of Glu-plasminogen (Violand, Sodetz & Castellino, 1975).
The crystal structure of Glu-plasminogen also reveals an additional mechanism by which the activation loop is protected from undesirable activation. The activation sequence is shielded by the position of the kringle 3/kringle 4 linker region while the O-linked glycan on Thr 346 (present in all plasminogen glycoforms) serves to partially shield the R561/Val562 bond and forms a polar interaction with the sidechain of R561 (Law, Caradoc-Davies, Cowieson, Horvath, Quek, Encarnacao, Steer, Cowan, Zhang, Lu, Pike, Smith, Coughlin & Whisstock, 2012; Xue, Bodin & Olsson, 2012)
3. General Characteristics of Plasminogen Receptors
3.1 Functional Consequences of Plasminogen Binding to Cells
Over twenty-five years ago it was first recognized that plasminogen specifically interacts with the surfaces of cells. This interaction is analogous to the interaction of plasminogen with fibrin in that plasminogen bound to cells is more efficiently activated by plasminogen activators compared with plasminogen in solution (Miles & Plow, 1985; Stricker, Wong, Tak Shiu, Reyes & Shuman, 1986; Hajjar, Harpel, Jaffe & Nachman, 1986; Loscalzo & Vaughan, 1987; Ellis, Behrendt & Dano, 1991; Gonias, Braud, Geary & VandenBerg, 1989; Duval-Jobe & Parmely, 1994; Félez, Miles, Fábregas, Jardi, Plow & Lijnen, 1996; Longstaff, Merton, Fabregas & Felez, 1999; Sinniger, Merton, Fabregas, Felez & Longstaff, 1999) (Figure 4). Furthermore, plasmin remains associated with the cell surface where it is relatively protected from inactivation by α2-antiplasmin (Plow, Freaney, Plescia & Miles, 1986; Hall, Humphries & Gonias, 1991). This results in association of the broad spectrum proteolytic activity of plasmin with the cell surface leading to a wide array of physiological and pathological sequelae.
Figure 4. Enhancement of plasminogen activation on the cell surface.

Activation of cell-associated plasminogen (Plg) to plasmin (Pm) by cell-associated plasminogen activators (PA) is markedly enhanced compared to the reaction in solution. The Pm formed remains on the cell surface where it is relatively protected from its inhibitor, α2-antiplasmin (α2-AP). This results in association of the broad spectrum proteolytic activity of plasmin with the cell surface leading to a wide array of physiological and pathological sequelae. This figure was originally published in Proteomics/Book 1: Human Diseases and Protein Functions, Miles, L.A., Andronicos, N.M., Chen, E.I., Baik, N., Bai, H., Parmer, C.M., Lighvani, S., Nangia, S., Kiosses, W.B., Kamps, M.P., Yates III, J.R. and Parmer, R.J. Identification of the novel plasminogen receptor, Plg-RKT. In: Proteomics/Book 1: Human Diseases and Protein Functions, ISBN 978-953-307-832-8, Man, T.K. and Flores, R.J.(Ed.), Intech, Chapter 10: 219–238, 2012.
Plasminogen receptors are very broadly distributed on both prokaryotic and eukaryotic cells, including monocytes (Miles & Plow, 1987), monocytoid cells (Plow, Freaney, Plescia & Miles, 1986), macrophages (Silverstein, Friedlander, Jr., Nicholas & Nachman, 1988), endothelial cells (Hajjar, Harpel, Jaffe & Nachman, 1986; Miles, Levin, Plescia, Collen & Plow, 1988b), fibroblasts (Gonzalez-Gronow, Gawdi & Pizzo, 1994), platelets (Miles & Plow, 1985), adrenal medullary cells (Parmer, Mahata, Gong, Mahata, Jiang, O’Connor, Xi & Miles, 2000; Jiang, Yasothornsrikul, Taupenot, Miles & Parmer, 2002) and carcinoma cells (Durliat, Komano, Correc, Bertrand, Cochet, Caignard, Martin & Burtin, 1992; Hembrough, Vasudevan, Allietta, Glass & Gonias, 1995) as well as on cell-derived microvesicles (Lacroix, Sabatier, Mialhe, Basire, Pannell, Borghi, Robert, Lamy, Plawinski, Camoin-Jau, Gurewich, Angles-Cano & Dignat-George, 2007; Lacroix, Plawinski, Robert, Doeuvre, Sabatier, Martinez, Mezzapesa, Anfosso, Leroyer, Poullin, Jourde, Njock, Boulanger, Angles-Cano & Dignat-George, 2012). Several distinct plasminogen receptors (see section 3.4 below) have been identified over the past decades, consistent with the high number of receptors determined/cell [ranging from 37,000/platelet (Miles & Plow, 1985) to > 107 sites/endothelial cell (Miles, Levin, Plescia, Collen & Plow, 1988b)].
When cells become armed with the broad-spectrum activity of plasmin, they acquire the ability to degrade extracellular matrices and activate matrix-associated growth factors and proteolytic enzymes that facilitate cell migration. Physiologic and pathophysiologic processes with plasmin-dependent cell migration as a central feature include inflammation (Ploplis, French, Carmeliet, Collen & Plow, 1998; Plow, Ploplis, Busuttil, Carmeliet & Collen, 1999; Busuttil, Ploplis, Castellino, Tang, Eaton & Plow, 2004), wound healing (Romer, Bugge, Pyke, Lund, Flick, Degen & Dano, 1996; Creemers, Cleutjens, Smits, Heymans, Moons, Collen, Daemen & Carmeliet, 2000), oncogenesis (Madureira, Hill, Miller, Giacomantonio, Lee & Waisman, 2011a; Madureira, O’Connell, Surette, Miller & Waisman, 2012), metastasis (Ranson, Andronicos, O’Mullane & Baker, 1998; Palumbo, Talmage, Liu, La Jeunesse, Witte & Degen, 2003), myogenesis, and muscle regeneration (Suelves, Lopez-Alemany, Lluis, Aniorte, Serrano, Parra, Carmeliet & Munoz-Canoves, 2002; Lopez-Alemany, Suelves & Munoz-Canoves, 2003; Lopez-Alemany, Suelves, Diaz-Ramos, Vidal & Munoz-Canoves, 2005). Cell surface plasmin also participates in neurite outgrowth (Jacovina, Zhong, Khazanova, Lev, Deora & Hajjar, 2001; Gutierrez-Fernandez, Gingles, Bai, Castellino, Parmer & Miles, 2009), and prohormone processing (Parmer, Mahata, Gong, Mahata, Jiang, O’Connor, Xi & Miles, 2000; Jiang, Taupenot, Mahata, Mahata, O’Connor, Miles & Parmer, 2001; Jiang, Yasothornsrikul, Taupenot, Miles & Parmer, 2002; Bai, Nangia & Parmer, 2012). Furthermore, plasmin-induced cell signaling also affects the functions of inflammatory cells, via production of cytokines, reactive oxygen species, and other mediators (Syrovets, Lunov & Simmet, 2012; Li, Syrovets & Simmet, 2012). Finally, plasminogen receptors regulate fibrinolysis (Ling, Jacovina, Deora, Febbraio, Simantov, Silverstein, Hempstead, Mark & Hajjar, 2004; Dassah, Deora, He & Hajjar, 2009; Surette, Madureira, Phipps, Miller, Svenningsson & Waisman, 2011; Madureira, Surette, Phipps, Taboski, Miller & Waisman, 2011b; Pluskota, Soloviev, Bdeir, Cines & Plow, 2004).
3.2 Structural Determinants within Plasminogen that Mediate its Interaction with Cells
The recent X-ray crystal structures of full-length human Glu-plasminogen (Law, Caradoc-Davies, Cowieson, Horvath, Quek, Encarnacao, Steer, Cowan, Zhang, Lu, Pike, Smith, Coughlin & Whisstock, 2012; Xue, Bodin & Olsson, 2012) have provided new insights into the structural determinants in the plasminogen molecule that mediate its interaction with cells. The interactions of plasminogen with all cell types evaluated are blocked by lysine and lysine analogs, e.g. EACA, as well as peptides with carboxyl terminal lysines. This implies that the lysine binding sites within the disulfide-bonded kringle domains of plasminogen are required for the interaction with cells. Indeed, isolated kringle-containing plasminogen domains can specifically bind to cells with the following order of potency: The kringle 1–3 domain > the kringle 5-protease domain > kringle 2 (Miles et al., 1988a). Nonetheless, the crystal structures of plasminogen reveal that only the lysine binding site of kringle 1 is available in the closed form of Glu-plasminogen, suggesting that kringle 1 mediates the initial recruitment of plasminogen to the cell surface (Law, Caradoc-Davies, Cowieson, Horvath, Quek, Encarnacao, Steer, Cowan, Zhang, Lu, Pike, Smith, Coughlin & Whisstock, 2012).
3.3. Conformational Changes Induced in Plasminogen upon Binding to Cells and Their Relationship to Plasminogen Activation
As discussed in section 2.2.1, Glu-plasminogen exists in a closed tight conformation in the presence of chloride ion (Castellino & Ploplis, 2005). In the presence of lysine and lysine analogs, Glu-plasminogen adopts a more open relaxed conformation that is much more readily activated by plasminogen activators (Violand, Sodetz & Castellino, 1975). In addition, plasmin catalyzes hydrolysis of the N-terminal 77 amino acids of Glu-plasminogen, resulting in formation of a truncated form with an N-terminal lysine, Lys-plasminogen. Lys-plasminogen exists in an open conformation and is, consequently, more readily activated by plasminogen activators than Glu-plasminogen in the closed conformation (Wiman, 1973; Wiman & Wallen, 1973; Violand & Castellino, 1976).
When Glu-plasminogen binds to cells, its activation is markedly enhanced, compared with the reaction in solution due to a reduction in the Km (by 11 to 60-fold) for the plasminogen activation reaction in solution (Miles & Plow, 1985; Stricker, Wong, Tak Shiu, Reyes & Shuman, 1986; Hajjar, Harpel, Jaffe & Nachman, 1986; Loscalzo & Vaughan, 1987; Ellis, Behrendt & Dano, 1991; Gonias, Braud, Geary & VandenBerg, 1989; Duval-Jobe & Parmely, 1994; Félez, Miles, Fábregas, Jardi, Plow & Lijnen, 1996; Longstaff, Merton, Fabregas & Felez, 1999; Sinniger, Merton, Fabregas, Felez & Longstaff, 1999). This suggests that Glu-plasminogen on the cell surface adopts a conformation distinct from its conformation in solution. Direct evidence for such a conformational change was obtained recently using monoclonal anti-plasminogen antibodies that recognize receptor-induced binding sites (RIBS) in Glu-plasminogen upon its interaction with cells, but react poorly with soluble Glu-plasminogen (Han et al., 2011b; Felez et al., 2012; Jardi et al., 2012). Previously, it had generally been accepted that Glu-plasminogen adopts a Lys-plasminogen-like open conformation when bound to the cell surface, to account for enhancement of activation of cell-associated Glu-plasminogen (Dejouvencel et al., 2010). However, soluble Lys-plasminogen did not compete for the interaction of anti-plasminogen RIBS mAbs with surface-associated Glu-plasminogen, suggesting that the conformation induced when Glu-plasminogen binds to cells is distinct from the conformation of Lys-plasminogen (Han, Baik, Kim, Yang, Han, Gong, Jardi, Castellino, Felez, Parmer & Miles, 2011b).
The crystal structure of full-length Glu-plasminogen has revealed that the plasmin cleavage site in the N-terminus (that produces Lys-plasminogen) is buried in the closed conformation of Glu-plasminogen, suggesting that a conformational rearrangement precedes the production of Lys-plasminogen (Law, Caradoc-Davies, Cowieson, Horvath, Quek, Encarnacao, Steer, Cowan, Zhang, Lu, Pike, Smith, Coughlin & Whisstock, 2012) (Figure 3). Plasmin proteolysis of Glu-plasminogen to Lys-plasminogen is promoted when Glu-plasminogen is bound to the cell surface (Gong et al., 2001; Miles et al., 2003; Zhang, Gong, Grella, Castellino & Miles, 2003). Taken together, these studies suggest that the conformation adopted by Glu-plasminogen when bound to cells is one that exposes the cleavage site for plasmin-mediated removal of the N-terminal 77 amino acids, as a mechanism for promoting the conversion of Glu-plasminogen to Lys-plasminogen on the cell surface. Thus, the mechanism by which plasminogen binding to the cell surface promotes plasminogen activation is by accelerating cleavage of Glu-plasminogen to the more readily activated Lys-plasminogen form. Analysis of the crystal structure of plasminogen (Law, Caradoc-Davies, Cowieson, Horvath, Quek, Encarnacao, Steer, Cowan, Zhang, Lu, Pike, Smith, Coughlin & Whisstock, 2012) as well as earlier studies (Thorsen, 1992; Marshall et al., 1994; Hasumi et al., 2010; Gutierrez-Fernandez, Gingles, Bai, Castellino, Parmer & Miles, 2009) suggest that this conformational rearrangement may be due to an interaction of the lysine binding site within plasminogen kringle 5 with plasminogen receptors on the cell surface. Notably, the kringle 5 domain interacts with plasminogen receptors on the cell surface, as discussed in Section 3.2, above.
3.4 Molecular Identity of Plasminogen Receptors-Role of Carboxyl Terminal Lysines in Plasminogen Binding to Cells
A key concept regarding the mechanism by which interaction of plasminogen with the eukaryotic cell surface promotes plasminogen activation is that a subset of carboxypeptidase B (CpB)-sensitive plasminogen binding proteins is responsible for enhancing plasminogen activation. When cells are treated with CpB, the ability to stimulate plasminogen activation is lost (Felez et al., 1996). Since CpB removes C-terminal basic residues, these results imply that plasminogen binding proteins exposing C-terminal basic residues on cell surfaces are responsible for stimulation of plasminogen activation.
Several distinct plasminogen receptors have been identified over the past decades, consistent with the high number of receptors determined/cell and also consistent with the diversity of cell types that bind plasminogen (see Section 3.1). Until recently, known CpB-sensitive cellular plasminogen receptors could be divided into two classes: 1) proteins synthesized with C-terminal basic residues and having well established intracellular functions, including α-enolase (Miles et al., 1991; Redlitz et al., 1995), cytokeratin 8 (Hembrough, Vasudevan, Allietta, Glass & Gonias, 1995; Hembrough et al., 1996), S100A10 (in complex with annexin A2 within the annexin A2 heterotetramer) (Kassam et al., 1998; Choi et al., 2003; Madureira, Surette, Phipps, Taboski, Miller & Waisman, 2011b), TIP49a (Hawley et al., 2001) and histone H2B (Herren et al., 2006) and; 2) proteins requiring proteolytic processing in order to reveal a C-terminal basic residue (lysine), including actin (Dudani & Ganz, 1996; Miles et al., 2006). It was initially proposed that the annexin A2 monomer functioned directly as a plasminogen receptor after a proteolytic cleavage event to liberate a new C-terminal lysine (Hajjar et al., 1994). However, recent data suggest that the profibrinolytic role of annexin A2 is to transport and localize the plasminogen regulatory protein, S100A10, to the cell surface within the annexin A2 heterotetramer [reviewed in (Madureira, Surette, Phipps, Taboski, Miller & Waisman, 2011b; Madureira, O’Connell, Surette, Miller & Waisman, 2012)]. It should be noted that there is a CpB-insensitive component of plasminogen binding to eukaryotic cells, as exemplified by tissue factor (Fan et al., 1998) and non-protein gangliosides (Miles et al., 1989). However, this CpB-insensitive class of plasminogen receptors does not appreciably promote activation of cell-bound plasminogen (Felez, Miles, Fabregas, Jardi, Plow & Lijnen, 1996). Integrins, including αIIbβ3 (Miles et al., 1986; Gonzalez-Gronow et al., 1993), αMβ2 (Pluskota, Soloviev, Bdeir, Cines & Plow, 2004; Lishko et al., 2004) and α5β1 (Lishko, Novokhatny, Yakubenko, Skomorovska-Prokvolit & Ugarova, 2004), as well as amphoterin (Parkkinen et al., 1993) and GP330 (Kanalas & Makker, 1991; Kanalas, 1992) are plasminogen binding proteins not synthesized with C-terminal basic residues. Whether this group of proteins undergoes proteolysis to reveal C-terminal basic residues and/or are susceptible to CpB proteolysis has not been investigated.
4. Role of Plasminogen in Macrophage Recruitment in vivo
4.1 Overview of the Thioglycollate-Stimulated Leukocyte Migration Model
In the acute inflammatory response, macrophages/monocytes accumulate at sites of injury where they participate in the innate immune response as professional antigen presenting cells, in cytokine secretion and as direct effector cells (Gordon, 1999) and later, participate in wound healing (Leibovich & Ross, 1975). Inflammation of the peritoneum induced by sterile injection of thioglycollate is a widely-used model to study acute inflammation. This is a convenient model because inflammatory cells can be obtained readily by lavage in reasonable numbers, as compared to other tissue sites such as the synovium. Thioglycollate injection results in the sequential recruitment of granulocytes (within hours), monocytes (within 2 days) and lymphocytes (after 2–3 days). The basis for this differential rate of cellular recruitment is that granulocytes are recruited from blood, whereas recruitment of monocytes and lymphocytes involves their release from the bone marrow. Monocytes traverse the endothelial barrier and arrive in the peritoneum where they differentiate into macrophages. Therefore, the inflammatory response to thioglycollate is multi-faceted and closely orchestrated, requiring the ability to respond to cytokines, to synthesize cytokines, to migrate, to degrade extracellular matrices, to adhere to endothelium and roll, and, once attached to the endothelium, to detach and penetrate the basement membrane. The enhanced recruitment to the peritoneum is counterbalanced by apoptosis of peritoneal macrophages (Kuhn et al., 2001).
4.2. Requirement for Plasminogen in Macrophage Migration
Genetic deletion of plasminogen has a profound effect on the ability of monocytes/macrophages to migrate to the peritoneum in response to thioglycollate. Monocyte recruitment is maximal at 72 hours and is severely compromised in plasminogen (Plg)−/− mice (decreased by 56% compared to the level in Plg+/+ mice) (Ploplis, French, Carmeliet, Collen & Plow, 1998; Gong et al., 2008). Furthermore, in the absence of plasminogen, monocyte recruitment reaches a plateau 24 hr after injection of thioglycollate, whereas monocyte recruitment continues to increase in Plg+/+ mice. It is noteworthy that monocyte recruitment achieves an intermediate level in heterozygous Plg+/− mice (decreased by 43% compared to the level in Plg+/+ mice), suggesting that even partial lack of plasminogen function can affect monocyte recruitment. In these experiments, important controls established that the decreased recruitment of monocytes to the peritoneum in Plg−/− mice is not due to a decreased source of monocytes in the circulation (Ploplis, French, Carmeliet, Collen & Plow, 1998).
Plasminogen deficiency also affects lymphocyte recruitment in the thioglycollate model (decreased by 38%in Plg−/− mice vs Plg+/+ mice) (Ploplis, French, Carmeliet, Collen & Plow, 1998). In contrast, there is no effect of plasminogen deficiency on neutrophil recruitment in response to thioglycollate (Ploplis, French, Carmeliet, Collen & Plow, 1998).
4.3 Requirement for Plasmin Formation in Macrophage Recruitment
Several lines of evidence suggest that formation of the active enzyme, plasmin, is required for plasminogen-dependent cell recruitment. First, plasmin activity is increased in the peritoneal lavage fluid following thioglycollate administration (Gong, Hart, Shchurin & Hoover-Plow, 2008). Second, aprotinin (a potent inhibitor of plasmin activity) suppresses the level of macrophage recruitment in wild-type mice to a level, similar to that found in Plg−/− mice (Gong, Hart, Shchurin & Hoover-Plow, 2008). Recently, mice expressing a plasmin-inactivating active site mutation have been generated (Iwaki et al., 2010). When these mice are tested in the thioglycollate recruitment model, the question of the requirement for active plasmin will be definitively resolved. In further support of a role for plasmin, tPA deficient mice exhibit impaired monocyte recruitment to the peritoneum (Ploplis & Castellino, 2002; Cook et al., 2006; Cao et al., 2006) and, conversely, macrophage recruitment is significantly increased in PAI-1−/− mice (Gong, Hart, Shchurin & Hoover-Plow, 2008). uPA−/− macrophages have been reported to both invade normally into the peritoneum in response to thioglycollate (Carmeliet, Schoonjans, Kieckens, Ream, Degen, Bronson, De Vos, Van den Oord, Collen & Mulligan, 1994; Cook, Vlahos, Massa, Braine, Lenzo, Turner, Way & Hamilton, 2006; Connolly et al., 2010) as well as to exhibit reduced migration (Ploplis & Castellino, 2002; Cao, Lawrence, Li, Von Arnim, Herz, Su, Makarova, Hyman, Strickland & Zhang, 2006; Han et al., 2011a). Nonetheless, the consensus of several studies is that recruitment of macrophages and neutrophils is not diminished in uPAR-deficient mice (Carmeliet, Schoonjans, Kieckens, Ream, Degen, Bronson, De Vos, Van den Oord, Collen & Mulligan, 1994; Ploplis & Castellino, 2002; Cook, Vlahos, Massa, Braine, Lenzo, Turner, Way & Hamilton, 2006; Connolly, Choi, Gardsvoll, Bey, Currie, Chavakis, Liu, Molinolo, Ploug, Leppla & Bugge, 2010). Taken together, the uPA-uPAR axis does not appear to play a detectable role in thioglycollate-dependent macrophage migration (Connolly, Choi, Gardsvoll, Bey, Currie, Chavakis, Liu, Molinolo, Ploug, Leppla & Bugge, 2010).
4.3.1 The Role of Plasmin-Dependent-Matrix Degradation in Macrophage Recruitment
A key step in plasmingen-dependent macrophage recruitment is transmigration of macrophages across the peritoneal membrane into the peritoneal cavity. Macrophage accumulation in peritoneal tissue of Plg−/− mice is 4-fold-higher than wild-type mice (Gong, Hart, Shchurin & Hoover-Plow, 2008). The accumulated macrophages are clearly localized on the submesothelial layer of the peritoneal tissue and this is accompanied by enhanced collagen deposition in the peritoneal tissue (Gong, Hart, Shchurin & Hoover-Plow, 2008), consistent with a defect the ability of macrophages to transmigrate through the extracellular matrix (ECM), due to decreased ability to degrade collagen within the matrix.
Activation of MMP-9 is also required for plasminogen-dependent transmigration of macrophages. First, the lack of ability to transmigrate is accompanied by decreased activation of MMP-9 in the peritoneal fluid of Plg−/− mice. Second, injection with an MMP-9 neutralizing antibody inhibits macrophage recruitment in Plg+/+, but not Plg-/ mice. Third, the number of recruited macrophages was restored to a significant extent when Plg−/− mice are treated with activated MMP-9, but there was no enhancement of recruitment in Plg+/+ mice. Fourth, MMP-9−/− mice exhibit a significant reduction in macrophage recruitment in response to thioglycollate (Gong, Hart, Shchurin & Hoover-Plow, 2008). Thus, these authors concluded that activation of MMP-9 is necessary for plasminogen-regulated macrophage migration (Gong, Hart, Shchurin & Hoover-Plow, 2008). They did raise the issue that MMP-9 is not an interstitial collagenase, and so may participate in the degradation of interstitial collagen indirectly (Gong, Hart, Shchurin & Hoover-Plow, 2008). Although, plasmin production clearly appears to lead to MMP-9 activation, plasmin proteolytically cleaves MMP-9, but does not produce an enzymatically active MMP-9 (Ramos-DeSimone et al., 1999). Thus, an intermediate enzyme that is activated by plasmin is likely to link plasmin production and MMP-9 activation. Notably, MMP-3 (stromelysin-1) is very efficiently activated by plasmin and activated MMP-3 is a potent activator of MMP-9 (Ramos-DeSimone, Hahn-Dantona, Sipley, Nagase, French & Quigley, 1999). These in vivo studies are buttressed by in vitro data indicating a requirement for plasmin and MMP-9 in macrophage migration across the representative ECM, Matrigel, and collagen IV (Gong, Hart, Shchurin & Hoover-Plow, 2008).
Other ECM components that are subject to plasmin proteolysis include laminin and fibronectin (Liotta et al., 1981b; Liotta et al., 1981a). Although laminin is a major component of the basement membrane underlying mesothelial cells within the peritoneal tissue (Nagy, 1996), there is no difference in the laminin content of peritoneal tissue of Plg++ and Plg−/− mice treated with thioglycollate (Gong, Hart, Shchurin & Hoover-Plow, 2008), suggesting that laminin degradation by plasmin may not be required for macrophage transmigration across the peritoneal membrane. Fibronectin degradation has not been examined in this model.
Extravascular fibrin functions as a provisional extracellular matrix at sites of inflammation (Szaba & Smiley, 2002). In response to an inflammatory stimulus, both resident and newly recruited macrophages participate in a phenomenon known as “the macrophage disappearance reaction” in which there is a large decrease in macrophages that are recovered from peritoneal exudates due to a concomitant increase in macrophages adhering to the peritoneal lining (Barth et al., 1995). This response is inhibited by heparin and warfarin and, therefore, appears to be dependent on the coagulation system (Nelson, 1965). Furthermore, cellular aggregates on the peritoneal wall are surrounded by fibrin filaments (Leak, 1983). Although macrophage recruitment to the peritoneum is not suppressed in fibrinogen −/− mice (Szaba & Smiley, 2002), macrophage adhesion to the peritoneal wall is suppressed in these mice (Szaba & Smiley, 2002). Interestingly, in tPA−/− mice increased numbers of macrophages expressing high levels of the integrin, Mac-I, are present on/in the lining of the peritoneal cavity and associated with areas of increased fibrin(ogen) staining, suggesting that the cells are adhering to fibrin (Cook, Vlahos, Massa, Braine, Lenzo, Turner, Way & Hamilton, 2006). Consistent with this interpretation, administration of plasmin significantly increased the numbers of macrophages present in the peritoneal cavity to the level of that in tPA+/+ mice, suggesting that plasmin had lysed the fibrin and caused release of the macrophages from the fibrin scaffold (Cook, Vlahos, Massa, Braine, Lenzo, Turner, Way & Hamilton, 2006). Adhesion of macrophages to fibrin on the peritoneal wall of Plg−/− mice could be an additional contributor to the low level of macrophages recovered in the peritoneal fluid in plasminogen−/− mice challenged with thioglycollate. This has not been addressed in the literature.
In a related mechanism, macrophage egress from the peritoneum to the lymph nodes in response to LPS is diminished in both tPA−/− and PAI-1−/− mice (Cao, Lawrence, Li, Von Arnim, Herz, Su, Makarova, Hyman, Strickland & Zhang, 2006). In combination with additional data demonstrating a requirement for Mac-1-dependent adhesion to fibrin and LDL Receptor Related Protein (LRP) in egress from the peritoneum, these results have been interpreted as indicating the requirement for initial complex formation of Mac-1, fibrin and tPA, that forms an adhesive complex on the wall of the peritoneum, with subsequent neutralization of tPA by PAI-1, leading to Mac-1 internalization by LRP and cell detachment to allow egress from the peritoneum and migration to the lymphatics. In this study, quantification of macrophages on the peritoneal wall was not performed and, indeed, the initial recruitment of tPA−/− macrophages to the peritoneum in response to thioglycollate was compromised (Cao, Lawrence, Li, Von Arnim, Herz, Su, Makarova, Hyman, Strickland & Zhang, 2006). Thus, it remains possible that detachment of macrophages from fibrin lining the peritoneal cavity may be the rate limiting step in macrophage egress from the peritoneum. As discussed in Section 2.1.4, spontaneous phenotypes of plasminogen deficient mice are rescued by concomitant knockout of fibrinogen (Bugge, Kombrinck, Flick, Daugherty, Danton & Degen, 1996). Challenge of plasminogen−/−/fibrinogen−/− double knockout mice could resolve whether the major role of tPA in inflammatory recruitment in response to thioglycollate is due to fibrinolysis and/or fibrin-dependent adhesion. As fibrin fragments are also chemotactic for macrophages, the plasminogen/fibrinogen double knockouts could also be studied to address the role of fibrin degradation products in thioglycollate-induced peritonitis.
4.3.2 The Role of Plasmin-Dependent Signal Transduction in Macrophage Recruitment
Plasmin exhibits potent and direct effects on monocytes and macrophages (Syrovets, Lunov & Simmet, 2012) that have the potential to regulate many steps in the proinflammatory responses of these cells. Plasmin induces chemotaxis and actin polymerization in monocytes that is dependent on PKC and cGMP (Syrovets et al., 1997). In addition, plasmin stimulates expression of pro-inflammatory cytokines TNF-α, IL-1, MCP-1 and tissue factor, that is dependent upon NF-κB, AP-1 and STAT (Syrovets et al., 2001; Burysek et al., 2002). Plasmin stimulates the 5-lipoxygenase pathway to trigger synthesis of cysteinyl-leukotrienes and leukotriene B4 (Weide et al., 1994; Weide et al., 1996). In addition, plasmin induces COX-2, PGE2 and MMP-1 production by monocytes (Zhang et al., 2007). The contribution of each of these pathways to plasminogen-dependent thioglycollate-induced macrophage recruitment to the peritoneum has not yet been tested.
It is also noteworthy that expression of the plasminogen gene is up-regulated in response to inflammatory cytokines (Jenkins et al., 1997; Bannach et al., 2002; Gutierrez-Fernandez et al., 2003; Bannach et al., 2004). This up-regulation could increase local levels of active plasmin [including macrophage-derived plasmin (Zhang, Gong, Grella, Castellino & Miles, 2003)] and provide a mechanism for further amplifying the pathway in which plasmin induces cell signaling.
Elimination of activated macrophages by apoptosis, serves as a mechanism for down-regulating the inflammatory response (Kuhn, Godshall, Scott, Franklin, Rowe, Peyton & Cheadle, 2001). Plasmin activity on the monocyte surface is cytoprotective in apoptosis induced by proinflammatory agents (Mitchell et al., 2006). Thus, the presence of plasminogen may regulate the number of peritoneal macrophages by this mechanism, also. The extent of apoptosis in Plg−/− macrophages that successfully migrate to the peritoneum has not been evaluated to our knowledge.
4.3.3 The Role of Plasmin(ogen) Receptors in Macrophage Recruitment
As discussed above, plasminogen binding to cellular receptors is blocked by lysine and lysine analogs (Miles et al., 2005). Notably, macrophage recruitment to the peritoneum is suppressed by the lysine analog, tranexamic acid (Gong, Hart, Shchurin & Hoover-Plow, 2008), suggesting that plasminogen binding to cellular receptors plays a role in macrophage recruitment. Most tellingly, plasminogen-dependent macrophage recruitment in vivo is mediated by CpB-sensitive plasminogen receptors (Swaisgood et al., 2002). The roles of specific plasminogen receptors in macrophage recruitment are discussed below in Section 9.1.
5. Proteomics-Based Discovery of Plg-RKT
5.1 Need for a Proteomics Approach
As discussed in Section 3.4, until recently, plasminogen binding proteins exposing C-terminal basic residues were comprised of two classes: 1) proteins with intracellular functions that are synthesized with C-terminal lysines and are also detected on cell surfaces and 2) proteins requiring processing to expose a C-terminal lysine. No integral membrane proteins synthesized with and exposing a C-terminal basic residue on the cell surface had been identified. The existence of a plasminogen receptor with such a structure would constitute a novel mechanism for stimulating plasminogen activation because its induction would endow cells with the ability to bind plasminogen and promote plasminogen activation, without requiring release and rebinding of intracellular proteins or proteolytic cleavage of a membrane protein to reveal a C-terminal basic residue. Previous methodologies and characteristics of plasminogen binding proteins may have precluded identification of an integral membrane plasminogen binding protein with a C-terminal basic residue. The identification of plasminogen receptors has relied previously on cell surface labeling followed by affinity chromatography on plasminogen-Sepharose columns and N-terminal sequencing of fractions eluted from SDS gels. Thus, many intracellular proteins that are also present on the cell surface were readily identified because protein fractions that bound to plasminogen-Sepharose included the labeled, surface-associated protein, as well as nonlabeled intracellular protein. Using these methods, a lower abundance integral membrane plasminogen binding protein might not have been detectable. In addition, many membrane proteins are not well resolved on SDS polyacrylamide gels. Therefore, we used an isolation method that used column chromatography instead of SDS polyacrylamide gel analysis: We took advantage of the exquisite sensitivity of multidimensional protein identification technology (MudPIT) to search for integral membrane plasminogen receptor(s) exposing a C-terminal basic residue on the cell surface and present on viable cells.
5.2 Discovery of Plg-RKT
We used a proteomics approach involving MudPIT [reviewed in (Eng et al., 1994)] to probe the membrane proteome of differentiated, macrophage colony stimulating factor (M-CSF)-treated murine monocyte progenitor cells [Hoxa9-ER4 (Wang et al., 2006)] for the presence of integral membrane plasminogen receptor(s) exposing a C-terminal basic residue on the cell surface (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010) (Figure 5). Intact cells were biotinylated using a biotinylation reagent that reacts with carboxyl groups, rather than basic groups (thus, avoiding potential interference with the plasminogen-binding function of C-terminal basic residues). Because early apoptotic and non-viable/necrotic cells exhibit markedly enhanced plasminogen binding ability (O’Mullane MJ & Baker MS, 1998; O’Mullane MJ & Baker MS, 1999; Mitchell, Baik, Castellino & Miles, 2006) we focussed on plasminogen receptors on viable cells and, therefore, passed the biotinylated cells over a dead cell removal column to enrich for live cells. The cells were then lysed and membrane fractions prepared and passed over a plasminogen-Sepharose affinity column and specifically eluted with EACA. Biotinylated cell surface proteins bound to the avidin column and were digested with trypsin while still on the column. The peptide digest was then subjected to MudPI. In MudPIT, the peptide mixtures were first resolved by strong cation exchange liquid chromatography prior to reversed phase liquid chromatography. The eluting peptides were electrosprayed onto an LTQ ion trap mass spectrometer and full MS spectra were recorded over a 400–1600 m/z range, followed by three tandem mass events. The resulting spectra were searched against a mouse protein database. Only one protein with a predicted transmembrane sequence and a C-terminal basic residue was identified: the hypothetical protein, C9orf46 homolog (IPI00136293), homologous to the protein predicted to be encoded by human chromosome 9, open reading frame 46. We have designated the protein, Plg-RKT, to indicate a plasminogen receptor with a C-terminal lysine and having a transmembrane domain (see Section 6.2 below).
Figure 5. Isolation of Plasminogen Receptors.

Monocyte (Hoxa9-ER4) progenitor cells were differentiated with macrophage colony stimulating factor (M-CSF), which induces plasminogen receptors (
 ) on these cells. Then intact cells were biotinylated (●) and passed over a dead cell removal column. Live cells were then lysed and membrane fractions prepared and passed over a plasminogen-Sepharose affinity column and specifically eluted. Biotinylated plasminogen receptors (
●) were then bound to an avidin column and digested with trypsin. This figure was originally published in Proteomics/Book 1: Human Diseases and Protein Functions, Miles, L.A., Andronicos, N.M., Chen, E.I., Baik, N., Bai, H., Parmer, C.M., Lighvani, S., Nangia, S., Kiosses, W.B., Kamps, M.P., Yates III, J.R. and Parmer, R.J. Identification of the novel plasminogen receptor, Plg-RKT. In:
Proteomics/Book 1: Human Diseases and Protein Functions, ISBN 978-953-307-832-8, Man, T.K. and Flores, R.J.(Ed.), Intech, Chapter 10: 219–238, 2012.
) on these cells. Then intact cells were biotinylated (●) and passed over a dead cell removal column. Live cells were then lysed and membrane fractions prepared and passed over a plasminogen-Sepharose affinity column and specifically eluted. Biotinylated plasminogen receptors (
●) were then bound to an avidin column and digested with trypsin. This figure was originally published in Proteomics/Book 1: Human Diseases and Protein Functions, Miles, L.A., Andronicos, N.M., Chen, E.I., Baik, N., Bai, H., Parmer, C.M., Lighvani, S., Nangia, S., Kiosses, W.B., Kamps, M.P., Yates III, J.R. and Parmer, R.J. Identification of the novel plasminogen receptor, Plg-RKT. In:
Proteomics/Book 1: Human Diseases and Protein Functions, ISBN 978-953-307-832-8, Man, T.K. and Flores, R.J.(Ed.), Intech, Chapter 10: 219–238, 2012.
6. Structure of Plg-RKT
The C9orf46 homolog/Plg-RKT murine DNA sequence encodes a protein of 147 amino acids with a molecular mass of 17,261 Da and a C-terminal lysine (Figure 6, Panel a). We blasted the C9orf46 homolog/Plg-RKT sequence against all species using NCBI Blast and obtained unique human, rat, dog, cow, dog, giant panda, gibbon, horse, pig, rabbit, and rhesus monkey predicted orthologs, which exhibited high identity (e.g. human vs. chimpanzee = 99% identity) and no gaps in the sequence. Of key importance, a C-terminal lysine was predicted for all of the mammalian orthologs obtained in the blast search. In a query of the Ensembl Gene Report, DNA sequences of all 10 other sequenced mammalian orthologs encoded a C-terminal lysine (Miles et al., 2012b).
Figure 6. High Interspecies Homology of Plg-RKT.

Alignment of predicted amino acid sequences of mouse, human, rat, dog and cow orthologs of Plg-RKT (a) and the structural model of Plg-RKT (b). Green indicates amino acids within the predicted primary transmembrane helix. Orange indicates amino acids within the predicted secondary transmembrane helix. Red indicates basic amino acids. This research was originally published in Blood, Andronicos, N.M., Chen, E.I., Baik, N., Bai, H., Parmer, C.M., Kiosses, W.B., Kamps, M.P., Yates, J.R., III, Parmer, R.J., Miles, L.A., Proteomics-based discovery of a novel, structurally unique, and developmentally regulated plasminogen receptor, Plg-RKT, a major regulator of cell surface plasminogen activation, Blood. 2010, 115: 1319-30.
It is noteworthy that the primary sequence of C9orf46/Plg-RKT is apparently tightly conserved in humans, with no validated coding polymorphisms (cSNPs) thus far identified within the 6 exons encoded by the gene (on chromosome 9p24.1) in the NCBI human genome sequence variation database (dbSNP, http://www.ncbi.nlm.nih.gov/SNP).
6.1 Conservation of the Plg-RKT Sequence Among Species
In addition to mammals, the DNA sequences of xenopus, the green lizard and zebrafish also encode a C-terminal lysine. The Plg-RKT sequence also encodes a putative conserved DUF2368 domain (encompassing amino acids 1–135), an uncharacterized protein with unknown function conserved from nematodes to humans. Notably, the DNA sequences of Plg-RKT orthologs of lower organisms (e.g. the sea urchin, Strongylocentrotus purpuratus, Drosophila and Paramecium) predicted proteins of different lengths and did not consistently predict C-terminal lysines. It is interesting to note that the evolutionary origin of plasminogen is currently believed to originate with protochordates (Liu & Zhang, 2009), so that lower organisms without plasminogen would not need the C-terminal lysine of Plg-RKT to bind plasminogen.
6.2 Topology of Plg-RKT
We analyzed the C9orf46 homolog/Plg-RKT sequence in the TMpred site (www.ch.embnet.org/cgi-bin/TMPRED). The strongly preferred model included two transmembrane helices extending from F53-L73 (secondary helix, oriented from outside the cell to inside the cell) and P78-Y99 (primary helix, oriented from inside the cell to outside the cell) (Figure 6, Panel b). Thus, a 52 amino acid N-terminal region and a 48 amino acid C-terminal tail with a C-terminal lysine were predicted to be exposed on the cell surface.
6.2.1 Experimental Evidence that Plg-RKT is a Transmembrane Protein Exposing a C-terminal Lysine on the Cell Surface
The predictions of the topology model were supported using several experimental approaches. 1) In Triton X-114 phase separation experiments Plg-RKT partitioned to the detergent phase, thus behaving as an integral membrane protein (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010; Bai et al., 2011). 2) When we treated intact cells with CpB, prior to performing our proteomic analysis, Plg-RKT was not recovered, consistent with cell surface exposure of the C-terminus of Plg-RKT (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010). 3) A mAb raised against the C-terminal peptide of Plg-RKT reacted with the cell surface (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010; Bai, Baik, Kiosses, Krajewski, Miles & Parmer, 2011). 4) Extracellular exposure of both N-and C-termini of Plg-RKT was supported by protease accessibility experiments (Miles, Lighvani, Baik, Andronicos, Chen, Parmer, Khaldoyanidi, Diggs, Kiosses, Kamps, Yates, III & Parmer, 2012b; Miles et al., 2012a).
7. Tissue and Cellular Distribution of Plg-RKT
Because the murine genome has been sequenced we searched for C9orf46 homolog/Plg-RKT mRNA microarray expression data at http://www.ebi.ac.uk/microarray-as/aew/. Plg-RKT mRNA is present in monocytes, leukocytes, NK cells, T cells, myeloid, dendritic, and plasmacytoid cells, breast cancer, acute lymphoblastic leukemia and Molt-4 acute lymphoblastic leukemia cells. These data are consistent with previous reports documenting expression of plasminogen binding sites on peripheral blood leukocytes (Miles & Plow, 1987), breast cancer cells (Correc et al., 1990; Ranson, Andronicos, O’Mullane & Baker, 1998) and other tissues [reviewed in (Miles, Hawley, Baik, Andronicos, Castellino & Parmer, 2005)]. The broad distribution in tissues that express plasminogen binding sites, suggest that Plg-RKT provides plasminogen receptor function that may serve to modulate plasmin proteolytic functions in these tissues, as well.
8. Plg-RKT and Plasminogen Receptor Functions
8.1 Role of the C-terminal Lysine of Plg-RKT in Plasminogen Binding to Cells
In order to study the function of the C-terminus of Plg-RKT we raised an anti-Plg-RKT mAb against the Plg-RKT C-terminal peptide (CEQSKLFSDK, with the C added for coupling). The mAb specifically blocks Plg-RKT binding to the immobilized peptide (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010; Lighvani et al., 2011). Confocal microscopy studies with anti-Plg-RKT mAb demonstrated that the C-terminus of Plg-RKT is exposed on the cell surface and that plasminogen binds to the C-terminal domain of Plg-RKT on the cell surface (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010) (Figure 7).
Figure 7. Plg-RKT binds plasminogen on the cell surface.

A. M-CSF-differentiated (Hoxa9-ER4) cells were grown on coverslips and preincubated with either phosphate buffered saline (− plasminogen) or 2 μM plasminogen (+ plasminogen), then fixed in 1% formaldehyde, washed and stained with polyclonal anti-plasminogen IgG or anti-Plg-RKT mAb and stained with a combination of Alexa 488- F(ab′)2 of goat anti-rabbit IgG and Alexa 568- F(ab′)2 fragment of goat anti-mouse IgG.
B. The number and size of each labeled aggregate was determined. The results reflect counts from over 40 cells in 2 independent experiments. Data represent mean ± SEM. *p < 0.001. This research was originally published in Blood, Andronicos, N.M., Chen, E.I., Baik, N., Bai, H., Parmer, C.M., Kiosses, W.B., Kamps, M.P., Yates, J.R., III, Parmer, R.J., Miles, L.A., Proteomics-based discovery of a novel, structurally unique, and developmentally regulated plasminogen receptor, Plg-RKT, a major regulator of cell surface plasminogen activation, Blood. 2010, 115: 1319-30.
In addition, the synthetic peptide, corresponding to the C-terminus of Plg-RKT interacts specifically with plasminogen. Glu-plasminogen binds to the immobilized peptide in a concentration dependent manner, and the soluble peptide competes for Glu-plasminogen binding to the immobilized peptide with an IC50 of 2 μM, similar to the Kd determined for the interaction of Glu-plasminogen with M-CSF treated monocyte progenitor (Hoxa9-ER4) cells (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010). Furthermore, a mutated peptide with the C-terminal lysine substituted with alanine does not compete for Glu-plasminogen binding, further supporting the role of the C-terminal lysine in the interaction of Plg-RKT with Glu-plasminogen. Lys-plasminogen also binds to the peptide with an affinity ~3-fold greater than that of Glu-plasminogen, consistent with the higher affinity of Lys-plasminogen compared to Glu-plasminogen for the cell surface (Hajjar et al., 1987; Miles, Dahlberg & Plow, 1988a). tPA also binds to the peptide in a concentration-manner (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010). This is consistent with earlier studies demonstrating that tPA and plasminogen share binding sites on monocytoid cells and tPA binding to monocytoid cells is sensitive to CpB (Felez et al., 1993).
8.2 Plg-RKT Regulates Cell Surface Plasminogen Activation
Plasminogen activation by tPA is stimulated 12.7-fold in the presence of M-CSF treated monocyte progenitor (Hoxa9-ER4) cells, compared to the reaction in the absence of cells (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010). Anti-Plg-RKT mAb substantially suppresses this activation, by 58%, demonstrating a key role of Plg-RKT in regulating cell surface plasminogen activation.
Plg-RKT is markedly colocalized with uPAR on the surfaces of M-CSF-differentiated monocyte progenitor Hoxa9-ER4 cells, as revealed by merged confocal images. The extent of colocalization of Plg-RKT with uPAR is 73 ± 3% (Figure 8) (Andronicos, Chen, Baik, Bai, Parmer, Kiosses, Kamps, Yates, III, Parmer & Miles, 2010), suggesting that Plg-RKT and uPAR are present in very close proximity on the cell surface in an orientation to promote plasminogen activation.
Figure 8. Plg-RKT colocalizes with uPAR.
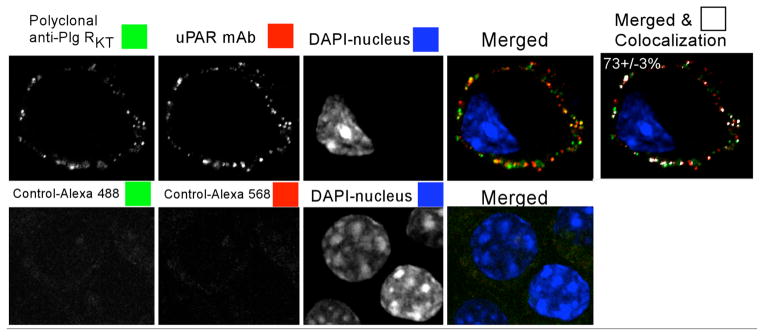
M-CSF-differentiated Hoxa9-ER4 cells were grown on coverslips and incubated with a combination of polyclonal rabbit anti-Plg-RKT IgG and mouse monoclonal anti-uPAR. Cells were washed, fixed in 1% formaldehyde and then stained with a combination of Alexa 488- F(ab′)2 of goat anti-rabbit IgG and Alexa 568-F(ab′)2 fragment of goat anti-mouse IgG. Controls (lower panel) are samples incubated without first antibody. The results reflect colocalization correlation coefficients (M1) values (last column) from over 40 cells in 2 independent experiments. *p < 0.001. This research was originally published in Blood, Andronicos, N.M., Chen, E.I., Baik, N., Bai, H., Parmer, C.M., Kiosses, W.B., Kamps, M.P., Yates, J.R., III, Parmer, R.J., Miles, L.A., Proteomics-based discovery of a novel, structurally unique, and developmentally regulated plasminogen receptor, Plg-RKT, a major regulator of cell surface plasminogen activation, Blood. 2010, 115: 1319-30.
The kinetically favored substrate for uPAR-bound uPA is cell-associated, rather than solution phase plasminogen (Ellis, Behrendt & Dano, 1991). uPA-dependent cell surface plasminogen activation was substantially inhibited anti-Plg-RKT mAb (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011). Therefore, Plg-RKT plays a major role in uPA-dependent plasminogen activation on monocytoid cells.
8.3 Regulation of Cell Migration by Plg-RKT
Invasion of the representative ECM, Matrigel, by monocytoid cells in response to the chemotactic stimulus, MCP-1, is enhanced in the presence of plasminogen and also requires active plasmin (Das et al., 2007; O’Connell et al., 2010a) and uPA (Das, Burke & Plow, 2007) and is markedly suppressed in the presence of EACA, suggesting a key role of plasminogen receptors in this function(Das, Burke & Plow, 2007; Wygrecka et al., 2009; O’Connell, Surette, Liwski, Svenningsson & Waisman, 2010a). Treatment of U937 cells or human peripheral blood monocytes with anti-Plg-RKT mAb markedly decreases migration of the cells through Matrigel (by 54% and 48%, respectively) (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011) demonstrating a major role for Plg-RKT in Matrigel invasion, consistent with the role of Plg-RKT in regulation of plasminogen activation.
Plasmin also promotes chemotactic cell migration across polycarbonate membranes in the absence of ECM (Syrovets, Tippler, Rieks & Simmet, 1997). Chemotactic migration of both U937 monocytoid cells and human peripheral blood monocytes was maximally reduced by 64%, and 39%, respectively, by treatment with anti-Plg-RKT mAb (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011). Regulation of chemotactic migration in the absence of ECM appears to be a unique property of Plg-RKT because other plasminogen receptors do not regulate chemotactic migration (Das, Burke & Plow, 2007). In addition to chemotaxis, Plg-RKT is involved in chemokinesis, with chemokinesis being suppressed almost to a background level in the presence of anti-Plg-RKT mAb (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011).
8.4 Regulation of the Inflammatory Response by Plg-RKT
Plg-RKT also plays a key role in monocyte migration in vivo. In the thioglycollate model or sterile peritonitis, macrophage recruitment is significantly (49%) impaired in mice injected with anti-Plg-RKT mAb compared with mice injected with the isotype control (2.46 X 105 ± 0.28 X 104 for mice injected with anti-Plg-RKT mAb vs 4.82 X 105 ± 0.33 X 104 for mice injected with isotype control, n=5, p=.00048) (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011). The decreased response in macrophage recruitment to the peritoneum was not due to a decreased level of monocytes in the circulation. In addition, similar to the results with plasminogen deficient mice, treatment with anti-Plg-RKT mAb also reduced lymphocyte recruitment to the peritoneum, but did not affect neutrophil and eosinophil recruitment (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011). Furthermore, treatment with anti-Plg-RKT mAb substantially reduce activation of pro-MMP-9 in the peritoneal fluid, similar to results with plasminogen deficient mice and consistent for the requirement for activation of pro-MMP-9 in plasminogen-dependent monocyte recruitment (Gong, Hart, Shchurin & Hoover-Plow, 2008).
The effect of anti-Plg-RKT mAb in the peritonitis model is consistent with the plasminogen binding function of Plg-RKT. Thioglycollate stimulated recruitment of macrophages in Plg−/− mice injected with isotype control is significantly decreased (by 73 %) in Plg−/− compared to Plg+/+ littermates, as reported [56% (Ploplis, French, Carmeliet, Collen & Plow, 1998) – 65% (Gong, Hart, Shchurin & Hoover-Plow, 2008)]. When Plg−/− mice are treated with anti-Plg-RKT mAb, there is no effect on the residual low level of macrophage recruitment in Plg−/− mice. Thus, the effect of the anti- Plg-RKT mAb is entirely dependent on plasminogen, consistent with Plg-RKT exhibiting plasminogen receptor function in vivo.
9. Interplay among Plasminogen Receptors in the Macrophage Migration in Experimentally Induced Peritonitis
The thioglycollate-induced model of macrophage recruitment originally demonstrated the key role of plasminogen receptors with C-terminal lysines in cell migration in the inflammatory response in vivo (Swaisgood, Schmitt, Eaton & Plow, 2002). The high number of plasminogen binding sites/cell taken together with the diversity of cell types that bind plasminogen (Miles, Levin, Plescia, Collen & Plow, 1988b) suggests that the plasminogen binding capacity of a given cell may be composed of contributions from a set of distinct cell surface proteins and that different cell types may utilize a different panel of plasminogen receptors [recently reviewed by Plow and colleagues in (Plow et al., 2012)]. The results of our proteomic analysis of differentiated monocyte progenitor cells were consistent with this concept: In addition to peptides corresponding to Plg-RKT, peptides corresponding to other proteins previously identified as plasminogen binding proteins on monocytes were also detected in the membrane preparations: a-enolase, gamma actin, S100A10, annexin 2 (that most likely bound to the plasminogen-Sepharose column via S100A10 in the annexin 2 heterotetramer), histone H2B, and β2 integrin.
Recent studies utilizing the thioglycollate-induced peritonitis model illustrate the interplay among distinct plasminogen receptors in vivo because the sum of the effects of functional blockade of specific plasminogen receptors, is greater than a 100% reduction in plasminogen-dependent macrophage recruitment. Intravenous injection of specific antibodies to histone H2B results in 48% less macrophage recruitment (Das, Burke & Plow, 2007), injection of specific antibodies to a-enolase results in 24% less recruitment (Das, Burke & Plow, 2007) and injection of mice with anti- Plg-RKT mAb results in 49% less macrophage recruitment (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011) (compared to injection of nonimmune control). In S100A10−/− mice, macrophage recruitment in response to thioglycollate is 53% less in S100A10 −/− mice, compared to wild type mice (O’Connell et al., 2010b). Thus, it is likely that each specific plasminogen receptor may be required at different steps in the inflammatory response, for example chemotactic migration to the peritoneum, or, perhaps, crossing different layers of peritoneal tissue at which different contributions of direct plasmic cleavage of the extracellular matrix or activation of MMP-9 for collagen degradation (Gong, Hart, Shchurin & Hoover-Plow, 2008). It is noteworthy that a reduction in pro-MMP-9 activation has been demonstrated in S100A10−/− peritoneal macrophages in culture (O’Connell, Surette, Liwski, Svenningsson & Waisman, 2010b) and by treatment of mice with anti-Plg-RKT mAb in vivo and there may be overlap in this function, as well. Although each of these receptors regulates Matrigel invasion in vitro (Das, Burke & Plow, 2007; Wygrecka, Marsh, Morty, Henneke, Guenther, Lohmeyer, Markart & Preissner, 2009; O’Connell, Surette, Liwski, Svenningsson & Waisman, 2010b; Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011), only Plg-RKT has been demonstrated to contribute to directed chemotaxis/chemokinesis (Lighvani, Baik, Diggs, Khaldoyanidi, Parmer & Miles, 2011). Thus, Plg-RKT may be the predominant modulator of this component of macrophage recruitment.
It should also be recognized that the annexin A2/S100A10 complex binds to anionic phospholipids in a Ca+2-dependent manner (Madureira, Surette, Phipps, Taboski, Miller & Waisman, 2011b; Madureira, O’Connell, Surette, Miller & Waisman, 2012; Rattner et al., 1991; Ross et al., 2003) and it has recently been shown that histone H2B is tethered to the cell surface via an electrostatic interaction with phosphatidyl serine (Das & Plow, 2011). Phosphatidyl serine exposure is a well established marker of apoptosis. When apoptosis is induced, cell surface expression of both histone H2B and S100A10 is increased (Plow, Doeuvre & Das, 2012; Das & Plow, 2011). In addition, differentiation of monocytes to macrophages (Marguet et al., 1999; Callahan et al., 2003) and cellular activation (Zwaal et al., 2005) are associated with phosphatidyl serine exposure and when cells are induced to differentiate, phosphatidyl serine exposure is increased on non-apoptotic cells (Das & Plow, 2011). Thus, differential exposure of phosphatidyl serine may play a role in determining which plasminogen receptors are utilized at a given stage in cell maturation and activation.
The contribution of distinct plasminogen receptors to macrophage recruitment may also be tissue- and stimulus- specific. For example in a model of monocyte recruitment to the alveolar compartment, α-enolase appears to play a predominant role (Wygrecka, Marsh, Morty, Henneke, Guenther, Lohmeyer, Markart & Preissner, 2009). α-enolase also plays a role in inflammatory cell infiltration required for muscle repair after injury (Diaz-Ramos et al., 2012).
10. Conclusions
Over the past 25 years, a broad spectrum of experimental approaches and areas of investigation have demonstrated the wide array of cellular events and functions that are mediated by the interaction of plasminogen with it cellular receptors. Here we have reviewed emerging data establishing a role for plasminogen, plasminogen receptors and specifically, the newly discovered plasminogen receptor, Plg-RKT, in macrophage recruitment in the inflammatory response. A proteomics approach has allowed us to identify a new protein, Plg-RKT, a novel plasminogen receptor with unique characteristics: integral to the cell membrane and exposing a C-terminal lysine on the cell surface in an orientation to bind plasminogen. Furthermore, the ability of Plg-RKT to bind tPA, as well as the colocalization of Plg-RKT with uPAR, bring the substrate, plasminogen, and its activators in close proximity on the cell surface in an orientation to promote plasminogen activation as shown in the model in Figure 9. Future studies with knockout mice should build on our initial results and further elucidate the role of Plg-RKT and the interplay with other plasminogen receptors in specific steps in the macrophage inflammatory response.
Figure 9. Model for Plg-RKT-dependent cell surface plasminogen activation.

Plg-RKT is located on the monocyte surface in close physical proximity to the uPAR. The uPAR brings uPA in close proximity to plasminogen bound to Plg-RKT, thus promoting activation of the bound plasminogen to plasmin. In addition, tPA also interacts specifically with Plg-RKT, thus mimicking the interaction of tPA with cellular binding sites. Despite sharing a binding site on Plg-RKT, the relative concentrations of tPA and plasminogen in the circulation should permit simultaneous binding of both ligands to the cell surface, and each tPA molecule should be bound proximally to several plasminogen molecules, thus promoting plasminogen activation to plasmin on the cell surface. Reprinted from Miles, Lighvani, Baik, Andronicos, Chen, Parmer, Khaldoyanidi, Diggs, Kiosses, Kamps, Yates, III & Parmer, 2012b, according Journal of Biomedicine and Biotechnology policies.
Acknowledgments
Supported by National Institutes of Health Grants (HL38272, H45934, and HL081046 to L.A.M., CA166473 to B.M. and L.A.M., T32 HL007195 to S.L., HL50398 to R.J.P., and Department of Veterans Affairs to R.J.P. We thank Ms. Linda Bonafede for manuscript =preparation. This is manuscript #25011 from The Scripps Research Institute.
Reference List
- Andronicos NM, Chen EI, Baik N, Bai H, Parmer CM, Kiosses WB, Kamps MP, Yates JR, III, Parmer RJ, Miles LA. Proteomics-based discovery of a novel, structurally unique, and developmentally regulated plasminogen receptor, Plg-RKT, a major regulator of cell surface plasminogen activation. Blood. 2010;115:1319–1330. doi: 10.1182/blood-2008-11-188938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Astrup T, STERNDORFF I. An activator of plasminogen in normal urine. Proc Soc Exp Biol Med. 1952;81:675–678. doi: 10.3181/00379727-81-19983. [DOI] [PubMed] [Google Scholar]
- Bai H, Baik N, Kiosses WB, Krajewski S, Miles LA, Parmer RJ. The novel plasminogen receptor, plasminogen receptor(KT) (Plg-R(KT)), regulates catecholamine release. J Biol Chem. 2011;286:33125–33133. doi: 10.1074/jbc.M111.218693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bai H, Nangia S, Parmer RJ. The plasminogen activation system and the regulation of catecholaminergic function. J Biomed Biotechnol. 2012;2012:721657. doi: 10.1155/2012/721657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bannach FG, Gutierrez A, Fowler BJ, Bugge TH, Degen JL, Parmer RJ, Miles LA. Localization of regulatory elements mediating constitutive and cytokine-stimulated plasminogen gene expression. J Biol Chem. 2002;277:38579–38588. doi: 10.1074/jbc.M202509200. [DOI] [PubMed] [Google Scholar]
- Bannach FG, Gutierrez-Fernandez A, Parmer RJ, Miles LA. Interleukin-6-induced plasminogen gene expression in murine hepatocytes is mediated by transcription factor CCAAT/enhancer binding protein beta (C/EBPbeta) J Thromb Haemost. 2004;2:2205–2212. doi: 10.1111/j.1538-7836.2004.01022.x. [DOI] [PubMed] [Google Scholar]
- Barth MW, Hendrzak JA, Melnicoff MJ, Morahan PS. Review of the macrophage disappearance reaction. J Leukoc Biol. 1995;57:361–367. doi: 10.1002/jlb.57.3.361. [DOI] [PubMed] [Google Scholar]
- Basham ME, Seeds NW. Plasminogen expression in the neonatal and adult mouse brain. J Neurochem. 2001;77:318–325. doi: 10.1046/j.1471-4159.2001.t01-1-00239.x. [DOI] [PubMed] [Google Scholar]
- Bhattacharya S, Ploplis VA, Castellino FJ. Bacterial plasminogen receptors utilize host plasminogen system for effective invasion and dissemination. J Biomed Biotechnol. 2012;2012:482096. doi: 10.1155/2012/482096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bohmfalk JF, Fuller GM. Plasminogen is synthesized by primary cultures of rat hepatocytes. Science. 1980;209:408–410. doi: 10.1126/science.7384814. [DOI] [PubMed] [Google Scholar]
- Bugge TH, Flick MJ, Daugherty CC, Degen JL. Plasminogen deficiency causes severe thrombosis but is compatible with development and reproducton. Genes and Development. 1995;9:794–807. doi: 10.1101/gad.9.7.794. [DOI] [PubMed] [Google Scholar]
- Bugge TH, Kombrinck KW, Flick MJ, Daugherty CC, Danton MJS, Degen JL. Loss of fibrinogen rescues mice from the pleiotropic effects of plasminogen deficiency. Cell. 1996;87:709–719. doi: 10.1016/s0092-8674(00)81390-2. [DOI] [PubMed] [Google Scholar]
- Burysek L, Syrovets T, Simmet T. The serine protease plasmin triggers expression of MCP-1 and CD40 in human primary monocytes via activation of p38 MAPK and janus kinase (JAK)/STAT signaling pathways. J Biol Chem. 2002;277:33509–33517. doi: 10.1074/jbc.M201941200. [DOI] [PubMed] [Google Scholar]
- Busuttil SJ, Ploplis VA, Castellino FJ, Tang L, Eaton JW, Plow EF. A central role for plasminogen in the inflammatory response to biomaterials. J Thromb Haemost. 2004;2:1798–1805. doi: 10.1111/j.1538-7836.2004.00916.x. [DOI] [PubMed] [Google Scholar]
- Callahan MK, Halleck MS, Krahling S, Henderson AJ, Williamson P, Schlegel RA. Phosphatidylserine expression and phagocytosis of apoptotic thymocytes during differentiation of monocytic cells. J Leukoc Biol. 2003;74:846–856. doi: 10.1189/jlb.0902433. [DOI] [PubMed] [Google Scholar]
- Cao C, Lawrence DA, Li Y, Von Arnim CA, Herz J, Su EJ, Makarova A, Hyman BT, Strickland DK, Zhang L. Endocytic receptor LRP together with tPA and PAI-1 coordinates Mac-1-dependent macrophage migration. EMBO J. 2006;25:1860–1870. doi: 10.1038/sj.emboj.7601082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmeliet P, Schoonjans L, Kieckens L, Ream B, Degen J, Bronson R, De Vos R, Van den Oord JJ, Collen D, Mulligan RC. Physiological consequences of loss of plasminogen activator gene function in mice. Nature. 1994;368:419–424. doi: 10.1038/368419a0. [DOI] [PubMed] [Google Scholar]
- Castellino FJ, Ploplis VA. Structure and function of the plasminogen/plasmin system. Thromb Haemost. 2005;93:647–654. doi: 10.1160/TH04-12-0842. [DOI] [PubMed] [Google Scholar]
- Choi KS, Fogg DK, Yoon CS, Waisman DM. p11 regulates extracellular plasmin production and invasiveness of HT1080 fibrosarcoma cells. FASEB J. 2003;17:235–246. doi: 10.1096/fj.02-0697com. [DOI] [PubMed] [Google Scholar]
- Christensen LR. STREPTOCOCCAL FIBRINOLYSIS: A PROTEOLYTIC REACTION DUE TO A SERUM ENZYME ACTIVATED BY STREPTOCOCCAL FIBRINOLYSIN. J Gen Physiol. 1945;28:363–383. doi: 10.1085/jgp.28.4.363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collen D. The plasminogen (fibrinolytic) system. Thromb Haemost. 1999;82:259–270. [PubMed] [Google Scholar]
- Collen D, Verstraete M. Molecular biology of human plasminogen. II Metabolism in physiological and some pathophysiological conditions in man. Thromb Diath Haemorrh Suppl. 1975;34:403–408. [PubMed] [Google Scholar]
- Colman RW. Activation of plasminogen by human plasma kallikrein. Biochem Biophys Res Commun. 1969;35:273–279. doi: 10.1016/0006-291x(69)90278-2. [DOI] [PubMed] [Google Scholar]
- Connolly BM, Choi EY, Gardsvoll H, Bey AL, Currie BM, Chavakis T, Liu S, Molinolo A, Ploug M, Leppla SH, Bugge TH. Selective abrogation of the uPA-uPAR interaction in vivo reveals a novel role in suppression of fibrin-associated inflammation. Blood. 2010;116:1593–1603. doi: 10.1182/blood-2010-03-276642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cook AD, Vlahos R, Massa CM, Braine EL, Lenzo JC, Turner AL, Way KJ, Hamilton JA. The effect of tissue type-plasminogen activator deletion and associated fibrin(ogen) deposition on macrophage localization in peritoneal inflammation. Thromb Haemost. 2006;95:659–667. [PubMed] [Google Scholar]
- Correc P, Fondanèche M-C, Bracke M, Burtin P. The presence of plasmin receptors on three mammary carcinoma MCF-7 sublines. Int J Cancer. 1990;46:745–750. doi: 10.1002/ijc.2910460432. [DOI] [PubMed] [Google Scholar]
- Creemers E, Cleutjens J, Smits J, Heymans S, Moons L, Collen D, Daemen M, Carmeliet P. Disruption of the plasminogen gene in mice abolishes wound healing after myocardial infarction. Am J Pathol. 2000;156:1865–1873. doi: 10.1016/S0002-9440(10)65060-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das R, Burke T, Plow EF. Histone H2B as a functionally important plasminogen receptor on macrophages. Blood. 2007;110:3763–3772. doi: 10.1182/blood-2007-03-079392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das R, Plow EF. Phosphatidylserine as an anchor for plasminogen and its plasminogen receptor, histone H2B, to the macrophage surface. J Thromb Haemost. 2011;9:339–349. doi: 10.1111/j.1538-7836.2010.04132.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dassah M, Deora AB, He K, Hajjar KA. The endothelial cell annexin A2 system and vascular fibrinolysis. Gen Physiol Biophys. 2009;28:F20–F28. Spec No Focus. [PMC free article] [PubMed] [Google Scholar]
- Davidson DJ, Higgins DL, Castellino FJ. Plasminogen activator activities of equimolar complexes of streptokinase with variant recombinant plasminogens. Biochemistry. 1990;29:3585–3590. doi: 10.1021/bi00466a023. [DOI] [PubMed] [Google Scholar]
- Dejouvencel T, Doeuvre L, Lacroix R, Plawinski L, Dignat-George F, Lijnen HR, Angles-Cano E. Fibrinolytic cross-talk: a new mechanism for plasmin formation. Blood. 2010;115:2048–2056. doi: 10.1182/blood-2009-06-228817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz-Ramos A, Roig-Borrellas A, Garcia-Melero A, Lopez-Alemany R. alpha-Enolase, a Multifunctional Protein: Its Role on Pathophysiological Situations. J Biomed Biotechnol. 2012;2012:156795. doi: 10.1155/2012/156795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drew AF, Kaufman AH, Kombrinck KW, Danton MJS, Daugherty CC, Degen JL, Bugge TH. Ligneous conjunctivitis in plasminogen-deficient mice. Blood. 1998;91:1616–1624. [PubMed] [Google Scholar]
- Dudani AK, Ganz PR. Endothelial cell surface actin serves as a binding site for plasminogen, tissue plasminogen activator and lipoprotein(a) Br J Haematol. 1996;95:168–178. doi: 10.1046/j.1365-2141.1996.7482367.x. [DOI] [PubMed] [Google Scholar]
- Durliat M, Komano O, Correc P, Bertrand O, Cochet S, Caignard A, Martin F, Burtin P. Plasminogen receptors on rat colon carcinoma cells. Br J Cancer. 1992;66:51–56. doi: 10.1038/bjc.1992.215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duval-Jobe C, Parmely MJ. Regulation of plasminogen activation by human U937 promonocytic cells. J Biol Chem. 1994;269:21353–21357. [PubMed] [Google Scholar]
- Ellis V, Behrendt N, Dano K. Plasminogen activation by receptor-bound urokinase. A kinetic study with both cell-associated and isolated receptor. J Biol Chem. 1991;266:12752–12758. [PubMed] [Google Scholar]
- Eng JK, McCormick AL, Yates JRI. An approach to correlate tandem mass spectral data of peptides with amino acid sequences in a protein database. J Am Soc Mass Spectrom. 1994;5:976–989. doi: 10.1016/1044-0305(94)80016-2. [DOI] [PubMed] [Google Scholar]
- Eriksson PO, Li J, Ny T, Hellstrom S. Spontaneous development of otitis media in plasminogen-deficient mice. Int J Med Microbiol. 2006;296:501–509. doi: 10.1016/j.ijmm.2006.04.002. [DOI] [PubMed] [Google Scholar]
- Fan ZQ, Larson PJ, Bognacki J, Raghunath PN, Tomaszewski JE, Kuo A, Canziani G, Chaiken I, Cines DB, Higazi AAR. Tissue factor regulates plasminogen binding and activation. Blood. 1998;91:1987–1998. [PubMed] [Google Scholar]
- Felez J, Chanquias CJ, Fabregas P, Plow EF, Miles LA. Competition between plasminogen and tissue plasminogen activator for cellular binding sites. Blood. 1993;82:2433–2441. [PubMed] [Google Scholar]
- Felez J, Jardi M, Fabregas P, Parmer RJ, Miles LA. Monoclonal antibodies against receptor-induced binding sites detect cell-bound plasminogen in blood. Blood. 2012;120:678–681. doi: 10.1182/blood-2012-02-410480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Félez J, Miles LA, Fábregas P, Jardi M, Plow EF, Lijnen RH. Characterization of cellular binding sites and interactive regions within reactants required for enhancement of plasminogen activation by tPA on the surface of leukocytic cells. Thromb Haemost. 1996;76:577–584. [PubMed] [Google Scholar]
- Felez J, Miles LA, Fabregas P, Jardi M, Plow EF, Lijnen RJ. Characterization of cellular binding sites and interactive regions within reactants required for enhancement of plasminogen activation by tPA on the surface of leukocytic cells. Thromb Haemost. 1996;76:577–584. [PubMed] [Google Scholar]
- Ferguson JH. A NEW BLOOD-CLOTTING THEORY. Science. 1943;97:319–322. doi: 10.1126/science.97.2519.319. [DOI] [PubMed] [Google Scholar]
- Fleury V, Lijnen HR, Anglés-Cano E. Mechanism of the enhanced intrinsic activity of single-chain urokinase-type plasminogen activator during ongoing fibrinolysis. J Biol Chem. 1993;268:18554–18559. [PubMed] [Google Scholar]
- Forsgren M, Raden B, Israelsson M, Larsson K, Heden L-O. Molecular cloning and characterization of a full-length cDNA clone for human plasminogen. FEBS Lett. 1987;213:254–260. doi: 10.1016/0014-5793(87)81501-6. [DOI] [PubMed] [Google Scholar]
- Gong Y, Hart E, Shchurin A, Hoover-Plow J. Inflammatory macrophage migration requires MMP-9 activation by plasminogen in mice. J Clin Invest. 2008;118:3012–3024. doi: 10.1172/JCI32750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gong Y, Kim S-O, Felez J, Grella DK, Castellino FJ, Miles LA. Conversion of glu-plasminogen to lys-plasminogen is necessary for optimal stimulation of plasminogen activation on the endothelial cell surface. J Biol Chem. 2001;276:19078–19083. doi: 10.1074/jbc.M101387200. [DOI] [PubMed] [Google Scholar]
- Gonias SL, Braud LL, Geary WA, VandenBerg SR. Plasminogen binding to rat hepatocytes in primary culture and to thin slices of rat liver. Blood. 1989;74:729–736. [PubMed] [Google Scholar]
- Gonzalez-Gronow M, Gawdi G, Pizzo SV. Plasminogen activation stimulates an increase in intracellular calcium in human synovial fibroblasts. J Biol Chem. 1993;268:20791–20795. [PubMed] [Google Scholar]
- Gonzalez-Gronow M, Gawdi G, Pizzo SV. Characterization of the plasminogen receptors of normal and rheumatoid arthritis human synovial fibroblasts. J Biol Chem. 1994;269:4360–4366. [PubMed] [Google Scholar]
- Gonzalez-Gronow M, Grenett HE, Fuller GM, Pizzo SV. The role of carbohydrate in the function of human plasminogen: Comparison of the protein obtained from molecular cloning and expression in Escherichia coli and COS cells. Biochim Biophys Acta Protein Struct Mol Enzymol. 1990;1039:269–276. doi: 10.1016/0167-4838(90)90259-i. [DOI] [PubMed] [Google Scholar]
- Gordon S. Development and distribution of mononuclear phagocytes: relevance to inflammation. In: Gallin I, Snyderman R, editors. Inflammation: Basic Principles and Clinical Correlates. Lippincott Williams & Wilkins; Philadelphia: 1999. pp. 35–48. [Google Scholar]
- Gutierrez-Fernandez A, Gingles NA, Bai H, Castellino FJ, Parmer RJ, Miles LA. Plasminogen enhances neuritogenesis on laminin-1. J Neurosci. 2009;29:12393–12400. doi: 10.1523/JNEUROSCI.3553-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutierrez-Fernandez A, Gingles NA, Zhang L, Garcia Bannach F, Jenkins GR, Loskutoff DJ, Parmer RJ, Miles LA. Regulation of plasminogen gene expression. In: Waisman DM, editor. Plasminogen Regulation. Academic/Plenum Publishers; New York: 2003. pp. 67–80. [Google Scholar]
- Hajjar KA, Hamel NM, Harpel PC, Nachman RL. Binding of tissue plasminogen activator to cultured human endothelial cells. J Clin Invest. 1987;80:1712–1719. doi: 10.1172/JCI113262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajjar KA, Harpel PC, Jaffe EA, Nachman RL. Binding of plasminogen to cultured human endothelial cells. J Biol Chem. 1986;261:11656–11662. [PubMed] [Google Scholar]
- Hajjar KA, Jacovina AT, Chacko J. An endothelial cell receptor for plasminogen/tissue plasminogen activator. I Identity with annexin II. J Biol Chem. 1994;269:21191–21197. [PubMed] [Google Scholar]
- Hall SW, Humphries JE, Gonias SL. Inhibition of cell surface receptor-bound plasmin by a2- antiplasmin and a2-macroglobulin. J Biol Chem. 1991;266:12329–12336. [PubMed] [Google Scholar]
- Han J, Baik N, Kim K-H, Yang J-MHG, Gong Y, Jardi M, Felez J, Castellino FJ, Parmer RJ, Miles LA. Monoclonal antibodies detect receptor-induced binding sites (RIBS) in Glu-plasminogen. Blood Under Revision. 2011a doi: 10.1182/blood-2010-11-316943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Han J, Baik N, Kim KH, Yang JM, Han GW, Gong Y, Jardi M, Castellino FJ, Felez J, Parmer RJ, Miles LA. Monoclonal antibodies detect receptor-induced binding sites in Glu-plasminogen. Blood. 2011b;118:1653–1662. doi: 10.1182/blood-2010-11-316943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hasumi K, Yamamichi S, Harada T. Small-molecule modulators of zymogen activation in the fibrinolytic and coagulation systems. FEBS J. 2010;277:3675–3687. doi: 10.1111/j.1742-4658.2010.07783.x. [DOI] [PubMed] [Google Scholar]
- Hawley SB, Tamura T, Miles LA. Purification, cloning, and characterization of a profibrinolytic plasminogen-binding protein, TIP49a. J Biol Chem. 2001;276:179–186. doi: 10.1074/jbc.M004919200. [DOI] [PubMed] [Google Scholar]
- Hayes ML, Castellino FJ. Carbohydrate of the human plasminogen variants. II Structure of the asparagine-linked oligosaccharide unit. J Biol Chem. 1979a;254:8772–8776. [PubMed] [Google Scholar]
- Hayes ML, Castellino FJ. Carbohydrate of the human plasminogen variants. III Structure of the O-glycosidically linked oligosaccharide unit. J Biol Chem. 1979b;254:8777–8780. [PubMed] [Google Scholar]
- Hembrough TA, Kralovich KR, Li L, Gonias SL. Cytokeratin 8 released by breast carcinoma cells in vitro binds plasminogen and tissue-type plasminogen activator and promotes plasminogen activation. Biochem J. 1996;317:763–769. doi: 10.1042/bj3170763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hembrough TA, Vasudevan J, Allietta MM, Glass WF, Gonias SL. A cytokeratin 8-like protein with plasminogen-binding activity is present on the external surfaces of hepatocytes, HepG2 cells and breast carcinoma cell lines. J Cell Sci. 1995;108 (Pt 3):1071–1082. doi: 10.1242/jcs.108.3.1071. [DOI] [PubMed] [Google Scholar]
- Herren T, Burke TA, Das R, Plow EF. Identification of histone H2B as a regulated plasminogen receptor. Biochemistry. 2006;45:9463–9474. doi: 10.1021/bi060756w. [DOI] [PubMed] [Google Scholar]
- Higgins DL, Vehar GA. Interaction of one-chain and two-chain tissue plasminogen activator with intact and plasmin-degraded fibrin. Biochemistry. 1987;26:7786–7791. doi: 10.1021/bi00398a038. [DOI] [PubMed] [Google Scholar]
- Hoover-Plow J, Wang N, Ploplis VA. Growth and behavioral develoment in plaslminogen gene-targeted mice. Growth, Development and Aging. 1999;63:13–32. [PubMed] [Google Scholar]
- Horrevoets AJG, Smilde AE, Fredenburgh JC, Pannekoek H, Nesheim ME. The activation-resistant conformation of recombinant human plasminogen is stabilized by basic residues in the amino-terminal hinge region. J Biol Chem. 1995;270:15770–15776. doi: 10.1074/jbc.270.26.15770. [DOI] [PubMed] [Google Scholar]
- Hoylaerts M, Rijken DC, Lijnen HR, Collen D. Kinetics of the activation of plasminogen by human tissue plasminogen activator. J Biol Chem. 1982;257:2912–2919. [PubMed] [Google Scholar]
- Iwaki T, Malinverno C, Smith D, Xu Z, Liang Z, Ploplis VA, Castellino FJ. The generation and characterization of mice expressing a plasmin-inactivating active site mutation. J Thromb Haemost. 2010;8:2341–2344. doi: 10.1111/j.1538-7836.2010.03995.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacovina AT, Zhong F, Khazanova E, Lev E, Deora AB, Hajjar KA. Neuritogenesis and the nerve growth factor-induced differentiation of PC-12 cells requires annexin II-mediated plasmin generation. J Biol Chem. 2001;276:49350–49358. doi: 10.1074/jbc.M106289200. [DOI] [PubMed] [Google Scholar]
- Jardi M, Fabregas P, Sagarra-Tio M, Perez-Lucena MJ, Felez J. Characterization of Plasminogen Binding to NB4 Promyelocytic Cells Using Monoclonal Antibodies against Receptor-Induced Binding Sites in Cell-Bound Plasminogen. J Biomed Biotechnol. 2012;2012:984589. doi: 10.1155/2012/984589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkins GR, Seiffert D, Parmer RJ, Miles LA. Regulation of plasminogen gene expression by interleukin- 6. Blood. 1997;89:2394–2403. [PubMed] [Google Scholar]
- Jiang Q, Taupenot L, Mahata SK, Mahata M, O’Connor DT, Miles LA, Parmer RJ. Proteolytic cleavage of chromogranin A (CgA) by plasmin. Selective liberation of a specific bioactive CgA fragment that regulates catecholamine release. J Biol Chem. 2001;276:25022–25029. doi: 10.1074/jbc.M101545200. [DOI] [PubMed] [Google Scholar]
- Jiang Q, Yasothornsrikul S, Taupenot L, Miles LA, Parmer RJ. The local chromaffin cell plasminogen/plasmin system and the regulation of catecholamine secretion. Ann N Y Acad Sci. 2002;971:445–449. doi: 10.1111/j.1749-6632.2002.tb04506.x. [DOI] [PubMed] [Google Scholar]
- Kanalas JJ. Analysis of plasmin binding and urokinase activation of plasminogen bound to the Heymann nephritis autoantigen, gp330. Arch Biochem Biophys. 1992;299:255–260. doi: 10.1016/0003-9861(92)90272-x. [DOI] [PubMed] [Google Scholar]
- Kanalas JJ, Makker SP. Identification of the rat Heymann nephritis autoantigen (GP330) as a receptor site for plasminogen. J Biol Chem. 1991;266:10825–10829. [PubMed] [Google Scholar]
- Kaplan MH. Studies of streptococcal fibrinolysis; the dissimilarity of serum protease and trypsin as indicated by the separate specificities of their kinases, fibrinolysin and enterokinase. J Clin Invest. 1946;25:331–336. [PubMed] [Google Scholar]
- Kassam G, Le BH, Choi KS, Kang HM, Fitzpatrick SL, Louie P, Waisman DM. The p11 subunit of the annexin II tetramer plays a key role in the stimulation of tPA-dependent plasminogen activation. Biochemistry. 1998;37:16958–16966. doi: 10.1021/bi981713l. [DOI] [PubMed] [Google Scholar]
- Kilpatrick LM, Harris RL, Owen KA, Bass R, Ghorayeb C, Bar-Or A, Ellis V. Initiation of plasminogen activation on the surface of monocytes expressing the type II transmembrane serine protease matriptase. Blood. 2006;108:2616–2623. doi: 10.1182/blood-2006-02-001073. [DOI] [PubMed] [Google Scholar]
- Klammt J, Kobelt L, Aktas D, Durak I, Gokbuget A, Hughes Q, Irkec M, Kurtulus I, Lapi E, Mechoulam H, Mendoza-Londono R, Palumbo JS, Steitzer H, Tabbara KF, Ozbek Z, Pucci N, Sotomayor T, Sturm M, Drogies T, Ziegler M, Schuster V. Identification of three novel plasminogen (PLG) gene mutations in a series of 23 patients with low PLG activity. Thromb Haemost. 2011;105:454–460. doi: 10.1160/TH10-04-0216. [DOI] [PubMed] [Google Scholar]
- Kuhn JF, Godshall CJ, Scott MJ, Franklin GA, Rowe SA, Peyton JC, Cheadle WG. Immediate and delayed leukocyte apoptosis in two models of peritonitis. Inflammation. 2001;25:389–397. doi: 10.1023/a:1012854731259. [DOI] [PubMed] [Google Scholar]
- Kurtulus I, Kazmi B, Lewis MP, Solakoglu S, Schuster V, Scully C, Gokbuget A, Wahl G. Altered functional activity patterns of fibroblasts related to periodontitis by systemic plasminogen deficiency (ligneous periodontitis) Quintessence Int. 2011;42:601–609. [PubMed] [Google Scholar]
- Lacroix R, Plawinski L, Robert S, Doeuvre L, Sabatier F, Martinez dL, Mezzapesa A, Anfosso F, Leroyer A, Poullin P, Jourde N, Njock MS, Boulanger C, Angles-Cano E, Dignat-George F. Leukocyte- and endothelial-derived microparticles: a circulating source for fibrinolysis. Haematologica. 2012 doi: 10.3324/haematol.2012.066167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacroix R, Sabatier F, Mialhe A, Basire A, Pannell R, Borghi H, Robert S, Lamy E, Plawinski L, Camoin-Jau L, Gurewich V, Angles-Cano E, Dignat-George F. Activation of plasminogen into plasmin at the surface of endothelial microparticles: a mechanism that modulates angiogenic properties of endothelial progenitor cells in vitro. Blood. 2007;110:2432–2439. doi: 10.1182/blood-2007-02-069997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Law RH, Caradoc-Davies T, Cowieson N, Horvath AJ, Quek AJ, Encarnacao JA, Steer D, Cowan A, Zhang Q, Lu BG, Pike RN, Smith AI, Coughlin PB, Whisstock JC. The X-ray crystal structure of full-length human plasminogen. Cell Rep. 2012;1:185–190. doi: 10.1016/j.celrep.2012.02.012. [DOI] [PubMed] [Google Scholar]
- Leak LV. Interaction of mesothelium to intraperitoneal stimulation. I Aggregation of peritoneal cells. Lab Invest. 1983;48:479–491. [PubMed] [Google Scholar]
- Leibovich SJ, Ross R. The role of the macrophage in wound repair. A study with hydrocortisone and antimacrophage serum. Am J Pathol. 1975;78:71–100. [PMC free article] [PubMed] [Google Scholar]
- Li X, Syrovets T, Simmet T. The Serine Protease Plasmin Triggers Expression of the CC-Chemokine Ligand 20 in Dendritic Cells via Akt/NF-kappaB-Dependent Pathways. J Biomed Biotechnol. 2012;2012:186710. doi: 10.1155/2012/186710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lighvani S, Baik N, Diggs JE, Khaldoyanidi S, Parmer RJ, Miles LA. Regulation of macrophage migration by a novel plasminogen receptor Plg-R KT. Blood. 2011;118:5622–5630. doi: 10.1182/blood-2011-03-344242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lijnen HR, Van Hoef B, Nelles L, Collen D. Plasminogen activation with single-chain urokinase-type plasminogen activator (scuPA). Studies with active site mutagenized plasminogen (Ser740-->Ala) and plasmin-resistant scuPA (Lys158-->Glu) J Biol Chem. 1990;265:5232–5236. [PubMed] [Google Scholar]
- Ling Q, Jacovina AT, Deora A, Febbraio M, Simantov R, Silverstein RL, Hempstead B, Mark WH, Hajjar KA. Annexin II regulates fibrin homeostasis and neoangiogenesis in vivo. J Clin Invest. 2004;113:38–48. doi: 10.1172/JCI200419684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liotta LA, Goldfarb RH, Brundage R, Siegal GP, Terranova V, Garbisa S. Effect of plasminogen activator (urokinase), plasmin, and thrombin on glycoprotein and collagenous components of basement membrane. Cancer Res. 1981a;41:4629–4636. [PubMed] [Google Scholar]
- Liotta LA, Goldfarb RH, Terranova VP. Cleavage of laminin by thrombin and plasmin: Alpha thrombin selectively cleaves the bet a chain of laminin. Thromb Res. 1981b;21:663–673. doi: 10.1016/0049-3848(81)90268-1. [DOI] [PubMed] [Google Scholar]
- Lishko VK, Novokhatny VV, Yakubenko VP, Skomorovska-Prokvolit HV, Ugarova TP. Characterization of plasminogen as an adhesive ligand for integrins {alpha}M{beta}2 (Mac-1) and {alpha}5{beta}1(VLA-5) Blood. 2004 doi: 10.1182/blood-2003-09-3016. [DOI] [PubMed] [Google Scholar]
- Liu M, Zhang S. A kringle-containing protease with plasminogen-like activity in the basal chordate Branchiostoma belcheri. Biosci Rep. 2009;29:385–395. doi: 10.1042/BSR20080173. [DOI] [PubMed] [Google Scholar]
- Longstaff C, Merton RE, Fabregas P, Felez J. Characterization of cell-associated plasminogen activation catalyzed by urokinase-type plasminogen activator, but independent of urokinase receptor (uPAR, CD87) Blood. 1999;93:3839–3846. [PubMed] [Google Scholar]
- Lopez-Alemany R, Suelves M, Diaz-Ramos A, Vidal B, Munoz-Canoves P. Alpha-enolase plasminogen receptor in myogenesis. Front Biosci. 2005;10:30–36. doi: 10.2741/1503. [DOI] [PubMed] [Google Scholar]
- Lopez-Alemany R, Suelves M, Munoz-Canoves P. Plasmin generation dependent on alpha-enolase-type plasminogen receptor is required for myogenesis. Thromb Haemost. 2003;90:724–733. doi: 10.1160/TH03-04-0291. [DOI] [PubMed] [Google Scholar]
- Loscalzo J. Structural and kinetic comparison of recombinant human single- and two-chain tissue plasminogen activator. J Clin Invest. 1988;82:1391–1397. doi: 10.1172/JCI113743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loscalzo J, Vaughan DE. Tissue plasminogen activator promotes platelet disaggregation in plasma. J Clin Invest. 1987;79:1749–1755. doi: 10.1172/JCI113015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lund LR, Bjorn SF, Sternlicht MD, Nielsen BS, Solberg H, Usher PA, Osterby R, Christensen IJ, Stephens RW, Bugge TH, Dano K, Werb Z. Lactational competence and involution of the mouse mammary gland require plasminogen. Development. 2000;127:4481–4492. doi: 10.1242/dev.127.20.4481. [DOI] [PubMed] [Google Scholar]
- Madureira PA, Hill R, Miller VA, Giacomantonio C, Lee PW, Waisman DM. Annexin A2 is a novel cellular redox regulatory protein involved in tumorigenesis. Oncotarget. 2011a;2:1075–1093. doi: 10.18632/oncotarget.375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madureira PA, O’Connell PA, Surette AP, Miller VA, Waisman DM. The Biochemistry and Regulation of S100A10: A Multifunctional Plasminogen Receptor Involved in Oncogenesis. J Biomed Biotechnol. 2012;2012:353687. doi: 10.1155/2012/353687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Madureira PA, Surette AP, Phipps KD, Taboski MA, Miller VA, Waisman DM. The role of the annexin A2 heterotetramer in vascular fibrinolysis. Blood. 2011b;118:4789–4797. doi: 10.1182/blood-2011-06-334672. [DOI] [PubMed] [Google Scholar]
- Marguet D, Luciani MF, Moynault A, Williamson P, Chimini G. Engulfment of apoptotic cells involves the redistribution of membrane phosphatidylserine on phagocyte and prey. Nat Cell Biol. 1999;1:454–456. doi: 10.1038/15690. [DOI] [PubMed] [Google Scholar]
- Marshall JM, Brown AJ, Ponting CP. Conformational studies of human plasminogen and plasminogen fragments: Evidence for a novel third conformation of plasminogen. Biochemistry. 1994;33:3599–3606. doi: 10.1021/bi00178a017. [DOI] [PubMed] [Google Scholar]
- McCormack PL. Tranexamic acid: a review of its use in the treatment of hyperfibrinolysis. Drugs. 2012;72:585–617. doi: 10.2165/11209070-000000000-00000. [DOI] [PubMed] [Google Scholar]
- Mehart N, Sehl LC, Kelley RF, Castellino FJ. Construction, expression, and purification of recombinant kringle 1 of human plasminogen and analysis of its interaction with omega-amino acids. Biochemistry. 1991;19:1948–1957. doi: 10.1021/bi00221a031. [DOI] [PubMed] [Google Scholar]
- Menhart N, McCance SG, Sehl LC, Castellino FJ. Functional independence of the kringle 4 and kringle 5 regions of human plasminogen. Biochemistry. 1993;32:8799–8806. doi: 10.1021/bi00085a010. [DOI] [PubMed] [Google Scholar]
- Miles LA, Andronicos NM, Baik N, Parmer RJ. Cell-surface actin binds plasminogen and modulates neurotransmitter release from catecholaminergic cells. J Neurosci. 2006;26:13017–13024. doi: 10.1523/JNEUROSCI.2070-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miles LA, Andronicos NM, Chen EI, Baik N, Bai H, Parmer CM, Lighvani S, Nangia S, Kiosses WB, Kamps MP, Yates JR, III, Parmer RJ. Identification of the novel plasminogen receptor, Plg-RKT. In: Man TK, Flores RJ, editors. Proteomics/Book 1: Human Diseases and Protein Functions. Intech. 2012a. pp. 219–238. [Google Scholar]
- Miles LA, Castellino FJ, Gong Y. Critical role for conversion of glu-plasminogen to Lys-plasminogen for optimal stimulation of plasminogen activation on cell surfaces. Trends Cardiovasc Med. 2003;13:21–30. doi: 10.1016/s1050-1738(02)00190-1. [DOI] [PubMed] [Google Scholar]
- Miles LA, Dahlberg CM, Levin EG, Plow EF. Gangliosides interact directly with plasminogen and urokinase and may mediate binding of these components to cells. Biochemistry. 1989;280:9337–9343. doi: 10.1021/bi00450a014. [DOI] [PubMed] [Google Scholar]
- Miles LA, Dahlberg CM, Plescia J, Felez J, Kato K, Plow EF. Role of cell-surface lysines in plasminogen binding to cells: Identification of alpha-Enolase as a candidate plasminogen receptor. Biochemistry. 1991;30:1682–1691. doi: 10.1021/bi00220a034. [DOI] [PubMed] [Google Scholar]
- Miles LA, Dahlberg CM, Plow EF. The cell-binding domains of plasminogen and their function in plasma. J Biol Chem. 1988a;263:11928–11934. [PubMed] [Google Scholar]
- Miles LA, Ginsberg MH, White JG, Plow EF. Plasminogen interacts with human platelets through two distinct mechanisms. J Clin Invest. 1986;77:2001–2009. doi: 10.1172/JCI112529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miles LA, Greengard JS, Griffin JH. A comparison of the abilities of plasma kallikrein, beta-Factor XIIa, Factor XIa and urokinase to activate plasminogen. Thromb Res. 1983;29:407–417. doi: 10.1016/0049-3848(83)90244-x. [DOI] [PubMed] [Google Scholar]
- Miles LA, Hawley SB, Baik N, Andronicos NM, Castellino FJ, Parmer RJ. Plasminogen receptors: the sine qua non of cell surface plasminogen activation. Front Biosci. 2005;10:1754–1762. doi: 10.2741/1658. [DOI] [PubMed] [Google Scholar]
- Miles LA, Levin EG, Plescia J, Collen D, Plow EF. Plasminogen receptors, urokinase receptors and their modulation on human endothelial cells. Blood. 1988b;72:628–635. [PubMed] [Google Scholar]
- Miles LA, Lighvani S, Baik N, Andronicos NM, Chen EI, Parmer CM, Khaldoyanidi S, Diggs JE, Kiosses WB, Kamps MP, Yates JR, III, Parmer RJ. The Plasminogen Receptor, Plg-R(KT), and Macrophage Function. J Biomed Biotechnol. 2012b;2012:250464. doi: 10.1155/2012/250464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miles LA, Plow EF. Binding and activation of plasminogen on the platelet surface. J Biol Chem. 1985;260:4303–4311. [PubMed] [Google Scholar]
- Miles LA, Plow EF. Receptor mediated binding of the fibrinolytic components, plasminogen and urokinase, to peripheral blood cells. Thromb Haemost. 1987;58:936–942. [PubMed] [Google Scholar]
- Milstone H. A factor in normal human blood which participates in streptococcal fibrinolysis. J Immunol. 1941;42:109. [Google Scholar]
- Mingers AM, Heimburger N, Zeitler P, Kreth HW, Schuster V. Homozygous type I plasminogen deficiency. Semin Thromb Hemost. 1997;23:259–269. doi: 10.1055/s-2007-996099. [DOI] [PubMed] [Google Scholar]
- Mitchell JW, Baik N, Castellino FJ, Miles LA. Plasminogen inhibits TNF{alpha}-induced apoptosis in monocytes. Blood. 2006;107:4383–4390. doi: 10.1182/blood-2005-07-2872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy JA. Peritoneal membrane morphology and function. Kidney Int Suppl. 1996;56:S2–S11. [PubMed] [Google Scholar]
- Nelson DS. The effects of anticoagulants and other drugs on cellular and cutaneous reactions to antigen in guinea-pigs with delayed-type hypersensitivity. Immunology. 1965;9:219–234. [PMC free article] [PubMed] [Google Scholar]
- O’Connell PA, Surette AP, Liwski RS, Svenningsson P, Waisman DM. S100A10 regulates plasminogen-dependent macrophage invasion. 2010a doi: 10.1182/blood-2010-01-264754. [DOI] [PubMed] [Google Scholar]
- O’Connell PA, Surette AP, Liwski RS, Svenningsson P, Waisman DM. S100A10 regulates plasminogen-dependent macrophage invasion. Blood. 2010b;116:1136–1146. doi: 10.1182/blood-2010-01-264754. [DOI] [PubMed] [Google Scholar]
- O’Mullane MJ, Baker MS. Loss of cell viability dramatically elevates cell surface plasminogen binding and activation. Exp Cell Res. 1998;242:153–164. doi: 10.1006/excr.1998.4067. [DOI] [PubMed] [Google Scholar]
- O’Mullane MJ, Baker MS. Elevated plasminogen receptor expression occurs as a degradative phase even in cellular apoptosis. Immunol Cell Biol. 1999;77:249–255. doi: 10.1046/j.1440-1711.1999.00823.x. [DOI] [PubMed] [Google Scholar]
- Okada K, Ueshima S, Tanaka M, Fukao H, Matsuo O. Analysis of plasminogen activation by the plasmin-staphylokinase complex in plasma of alpha2-antiplasmin-deficient mice. Blood Coagul Fibrinolysis. 2000;11:645–655. doi: 10.1097/00001721-200010000-00009. [DOI] [PubMed] [Google Scholar]
- Palumbo JS, Talmage KE, Liu H, La Jeunesse CM, Witte DP, Degen JL. Plasminogen supports tumor growth through a fibrinogen-dependent mechanism linked to vascular patency. Blood. 2003;102:2819–2827. doi: 10.1182/blood-2003-03-0881. [DOI] [PubMed] [Google Scholar]
- Parkkinen J, Raulo E, merenmies J, Nolo R, Kajander EO, Baumann M, Rauvala H. Amphoterin, the 30-kDa protein in a family of HMG1-type polypeptides. Enhanced expression in transformed cells, leading edge localization, and interactions with plasminogen activation. J Biol Chem. 1993;268:19726–19738. [PubMed] [Google Scholar]
- Parmer RJ, Mahata M, Gong Y, Mahata SK, Jiang Q, O’Connor DT, Xi XP, Miles LA. Processing of chromogranin A by plasmin provides a novel mechanism for regulating catecholamine secretion. J Clin Invest. 2000;106:907–915. doi: 10.1172/JCI7394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parmer RJ, Miles LA. Targeting of tissue plasminogen activator to the regulated pathway of secretion. Trends Cardiovasc Med. 1998;8:306–312. doi: 10.1016/s1050-1738(98)00025-5. [DOI] [PubMed] [Google Scholar]
- Ploplis VA. Gene targeting in hemostasis plasminogen. Front Biosci. 2001;6:D555–D569. doi: 10.2741/ploplis. [DOI] [PubMed] [Google Scholar]
- Ploplis V, Carmeliet P, Vazirzadeh S, Van Vlanenderen I, Plow E, Collen D. Inactivation of the murine plasminogen gene. Physiological consequences. 1995a:1420. [Google Scholar]
- Ploplis VA, Carmeliet P, Vazirzadeh S, Van VI, Moons L, Plow EF, Collen D. Effects of disruption of the plasminogen gene on thrombosis, growth, and health in mice. Circulation. 1995b;92:2585–2593. doi: 10.1161/01.cir.92.9.2585. [DOI] [PubMed] [Google Scholar]
- Ploplis VA, Castellino FJ. Gene targeting of components of the fibrinolytic system. Thromb Haemost. 2002;87:22–31. [PubMed] [Google Scholar]
- Ploplis VA, French EL, Carmeliet P, Collen D, Plow EF. Plasminogen deficiency differentially affects recruitment of inflammatory cell populations in mice. Blood. 1998;91:2005–2009. [PubMed] [Google Scholar]
- Plow EF, Doeuvre L, Das R. So many plasminogen receptors: why? J Biomed Biotechnol. 2012;2012:141806. doi: 10.1155/2012/141806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plow EF, Freaney DE, Plescia J, Miles LA. The plasminogen system and cell surfaces: Evidence for plasminogen and urokinase receptors on the same cell type. J Cell Biol. 1986;103:2411–2420. doi: 10.1083/jcb.103.6.2411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plow EF, Ploplis VA, Busuttil S, Carmeliet P, Collen D. A role of plasminogen in atherosclerosis and restenosis models in mice. Thromb Haemost. 1999;82(Suppl 1):4–7. [PubMed] [Google Scholar]
- Pluskota E, Soloviev DA, Bdeir K, Cines DB, Plow EF. Integrin alphaMbeta2 orchestrates and accelerates plasminogen activation and fibrinolysis by neutrophils. J Biol Chem. 2004;279:18063–18072. doi: 10.1074/jbc.M310462200. [DOI] [PubMed] [Google Scholar]
- Ramos-DeSimone N, Hahn-Dantona E, Sipley J, Nagase H, French DL, Quigley JP. Activation of matrix metalloproteinase-9 (MMP-9) via a converging plasmin/stromelysin-1 cascade enhances tumor cell invasion. J Biol Chem. 1999;274:13066–13076. doi: 10.1074/jbc.274.19.13066. [DOI] [PubMed] [Google Scholar]
- Ranson M, Andronicos NM, O’Mullane MJ, Baker MS. Increased plasminogen binding is associated with metastatic breast cancer cells: differential expression of plasminogen binding proteins. Br J Cancer. 1998;77:1586–1597. doi: 10.1038/bjc.1998.261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rattner JB, Martin L, Waisman DM, Johnstone SA, Fritzler MJ. Autoantibodies to the centrosome (centriole) react with determinants present in the glycolytic enzyme enolase. J Immunol. 1991;146:2341–2344. [PubMed] [Google Scholar]
- Redlitz A, Fowler BJ, Plow EF, Miles LA. The role of an enolase-related molecule in plasminogen binding to cells. Eur J Biochem. 1995;227:407–415. doi: 10.1111/j.1432-1033.1995.tb20403.x. [DOI] [PubMed] [Google Scholar]
- REMMERT LF, COHEN PP. Partial purification and properties of a proteolytic enzyme of human serum. J Biol Chem. 1949;181:431–448. [PubMed] [Google Scholar]
- Rijken DC, Hoylaerts M, Collen D. Fibrinolytic properties of one-chain and two-chain extrinsic (tissue-type) plasminogen activator. J Biol Chem. 1982;257:2920–2925. [PubMed] [Google Scholar]
- Robbins KC, Summaria L, Hsieh B, Shah RJ. The peptide chains of human plasmin. Mechanism of activation of human plasminogen to plasmin. J Biol Chem. 1967;242:2333–2342. [PubMed] [Google Scholar]
- Romer J, Bugge TH, Pyke C, Lund LR, Flick MJ, Degen JL, Dano K. Impaired wound healing in mice with a disrupted plasminogen gene. Nat Med. 1996;2:287–292. doi: 10.1038/nm0396-287. [DOI] [PubMed] [Google Scholar]
- Ross M, Gerke V, Steinem C. Membrane composition affects the reversibility of annexin A2t binding to solid supported membranes: a QCM study. Biochemistry. 2003;42:3131–3141. doi: 10.1021/bi027069z. [DOI] [PubMed] [Google Scholar]
- Saito H, Hamilton SM, Tavill AS, Louis L, Ratnoff OD. Production and release of plasminogen by isolated perfused rat liver. Proc Natl Acad Sci USA. 1980;77:6837–6840. doi: 10.1073/pnas.77.11.6837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakai M, Watanuki M, Matsuo O. Mechanism of fibrin-specific fibrinolysis by staphylokinase: participation of alpha 2-plasmin inhibitor. Biochem Biophys Res Commun. 1989;162:830–837. doi: 10.1016/0006-291x(89)92385-1. [DOI] [PubMed] [Google Scholar]
- Sanderson-Smith ML, De Oliveira DM, Ranson M, McArthur JD. Bacterial plasminogen receptors: mediators of a multifaceted relationship. J Biomed Biotechnol. 2012;2012:272148. doi: 10.1155/2012/272148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sappino A-P, huarte J, Belin D, Vassalli J-D. Plasminogen activators in tissue remodeling and invasion: mRNA localization in mouse ovaries and implanting embryos. J Cell Biol. 1989;109:2471–2479. doi: 10.1083/jcb.109.5.2471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sappino A-P, Huarter J, Vassalli J-D, Belin D. Sites of synthesis of urokinase and tissue-type plasminogen activators in the murine kidney. J Clin Invest. 1991;87:962–970. doi: 10.1172/JCI115104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sappino A-P, Madani R, huarte J, Belin D, Kiss JZ, Wohlwend A, Vassalli J-D. Extracellular proteolysis in the adult murine brain. J Clin Invest. 1993;92:679–685. doi: 10.1172/JCI116637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schousboe I, Feddersen K, Rojkjaer R. Factor XIIa is a kinetically favorable plasminogen activator. Thromb Haemost. 1999;82:1041–1046. [PubMed] [Google Scholar]
- Schuster V, Hugle B, Tefs K. Plasminogen deficiency. J Thromb Haemost. 2007;5:2315–2322. doi: 10.1111/j.1538-7836.2007.02776.x. [DOI] [PubMed] [Google Scholar]
- Schuster V, Seregard S. Ligneous conjunctivitis. Surv Ophthalmol. 2003;48:369–388. doi: 10.1016/s0039-6257(03)00056-0. [DOI] [PubMed] [Google Scholar]
- Sehl LC, Castellino FJ. Thermodynamic properties of the binding of alpha-, omega-amino acids to the isolated kringle 4 region of human plasminogen as determined by high sensitivity titration calorimetry. J Biol Chem. 1990;265:5482–5486. [PubMed] [Google Scholar]
- Silverstein RL, Friedlander RJ, Jr, Nicholas RL, Nachman RL. Binding of lys-plasminogen to monocytes/macrophages. J Clin Invest. 1988;82:1948–1955. doi: 10.1172/JCI113814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinniger V, Merton RE, Fabregas P, Felez J, Longstaff C. Regulation of tissue plasminogen activator activity by cells. J Biol Chem. 1999;274:12414–12422. doi: 10.1074/jbc.274.18.12414. [DOI] [PubMed] [Google Scholar]
- Sottrup-Jensen L, Claeys H, Zajdel M, Petersen TE, Magnusson S. The primary structure of human plasminogen: Isolation of two lysine-binding fragments and one “mini-”plasminogen (MW 38,000) by elastase-catalyzed-specific limited proteolysis. In: Davidson JF, Rowan RM, Samama MM, Desnoyers PC, editors. Progress in Chemical Fibrinolysis and Thrombolysis. Vol. 3. Raven Press; New York: 1978. pp. 191–209. [Google Scholar]
- Stricker RB, Wong D, Tak Shiu D, Reyes PT, Shuman MA. Activation of plasminogen by tissue plasminoagen activator on normal and thrombasthenic platelets: Effects on surface proteins and platelet aggregation. Blood. 1986;68:275–280. [PubMed] [Google Scholar]
- Strickland S, Reich E, Sherman MI. Plasminogen activator in early embryogenesis: enzyme production by trophoblast and parietal endoderm. Cell. 1976;9:231–240. doi: 10.1016/0092-8674(76)90114-8. [DOI] [PubMed] [Google Scholar]
- Suelves M, Lopez-Alemany R, Lluis F, Aniorte G, Serrano E, Parra M, Carmeliet P, Munoz-Canoves P. Plasmin activity is required for myogenesis in vitro and skeletal muscle regeneration in vivo. Blood. 2002;99:2835–2844. doi: 10.1182/blood.v99.8.2835. [DOI] [PubMed] [Google Scholar]
- Sulniute R, Lindh T, Wilczynska M, Li J, Ny T. Plasmin is essential in preventing periodontitis in mice. Am J Pathol. 2011;179:819–828. doi: 10.1016/j.ajpath.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surette AP, Madureira PA, Phipps KD, Miller VA, Svenningsson P, Waisman DM. Regulation of fibrinolysis by S100A10 in vivo. Blood. 2011;118:3172–3181. doi: 10.1182/blood-2011-05-353482. [DOI] [PubMed] [Google Scholar]
- Swaisgood CM, Schmitt D, Eaton D, Plow EF. In vivo regulation of plasminogen function by plasma carboxypeptidase B. J Clin Invest. 2002;110:1275–1282. doi: 10.1172/JCI15082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Syrovets T, Jendrach M, Rohwedder A, Schule A, Simmet T. Plasmin-induced expression of cytokines and tissue factor in human monocytes involves AP-1 and IKKbeta-mediated NF-kappaB activation. Blood. 2001;97:3941–3950. doi: 10.1182/blood.v97.12.3941. [DOI] [PubMed] [Google Scholar]
- Syrovets T, Lunov O, Simmet T. Plasmin as a proinflammatory cell activator. J Leukoc Biol. 2012;92:509–519. doi: 10.1189/jlb.0212056. [DOI] [PubMed] [Google Scholar]
- Syrovets T, Tippler B, Rieks M, Simmet T. Plasmin is a potent and specific chemoattractant for human peripheral monocytes acting via a cyclic guanosine monophosphate-dependent pathway. Blood. 1997;89:4574–4583. [PubMed] [Google Scholar]
- Szaba FM, Smiley ST. Roles for thrombin and fibrin(ogen) in cytokine/chemokine production and macrophage adhesion in vivo. Blood. 2002;99:1053–1059. doi: 10.1182/blood.v99.3.1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorsen S. The mechanism of plasminogen activation and the variability of the fibrin effector during tissue-type plasminogen activator-mediated fibrinolysis. Ann N Y Acad Sci. 1992;667:52–63. doi: 10.1111/j.1749-6632.1992.tb51597.x. [DOI] [PubMed] [Google Scholar]
- Tillett WS, Garner RL. THE FIBRINOLYTIC ACTIVITY OF HEMOLYTIC STREPTOCOCCI. J Exp Med. 1933;58:485–502. doi: 10.1084/jem.58.4.485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd AS. The histological localisation of fibrinolysin activator. J Pathol Bact. 1959;78:281–283. doi: 10.1002/path.1700780131. [DOI] [PubMed] [Google Scholar]
- Tordai H, Banyai L, Patthy L. The PAN module: the N-terminal domains of plasminogen and hepatocyte growth factor are homologous with the apple domains of the prekallikrein family and with a novel domain found in numerous nematode proteins. FEBS Lett. 1999;461:63–67. doi: 10.1016/s0014-5793(99)01416-7. [DOI] [PubMed] [Google Scholar]
- Tsirka SE, Rogove AD, Bugge TH, Degen JL, Strickland S. An extracellular proteolytic cascade promotes neuronal degeneration in the mouse hippocampus. J Neurosci. 1997;17:543–552. doi: 10.1523/JNEUROSCI.17-02-00543.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Twining SS, Wilson PM, Ngamkitidechakul C. Extrahepatic synthesis of plasminogen in the human cornea is up- regulated by interleukins-1alpha and -1beta. Biochem J. 1999;339 (Pt 3):705–712. [PMC free article] [PubMed] [Google Scholar]
- Unkeless JC, Gordon S, Reich E. Secretion of plasminogen activator by stimulated macrophages. J Exp Med. 1974;139:834. doi: 10.1084/jem.139.4.834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urano T, Chibber BAK, Castellino FJ. The reciprocal effects of E-aminohexanoic acid and chloride ion on the activation of human [Glu1]plasminogen by human urokinase. Proc Natl Acad Sci USA. 1987a;84:4031–4034. doi: 10.1073/pnas.84.12.4031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urano T, de Serrano VS, Chibber BAK, Castellino FJ. The control of the urokinase-catalyzed activation of human glutamic acid 1-plasminogen by positive and negative effectors. J Biol Chem. 1987b;262:15959–15964. [PubMed] [Google Scholar]
- Urano T, de Serrano VS, Gaffney PJ, Castellino FJ. Effectors of the activation of human glu-plasminogen by human tissue plasminogen activator. Biochemistry. 1988;27:6522–6528. doi: 10.1021/bi00417a049. [DOI] [PubMed] [Google Scholar]
- Vassalli J, Dayer J, Wohlwend A, Belin D. Concomitant secretion of prourokinase and of a plasminogen activator specific inhibitor by cultured human monocytes-macrophages. J Exp Med. 1984;159:1653. doi: 10.1084/jem.159.6.1653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Violand BN, Byrne R, Castellino FJ. The effect of alpha-, omega-amino acids on human plasminogen structure and activation. J Biol Chem. 1978;253:5395–5401. [PubMed] [Google Scholar]
- Violand BN, Castellino FJ. Mechanism of the urokinase-catalyzed activation of human plasminogen. J Biol Chem. 1976;251:3906–3912. [PubMed] [Google Scholar]
- Violand BN, Sodetz JM, Castellino FJ. The effect of epsilon-amino caproic acid on the gross conformation of plasminogen and plasmin. Arch Biochem Biophys. 1975;170:300–305. doi: 10.1016/0003-9861(75)90121-6. [DOI] [PubMed] [Google Scholar]
- Wang GG, Calvo KR, Pasillas MP, Sykes DB, Hacker H, Kamps MP. Quantitative production of macrophages or neutrophils ex vivo using conditional Hoxb8. Nat Methods. 2006;3:287–293. doi: 10.1038/nmeth865. [DOI] [PubMed] [Google Scholar]
- Wang H, Prorok M, Bretthauer RK, Castellino FJ. Serine-578 is a major phosphorylation locus in human plasma plasminogen. Biochemistry. 1997a;36:8100–8106. doi: 10.1021/bi970328d. [DOI] [PubMed] [Google Scholar]
- Wang H, Prorok M, Bretthauer RK, Castellino FJ. Serine-578 is a major phosphorylation locus in human plasma plasminogen. Biochemistry. 1997b;36:8100–8106. doi: 10.1021/bi970328d. [DOI] [PubMed] [Google Scholar]
- Weide I, Römisch J, Simmet T. Contact activation triggers stimulation of the monocyte 5-lipoxygenase pathway via plasmin. Blood. 1994;83:1941–1951. [PubMed] [Google Scholar]
- Weide I, Tippler B, Syrovets T, Simmet T. Plasmin is a specific stimulus of the 5-lipoxygenase pathway of human peripheral monocytes. Thromb Haemost. 1996;76:561–568. [PubMed] [Google Scholar]
- Wilson EL, Becker ML, Hoal EG, Dowdle EB. Molecular species of plasminogen activators secreted by normal and neoplastic human cells. Cancer Res. 1980;40:933–938. [PubMed] [Google Scholar]
- Wiman B. Primary structure of peptides released during activation of human plasminogen by urokinase. Eur J Biochem. 1973;39:1–9. doi: 10.1111/j.1432-1033.1973.tb03096.x. [DOI] [PubMed] [Google Scholar]
- Wiman B, Wallen P. Activation of human plasminogen by an insoluble derivative or urokinase. Structural changes of plasminogen in the course of activation to plamsin and demonstration of a possible intermediate complex. Eur J Biochem. 1973;36:25–31. doi: 10.1111/j.1432-1033.1973.tb02880.x. [DOI] [PubMed] [Google Scholar]
- Wygrecka M, Marsh LM, Morty RE, Henneke I, Guenther A, Lohmeyer J, Markart P, Preissner KT. Enolase-1 promotes plasminogen-mediated recruitment of monocytes to the acutely inflamed lung. Blood. 2009;113:5588–5598. doi: 10.1182/blood-2008-08-170837. [DOI] [PubMed] [Google Scholar]
- Xue Y, Bodin C, Olsson K. Crystal structure of the native plasminogen reveals an activation-resistant compact conformation. J Thromb Haemost. 2012;10:1385–1396. doi: 10.1111/j.1538-7836.2012.04765.x. [DOI] [PubMed] [Google Scholar]
- Zhang L, Gong Y, Grella DK, Castellino FJ, Miles LA. Endogenous plasmin converts Glu-plasminogen to Lys-plasminogen on the monocytoid cell surface. J Thromb Haemost. 2003;1:1264–1270. doi: 10.1046/j.1538-7836.2003.00155.x. [DOI] [PubMed] [Google Scholar]
- Zhang L, Seiffert D, Fowler B, Jenkins GR, Thinnes T, Loskutoff DJ, Parmer RJ, Miles LA. Plasminogen has a broad extrahepatic distribution. Thromb Haemost. 2002a;87:493–501. [PubMed] [Google Scholar]
- Zhang L, Seiffert D, Fowler BJ, Jenkins GR, Thinnes TC, Loskutoff DJ, Parmer RJ, Miles LA. Plasminogen has a broad extrahepatic distribution. Thromb Haemost. 2002b;87:493–501. [PubMed] [Google Scholar]
- Zhang Y, Zhou ZH, Bugge TH, Wahl LM. Urokinase-type plasminogen activator stimulation of monocyte matrix metalloproteinase-1 production is mediated by plasmin-dependent signaling through annexin A2 and inhibited by inactive plasmin. J Immunol. 2007;179:3297–3304. doi: 10.4049/jimmunol.179.5.3297. [DOI] [PubMed] [Google Scholar]
- Zwaal RF, Comfurius P, Bevers EM. Surface exposure of phosphatidylserine in pathological cells. Cell Mol Life Sci. 2005;62:971–988. doi: 10.1007/s00018-005-4527-3. [DOI] [PMC free article] [PubMed] [Google Scholar]