

Abstract
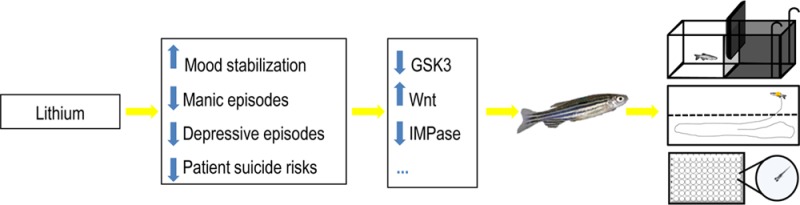
Lithium is the paradigmatic treatment for bipolar disorder and has been widely used as a mood stabilizer due to its ability to reduce manic and depressive episodes, efficiency in long-term mood stabilization, and effectiveness in reducing suicide risks. Despite many decades of clinical use, the molecular targets of lithium are not completely understood. However, they are credited at least partially to glycogen synthase kinase 3 (GSK3) inhibition, mimicking and exacerbating Wnt signaling pathway activation. There has been a great effort to characterize lithium cellular and system actions, aiming to improve treatment effectiveness and reduce side effects. There is also a growing concern about lithium’s impact as an environmental contaminant and its effects on development. In this scenario, zebrafish is a helpful model organism to gather more information on lithium’s effects in different systems and developmental stages. The rapid external development, initial transparency, capacity to easily absorb substances, and little space required for maintenance and experimentation, among other advantages, make zebrafish a suitable model. In addition, zebrafish has been established as an effective model organism in behavioral and neuropharmacological studies, reacting to a wide range of psychoactive drugs, including lithium. So far only a limited number of studies evaluated the toxicological impact of lithium on zebrafish development and demonstrated morphological, physiological, and behavioral effects that may be informative regarding human findings. Further studies dedicated to characterize and evaluate the underlying mechanisms of the toxic effects and the potential impact of exposure on developing and adult individuals are necessary to establish safe clinical management guidelines for women with bipolar disorder of childbearing age and safety disposal guidelines for pharmaceutical neuroactive compounds.
Keywords: Lithium, zebrafish, toxicity, bipolar disorder, GSK3, Wnt
1. Introduction
Lithium remains, after more than 60 years from its therapeutic discovery, the paradigmatic treatment for bipolar disorder and has been widely used as a mood stabilizer due to its ability to reduce manic and depressive episodes, efficiency in long-term mood stabilization, and effectiveness in reducing patient suicide risks.1 Despite the effectiveness of lithium on bipolar disorders, its underlying mechanisms of action are not completely characterized but are mostly attributed to inhibition of glycogen synthase kinase 3 (GSK3) and inositol monophosphatase (IMPase).2,3 Due to GSK3 inhibition, lithium mimics and exacerbates Wnt signaling pathway activation, which is critical for central nervous system development and plasticity.4 There has been a great effort to characterize lithium cellular and system actions, aiming to improve treatment effectiveness and reduce side effects. In order to fully assess the therapeutic and toxicological effects of lithium, different experimental models have been proposed.
The teleost zebrafish Danio rerio has already been established as an effective model organism in several areas, from developmental biology and genetics to behavior and neuropharmacology. Both larval and adult zebrafish have been used to investigate brain function, behavior disorders and genetic and pharmacological manipulation.5,6 Zebrafish presents significant physiological similarities to humans, in addition to a central nervous system (CNS) with the main homologous areas present.6 Furthermore, a direct comparison of protein-coding genes in humans and zebrafish recently revealed that approximately 84% of human disease-related genes have at least one ortholog in zebrafish.7 Zebrafish has sensorial, motor, and endocrine systems that are well-developed, high sensitivity to environmental changes and pharmacological manipulation, evident cognitive abilities, and a sophisticated behavioral repertoire.6,8,9 Several studies have shown the behavioral effects of psychotropic compounds, including anxiolytics and antidepressives in zebrafish.10−12 Additionally, zebrafish has the capacity to easily absorb substances and present rapid external development, allowing easy manipulation, final advantages that make this animal organism highly suitable for genetic and chemical screens.13
2. Zebrafish
2.1. Zebrafish Development
Zebrafish development is fast, comprising an embryological period restricted to only 3 days after which a well formed larva eclodes and achieves adulthood within a few months.14,15 Notably, zebrafish is particularly well-suited for development studies because embryos develop externally and remain transparent throughout their initial development stages. These conditions allow embryo manipulation and morphological analysis during development in vivo.15 At early larval stages, on 5 days postfertilization (dpf), zebrafish already displays sophisticated behavioral manifestations and complete neurotransmitter systems.15
The developmental life stages of zebrafish are classified, in progressive order, as embryonic (from fertilization until the hatching), larval (after hatching, yet without adult characteristics), juvenile (presents adult characteristics but not sexual maturity) and adult (produces gametes, presents secondary sexual characteristics and is able to breed).14,15 The zebrafish embryogenesis is well described and defined in seven stages: zygote, cleavage, blastula, gastrula, segmentation, pharyngula, hatching, and early larva (Figure 1). These stages occur within the first 72 h postfertilization (hpf).14,15 At the end of embryogenesis, the animal is a larva with almost complete morphogenesis, moving eyes and fins, inflated swim bladder, and well-formed jaw.14 The larval stage is marked by food seeking, avoidance behavior, swimming activity, and absence of social behavior.14,15 The zebrafish larval stage lasts up to 1 month until the animal becomes a juvenile.14,15 The process of larval metamorphosis is characterized by the loss of fin fold, nervous system further development and acquisition of scales.15 The juvenile stage is evidenced by the start of zebrafish shoaling behavior.16 Finally, zebrafish is termed “adult” when it becomes sexually mature, which occurs at 3–4 months. Adult zebrafish present shoaling behavior, exploratory activity and a sophisticated behavioral repertory.6,17,18
Figure 1.

Stages of zebrafish early development. From single-cell fertilized zygote to a hatched larva takes only 72 hpf.
2.2. Zebrafish Behavior
Both larval and adult zebrafish have been regularly used as models in studies of depression, bipolar and autism disorder, among others CNS conditions.6,19,20 These studies are based on the well characterized behavior of zebrafish, which is marked by group member interdependence and social interaction, thigmotaxis, scototaxis and exploratory activity.6,16,20,21 Accordingly, the use of zebrafish for studying behavioral disorders and drugs effects has received a growing experimental support. Different behavior test protocols were established and adjusted to zebrafish, including social interaction, novel environment/tank and inhibitory avoidance task, among others.9,22−24
Zebrafish is a social species, living in shoals with ordered social relationships, including dominance hierarchies and territoriality.25,26 Shoaling behavior reduces predation and enhances foraging efficiency, and, accordingly, this species shows increased cohesion in the presence of a predator and decreased cohesion during feeding.27 Shoaling is an innate behavior that appears early in development, and shoal cohesion significantly increases from the first days of free swimming stage to adulthood.25,26,28 Gerlai et al. showed animals’ preferences for a group of conspecifics regardless of being in isolation or in groups of up to five individuals.23 In a zebrafish model of schizophrenia, the NMDA receptor antagonist MK-801 significantly decreased the time of social interaction while the atypical antipsychotic drugs sulpiride and olanzapine reversed the MK-801- induced social interaction deficits.29
Zebrafish demonstrates robust response to novelty, which is marked by constant swimming activity and exploratory behavior. When placed in a novel environment, this teleost shows initial increased exploratory activity, which significantly decreases throughout time while animals habituate.20 This process is sensitive to environment manipulation, drugs effects and pathological conditions. Piato et al. (2011) showed that unpredictable chronic stress reduces the exploratory activity, which may reflect a resilience loss and depressive-like behavior induced by chronic stress.30 Treatment with anxiolytic drugs such as benzodiazepines restores the increased exploratory activity.9
This teleost presents evident defensive/antipredatory behaviors, having response patterns of scototaxis, thigmotaxis and diving well characterized.21,31 Thigmotaxis, also known as “wall-following”, is the tendency to stay close to the borders in a novel environment aiming to facilitate the search of shelter and escape routes.32 The thigmotaxis tests have been used to investigate mechanisms of fear and anxiety and also to screen psychotropic drugs.31 In addition to thigmotaxis, zebrafish demonstrates a marked scototaxis behavior, which is the preference for dark environments.21 As thigmotaxis, scototaxis can be used to assess anxiety and fear in zebrafish.21 The increased time spent in a dark environment may reflect anxiety.21 Piato et al. showed that stressed animals spend more time in a dark chamber comparing with nonstressed controls.30 Like thigmotaxis and scototaxis, bottom diving response is a defensive/antipredatory behavior that is evident in a new environment. During novelty habituation, zebrafish tends to initially dive to the bottom and gradually swim to upper areas of the tank (reviewed by Blaser and Rosemberg).33 This behavior pattern can be modified in behavioral disorders and is sensitive to pharmacological manipulation. Anxiolytic and antidepressive drugs significantly reduce the time spent in the bottom of a novel tank,34 and animals treated with fluoxetine and tranylcypromine remain longer periods at the top of the tank and show decreased locomotor activity, characteristic effects of increased serotonergic signaling.35
Zebrafish has evident cognitive abilities and several studies have shown that zebrafish present excellent short-term and long-term memory and respond to different learning tasks.6 Aiming to evaluate rapidly acquired and consistent paradigms of learning and memory in zebrafish, Blank et al. proposed an inhibitory avoidance task in which animals learn to refrain from swimming from a white into a dark chamber in order to avoid an electric shock in a single-trial training. The resulting memory is robust, long-lasting and sensitive to NMDA-receptor antagonist MK-801.22 In an animal model of schizophrenia induced by the NMDA antagonist MK-801, Seibt et al. showed that MK-801 treatment induced memory deficits in zebrafish and that this effect was completely reversed by acute administration of atypical antipsychotics olanzapine and sulpiride.29
2.3. Drug Screening in Zebrafish
The rapid development, capacity to absorb easily substances, and little space required for their maintenance and experimentation, among other advantages, make zebrafish a great animal model for drug screening.6,36,37 Zebrafish have been widely used to study in vivo effects of several drugs and identification of their targets. Different drugs can be added directly to the water and taken up by both embryo, larvae and adult zebrafish through gills and skin. Furthermore, the embryos and larvae can be individually treated and simultaneously analyzed into 96-well plates, facilitating large scale drug screening (Figure 2).38,39
Figure 2.
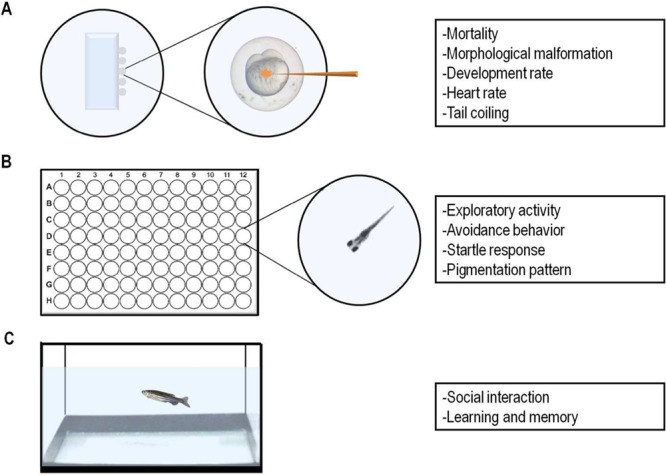
Parameters progressively studied in drug screening assays at different developmental stages in zebrafish: (A) Embryonic stages exposure or injection. (B) Larvae treatment in 96-well cell culture dishes. (C) Adult zebrafish treated in tank water.
When zebrafish embryos and larvae are used to study the developmental toxicity of chemical substances, different parameters can be analyzed, including: survival/mortality, morphological malformations, and functional deficits or potential target organs and systems.40 Studies may also evaluate the behavioral ability of larvae, commonly based on parameters associated with swimming such as distance traveled and mean speed. Recently, more complex behavioral protocols have been validated for this life stage and include light induced changes,41 thigmotaxis,42,43 and avoidance.44
In order to understand the therapeutic mechanisms and toxicological effects of drugs of clinical use such as lithium, it is strategically relevant to use experimental models that allow the characterization of behavioral and toxicological parameters. In the last 10 years, zebrafish has been considered a valuable model for studies evaluating anxiety, stress, depression, and psychosis-like behaviors.30,45−48
Zebrafish reacts to a wide range of psychoactive drugs, such as antidepressants and antipsychotics (reviewed by Stewart et al.).49 Among antidepressant drugs, the selective serotonin reuptake inhibitor (SSRI) fluoxetine induces decreased swimming activity in specific stages (6–14 dpf) of developing zebrafish.50 Egan et al. have demonstrated that chronic exposure to fluoxetine promotes an anxiolytic behavior paralleled with reduced whole-body cortisol concentrations when compared to controls.8 Studies have shown that while acute fluoxetine alters locomotion in higher doses, chronic treatment for 2 weeks induces anxiolytic effects with no changes on locomotor activity.8,51 Accordingly, behavioral alterations and transcriptome changes have been observed after racemic and stereoisomeric fluoxetine treatment in zebrafish.52 When treated with other SSRI drugs such as citalopram, which binds with high affinity to serotonin transporters in zebrafish brain (Ki = 9 ± 5 nM) fish spent more time in tank top zone.53 Desipramine, a tricyclic antidepressant, also induced a anxiolytic effects and high affinity for serotonin transporters (Ki = 7 ± 5 nM). Amitriptyline, another tricyclic antidepressant drug, produces marked toxicological effects, reducing the hatching time and body length of embryos accompanied by changes in adrenocorticotropic hormone (ACTH) level, oxidative stress and antioxidant parameters, as well as nitric oxide (NO) production and total nitric oxide synthase (TNOS) activity.54
Zebrafish has also demonstrated behavioral responses to several anxiolytic drugs (reviewed in Stewart et al.).49 For example, it has been shown that buspirone increased zebrafish exploration of higher portions of the tank and the time spent in a bright-lighted compartment.11 Antipsychotics such as olanzapine and sulpiride also increase the time spent in the top zone of the tank, which is commonly interpreted as an anxiolytic effect.47
3. Effects of Lithium in Zebrafish
Despite the increasing interest about the effects of antidepressants and anxiolytics on behavioral responses in zebrafish, few studies have been performed evaluating the actions of lithium on this experimental model. Studies have demonstrated that drugs used for treatment of mood disorders, such as lithium, are able to modulate circadian rhythm.55 Lithium chloride (LiCl) promoted a 0.7 h lengthening of the circadian period in a specific zebrafish line, Tg(4xEbox:Luc), able to detect maturing and developmental activity of the circadian clock.56 LiCl and LiCl plus forskolin treatments promoted a significant increase in the pigmentation in zebrafish embryos, due to synthesis of melanin in neural crest-derived melanophores.57
Despite the need of further studies characterizing lithium targets, its effects were investigated in neurotransmitter systems involved in depression and anxiety. Among them, we may highlight the purinergic and cholinergic systems. Previous studies have demonstrated that the exposure to 5 and 10 mg/L lithium chloride during 7 days reduced ADP hydrolysis promoted by nucleoside triphosphate diphosphohydrolases (NTPDases), whereas ecto-5′-nucleotidase and acetylcholinesterase activities were only decreased at 10 mg/L lithium chloride.58 These inhibitory effects may be related to an indirect effect of lithium on NTPDase, 5′-nucleotidase, and acetylcholinesterase, since lithium did not significantly affect these enzymes activities in vitro (when added directly to reaction medium).58 The control of ATP, adenosine, and acetylcholine levels may represent a new mechanism underlying the neuroprotective and therapeutic effects of lithium.
4. Toxicological Effects of Lithium
The potential toxicological effects of lithium due to environmental exposure and therapeutic overload have already been studied (reviewed by Aral and Vecchio-Sadus).59 Aral and Vecchio-Sadus suggested that lithium is not expected to bioaccumulate, and that its human and environmental toxicities are low. Lithium intake neither from food and water nor from occupational exposure presents a toxicological hazard. However, additional research is required to determine the amount of lithium that is beneficial to health and to obtain a better understanding of the mechanisms by which lithium alters behavior and its potential side effects.59
McKnight et al. delineated a toxicity profile of lithium through a systematic review and meta-analysis of randomized controlled trials and observational studies. The study suggested that lithium is associated with hypothyroidism, hyperparathyroidism, reduced urinary concentrating ability, and weight gain. Also, the study showed that there is little evidence for a clinically significant reduction in renal activity and that the risk of end-stage renal failure is low. In addition, the risk of congenital malformations was unclear.60
Despite lithium classification as a low teratogenic agent,61,62 concerns about its potential toxic effects on developing individuals are accentuated by the lack of studies supporting definitive management guidelines during pregnancy.63 Bipolar patient gestations are commonly accompanied by a conflict between therapy interruption induced-relapse risk and the potential deleterious effects of continued therapy on development. Reports of teratogenic effects of lithium exposure during early human intrauterine development and breastfeeding are controversial and include reports of cardiac, renal, and hepatic abnormalities, infant diabetes insipidus, and hypothyroidism.64−70 Lithium exposure during development has also been associated with neurobehavioral effects including depressed neuromuscular function manifested as lethargy, hypotonicity, slower reflexes, and respiratory difficulties in neonates,71 symptoms collectively described as Floppy Baby Syndrome. The relative incidences of these effects are not clear and need to be systematically investigated as much as its underlying mechanisms.70,72 In addition to observational studies in humans, a key strategy to obtain lacking information is the experimental use of animal models. Previous studies with this objective including rodents and rabbits showed higher incidences of lithium-induced teratogenesis than those observed in humans, what was later attributed to excessively high doses used and species specific sensitivity (discussed by Cohen and collaborators).61
Reports of the lithium exposure effects during zebrafish development vary significantly in terms of treatment duration, period, doses, and conditions, and have revealed this drug impact on development. Lithium has long been known to affect one of the most critical and earliest events on vertebrate embryo development, the formation of the dorsal-ventral body axis. Acute exposure of zebrafish embryos has a dorsalizing effect, severely impacting body-plan formation and impeding normal development. This effect was initially described for exposure during the cleavage stage,73 and recently also shown to occur in the initial first minutes after fertilization (mpf).74 Despite the fact that different cellular and molecular effectors may mediate this phenotype at the different time periods, the negative impact on development is massive and the evolutionary conservation of basic vertebrate development mechanisms alerts to potential deleterious effects in other species including humans.
Due to zebrafish advantages to toxicological-screenings during development, toxic effects of lithium during the organogenesis period have also been reported and include several systems and target organs. Selderslaghs and colleagues used lithium chloride among other substances for the establishment of a new screening procedure for embryotoxic and teratogenic effects after a 0–144 hpf exposure and argued that, in addition to the classic advantages of zebrafish for such screenings, the longer survival of malformed individuals allows an extended analysis. Using concentrations of lithium chloride between 0.05 and 235.9 mM they found no obvious teratogenic effects on the initial 48 hpf, except for occasional kinks in the chorda. According to their results, lithium deleterious effects, including delayed development, skeletal deformities and reduced swimming were observed only at higher doses and at later time points, suggesting a teratogenic potential at later intrachorionic developmental periods. In accordance to what is commonly observed in rat and mice, skeletal deformities were the most frequently morphological effect observed.75
A complementary study by Pruvot et al. exposed animals during the third developmental day confirming the higher sensitivity to lithium at later developmental stages. When embryos were exposed to lithium starting at 48 hpf and lasting 24 h to concentrations ranging from 0 to 250 μM a significant toxic effect was observed.76 While survival was minimally affected, important morphological, physiological, and behavioral defects such as dorsal curvature, pericardial edema, decreased heart rate, decreased swimming activity, and velocity were evident in lithium treated animals.
Recent work by Nery et al.77 demonstrates specific and sustained neurobehavioral effects of lithium exposure during the initial 72 hpf that were not accompanied by teratogenic effects. In this study, embryos were exposed throughout the initial 3 dpf to LiCl concentrations ranging from potentially neuroprotective,78,79 therapeutical, and representative of intoxication during human therapy.80 While survival was unaffected, at the end of treatment period, Western blot analysis of selected Wnt-β-catenin system components showed increased β-catenin and decreased N-cadherin protein levels in animals treated with intermediate and higher lithium doses, supporting GSK-3 inhibition as a central target of lithium’s effects. Interestingly, at 10 dpf, previously altered proteins returned to control levels in lithium treated animals, but sustained behavioral effects were observed including lethargy, new environment exploratory deficits, and thigmotatic behavior.
Additional to developing infants, aquatic animals may have their development affected by lithium environmental contamination associated with the growing consumption of neuroactive drugs and their subsequent presence in wastewater.62 In this scenario, zebrafish is a helpful model organism to gather more information on lithium’s effects on vertebrate development and its underlying cellular and molecular mechanisms, critical information to establish safe clinical management guidelines for woman with bipolar disorder of childbearing age and safety disposal guidelines for pharmaceutical neuroactive compounds.
5. Wnt Signaling Pathway Is a Target of Lithium
Lithium has profound effects on several organisms, affecting development, metabolism, and several physiological parameters. The molecular targets of lithium are not clearly understood and may vary depending on organism and system affected, in addition to developmental stage and genetic background. Several molecules have been shown to have their activities modulated by lithium, including enzymes, apoptotic factors and receptors. (reviewed by Shaldubina et al.).81 The different doses to which organisms and systems are exposed may also encompass specific effectors and explain opposing properties such as neuroprotection and neurotoxicity.82
Despite many decades of clinical use, the molecular targets underlying lithium effects on bipolar patients are only now being outlined.83 Among the enzymes proposed as lithium targets, the IMPase family of structurally related phosphomonoesterases now have its contribution under dispute. IMPase inhibition promotes inositol depletion, a decrease in the resynthesis of phosphatidylinositol bisphosphate (PIP2), which results in a reduction of the second messenger inositol-1,4,5, triphosphate (IP3).3 Altered phosphatydilinositol cycle activity and enhanced myo-inositol levels have been observed in patients with bipolar disorders and could account, at least partially, to lithium effects in bipolar patients.3,84 Despite in vivo data obtained mostly in rodents, no causal relation of its inhibition and therapeutic effects were effectively demonstrated in humans (reviewed by Gould and Manji).85
In 1996 lithium was shown to inhibit the ubiquitous GSK3.86,87 Subsequent demonstrations associated lithium therapeutic doses to GSK3 inhibition and mood stabilization, suggesting this enzyme may also participate in bipolar patients mood stabilization. Most evidence concerning lithium effects on GSK3, including brain plasticity and metabolism, modulation of monoaminergic signaling88,89 stem cell activity and neuroprotection,82,90 however, were obtained in model organisms and cell cultures. Polter et al. observed changes on GSK3 activity in blood cells from patients with bipolar disorder under lithium regimen at therapeutically effective doses.91 Moreover, other GSK3 inhibitors promote behavioral effects similar to those induced by lithium in rodents92 and mice with reduced GSK3 activity due to genetic modification demonstrated a behavioral profile that mimics the effects observed after chronic lithium treatment.93
GSK3 affects uncountable cellular events in different tissues due to its central regulatory actions on the canonical Wnt-beta-catenin signaling pathway (Figure 3). The Wnt family of secreted glycoproteins modulates several cellular events through the regulation of catenin-mediated gene expression of DNA-bound T cell factor/lymphoid enhancement factor (TCF/LEF) family of transcriptional factors. This pathway activation begins with Wnt binding to frizzled (Fz or FZd) receptor and its low-density lipoprotein receptor-related protein coreceptor (LPR) which leads to a series of events that alters the activation state of a multiprotein system referred as “β-catenin destruction-complex”. This in turn prevents GSK3 phosphorylation of β-catenin residues Thr41, Ser37, and Ser33 and subsequent ubiquitination and proteosomal degradation.94,95 β-catenin stabilization due to Wnt signaling or lithium-mediated GSK3 inhibition results in higher nuclear levels and differential modulation of TCF/LEF-mediated gene expression. β-catenin may also translocate to the membrane and associate to members of the cadherin family of adhesion molecules.
Figure 3.

Lithium mimics and exacerbates Wnt/β-catenin signaling pathway activation. (A) Schematic representation of the canonical Wnt/β-catenin signaling pathway off state when β-catenin is ubiquitinated and degraded by proteosome. (B) Lithium mimics Wnt/β-catenin signaling by GSK3 inhibition and allows β-catenin nuclear modulation of transcription.
Wnt plays critical roles on development, including actions as morphogens and controlling stem cell proliferation, cell fate determination and differentiation, migration, growth cone guidance, synaptogenesis, and sustained synaptic plasticity through adulthood.96,97 Lithium inhibition of GSK3 mimics and exacerbates Wnt-β-catenin signaling89,95 and has been used as a tool to study the system contribution to several biological processes in vitro and in vivo.
In addition to the already known events mediated by the Wnt-catenin signaling pathway potentially involved in lithium’s impact on zebrafish development, some downstream effectors are not yet characterized and deserve future studies. The dorsalizing effect of lithium on early development is mediated by GSK3 inhibition, but the maternal effectors on the initial lithium sensitive minutes involved in the microtubule remodeling are not yet characterized. In contrast, at the late cleavage stage of 32-cells, the dorsalized phenotype is attributed to β-catenin nuclear accumulation and altered gene expression modulation.74
6. Perspectives
The benefits of lithium to bipolar patients are unequivocal and reflected by its ubiquitous use despite the need for constant monitoring of Li+ plasma levels required to reach effectiveness and avoid toxicity. Detailed information and comprehensive data about its cellular and molecular targets are necessary to further advance the current knowledge about this drug potential impact in other systems. The growing concern about pharmaceuticals as environmental contaminants increasingly found in aquatic systems reinforces the need for more studies dedicated to lithium toxic potential in broader ranges of concentration and developmental stages. Finally, the lack of conclusive data supporting clinical management guidelines for bipolar woman of childbearing age and safety disposal guidelines for pharmaceutical neuroactive compounds support the necessity of further studies. In this context, zebrafish has proven to be advantageous and congregates the possibilities of toxicological, molecular, behavioral, and developmental studies.
Acknowledgments
The authors would like to thank Fabiano Peres and Natália Eltz Silva, Zeblab, for images used in Figure 1.
Glossary
Abbreviations
- dpf
days postfertilization
- Dsh
dishvelled
- Fz or FZd
frezzled receptor
- GSK3
glycogen synthase kinase 3
- hpf
hours postfertilization
- IMPase
inositol monophosphatase
- LPR
low-density lipoprotein receptor-related protein coreceptor
- mpf
minutes postfertilization
- NMDA
N-methyl-d-Aspartate
- TCF/LEF
DNA-bound T cell factor/lymphoid enhancement factor family of transcriptional factors
Author Contributions
The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
Author Contributions
A.M.S. participated in the conception and manuscript design, data collection and review, discussion, writing, critical revision and final approval of the article. M.R.V. participated in the conception and manuscript design, data collection and review, discussion, writing, critical revision and final approval of the article. C.D.B. participated in the conception and manuscript design, data collection and review, discussion, writing, critical revision and final approval of the article.
A.M. Siebel receives a postdoctoral fellowship from CNPq. M.R.V. receives support from CNPq 484124/2013-7 and 308424/2012-3. C.D. Bonan receives a fellowship from CNPq. All authors are supported by DECIT/SCTIE-MS through the CNPq AND FAPERGS (Proc. 10/0036-5, conv. n. 700545/2008-PRONEX).
The authors declare no competing financial interest.
References
- Grof P. (2010) Sixty years of lithium responders. Neuropsychobiology 62, 8–16. [DOI] [PubMed] [Google Scholar]
- De Sarno P.; Li X.; Jope R. S. (2002) Regulation of Akt and glycogen synthase kinase-3 beta phosphorylation by sodium valproate and lithium. Neuropharmacology 43, 1158–1164. [DOI] [PubMed] [Google Scholar]
- Haimovich A.; Eliav U.; Goldbourt A. (2012) Determination of the lithium binding site in inositol monophosphatase, the putative target for lithium therapy, by magic-angle-spinning solid-state NMR. J. Am. Chem. Soc. 134(12), 5647–5651. [DOI] [PubMed] [Google Scholar]
- Meffre D.; Grenier J.; Bernard S.; Courtin F.; Dudev T.; Shackleford G.; Jafarian-Tehrani M.; Massaad C. (2013) Wnt and lithium: a common destiny in the therapy of nervous system pathologies?. Cell. Mol. Life Sci. 71, 1123–1148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Norton W. H. (2013) Toward developmental models of psychiatric disorders in zebrafish. Front. Neural Circuits 7, 79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalueff A. V.; Stewart A. M.; Gerlai R. (2014) Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol. Sci. 35(2), 63–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howe K.; et al. (2013) The zebrafish reference genome sequence and its relationship to the human genome. Nature 496(7446), 498–503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Egan R. J.; Bergner C. L.; Hart P. C.; Cachat J. M.; Canavello P. R.; Elegante M. F.; Elkhayat S. I.; Bartels B. K.; Tien A. K.; Tien D. H.; Mohnot S.; Beeson E.; Glasgow E.; Amri H.; Zukowska Z.; Kalueff A. V. (2009) Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 205(1), 38–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cachat J.; Stewart A.; Grossman L.; Gaikwad S.; Kadri F.; Chung K. M.; Wu N.; Wong K.; Roy S.; Suciu C.; Goodspeed J.; et al. (2010) Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 5(11), 1786–1799. [DOI] [PubMed] [Google Scholar]
- Lau B. Y.; Mathur P.; Gould G. G.; Guo S. (2011) Identification of a brain center whose activity discriminates a choice behavior in zebrafish. Proc. Natl. Acad. Sci. U.S.A. 108(6), 2581–2586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gebauer D. L.; Pagnussat N.; Piato A. L.; Schaefer I. C.; Bonan C. D.; Lara D. R. (2011) Effects of anxiolytics in zebrafish: similarities and differences between benzodiazepines, buspirone and ethanol. Pharmacol., Biochem. Behav. 99(3), 480–486. [DOI] [PubMed] [Google Scholar]
- Boehmler W.; Carr T.; Thisse C.; Thisse B.; Canfield V. A.; Levenson R. (2007) D4 Dopamine receptor genes of zebrafish and effects of the antipsychotic clozapine on larval swimming behaviour. Genes Brain Behav. 6(2), 155–166. [DOI] [PubMed] [Google Scholar]
- Zon L. I.; Peterson R. T. (2005) In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discovery 4(1), 35–44. [DOI] [PubMed] [Google Scholar]
- Kimmel C. B.; Ballard W. W.; Kimmel S. R.; Ullmann B.; Schilling T. F. (1995) Stages of embryonic development of the zebrafish. Dev. Dyn. 203(3), 253–310. [DOI] [PubMed] [Google Scholar]
- Parichy D. M.; Elizondo M. R.; Mills M. G.; Gordon T. N.; Engeszer R. E. (2009) Normal table of postembryonic zebrafish development: staging by externally visible anatomy of the living fish. Dev. Dyn. 238(12), 2975–3015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buske C.; Gerlai R. (2012) Maturation of shoaling behavior is accompanied by changes in the dopaminergic and serotoninergic systems in zebrafish. Dev. Psychobiol. 54(1), 28–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Falk-Petersen I. B. (2005) Comparative organ differentiation during early life stages of marine fish. Fish Shellfish Immunol. 19(5), 397–412. [DOI] [PubMed] [Google Scholar]
- Dahm R.; Geisler R. (2006) Learning from small fry: the zebrafish as a genetic model organism for aquaculture fish species. Mar. Biotechnol. (NY) 8(4), 329–345. [DOI] [PubMed] [Google Scholar]
- Ellis L. D.; Soanes K. H. (2012) A larval zebrafish model of bipolar disorder as a screening platform for neuro-therapeutics. Behav. Brain Res. 233(2), 450–457. [DOI] [PubMed] [Google Scholar]
- Stewart A. M.; Gaikwad S.; Kyzar E.; Kalueff A. V. (2012) Understanding spatio-temporal strategies of adult zebrafish exploration in the open field test. Brain Res. 1451, 44–52. [DOI] [PubMed] [Google Scholar]
- Maximino C.; Marques de; Brito T.; Dias C. A.; Gouveia A.; Morato S. (2010) Scototaxis as anxiety-like behavior in fish. Nat. Protoc. 5(2), 209–216. [DOI] [PubMed] [Google Scholar]
- Blank M.; Guerim L. D.; Cordeiro R. F.; Vianna M. R. (2009) A one-trial inhibitory avoidance task to zebrafish: rapid acquisition of an NMDA-dependent long-term memory. Neurobiol. Learn. Mem. 92(4), 529–534. [DOI] [PubMed] [Google Scholar]
- Gerlai R.; Lahav M.; Guo S.; Rosenthal A. (2000) Drinks like a fish: zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects. Pharmacol., Biochem. Behav. 67(4), 773–782. [DOI] [PubMed] [Google Scholar]
- Rosemberg D. B.; Rico E. P.; Mussulini B. H.; Piato A. L.; Calcagnotto M. E.; Bonan C. D.; et al. (2011) Differences in spatio-temporal behavior of zebrafish in the open tank paradigm after a short-period confinement into dark and bright environments. PLoS One 6(5), e19397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spence R.; Gerlach G.; Lawrence C.; Smith C. (2008) The behaviour and ecology of the zebrafish, Danio rerio. Biol. Rev. Cambridge Philos. Soc. 83(1), 13–34. [DOI] [PubMed] [Google Scholar]
- Spence R.; Magurran A. E.; Smith C. (2011) Spatial cognition in zebrafish: the role of strain and rearing environment. Anim Cognit. 14(4), 607–612. [DOI] [PubMed] [Google Scholar]
- Miller N.; Gerlai R. (2007) Quantification of shoaling behaviour in zebrafish (Danio rerio). Behav. Brain Res. 184(2), 157–166. [DOI] [PubMed] [Google Scholar]
- Buske C.; Gerlai R. (2011) Shoaling develops with age in Zebrafish (Danio rerio). Prog. Neuropsychopharmacol. Biol. Psychiatry 35(6), 1409–1415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seibt K. J.; Piato A. L.; da Luz Oliveira R.; Capiotti K. M.; Vianna M. R.; Bonan C. D. (2011) Antipsychotic drugs reverse MK-801-induced cognitive and social interaction deficits in zebrafish (Danio rerio). Behav. Brain Res. 224(1), 135–139. [DOI] [PubMed] [Google Scholar]
- Piato Â.L.; Capiotti K. M.; Tamborski A. R.; Oses J. P.; Barcellos L. J.; Bogo M. R.; Lara D. R.; Vianna M. R.; Bonan C. D. (2011) Unpredictable chronic stress model in zebrafish (Danio rerio): behavioral and physiological responses. Prog. Neuropsychopharmacol. Biol. Psychiatry. 35(2), 561–567. [DOI] [PubMed] [Google Scholar]
- Champagne D. L.; Hoefnagels C. C.; de Kloet R. E.; Richardson M. K. (2010) Translating rodent behavioral repertoire to zebrafish (Danio rerio): relevance for stress research. Behav. Brain Res. 214(2), 332–342. [DOI] [PubMed] [Google Scholar]
- Sharma S.; Coombs S.; Patton P.; Burt de Perera T. (2009) The function of wall-following behaviors in the Mexican blind cavefish and a sighted relative, the Mexican tetra (Astyanax). J. Comp. Physiol., A 195(3), 225–240. [DOI] [PubMed] [Google Scholar]
- Blaser R. E.; Rosemberg D. B. (2012) Measures of anxiety in zebrafish (Danio rerio): dissociation of black/white preference and novel tank test. PLoS One 7(5), e36931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bencan Z.; Sledge D.; Levin E. D. (2009) Buspirone, chlordiazepoxide and diazepam effects in a zebrafish model of anxiety. Pharmacol., Biochem. Behav. 94(1), 75–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stewart A. M.; Cachat J.; Gaikwad S.; Robinson K. S.; Gebhardt M.; Kalueff A. V. (2013) Perspectives on experimental models of serotonin syndrome in zebrafish. Neurochem. Int. 62(6), 893–902. [DOI] [PubMed] [Google Scholar]
- Berghmans S.; Hunt J.; Roach A.; Goldsmith P. (2007) Zebrafish offer the potential for a primary screen to identify a wide variety of potential anticonvulsants. Epilepsy Res. 75(1), 18–28. [DOI] [PubMed] [Google Scholar]
- Gerlai R. (2010) High-throughput behavioral screens: the first step towards finding genes involved in vertebrate brain function using zebrafish. Molecules 15(4), 2609–2622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rihel J.; Schier A. F. (2011) Behavioral screening for neuroactive drugs in zebrafish. Dev. Neurobiol. 72(3), 373–385. [DOI] [PubMed] [Google Scholar]
- Mirat O.; Sternberg J. R.; Severi K. E.; Wyart C. (2013) ZebraZoom: an automated program for high-throughput behavioral analysis and categorization. Front. Neural Circuits 7, 107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selderslaghs I. W.; Blust R.; Witters H. E. (2012) Feasibility study of the zebrafish assay as an alternative method to screen for developmental toxicity and embryotoxicity using a training set of 27 compounds. Reprod. Toxicol. 33(2), 142–154. [DOI] [PubMed] [Google Scholar]
- Rihel J.; Prober D. A.; Arvanites A.; Lam K.; Zimmerman S.; Jang S.; Haggarty S. J.; Kokel D.; Rubin L. L.; Peterson R. T.; Schier A. F. (2010) Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation. Science 327(5963), 348–351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richendrfer H.; Pelkowski S. D.; Colwill R. M.; Creton R. (2012) On the edge: pharmacological evidence for anxiety-related behavior in zebrafish larvae. Behav. Brain Res. 228(1), 99–106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schnörr S. J.; Steenbergen P. J.; Richardson M. K.; Champagne D. L. (2012) Measuring thigmotaxis in larval zebrafish. Behav. Brain Res. 228(2), 367–374. [DOI] [PubMed] [Google Scholar]
- Pelkowski S. D.; Kapoor M.; Richendrfer H. A.; Wang X.; Colwill R. M.; et al. (2011) A novel high-throughput imaging system for automater analyses of avoidance behavior in zebrafish larvae. Behav. Brain Res. 223, 135–144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths B. B.; Schoonheim P. J.; Ziv L.; Voelker L.; Baier H.; Gahtan E. (2012) A zebrafish model of glucocorticoid resistance shows serotonergic modulation of the stress response. Front. Behav. Neurosci. 6, 68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyzar E.; Stewart A. M.; Landsman S.; Collins C.; Gebhardt M.; Robinson K.; Kalueff A. V. (2013) Behavioral effects of bidirectional modulators of brain monoamines reserpine and d-amphetamine in zebrafish. Brain Res. 1527, 108–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seibt K. J.; Oliveira R.; da L.; Zimmermann F. F.; Capiotti K. M.; Bogo M. R.; Ghisleni G.; Bonan C. D. (2010) Antipsychotic drugs prevent the motor hyperactivity induced by psychotomimetic MK-801 in zebrafish (Danio rerio). Behav. Brain Res. 214(2), 417–422. [DOI] [PubMed] [Google Scholar]
- Teles M. C.; Dahlbom S. J.; Winberg S.; Oliveira R. F. (2013) Social modulation of brain monoamine levels in zebrafish. Behav. Brain Res. 253, 17–24. [DOI] [PubMed] [Google Scholar]
- Stewart A.; Wu N.; Cachat J.; Hart P.; Gaikwad S.; Wong K.; Utterback E.; Gilder T.; Kyzar E.; Newman A.; Carlos D.; Chang K.; Hook M.; Rhymes C.; Caffery M.; Greenberg M.; Zadina J.; Kalueff A. V. (2011) Pharmacological modulation of anxiety-like phenotypes in adult zebrafish behavioral models. Prog Neuropsychopharmacol. Biol. Psychiatry 35(6), 1421–1431. [DOI] [PubMed] [Google Scholar]
- Airhart M. J.; Lee D. H.; Wilson T. D.; Miller B. E.; Miller M. N.; Skalko R. G. (2007) Movement disorders and neurochemical changes in zebrafish larvae after bath exposure to fluoxetine (PROZAC). Neurotoxicol. Teratol. 29(6), 652–664. [DOI] [PubMed] [Google Scholar]
- Maximino C.; da Silva A. W.; Gouveia A. Jr; Herculano A. M. (2011) Pharmacological analysis of zebrafish (Danio rerio) scototaxis. Prog. Neuropsychopharmacol. Biol. Psychiatry 35(2), 624–631. [DOI] [PubMed] [Google Scholar]
- Wong R. Y.; Oxendine S. E.; Godwin J. (2013) Behavioral and neurogenomic transcriptome changes in wild-derived zebrafish with fluoxetine treatment. BMC Genomics 14, 348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sackerman J.; Donegan J. J.; Cunningham C. S.; Nguyen N. N.; Lawless K.; Long A.; Benno R. H.; Gould G. G. (2010) Zebrafish behavior in novel environments: effects of acute exposure to anxiolytic compounds and choice of Danio rerio line. Int. J. Comp. Psychol. 23(1), 43–61. [PMC free article] [PubMed] [Google Scholar]
- Yang M.; Qiu W.; Chen J.; Zhan J.; Pan C.; Lei X.; Wu M. (2014) Growth inhibition and coordinated physiological regulation of zebrafish (Danio rerio) embryos upon sublethal exposure to antidepressant amitriptyline. Aquat. Toxicol. 151, 68–76. [DOI] [PubMed] [Google Scholar]
- Li J.; Lu W. Q.; Beesley S.; Loudon A. S.; Meng Q. J. (2012) Lithium impacts on the amplitude and period of the molecular circadian clockwork. PLoS One 7, e33292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weger M.; Weger B. D.; Diotel N.; Rastegar S.; Hirota T.; Kay S. A.; Strähle U.; Dickmeis T. (2013) Real-time in vivo monitoring of circadian E-box enhancer activity: a robust and sensitive zebrafish reporter line for developmental, chemical and neural biology of the circadian clock. Dev. Biol. 380(2), 259–73. [DOI] [PubMed] [Google Scholar]
- Jin E. J.; Thibaudeau G. (1999) Effects of lithium on pigmentation in the embryonic zebrafish (Brachydanio rerio). Biochim. Biophys. Acta 1449(1), 93–99. [DOI] [PubMed] [Google Scholar]
- da Luz Oliveira R.; Seibt K. J.; Rico E. P.; Bogo M. R.; Bonan C. D. (2011) Inhibitory effect of lithium on nucleotide hydrolysis and acetylcholinesterase activity in zebrafish (Danio rerio) brain. Neurotoxicol. Teratol. 33(6), 651–657. [DOI] [PubMed] [Google Scholar]
- Aral H.; Vecchio-Sadus A. (2008) Toxicity of lithium to humans and the environment--a literature review. Ecotoxicol. Environ. Saf. 70, 349–356. [DOI] [PubMed] [Google Scholar]
- McKnight R. F.; Adida M.; Budge K.; Stockton S.; Goodwin G. M.; Geddes J. R. (2012) Lithium toxicity profile: a systematic review and meta-analysis. Lancet 379, 721–728. [DOI] [PubMed] [Google Scholar]
- Cohen L. S.; Friedman J. M.; Jefferson J. W.; Johnson E. M.; Weiner M. L. (1994) A reevaluation of risk of in utero exposure of lithium. JAMA, J. Am. Med. Assoc. 271, 146–150. [PubMed] [Google Scholar]
- Gentile S. (2011) Drug treatment for mood disorders in pregnancy. Curr. Opin. Psychiatry 24, 34–40. [DOI] [PubMed] [Google Scholar]
- Yonkers K. A.; Wisner K. L.; Stowe Z.; Leibenluft E.; Cohen L.; Miller L.; et al. (2004) Management of bipolar disorder during pregnancy and the postpartum period. Am. J. Psychiatry 161, 608–620. [DOI] [PubMed] [Google Scholar]
- Schou M.; Goldfield M. D.; Weinstein M. R.; Villeneuve A. (1973) Lithium and pregnancy, I: report from the Register of lithium Babies. Br. Med. J. 2, 135–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schou M.; Amdisen A. (1973) Lithium and pregnancy, 3: lithium ingestion by children breast-fed by women on lithium treatment. Br. Med. J. 2, 138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schou M. (1976) What happened later to the lithium babies? A follow up study of children born without malformations. Acta Psychiatr. Scand. 54, 193–197. [DOI] [PubMed] [Google Scholar]
- Nars P. W.; Girard J. (1977) Lithium carbonate intake during pregnancy leading to large goiter in a premature infant. Am. J. Dis. Child. 131, 924–925. [DOI] [PubMed] [Google Scholar]
- Ananth J. (1978) Side effects in the neonate from psychotropic agents excreted through breast-feeding. Am. J. Psychiatry 135, 801–805. [DOI] [PubMed] [Google Scholar]
- Mizrahi E. M.; Hobbs J. F.; Goldsmith D. I. (1979) Nephrogenic diabetes insipidus in transplacental lithium intoxication. J. Pediatr. 94, 493–495. [DOI] [PubMed] [Google Scholar]
- Jacobson S. J.; Jones K.; Johnson K.; Ceolin L.; Kauer P.; Sah K.; et al. (1992) Prospective multicentre study of pregnancy outcome after lithium exposure during first trimester. Lancet 339, 530–533. [DOI] [PubMed] [Google Scholar]
- Kozma C. (2005) Neonatal toxicity and transient neurodevelopmental deficits following prenatal exposure to lithium: another clinical report and a review of the Literature American Journal of Medical Genetics. Am. J. Med. Genet. 132, 441–444. [DOI] [PubMed] [Google Scholar]
- Zegers B.; Andriessen P. (2003) Maternal lithium therapy and neonatal morbidity. Eur. J. Pediatr. 162, 348–349. [DOI] [PubMed] [Google Scholar]
- Stachel S. E.; Grunwald D. J.; Myers P. Z. (1993) Lithium perturbation and goosecoid expression identify a dorsal specification pathway in the pregastrula zebrafish. Development. 117(4), 1261–1274. [DOI] [PubMed] [Google Scholar]
- Shao M.; Lin Y.; Liu Z.; Zhang Y.; Wang L.; et al. (2012) GSK-3 activity is critical for the orientation of the cortical microtubules and the dorsoventral axis determination in zebrafish embryos. PLoS One 7(5), e36655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selderslaghs I. W. T.; Van Rompay A. R.; De Coen W.; Witters H. E. (2009) Development of a screening assay to identify teratogenic and embryotoxic chemicals using the zebrafish embryo. Reprod. Toxicol. 3, 308–320. [DOI] [PubMed] [Google Scholar]
- Pruvot B.; Quiroz Y.; Voncken A.; Jeanray N.; Piot A.; Martial J. A.; Muller M. (2012) A panel of biological tests reveals developmental effects of pharmaceutical pollutants on late stage zebrafish embryos. Reprod. Toxicol. 34(4), 568–583. [DOI] [PubMed] [Google Scholar]
- Nery L. R.; Eltz N. S.; Martins L.; Guerim L. D.; Pereira T. C.; Bogo M. R.; Vianna M. R. (2014) Sustained behavioral effects of lithium exposure during early development in zebrafish: involvement of the Wnt-β-catenin signaling pathway. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 10.1016/j.pnpbp.2014.04.011. [DOI] [PubMed] [Google Scholar]
- Leng Y.; Liang M.; Ren M.; Marinova Z.; Leeds P.; Chuang D. (2008) Synergistic Neuroprotective Effects of lithium and Valproic Acid or Other Histone Deacetylase Inhibitors in Neurons: Roles of Glycogen Synthase Kinase-3 Inhibition. J. Neurosci. 28, 2576–2588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maguschak K. A.; Ressler K. J. (2008) Beta-catenin is required for memory consolidation. Nat. Neurosci. 11, 1319–1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newport J.; Viguera A. C.; Beach A. J.; Ritchie J. C.; Cohen L. S.; Stowe Z. N. (2005) Lithium placental passage and obstetrical outcome: Implications for clinical management during late pregnancy. Am. J. Psychiatry 162, 2162–2170. [DOI] [PubMed] [Google Scholar]
- Shaldubina A.; Agam G.; Belmaker R. H. (2001) The mechanism of lithium action: state of the art, ten years later. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 4, 855–866. [DOI] [PubMed] [Google Scholar]
- Toledo E. M.; Colombres M.; Inestrosa N. C. (2008) Wnt signaling in neuroprotection and stem cell differentiation. Prog. Neurobiol. 86, 281–296. [DOI] [PubMed] [Google Scholar]
- Phiel C. J.; Klein P. S. (2001) Molecular targets of lithium action. Annu. Rev. Pharmacol. Toxicol. 41, 789–813. [DOI] [PubMed] [Google Scholar]
- Silverstone P. (2005) What is the best way to treat bipolar depression?. J. Psychiatry Neurosci. 30(5), 384. [PMC free article] [PubMed] [Google Scholar]
- Gould T. D.; Manji H. K. (2005) Glycogen synthase kinase-3: a putative molecular target for lithium mimetic drugs. Neuropsychopharmacology 30, 1223–1237. [DOI] [PubMed] [Google Scholar]
- Klein P. S.; Melton D. A. (1996) A molecular mechanism for the effect of lithium on development. Proc. Natl. Acad. Sci. U.S.A. 93, 8455–8459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stambolic V.; Ruel L.; Woodget J. R. (1996) Lithium inhbits glycogen synthase kinase-3 activity and mimics Wingless signaling in intact cells. Curr. Biol. 6, 1664–1668. [DOI] [PubMed] [Google Scholar]
- Wu D.; Pan W. (2009) GSK3: a multifaceted kinase in Wnt signalling. Trends Biochem. Sci. 35(3), 161–168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada A. (2009) Lithium and neuropsychiatric therapeutics: neuroplasticity via glycogen synthase kinase-3β, β-catenin, and neurotrophin cascades. J. Pharmacol. Sci. 110, 14–28. [DOI] [PubMed] [Google Scholar]
- Willert K.; Brown J. D.; Danenberg E.; Duncan A. W.; Weissman I. L.; Reya T.; Yates J. R. 3rd; Nusse R. (2003) Wnt proteins are lipid-modified and can act as stem cell growth factors. Nature 423, 448–452. [DOI] [PubMed] [Google Scholar]
- Polter A.; Beurel E.; Yang S.; Garner R.; Song L.; Miller C. A.; Sweatt J. D.; McMahon L.; Bartolucci A. A.; Li X.; Jope R. S. (2010) Deficiency in the inhibitory serine-phosphorylation of glycogen synthase kinase-3 increases sensitivity to mood disturbances. Neuropsychopharmacology 35(8), 1761–1774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gould T. D.; Einat H.; Bhat R.; Manji H. K. (2004) AR-A014418, a selective GSK-3 inhibitor, produces antidepressant-like effects in the forced swim test. Int. J. Neuropsychopharmacol. 7, 387–390. [DOI] [PubMed] [Google Scholar]
- Kaidanovich-Beilin O.; Lipina T. V.; Takao K.; van Eede M.; Hattori S.; Laliberté C.; Khan M.; Okamoto K.; Chambers J. W.; Fletcher P. J.; MacAulay K.; Doble B. W.; Henkelman M.; Miyakawa T.; Roder J.; Woodgett J. R. (2009) Abnormalities in brain structure and behavior in GSK-3α mutant mice. Mol. Brain 2, 35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clevers H. (2006) Wnt/ß-catenin signaling in development and disease. Cell 125, 469–480. [DOI] [PubMed] [Google Scholar]
- MacDonald B. T.; Tamai K.; He X. (2009) Wnt/β-Catenin Signaling: components, mechanisms, and diseases. Dev. Cell 17, 9–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farías G. G.; Godoy J. A.; Cerpa W.; Varela-Nallar L.; Inestrosa N. C. (2010) Wnt signaling modulates pre- and postsynaptic maturation: Therapeutic considerations. Dev. Dyn. 239(1), 94–101. [DOI] [PubMed] [Google Scholar]
- Inestrosa N. C.; Arenas E. (2010) Emerging roles of wnts inthe adult nervous system. Nat. Rev. Neurosci. 11, 77–86. [DOI] [PubMed] [Google Scholar]