

Abstract
DNA methylation, one of the best-characterized epigenetic modifications, plays essential roles in development, aging and diseases. The de novo DNA methyltransferase DNMT3A is responsible for the establishment of de novo genomic DNA methylation patterns and, as such, involved in normal development as well as in many diseases including cancer. In recent years, our understanding of this important protein has made significant progress, which was facilitated by stunning development in the analysis of the DNA methylome of multiple organs and cell types. In this review, recent developments in the characterization of DNMT3A were discussed with special emphasis on the roles of DNMT3A in development and cancer.
Keywords: epigenetics, DNMT3A, DNA methylation, development, cancer
Introduction
Epigenetic regulation is regarded as the key factor that controls chromatin structure and supports functional organization of the eukaryotic genome through cell divisions.1 As a key epigenetic mark, DNA methylation plays a crucial role in normal development.2 It has been shown to participate in many physiological processes, including cellular differentiation,3 oncogenic transformation,4 and long-term memory.5 DNA methyltransferases (DNMTs) are responsible for the transfer of a methyl group from the universal methyl donor, S-adenosyl-methionine (SAM), to carbon-5 (C5) of cytosine. In mammals, three families of DNMTs have been shown to be involved in DNA methylation. DNMT1 is predominately responsible for the maintenance of genomic DNA methylation patterns. DNMT2 transfers methyl groups to RNA not DNA, and hence has been renamed tRNA aspartic acid methyltransferase 1 (TRDMT1). DNMT3s (de novo DNA methyltransferases, including DNMT3A, DNMT3B, and the regulatory subunit DNMT3L) are responsible for the establishment of genomic DNA methylation patterns.6 The role of the de novo DNMT3s family as a whole in mammalian development has been described.7,8 However, the selectivity and specificity of substrates (such as DNA sequence), and the spatial and temporal activators and regulators may be unique to each DNMT family member. Therefore, their functional and pathological roles in development and disease could be markedly different. Very recently, novel roles of DNMT3A in mammalian development have been identified. This makes it necessary for us to re-assess each member of the DNMT3 family individually. Thus, this review focuses on DNMT3A, a unique member of the DNMT3 family that establishes DNA methylation patterns de novo.
As a de novo DNA methyltransferase, DNMT3A is traditionally considered to be playing an important role in the establishment of methylation patterns during primordial germ cell development and early embryogenesis,9 and behaving as an oncogene in human cancers.10 However, several inactivating mutations of DNMT3A in myeloid malignancies11-23 and loss of DNMT3A activity at advanced tumor stages24 were recently identified. In addition, novel roles of DNMT3A in the hematopoiesis system25 and in the age-related decline of cognition26 have been revealed. These new data suggested that the role of DNMT3A in mammalian development may be more complex than was previously believed, and indicated that in addition to acting as an oncogene in some circumstances, DNMT3A may also behave as a tumor suppressor in other circumstances.
Interestingly, human DNMT3A protein shows 98% amino acid sequence identity to its murine homolog, and contains similar structural domains as mouse Dnmt3a protein. In addition, both human DNMT3A and mouse Dnmt3a possess de novo DNA methylation activity.9,27,28 Therefore, it is reasonable to assume that the functional role of DNMT3A is the same between human and murine. In this review, we provide an overview of recent observations in the characterization and functional studies of DNMT3A in human and murine development and summarize the potential dual roles of DNMT3A protein in cancer.
Molecular Characteristics of DNMT3A
Genetic studies of the DNMT3A gene
Dnmt3a was first cloned and characterized in 1998 by Okano et al.28 In human, the gene encoding DNMT3A is localized at 2p23, and is comprised of 26 exons/25 introns, spanning nearly 110 kb in the genome. In mouse, the Dnmt3a gene, localized in chromosome 12A2–A3, is comprised of 23 exons/22 introns, extending approximately 79 kb in the genome.27,29
DNMT3A variants
In both human and mouse, the DNMT3A (Dnmt3a) gene gives rise to multiple alternative transcripts. Northern blot analyses revealed three distinct tissue-specific splicing variants in human (4.0 kb, 4.4 kb, and 9.5 kb) and in mouse (4.0 kb, 4.2 kb, and 9.5 kb).30 Recently, another variant similar to the 9.5 kb transcript was identified. This variant was named Dnmt3a-β transcript because it was the result of the replacement of exon 1α in the 9.5 kb transcript by exon 1β. Further study revealed that the Dnmt3a-β transcript may be important for establishing DNA methylation patterns during development.31 However, it is still not clear whether these splicing variants could give rise to functional proteins. Chen et al. recently identified a novel short isoform-DNMT3A2 protein (about 82 kD). Transcription of this isoform is initiated from a different promoter in the sixth intron of the DNMT3A gene, which encodes the full-length DNMT3A protein (DNMT3A1, approximately 120 kDa). The mechanism of generation of the other splicing variants remains to be determined.32 Previous studies with knockout technologies addressed effects of the deletion of the Dnmt3a gene without differentiating the two isoforms. Consequently, there is lack of information in the literature describing the action of the two isoforms individually. Therefore, in this review, when only DNMT3A (Dnmt3a) is mentioned, it means the whole DNMT3A (Dnmt3a) gene or the two isoforms (DNMT3A1/2, Dnmt3a1/2) collectively.
The expression pattern of DNMT3A
In mammals, the initial DNA methylation pattern is generated during the early stage of embryonic development. The second wave of de novo DNA methylation occurs during primordial germ cell development.33 Surprisingly, Dnmt3a is indeed highly expressed during primordial germ cell development and blastocyst stages,34 confirming its role as a de novo DNA methyltransferase in mammalian development. However, studies revealed that Dnmt3a1 and Dnmt3a2 have different expression patterns. During male mouse spermatogenesis, Dnmt3a1 and Dnmt3a2 were dynamically expressed (Fig. 1A).35 As development progresses, Dnmt3a2 expression gradually decreases upon differentiation and specifically expressed in testis, spleen and thymus, but silenced in other somatic tissues while Dnmt3a1 is expressed ubiquitously at low levels (Fig. 1B and C).32 So far, little is known about the regulatory mechanisms of the differential expression patterns of DNMT3A1 and DNMT3A2. The only case in testicular cancer study showed that the de-methylation of most CpGs in intron 25 of the DNMT3A gene specifically contributed to the upregulation of DNMT3A2 but not DNMT3A1 isoform. However, it was unclear why the CpGs in intron 25 of the DNMT3A gene were selectively associated with DNMT3A2 expression.36
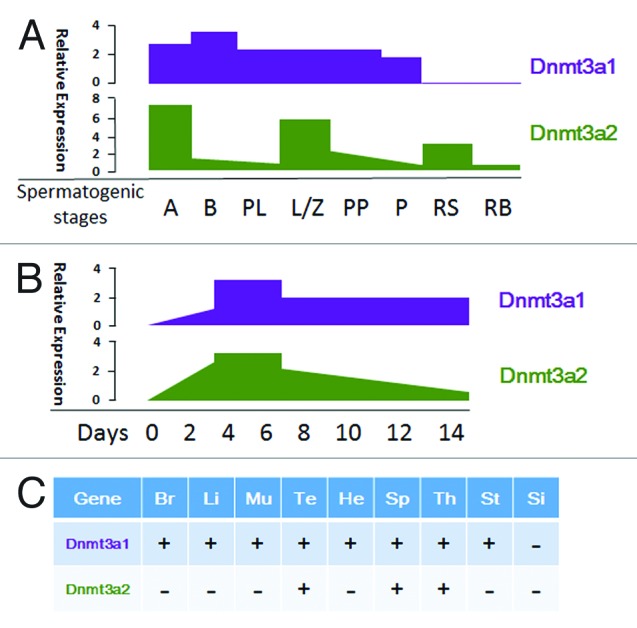
Figure 1. The expression pattern of DNMT3A. (A) The expression of Dnmt3a1 and Dnmt3a2 in male germ cell development. Dnmt3a2 expression was highest in type A spermatogonia, decreased sharply in type B spermatogonia and preleptotene spermatocytes, then went up in leptotene/zygotene spermatocytes to the similar level in type A spermatogonia, followed with a drop again in prepubertal pachytene spermatocytes and pachytene spermatocytes. Moreover, Dnmt3a2 was also expressed in round spermatids and residual bodies/elongating spermatids. Whereas, Dnmt3a1 expression was mostly constant, except for a moderate increase in type B spermatogonia, and showed extremely low levels in round spermatids and residual bodies/elongating spermatids.35 A, type A spermatogonia; B, type B spermatogonia; PL, preleptotene; L/Z, leptotene/zygotene; PP, prepubertal pachytene spermatocytes; P, pachytene spermatocytes; RS, round spermatids; RB, residual bodies/elongating spermatids. (B) The expression of Dnmt3a1 and Dnmt3a2 during mouse ES cells differentiation. Dnmt3a1 and Dnmt3a2 were all upregulated upon differentiation, with the highest level observed in embryoid bodies (EBs) (4–6 d). However, from 6 d onward, Dnmt3a2 expression decreased dramatically, whereas the level of Dnmt3a1 sustained throughout the examined time-course.32 (C) Dnmt3a1 and Dnmt3a2 expression in mouse tissues (3 wks. old).32 Br, brain; Li, liver; Mu, muscle; Te, testis; Ht, heart; Sp, spleen; Th, thymus; St, stomach; Si, small intestine; +, express; -, not express.
The sub-nuclear localization of DNMT3A
DNMT3A1 is found stably associated with chromatin containing methylated CGIs (CpG islands) and repetitive DNA elements,37 and concentrated in pericentromeric heterochromatin, which is considered to be transcriptionally silent.38,39 These observations suggest that DNMT3A1 may function primarily as a transcription repressor. In contrast, DNMT3A2 is associated with transcriptionally active euchromatin.32 Since euchromatin contains mostly actively transcribed genes, and silencing of some transcribed genes (such as tumor suppressor genes) for hypermethylated gene promoter and/or hypomethylated gene body contributes to diseases such as human cancers, it is exciting to observe the association of DNMT3A2 with euchromatin, suggesting a potential role of DNMT3A2 in human diseases.32,40 However, the mechanism underlying the different sub-nuclear localization of DNMT3A1 and DNMT3A2 remains largely unknown. Recently, Chantalat et al. revealed that H3K36me3, previously identified to be associated with actively transcribed regions, may also contribute to the composition of heterochromatin.41 Considering the DNMT3A PWWP domain-H3K36me3 interaction,42 this finding may partially explain why DNMT3A1 and DNMT3A2 have different sub-nuclear localization.
Structural domains of DNMT3A protein
Overall, the major architectural features of DNMT3A are the N-terminal regulatory region and the C-terminal catalytic region.43 Figure 2 shows the structural domains of DNMT3A and their interacting partners.
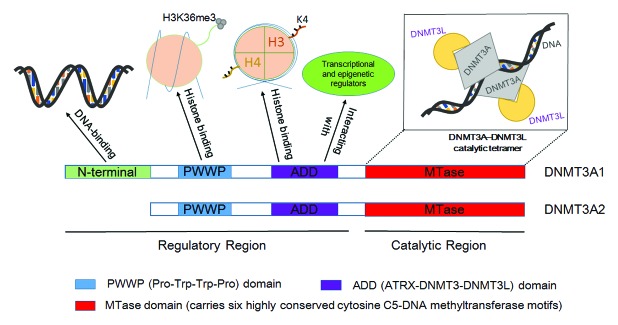
Figure 2. Structural domains of DNMT3A protein. Both DNMT3A isoforms (DNMT3A1 and DNMT3A2) have N-terminal regulatory region and C-terminal catalytic region. A 223 (human) or 219 (mouse) amino acid N-terminal domain, which was shown to be able to bind to DNA sequence, is present only in DNMT3A1.32 The N-terminal regulatory region consists of a moderately conserved PWWP domain and a cysteine-rich (Cys-rich) ADD domain. The PWWP domain of DNMT3A is showed to directly interact with the H3K36m3 mark,42 while the ADD domain associates with transcriptional factors and epigenetic regulators, and interacts specifically with unmethylated H3K4.57,58 Finally, the MTase domain resides in the C-terminal region. The oligomerization of DNMT3A and DNMT3L at this structurally conserved domain controls the catalytic mechanism of de novo DNA methylation.69
The regulatory region of both DNMT3A isoforms contains two defined domains: the PWWP (Pro-Trp-Trp-Pro) domain, and the ADD (ATRX-DNMT3-DNMT3L) domain. However, a 223 (human) or 219 (mouse) amino acid N-terminal non-catalytic domain is present only in DNMT3A1.32 To elucidate the properties of the different N-terminal domain in Dnmt3a1, Suetake et al. purified the 219 amino acid residues of Dnmt3a1 N-terminal domain and found that the N-terminal 1–211 polypeptide was able to bind to DNA, irrespective of the methylation status of targeted DNA. Thus, DNMT3A1 showed higher DNA-binding and DNA-methylation activities than DNMT3A2.44 Moreover, Choi et al. recently conducted a high-throughput gene-specific DNA methylation analysis to compare the methylome of HEK 293T cells that were stably transfected with the two DNMT3A isoforms separately. They revealed that DNMT3A1 and DNMT3A2 shared 93% de novo DNA methylation target sites,45 indicating that the two isoforms, albeit behaving differently, shared catalytic function as de novo DNA methyltransferase.
The PWWP domain is located at the N-terminal of the cysteine-rich (Cys-rich) domain and spans 100–150 amino acids, containing a conserved proline-tryptophan motif (hence the name PWWP). This domain has been found in over 60 eukaryotic proteins, especially in chromatin associated proteins.46 It is generally accepted that the PWWP domain is essential for targeting DNMT3A to pericentric heterochromatin and metaphase chromosomes, although the mechanism remains unknown.38,47 Surprisingly, similar to the ADD domain, the PWWP domain of Dnmt3a has also been shown to directly interact with the H3K36me3 mark,42 which contributes to the composition of heterochromatin.41 These observations may explain the role of the PWWP domain in guiding DNMT3A to pericentric heterochromatin. Furthermore, it was recently reported that transcribed genes carrying intragenic H3K36me3 are totally unmethylated in the promoter and hypermethylated within the gene body,48 and Dnmt3a regulates gene expression via gene body DNA methylation.49 Thus, PWWP-H3K36me3 interaction may also be partially responsible for Dnmt3a-mediated gene body DNA methylation in gene expression regulation.
The ADD domain is a Cys-rich zinc-finger binding domain related to the plant homeodomain (PHD) family, which processes a C2-C2 zinc-finger that binds zinc ions and contains a PHD motif involved in various protein–protein interactions.50 Recent evidence suggests that this cysteine-rich domain mediates the interaction of DNMT3A with several essential transcriptional factors and epigenetic regulators, including PU.1,51 Myc,52 p53,53 RP58,54 HDAC1,54 HP1,55 EZH2,56 SUV39H1,55 contributing to DNA methylation at specific sites during development. In addition, the ADD domain of Dnmt3a has been shown to interact specifically with histone H3 tails unmethylated at the N-terminal lysine 4 (unmethylated H3K4). The interaction could be disrupted by the methylation of H3K4, indicating that the ADD domain of DNMT3A could be a guide for DNA methylation in response to specific histone modifications.57,58
The C-terminal catalytic region of DNMT3A harbors six conserved motifs characteristic of cytosine C5-DNA methyltransferase. Unlike DNMT1, the isolated C-terminal domain of DNMT3A is catalytically active.59 Cheng et al. recently showed that the structure of Dnmt3a C-terminal domain is remarkably similar to the bacterial M. HhaI enzyme, a well-studied prokaryotic DNA methyltransferase.8 It is questionable whether the same catalytic mechanism applies to DNMT3A. Strikingly, it was indeed reported that the Dnmt3a C-terminal could bind to DNA sequence with a flipped target cytosine (Cyt), and the target Cyt precisely locates between the catalytic nucleophile (Cys706) of Dnmt3a C-terminal and methyl donor SAM. Then the Cys706 and SAM would attack the target Cyt C6 from opposite directions, similar to how M. HhaI acts. Therefore, the mammalian DNMT3A has the same catalytic mechanism as the bacterial homolog.8 Moreover, the DNA methyltransferase-like protein Dnmt3l, the third member of Dnmt3 family, has been shown to directly interact with the C-terminal catalytic domains of Dnmt3a via its C-terminal, and stimulate the enzymatic activity of Dnmt3a both in vivo and in vitro.60-62
DNMT3A and DNMT3L Cooperate to Establish DNA Methylation Patterns
In mammals, the de novo DNA methyltransferases DNMT3A and DNMT3B contribute to the establishment of DNA methylation patterns.9 On the other hand, DNMT3L, the third member of DNMT3 family, discovered later, was shown to cooperate with DNMT3A/3B to set de novo DNA methylation patterns.60,63
It was indeed reported that Dnmt3l co-localized and directly interacted with Dnmt3a,60 and co-expression of human DNMT3L with murine Dnmt3a resulted in a striking stimulation of de novo methylation by Dnmt3a.61 Subsequent study investigating the mechanism underlying this stimulation revealed that DNMT3L significantly increased the affinity of DNMT3A for SAM (methyl donor) and the processivity of DNMT3A up to 3-fold, but did not modify its binding activity to targeted DNA sequence.64,65 Interestingly, the interaction of DNMT3L with DNMT3A in the de novo DNA methylation process contributed to more methylated sites, of which most newly methylated sites were intrinsically poorly methylated by DNMT3A.66 Structural studies showed that the C-terminal domain of Dnmt3l directly interact with the C-catalytic domain of Dnmt3a to form a Dnmt3a-Dnmt3l heterodimer. Since Dnmt3a C-terminal also mediates the Dnmt3a-Dnmt3a contact, two Dnmt3a-3l heterodimer could further form a Dnmt3l:Dnmt3a:Dnmt3a:Dnmt3l heterotetramer (Fig. 2).67,68 Moreover, DNMT3A homodimer of the heterotetramer complex showed specific contacts with the DNA substrate and substitution of key non-catalytic residues at the Dnmt3a-Dnmt3l interface or the Dnmt3a-Dnmt3a interface eliminated enzymatic activity.14,68,69 However, the functional significance of DNMT3A oligomerization remains largely unknown. Recently, taking advantage of the homotetramer DNMT3A model, Holz-Schietinger et al. designed mutations at the tetramer interface (DNMT3A-3L interface) to disrupt tetramerization, and found that mutant DNMT3A could form homodimer with comparable activity to the wild type homotetramer, but with altered catalytic properties. The homotetramer DNMT3A with disrupted tetramer interface hold faster rate of product release. However, binding of DNMT3L to the mutated DNMT3A dimer would decrease the rate of product release and restored the catalytic properties.69 Since the interactions of DNMT3A with accessory proteins were critical for the setting of DNA methylation patterns,60,70 and tetramer interface of the DNMT3A homodimer was essential for DNMT3L binding, the mutations at the tetramer interface would disrupt the binding of DNMT3L to DNMT3A homodimer and lead to aberrant DNA methylation. Therefore, the oligomerization of DNMT3A with DNMT3L controls the catalytic mechanism of de novo DNA methylation.69
Collectively, these data suggest that DNMT3L cooperates with DNMT3A in establishing DNA methylation patterns, and DNMT3L stimulates the DNA methylation activity of DNMT3A via direct interaction. However, the stimulation of the DNMT3A catalytic activity by DNMT3L still leaves an open question, that whether the interactions of DNMT3A with other cellular proteins or nucleic acids also contribute to the regulation of its enzyme activity.
The Role of DNMT3A During Human and Mouse Development
Development from a zygote into a complex, multicellular adult organism requires an array of shared and specific cellular processes.71 The refined regulation of gene expression is required to shape developmental decisions. As a well-studied epigenetic modification, DNA methylation is essential for mammalian development.9 Recently, knockout technologies were used to study the roles of DNMT3A in global, as well as development of special organ and special cell type (see Table 1). The developmental roles of DNMT3A at different stages will be reviewed in the following paragraph.
Table 1. Phenotypes of Dnmt3a knockout (KO).
| Genotype | Organ type/Cell type | Phenotype | References | ||
|---|---|---|---|---|---|
| Preimplantation development |
Postimplantation development |
Postnatal development | |||
| Dnmt3a+/− | Whole body | Normal | Normal | Grossly normal and fertile | 9 |
| Dnmt3a−/− | Whole body | Normal | Normal | Runted, died at about 4 wks of age, gastrointestinal dysfunction, gait ataxia, reduced germ cell counts in male. | 9 |
| Dnmt3a−/− | Conditional KO in primordial germ cells | Mutant female: Offspring (mutant oocyte was fertilized with wild-type sperm) will die in utero. | Mutant males: Impaired spermatogenesis. | 74 | |
| Dnmt3a−/− | Conditional KO in forebrain excitatory neurons | No defects in learning and memory | 82 | ||
| Dnmt3a−/− | Conditional KO in nervous system | Motor neuron defects and die prematurely | 78 | ||
|
K-rasLSL-G12D /Dnmt3a2Lox/2Lox |
Conditional KO in lung of mice with lung cancer | Dnmt3a deletion accelerates mutant K-ras induced lung tumor growth | 24 | ||
| Dnmt3a−/− | mESCs | Precocious Gliogenesis, globally hypomethylated and increased proliferation from passage 6 onward | 76 | ||
| Dnmt3a−/− | HSCs | Increased to differentiated hematopoietic lineages | 25 | ||
| Dnmt3a−/− | NSCs | Impaired neurogenic potential | 79 | ||
mESCs, mouse embryonic stem cells; HSCs, hematopoietic stem cells; NSCs, neural stem cells
DNMT3A in primordial germ cells
The methylome of germ cells changes dynamically during development. These epigenetic changes contributed to the re-establishment of parental imprints that are required for the regulation of meiosis and genomic integrity.72 During male germ cell development Dnmt3a2 was dynamically expressed while Dnmt3a1 expression was mostly constant, except in type B spermatogonia where it increased.35 In addition, Dnmt3l, which is closely related to spermatogenesis and stimulates the DNA methylation catalytic activity of Dnmt3a, was shown to co-express with Dnmt3a2 in mouse germ cells at different stages.73 Evidence from Dnmt3a conditional knockout study revealed that both Dnmt3a-deficient male and female mice presented with impaired reproductive potential. Dnmt3a conditional knockout males were sterile due to azoospermia and lacked methylation at the H19 DMR (H19 differentially methylated region) and IG-DMR (intergenic-differentially methylated region) paternally imprinted loci in spermatogonia, indicating deficiency of Dnmt3a impaired spermatogenesis. Although the Dnmt3a conditional knockout females produced mature oocytes, their offspring (mutant oocyte was fertilized with wild type sperm) died in utero. In addition, Dnmt3a-knockout oocyte showed loss of methylation at dispersed IAP (inhibitors of apoptosis) repeats, indicating that this class of repeated elements may be the specific target of Dnmt3a.7,74 Collectively, these data provided evidence that Dnmt3a is critical for normal spermatogenesis and production of functional oocytes. Importantly, the Dnmt3a isoforms may function differentially in the establishment and/or maintenance of methylation patterns in mouse germ cells.
DNMT3A in embryos and embryonic stem cells (ESCs)
In post-implantation embryos of the mouse, demethylation of minor satellite repeats and dispersed repetitive elements (including C-type retroviral DNA and IAP) were not observed in Dnmt3a−/− embryos, but showed in double knockout embryos (Dnmt3a−/−, Dnmt3b−/−). Similar results were also observed in ESCs with Dnmt3a deficiency, which showed no effect on global DNA methylation.9 However, in highly demethylated ESCs (DNMT3A−/−, DNMT3B−/−), re-expression of DNMT3A1 or DNMT3A2 would re-methylate the tandem and dispersed repeat elements, as well as several single-copy genes.75 These observations confirmed that DNMT3A is the engine of de novo DNA methylation, and indicated that minor satellite repeats may not be the specific targets of DNMT3A.7
Furthermore, in vitro differentiation of mouse embryonic stem cells (mESCs) was affected when Dnmt3a was knocked out. Wu et al. found that at passage 6 (P6) when wild type mESCs derived neural stem cells (NSCs) became gliogenic, Dnmt3a-deficient mESCs derived NSCs showed a robust phenotype of precocious astrocyte and oligodendrocyte differentiation. From P6 onward, Dnmt3a-deficient mESCs derived NSCs had a significantly increased proliferation rate compared with wild type mESCs derived NSCs. In addition, global levels of methylcytosine reduced in P6 Dnmt3a-deficient mESCs derived NSCs.76
DNMT3A in postnatal development
Dynamic changes of the DNA methylome play a critical role in establishing and maintaining cellular phenotype during differentiation. As postnatal development progresses, tissue-specific DNA methylomes are established in multiple organs and tissues. With the help of conditional knockout technology and stunning development in the analysis of the human methylome, the developmental roles of DNMT3A have been evaluated in special organs and cell types.
In the mouse central nervous system (CNS), Dnmt3a was highly expressed in multiple neuronal tissues during early stage of postnatal development (first 3 wk), and then declined to a relatively low level in adulthood, suggesting an important role for Dnmt3a in mouse CNS maturation.77 Subsequently, Nguyen et al. conditionally ablated Dnmt3a in nervous system to clarify its specific role. Strikingly, the conditional mutant mouse were born healthy but died prematurely, and specifically showed motion abnormalities, which were attributed to neuromuscular defects caused by Dnmt3a loss.78 Recently, it was revealed that loss of Dnmt3a impaired the neurogenic potential of neural stem cells (NSCs). Further ChIP-chip results indicated that Dnmt3a promoted the transcription of neurogenic pluripotency genes (including Dlx2, Sp8, and Neurog2) via Dnmt3a-dependent non-proximal promoter DNA methylation. However, the mechanism underlying non-promoter DNA methylation by Dnmt3a remains to be clarified.79 In human, the full-length isoform DNMT3A1, but not the short-length isoform DNMT3A2, was expressed in mature cerebral cortex across the whole lifespan (fetal, child, and adult), which was consistent with the reported different expression patterns of DNMT3A1 and DNMT3A2.32,80
Since the formation and storage of memory require long-lasting changes in memory-related neuronal circuits,81 it is questionable whether DNMT3A is involved in this complex process. However, Dnmt3a deficiency has not been shown to have deleterious effects on synaptic plasticity, learning and memory.82 In contrast, Oliveira et al. reported recently that Dnmt3a2, the shorter isoform encoded by the Dnmt3a gene, controlled cognitive abilities in mice.26 This effect of Dnmt3a2 might not have been investigated in previous studies.82 Using gain- and loss-of-function strategies, they found that overexpression of Dnmt3a2 in the hippocampal brain area of elderly mice restored their cognitive skills in long-term-memory test. Reducing Dnmt3a2 expression in the hippocampus caused poor performance in young adult mice in long-term-memory test when compared with controls of the same age.26 These findings suggested that the special isoform Dnmt3a2 played a predominant role in the age-related decline of cognition.
Recent evidence suggested that Dnmt3a is also associated with postnatal development of the hematopoietic lineage. Dnmt3a has been previously reported to be dispensable for both hematopoietic stem cells (HSCs) self-renewal and differentiation.83 However, using a conditional knockout mouse model coupled with serial transplantation, Challen at al. re-evaluated the role of DNMT3A in HSCs.25 In primary transplant recipients, Dnmt3a−/− HSCs contributed normally to hematopoiesis, as reported in previous studies.83 Whereas in the secondary transplant recipients, Dnmt3a−/− HSCs showed a parallel increase in contribution to differentiated hematopoietic lineages, suggesting that an intrinsic self-renewal program was activated. Transcriptome analysis showed that Dnmt3a loss led to upregulated expression of “HSC fingerprint” genes (including Gata3, Runx1, Pbx1, and Cdkn1a) and decreased expression of genes related to HSC differentiation (such as Flk2, Ikaros, PU.1, and Mef2c).25 This study highlighted a previously unknown function of Dnmt3a in the repression of the stem cell program, i.e., by silencing HSC genes (multipotency genes) and permitting differentiation, presumably via regulation of DNA methylation.
The Dual Roles of DNMT3A in Cancer
In cancer, global DNA methylation patterns are altered, accompanied by the hypermethylation of CpG islands and the hypomethylation of non-CpG islands at promoters of specific genes.84 This alteration is largely dependent on the action of de novo DNMTs during early tumor progression.85 De novo DNMTs were previously suggested to promote tumorigenesis.86 Early studies reported that the DNMT3A overexpressed in multiple tumor types,87-90 and a poorer recurrence-free survival was noted in HCC (Hepatic Cellular Cancer) patients with >4-fold increase in DNMT3A mRNA.90 However, the mechanism underlying the promotion of tumorigenesis by DNMT3A is still unknown. One hypothesis is that elevated DNMT3A may contribute to the silencing of some critical tumor suppressor genes (TSGs) during tumorigenesis. Indeed, in mouse melanoma models, it was demonstrated that after knocking down DNMT3A using small interfering RNA (siRNA), melanoma growth and metastasis were dramatically inhibited. Genome-wide expression profiling revealed inhibition of DNMT3A upregulated the expression of various genes involved in the immune response in melanoma.87 Similarly, depletion of DNMT3A was shown to suppress HCC cell proliferation and colony-formation. This was believed to be attributed to the DNMT3A-induced upregulation of various TSGs (including PTEN) that are involved in cell cycle regulation, transcription regulation, and signal transduction.89
Although previous studies clearly pointed to an oncogenic role of DNMT3A in cancer, the recent discovery of genetic mutations of DNMT3A in acute myeloid leukemia (AML) suggested that its role in cancers may be more complex than was previously believed.11-23 These highly recurrent DNMT3A mutations were closely associated with poor outcome in patients with AML. However, the functional implications of these mutations are still poorly understood. Notably, in vitro studies using purified mutant protein revealed that the majority of somatic DNMT3A mutations resulted either in premature truncation of the protein product (nonsense or frame shift mutations), or decreased catalytic activity and DNA binding affinity (mutation occurs at a single amino acid, R882).14,22 Such DNMT3A-mutation-caused alternations would affect global DNA methylation pattern and expression of HOXA genes,22 which had been reported to have an important role in the pathogenesis of AML.91 Intriguingly, DNMT3A-mutant AMLs did not result in differences in global 5mC levels when compared with wild type DNMT3A AMLs. It was recently reported that 90.8% of de novo methylated loci of DNMT3A and DNMT3B overlapped, indicating that the two enzymes have overlapping function. Therefore, the supplement of DNMT3B in the case of DNMT3A mutation may explain the phenomenon.45 Further MeDIP-chip analysis and genome-wide gene expression profiling revealed a total of 182 genomic regions with significantly altered methylation levels and multiple differentially expressed gene clusters for DNMT3A mutation-AML genomes when compared with matched AML genomes. Whereas, the differentially methylated regions were not associated with local changes in gene expression, and DNMT3A mutation-AMLs did not define any identical differentially expressed gene clusters. These observations led Ley et al. to speculate that DNMT3A mutations might not affect the cytosine methyltransferase properties of DNMT3A.14 In addition to the inactivating mutations of DNMT3A in AML, two recently published studies showed that loss of Dnmt3a activity promoted mutant K-ras induced mouse lung tumor progression.24,49 To clarify the role of Dnmt3a, Gao et al. compared the transcriptome between Dnmt3a-deficient and wild type in mutant K-ras induced mouse lung tumors, and revealed that Dnmt3a was responsible for the maintenance of the methylation-dependent repression of specific oncogenes which were involved in key steps of lung tumor progression, such as angiogenesis, cell adhesion, and cell motion in cancer cells.24 The same group subsequently applied whole-genome bisulfite sequencing to comprehensively characterize the methylomes of wild type and Dnmt3a-deficient mutant K-ras induced mouse lung tumors. Of note, the tumor-specific PMDs (partially methylated domains), identified from control lung vs. Dnmt3a wild type mutant K-ras induced mouse lung tumor, were uniformly hypomethylated in Dnmt3a-deficient mutant K-ras induced mouse lung tumor, thus Dnmt3a may protect these PMDs from hypomethylation. Further analysis of Dnmt3a-protected domains revealed amount of dysregulated genes with characterization of upregulated genes showed hypomethylation in 5′ UTRs and downregulated genes with hypomethylation in exons. These data unveiled a previously unknown role of Dnmt3a in maintaining DNA methylation patterns and suggested a novel gene regulation mechanism of Dnmt3a-dependent gene body DNA methylation in cancer.49
Taken together, these studies showed that the dual roles of DNMT3A in cancer suppression or growth promotion much depend on that DNMT3A represses or activates specific targeted genes by affecting DNA methylation patterns.
Future Prospects and Challenges
The knowledge of the function of DNMT3A has increased considerably since its discovery about 15 y ago. We tried to provide an up-to-date overview of our understanding of the functional roles of DNMT3A in development and cancer. It is now clear that the de novo DNA methyltransferase DNMT3A plays key roles in setting de novo DNA methylation patterns genome-wide during mammalian development, while DNMT3A alterations and associated aberrant DNA methylation patterns would cause multiple types of cancer. Further investigations are needed to explore the molecular mechanisms of DNMT3A alteration in different types of cancer, such as miR-29 family members targeting DNMT3A in AML,92,93 and miR-199a-3p targeting DNMT3A2 in testicular cancer,94 which will be of significance in the understanding of the relevance and mechanisms of methylation changes seen in human cancers. Moreover, DNMT3A1 and DNMT3A2 shared 93% DNA methylation target sites,45 but showed different expression patterns as postnatal development progresses.32,77 Therefore, it is conceivable to propose that DNMT3A1 and DNMT3A2 may possess common and unique functions in development. However, previous studies have only addressed deletion of DNMT3A as a whole by gene targeting in conditional knockout (or knockout) models, it remains unknown which of the isoforms or both are the key factors of relevant phenotypes. Thus, more studies using conditional knockdown of DNMT3A1 or DNMT3A2 are necessary to decipher more stage- and organ- specific function of one or both specific isoforms of DNMT3A. Finally, based on the current data from animal models, more translation studies are needed to confirm the clinical feasibility of alleviating diseases by pharmacologically affecting levels of DNMT3A1, DNMT3A2 or both.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
This research was supported by funds provided by the Chinese University of Hong Kong, Vigconic Ltd, International, and the CUHK-Shandong University Joint Laboratory.
Acknowledgments
We gratefully acknowledge the assistance of Dr Yick-Keung Suen in the laboratory and Mr Chun-Shui Luk in carefully reading our manuscript.
References
- 1.Chen T, Hevi S, Gay F, Tsujimoto N, He T, Zhang B, Ueda Y, Li E. Complete inactivation of DNMT1 leads to mitotic catastrophe in human cancer cells. Nat Genet. 2007;39:391–6. doi: 10.1038/ng1982. [DOI] [PubMed] [Google Scholar]
- 2.Goll MG, Bestor TH. Eukaryotic cytosine methyltransferases. Annu Rev Biochem. 2005;74:481–514. doi: 10.1146/annurev.biochem.74.010904.153721. [DOI] [PubMed] [Google Scholar]
- 3.Reik W, Dean W, Walter J. Epigenetic reprogramming in mammalian development. Science. 2001;293:1089–93. doi: 10.1126/science.1063443. [DOI] [PubMed] [Google Scholar]
- 4.Esteller M. Epigenetics provides a new generation of oncogenes and tumour-suppressor genes. Br J Cancer. 2007;96(Suppl):R26–30. [PubMed] [Google Scholar]
- 5.Miller CA, Sweatt JD. Covalent modification of DNA regulates memory formation. Neuron. 2007;53:857–69. doi: 10.1016/j.neuron.2007.02.022. [DOI] [PubMed] [Google Scholar]
- 6.Lechner M, Boshoff C, Beck S. Cancer epigenome. Adv Genet. 2010;70:247–76. doi: 10.1016/B978-0-12-380866-0.60009-5. [DOI] [PubMed] [Google Scholar]
- 7.Chédin F. The DNMT3 family of mammalian de novo DNA methyltransferases. Prog Mol Biol Transl Sci. 2011;101:255–85. doi: 10.1016/B978-0-12-387685-0.00007-X. [DOI] [PubMed] [Google Scholar]
- 8.Cheng X, Blumenthal RM. Mammalian DNA methyltransferases: a structural perspective. Structure. 2008;16:341–50. doi: 10.1016/j.str.2008.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Okano M, Bell DW, Haber DA, Li E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell. 1999;99:247–57. doi: 10.1016/S0092-8674(00)81656-6. [DOI] [PubMed] [Google Scholar]
- 10.Fernandez AF, Huidobro C, Fraga MF. De novo DNA methyltransferases: oncogenes, tumor suppressors, or both? Trends Genet. 2012;28:474–9. doi: 10.1016/j.tig.2012.05.006. [DOI] [PubMed] [Google Scholar]
- 11.Singh RR, Bains A, Patel KP, Rahimi H, Barkoh BA, Paladugu A, Bisrat T, Ravandi-Kashani F, Cortes JE, Kantarjian HM, et al. Detection of high-frequency and novel DNMT3A mutations in acute myeloid leukemia by high-resolution melting curve analysis. J Mol Diagn. 2012;14:336–45. doi: 10.1016/j.jmoldx.2012.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lu Q, Chen Y, Wang H, Li Z. DNMT3A mutations and clinical features in Chinese patients with acute myeloid leukemia. Cancer Cell Int. 2013;13:1. doi: 10.1186/1475-2867-13-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Metzeler KH, Walker A, Geyer S, Garzon R, Klisovic RB, Bloomfield CD, Blum W, Marcucci G. DNMT3A mutations and response to the hypomethylating agent decitabine in acute myeloid leukemia. Leukemia. 2012;26:1106–7. doi: 10.1038/leu.2011.342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ley TJ, Ding L, Walter MJ, McLellan MD, Lamprecht T, Larson DE, Kandoth C, Payton JE, Baty J, Welch J, et al. DNMT3A mutations in acute myeloid leukemia. N Engl J Med. 2010;363:2424–33. doi: 10.1056/NEJMoa1005143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hou HA, Kuo YY, Liu CY, Chou WC, Lee MC, Chen CY, Lin LI, Tseng MH, Huang CF, Chiang YC, et al. DNMT3A mutations in acute myeloid leukemia: stability during disease evolution and clinical implications. Blood. 2012;119:559–68. doi: 10.1182/blood-2011-07-369934. [DOI] [PubMed] [Google Scholar]
- 16.Fried I, Bodner C, Pichler MM, Lind K, Beham-Schmid C, Quehenberger F, Sperr WR, Linkesch W, Sill H, Wölfler A. Frequency, onset and clinical impact of somatic DNMT3A mutations in therapy-related and secondary acute myeloid leukemia. Haematologica. 2012;97:246–50. doi: 10.3324/haematol.2011.051581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liang DC, Liu HC, Yang CP, Jaing TH, Hung IJ, Yeh TC, Chen SH, Hou JY, Huang YJ, Shih YS, et al. Cooperating gene mutations in childhood acute myeloid leukemia with special reference on mutations of ASXL1, TET2, IDH1, IDH2, and DNMT3A. Blood. 2013;121:2988–95. doi: 10.1182/blood-2012-06-436782. [DOI] [PubMed] [Google Scholar]
- 18.Ostronoff F, Othus M, Ho PA, Kutny M, Geraghty DE, Petersdorf SH, Godwin JE, Willman CL, Radich JP, Appelbaum FR, et al. Mutations in the DNMT3A exon 23 independently predict poor outcome in older patients with acute myeloid leukemia: a SWOG report. Leukemia. 2013;27:238–41. doi: 10.1038/leu.2012.168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pezzi A, Moraes L, Valim V, Amorin B, Melchiades G, Oliveira F, da Silva MA, Matte U, Pombo-de-Oliveira MS, Bittencourt R, et al. DNMT3A Mutations in Patients with Acute Myeloid Leukemia in South Brazil. Adv Hematol. 2012;2012:697691. doi: 10.1155/2012/697691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wakita S, Yamaguchi H, Omori I, Terada K, Ueda T, Manabe E, Kurosawa S, Iida S, Ibaraki T, Sato Y, et al. Mutations of the epigenetics-modifying gene (DNMT3a, TET2, IDH1/2) at diagnosis may induce FLT3-ITD at relapse in de novo acute myeloid leukemia. Leukemia. 2013;27:1044–52. doi: 10.1038/leu.2012.317. [DOI] [PubMed] [Google Scholar]
- 21.Thol F, Damm F, Lüdeking A, Winschel C, Wagner K, Morgan M, Yun H, Göhring G, Schlegelberger B, Hoelzer D, et al. Incidence and prognostic influence of DNMT3A mutations in acute myeloid leukemia. J Clin Oncol. 2011;29:2889–96. doi: 10.1200/JCO.2011.35.4894. [DOI] [PubMed] [Google Scholar]
- 22.Yan XJ, Xu J, Gu ZH, Pan CM, Lu G, Shen Y, Shi JY, Zhu YM, Tang L, Zhang XW, et al. Exome sequencing identifies somatic mutations of DNA methyltransferase gene DNMT3A in acute monocytic leukemia. Nat Genet. 2011;43:309–15. doi: 10.1038/ng.788. [DOI] [PubMed] [Google Scholar]
- 23.Walter MJ, Ding L, Shen D, Shao J, Grillot M, McLellan M, Fulton R, Schmidt H, Kalicki-Veizer J, O’Laughlin M, et al. Recurrent DNMT3A mutations in patients with myelodysplastic syndromes. Leukemia. 2011;25:1153–8. doi: 10.1038/leu.2011.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gao Q, Steine EJ, Barrasa MI, Hockemeyer D, Pawlak M, Fu D, Reddy S, Bell GW, Jaenisch R. Deletion of the de novo DNA methyltransferase Dnmt3a promotes lung tumor progression. Proc Natl Acad Sci U S A. 2011;108:18061–6. doi: 10.1073/pnas.1114946108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Challen GA, Sun D, Jeong M, Luo M, Jelinek J, Berg JS, Bock C, Vasanthakumar A, Gu H, Xi Y, et al. Dnmt3a is essential for hematopoietic stem cell differentiation. Nat Genet. 2012;44:23–31. doi: 10.1038/ng.1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Oliveira AM, Hemstedt TJ, Bading H. Rescue of aging-associated decline in Dnmt3a2 expression restores cognitive abilities. Nat Neurosci. 2012;15:1111–3. doi: 10.1038/nn.3151. [DOI] [PubMed] [Google Scholar]
- 27.Xie S, Wang Z, Okano M, Nogami M, Li Y, He WW, Okumura K, Li E. Cloning, expression and chromosome locations of the human DNMT3 gene family. Gene. 1999;236:87–95. doi: 10.1016/S0378-1119(99)00252-8. [DOI] [PubMed] [Google Scholar]
- 28.Okano M, Xie S, Li E. Cloning and characterization of a family of novel mammalian DNA (cytosine-5) methyltransferases. Nat Genet. 1998;19:219–20. doi: 10.1038/890. [DOI] [PubMed] [Google Scholar]
- 29.Okano M, Takebayashi S, Okumura K, Li E. Assignment of cytosine-5 DNA methyltransferases Dnmt3a and Dnmt3b to mouse chromosome bands 12A2-A3 and 2H1 by in situ hybridization. Cytogenet Cell Genet. 1999;86:333–4. doi: 10.1159/000015331. [DOI] [PubMed] [Google Scholar]
- 30.Robertson KD, Uzvolgyi E, Liang G, Talmadge C, Sumegi J, Gonzales FA, Jones PA. The human DNA methyltransferases (DNMTs) 1, 3a and 3b: coordinate mRNA expression in normal tissues and overexpression in tumors. Nucleic Acids Res. 1999;27:2291–8. doi: 10.1093/nar/27.11.2291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Weisenberger DJ, Velicescu M, Preciado-Lopez MA, Gonzales FA, Tsai YC, Liang G, Jones PA. Identification and characterization of alternatively spliced variants of DNA methyltransferase 3a in mammalian cells. Gene. 2002;298:91–9. doi: 10.1016/S0378-1119(02)00976-9. [DOI] [PubMed] [Google Scholar]
- 32.Chen T, Ueda Y, Xie S, Li E. A novel Dnmt3a isoform produced from an alternative promoter localizes to euchromatin and its expression correlates with active de novo methylation. J Biol Chem. 2002;277:38746–54. doi: 10.1074/jbc.M205312200. [DOI] [PubMed] [Google Scholar]
- 33.Jaenisch R, Harbers K, Jähner D, Stewart C, Stuhlmann H. DNA methylation, retroviruses, and embryogenesis. J Cell Biochem. 1982;20:331–6. doi: 10.1002/jcb.240200403. [DOI] [PubMed] [Google Scholar]
- 34.Li E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat Rev Genet. 2002;3:662–73. doi: 10.1038/nrg887. [DOI] [PubMed] [Google Scholar]
- 35.La Salle S, Trasler JM. Dynamic expression of DNMT3a and DNMT3b isoforms during male germ cell development in the mouse. Dev Biol. 2006;296:71–82. doi: 10.1016/j.ydbio.2006.04.436. [DOI] [PubMed] [Google Scholar]
- 36.Ishii T, Kohu K, Yamada S, Ishidoya S, Kanto S, Fuji H, Moriya T, Satake M, Arai Y. Up-regulation of DNA-methyltransferase 3A expression is associated with hypomethylation of intron 25 in human testicular germ cell tumors. Tohoku J Exp Med. 2007;212:177–90. doi: 10.1620/tjem.212.177. [DOI] [PubMed] [Google Scholar]
- 37.Jeong S, Liang G, Sharma S, Lin JC, Choi SH, Han H, Yoo CB, Egger G, Yang AS, Jones PA. Selective anchoring of DNA methyltransferases 3A and 3B to nucleosomes containing methylated DNA. Mol Cell Biol. 2009;29:5366–76. doi: 10.1128/MCB.00484-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Chen T, Tsujimoto N, Li E. The PWWP domain of Dnmt3a and Dnmt3b is required for directing DNA methylation to the major satellite repeats at pericentric heterochromatin. Mol Cell Biol. 2004;24:9048–58. doi: 10.1128/MCB.24.20.9048-9058.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Renauld H. Heterochromatin: a meiotic matchmaker? Trends Cell Biol. 1997;7:201–5. doi: 10.1016/S0962-8924(97)01034-9. [DOI] [PubMed] [Google Scholar]
- 40.Jones PA. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet. 2012;13:484–92. doi: 10.1038/nrg3230. [DOI] [PubMed] [Google Scholar]
- 41.Chantalat S, Depaux A, Héry P, Barral S, Thuret JY, Dimitrov S, Gérard M. Histone H3 trimethylation at lysine 36 is associated with constitutive and facultative heterochromatin. Genome Res. 2011;21:1426–37. doi: 10.1101/gr.118091.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Dhayalan A, Rajavelu A, Rathert P, Tamas R, Jurkowska RZ, Ragozin S, Jeltsch A. The Dnmt3a PWWP domain reads histone 3 lysine 36 trimethylation and guides DNA methylation. J Biol Chem. 2010;285:26114–20. doi: 10.1074/jbc.M109.089433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Xu F, Mao C, Ding Y, Rui C, Wu L, Shi A, Zhang H, Zhang L, Xu Z. Molecular and enzymatic profiles of mammalian DNA methyltransferases: structures and targets for drugs. Curr Med Chem. 2010;17:4052–71. doi: 10.2174/092986710793205372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Suetake I, Mishima Y, Kimura H, Lee YH, Goto Y, Takeshima H, Ikegami T, Tajima S. Characterization of DNA-binding activity in the N-terminal domain of the DNA methyltransferase Dnmt3a. Biochem J. 2011;437:141–8. doi: 10.1042/BJ20110241. [DOI] [PubMed] [Google Scholar]
- 45.Choi SH, Heo K, Byun HM, An W, Lu W, Yang AS. Identification of preferential target sites for human DNA methyltransferases. Nucleic Acids Res. 2011;39:104–18. doi: 10.1093/nar/gkq774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Stec I, Nagl SB, van Ommen GJ, den Dunnen JT. The PWWP domain: a potential protein-protein interaction domain in nuclear proteins influencing differentiation? FEBS Lett. 2000;473:1–5. doi: 10.1016/S0014-5793(00)01449-6. [DOI] [PubMed] [Google Scholar]
- 47.Ge YZ, Pu MT, Gowher H, Wu HP, Ding JP, Jeltsch A, Xu GL. Chromatin targeting of de novo DNA methyltransferases by the PWWP domain. J Biol Chem. 2004;279:25447–54. doi: 10.1074/jbc.M312296200. [DOI] [PubMed] [Google Scholar]
- 48.Hahn MA, Wu X, Li AX, Hahn T, Pfeifer GP. Relationship between gene body DNA methylation and intragenic H3K9me3 and H3K36me3 chromatin marks. PLoS One. 2011;6:e18844. doi: 10.1371/journal.pone.0018844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Raddatz G, Gao Q, Bender S, Jaenisch R, Lyko F. Dnmt3a protects active chromosome domains against cancer-associated hypomethylation. PLoS Genet. 2012;8:e1003146. doi: 10.1371/journal.pgen.1003146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bienz M. The PHD finger, a nuclear protein-interaction domain. Trends Biochem Sci. 2006;31:35–40. doi: 10.1016/j.tibs.2005.11.001. [DOI] [PubMed] [Google Scholar]
- 51.Suzuki M, Yamada T, Kihara-Negishi F, Sakurai T, Hara E, Tenen DG, Hozumi N, Oikawa T. Site-specific DNA methylation by a complex of PU.1 and Dnmt3a/b. Oncogene. 2006;25:2477–88. doi: 10.1038/sj.onc.1209272. [DOI] [PubMed] [Google Scholar]
- 52.Hervouet E, Vallette FM, Cartron PF. Dnmt3/transcription factor interactions as crucial players in targeted DNA methylation. Epigenetics. 2009;4:487–99. doi: 10.4161/epi.4.7.9883. [DOI] [PubMed] [Google Scholar]
- 53.Wang YA, Kamarova Y, Shen KC, Jiang Z, Hahn MJ, Wang Y, Brooks SC. DNA methyltransferase-3a interacts with p53 and represses p53-mediated gene expression. Cancer Biol Ther. 2005;4:1138–43. doi: 10.4161/cbt.4.10.2073. [DOI] [PubMed] [Google Scholar]
- 54.Fuks F, Burgers WA, Godin N, Kasai M, Kouzarides T. Dnmt3a binds deacetylases and is recruited by a sequence-specific repressor to silence transcription. EMBO J. 2001;20:2536–44. doi: 10.1093/emboj/20.10.2536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Fuks F, Hurd PJ, Deplus R, Kouzarides T. The DNA methyltransferases associate with HP1 and the SUV39H1 histone methyltransferase. Nucleic Acids Res. 2003;31:2305–12. doi: 10.1093/nar/gkg332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rush M, Appanah R, Lee S, Lam LL, Goyal P, Lorincz MC. Targeting of EZH2 to a defined genomic site is sufficient for recruitment of Dnmt3a but not de novo DNA methylation. Epigenetics. 2009;4:404–14. doi: 10.4161/epi.4.6.9392. [DOI] [PubMed] [Google Scholar]
- 57.Zhang Y, Jurkowska R, Soeroes S, Rajavelu A, Dhayalan A, Bock I, Rathert P, Brandt O, Reinhardt R, Fischle W, et al. Chromatin methylation activity of Dnmt3a and Dnmt3a/3L is guided by interaction of the ADD domain with the histone H3 tail. Nucleic Acids Res. 2010;38:4246–53. doi: 10.1093/nar/gkq147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Otani J, Nankumo T, Arita K, Inamoto S, Ariyoshi M, Shirakawa M. Structural basis for recognition of H3K4 methylation status by the DNA methyltransferase 3A ATRX-DNMT3-DNMT3L domain. EMBO Rep. 2009;10:1235–41. doi: 10.1038/embor.2009.218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Gowher H, Jeltsch A. Molecular enzymology of the catalytic domains of the Dnmt3a and Dnmt3b DNA methyltransferases. J Biol Chem. 2002;277:20409–14. doi: 10.1074/jbc.M202148200. [DOI] [PubMed] [Google Scholar]
- 60.Hata K, Okano M, Lei H, Li E. Dnmt3L cooperates with the Dnmt3 family of de novo DNA methyltransferases to establish maternal imprints in mice. Development. 2002;129:1983–93. doi: 10.1242/dev.129.8.1983. [DOI] [PubMed] [Google Scholar]
- 61.Chedin F, Lieber MR, Hsieh CL. The DNA methyltransferase-like protein DNMT3L stimulates de novo methylation by Dnmt3a. Proc Natl Acad Sci U S A. 2002;99:16916–21. doi: 10.1073/pnas.262443999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Suetake I, Morimoto Y, Fuchikami T, Abe K, Tajima S. Stimulation effect of Dnmt3L on the DNA methylation activity of Dnmt3a2. J Biochem. 2006;140:553–9. doi: 10.1093/jb/mvj185. [DOI] [PubMed] [Google Scholar]
- 63.Aapola U, Kawasaki K, Scott HS, Ollila J, Vihinen M, Heino M, Shintani A, Kawasaki K, Minoshima S, Krohn K, et al. Isolation and initial characterization of a novel zinc finger gene, DNMT3L, on 21q22.3, related to the cytosine-5-methyltransferase 3 gene family. Genomics. 2000;65:293–8. doi: 10.1006/geno.2000.6168. [DOI] [PubMed] [Google Scholar]
- 64.Kareta MS, Botello ZM, Ennis JJ, Chou C, Chédin F. Reconstitution and mechanism of the stimulation of de novo methylation by human DNMT3L. J Biol Chem. 2006;281:25893–902. doi: 10.1074/jbc.M603140200. [DOI] [PubMed] [Google Scholar]
- 65.Holz-Schietinger C, Reich NO. The inherent processivity of the human de novo methyltransferase 3A (DNMT3A) is enhanced by DNMT3L. J Biol Chem. 2010;285:29091–100. doi: 10.1074/jbc.M110.142513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wienholz BL, Kareta MS, Moarefi AH, Gordon CA, Ginno PA, Chédin F. DNMT3L modulates significant and distinct flanking sequence preference for DNA methylation by DNMT3A and DNMT3B in vivo. PLoS Genet. 2010;6:e1001106. doi: 10.1371/journal.pgen.1001106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Jurkowska RZ, Rajavelu A, Anspach N, Urbanke C, Jankevicius G, Ragozin S, Nellen W, Jeltsch A. Oligomerization and binding of the Dnmt3a DNA methyltransferase to parallel DNA molecules: heterochromatic localization and role of Dnmt3L. J Biol Chem. 2011;286:24200–7. doi: 10.1074/jbc.M111.254987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Jia D, Jurkowska RZ, Zhang X, Jeltsch A, Cheng X. Structure of Dnmt3a bound to Dnmt3L suggests a model for de novo DNA methylation. Nature. 2007;449:248–51. doi: 10.1038/nature06146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Holz-Schietinger C, Matje DM, Harrison MF, Reich NO. Oligomerization of DNMT3A controls the mechanism of de novo DNA methylation. J Biol Chem. 2011;286:41479–88. doi: 10.1074/jbc.M111.284687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Eckhardt F, Lewin J, Cortese R, Rakyan VK, Attwood J, Burger M, Burton J, Cox TV, Davies R, Down TA, et al. DNA methylation profiling of human chromosomes 6, 20 and 22. Nat Genet. 2006;38:1378–85. doi: 10.1038/ng1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Smith ZD, Meissner A. DNA methylation: roles in mammalian development. Nat Rev Genet. 2013;14:204–20. doi: 10.1038/nrg3354. [DOI] [PubMed] [Google Scholar]
- 72.Sasaki H, Matsui Y. Epigenetic events in mammalian germ-cell development: reprogramming and beyond. Nat Rev Genet. 2008;9:129–40. doi: 10.1038/nrg2295. [DOI] [PubMed] [Google Scholar]
- 73.Sakai Y, Suetake I, Shinozaki F, Yamashina S, Tajima S. Co-expression of de novo DNA methyltransferases Dnmt3a2 and Dnmt3L in gonocytes of mouse embryos. Gene Expr Patterns. 2004;5:231–7. doi: 10.1016/j.modgep.2004.07.011. [DOI] [PubMed] [Google Scholar]
- 74.Kaneda M, Okano M, Hata K, Sado T, Tsujimoto N, Li E, Sasaki H. Essential role for de novo DNA methyltransferase Dnmt3a in paternal and maternal imprinting. Nature. 2004;429:900–3. doi: 10.1038/nature02633. [DOI] [PubMed] [Google Scholar]
- 75.Chen T, Ueda Y, Dodge JE, Wang Z, Li E. Establishment and maintenance of genomic methylation patterns in mouse embryonic stem cells by Dnmt3a and Dnmt3b. Mol Cell Biol. 2003;23:5594–605. doi: 10.1128/MCB.23.16.5594-5605.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Wu Z, Huang K, Yu J, Le T, Namihira M, Liu Y, Zhang J, Xue Z, Cheng L, Fan G. Dnmt3a regulates both proliferation and differentiation of mouse neural stem cells. J Neurosci Res. 2012;90:1883–91. doi: 10.1002/jnr.23077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Feng J, Chang H, Li E, Fan G. Dynamic expression of de novo DNA methyltransferases Dnmt3a and Dnmt3b in the central nervous system. J Neurosci Res. 2005;79:734–46. doi: 10.1002/jnr.20404. [DOI] [PubMed] [Google Scholar]
- 78.Nguyen S, Meletis K, Fu D, Jhaveri S, Jaenisch R. Ablation of de novo DNA methyltransferase Dnmt3a in the nervous system leads to neuromuscular defects and shortened lifespan. Dev Dyn. 2007;236:1663–76. doi: 10.1002/dvdy.21176. [DOI] [PubMed] [Google Scholar]
- 79.Wu H, Coskun V, Tao J, Xie W, Ge W, Yoshikawa K, Li E, Zhang Y, Sun YE. Dnmt3a-dependent nonpromoter DNA methylation facilitates transcription of neurogenic genes. Science. 2010;329:444–8. doi: 10.1126/science.1190485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Siegmund KD, Connor CM, Campan M, Long TI, Weisenberger DJ, Biniszkiewicz D, Jaenisch R, Laird PW, Akbarian S. DNA methylation in the human cerebral cortex is dynamically regulated throughout the life span and involves differentiated neurons. PLoS One. 2007;2:e895. doi: 10.1371/journal.pone.0000895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Day JJ, Sweatt JD. DNA methylation and memory formation. Nat Neurosci. 2010;13:1319–23. doi: 10.1038/nn.2666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Feng J, Zhou Y, Campbell SL, Le T, Li E, Sweatt JD, Silva AJ, Fan G. Dnmt1 and Dnmt3a maintain DNA methylation and regulate synaptic function in adult forebrain neurons. Nat Neurosci. 2010;13:423–30. doi: 10.1038/nn.2514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Tadokoro Y, Ema H, Okano M, Li E, Nakauchi H. De novo DNA methyltransferase is essential for self-renewal, but not for differentiation, in hematopoietic stem cells. J Exp Med. 2007;204:715–22. doi: 10.1084/jem.20060750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Li W, Chen BF. Aberrant DNA methylation in human cancers. J Huazhong Univ Sci Technolog Med Sci. 2013;33:798–804. doi: 10.1007/s11596-013-1201-0. [DOI] [PubMed] [Google Scholar]
- 85.Fernandez AF, Assenov Y, Martin-Subero JI, Balint B, Siebert R, Taniguchi H, Yamamoto H, Hidalgo M, Tan AC, Galm O, et al. A DNA methylation fingerprint of 1628 human samples. Genome Res. 2012;22:407–19. doi: 10.1101/gr.119867.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Esteller M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat Rev Genet. 2007;8:286–98. doi: 10.1038/nrg2005. [DOI] [PubMed] [Google Scholar]
- 87.Deng T, Kuang Y, Wang L, Li J, Wang Z, Fei J. An essential role for DNA methyltransferase 3a in melanoma tumorigenesis. Biochem Biophys Res Commun. 2009;387:611–6. doi: 10.1016/j.bbrc.2009.07.093. [DOI] [PubMed] [Google Scholar]
- 88.Mizuno S, Chijiwa T, Okamura T, Akashi K, Fukumaki Y, Niho Y, Sasaki H. Expression of DNA methyltransferases DNMT1, 3A, and 3B in normal hematopoiesis and in acute and chronic myelogenous leukemia. Blood. 2001;97:1172–9. doi: 10.1182/blood.V97.5.1172. [DOI] [PubMed] [Google Scholar]
- 89.Zhao Z, Wu Q, Cheng J, Qiu X, Zhang J, Fan H. Depletion of DNMT3A suppressed cell proliferation and restored PTEN in hepatocellular carcinoma cell. J Biomed Biotechnol. 2010;2010:737535. doi: 10.1155/2010/737535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Oh BK, Kim H, Park HJ, Shim YH, Choi J, Park C, Park YN. DNA methyltransferase expression and DNA methylation in human hepatocellular carcinoma and their clinicopathological correlation. Int J Mol Med. 2007;20:65–73. [PubMed] [Google Scholar]
- 91.Eklund EA. The role of HOX genes in malignant myeloid disease. Curr Opin Hematol. 2007;14:85–9. doi: 10.1097/MOH.0b013e32801684b6. [DOI] [PubMed] [Google Scholar]
- 92.Fabbri M, Garzon R, Cimmino A, Liu Z, Zanesi N, Callegari E, Liu S, Alder H, Costinean S, Fernandez-Cymering C, et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc Natl Acad Sci U S A. 2007;104:15805–10. doi: 10.1073/pnas.0707628104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Garzon R, Liu S, Fabbri M, Liu Z, Heaphy CE, Callegari E, Schwind S, Pang J, Yu J, Muthusamy N, et al. MicroRNA-29b induces global DNA hypomethylation and tumor suppressor gene reexpression in acute myeloid leukemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood. 2009;113:6411–8. doi: 10.1182/blood-2008-07-170589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Chen B-F, Gu S, Suen Y-K, Li L, Chan W-Y. microRNA-199a-3p, DNMT3A, and aberrant DNA methylation in testicular cancer. Epigenetics. 2013;9:1. doi: 10.4161/epi.25799. [DOI] [PMC free article] [PubMed] [Google Scholar]