

Abstract
Pancreatic cancer is a highly lethal tumor type for which there are few viable therapeutic options. It is also caused by the accumulation of mutations in a variety of genes. These genetic alterations can be grouped into those that accumulate during pancreatic intraepithelial neoplasia (precursor lesions) and thus are present in all cells of the infiltrating carcinoma, and those that accumulate specifically within the infiltrating carcinoma during subclonal evolution, resulting in genetic heterogeneity. Despite this heterogeneity there are nonetheless commonly altered cellular functions, such as pathways controlling the cell cycle, DNA damage repair, intracellular signaling and development, which could provide for a variety of drug targets. This review aims to summarize current knowledge of the genetics and genomics of pancreatic cancer from its inception to metastatic colonization, and to provide examples of how this information can be translated into the clinical setting for therapeutic benefit and personalized medicine.
The promise of genetics and genomics for targeted therapies of pancreatic cancer
Pancreatic ductal adenocarcinoma is a significant health concern worldwide. According to the International Agency for Research on Cancer (IARC), it is the 15th cancer in overall incidence in the world, with an estimated 277,000 new cases diagnosed per year [1]. It is one of the few diseases in which the mortality rate equals the incidence rate; as a result the five-year survival rate for this disease remains a dismal 5%, and this has remained constant over many years. In the US, pancreatic cancer mortality is projected to exceed that of breast cancer in the coming decade [2,3]. Pancreatic ductal adenocarcinoma is the most common form of cancer affecting the pancreas, and this is the form that we discuss here.
There are two predominant reasons why pancreatic cancer is so lethal. First, there are currently no screening methods for identifying it at stages when it could be cured [4]; it remains largely asymptomatic and thus undetected until it reaches an advanced stage, when surgical resection, the only potentially curative treatment, is not possible [5-7]. The search for sensitive and specific biomarkers of early stage disease is therefore of utmost importance [8]. Second, the chemotherapeutic options for treating it are limited. For many years the standard of care for patients with advanced stage disease has been gemcitabine, even though this drug confers only modest survival advantages on its own [5,7,9]. When used in combination with other agents gemcitabine has shown increased effectiveness; for example, the combination of gemcitabine with the epidermal growth factor receptor (EGFR) inhibitor erlotinib has been shown to provide a survival advantage in pancreatic cancer patients compared with gemcitabine alone, although the overall response rate is still low [10]. Gemcitabine in combination with agents to target desmoplastic (fibrosis-causing) stroma, such as Nab-paclitaxel (albumin-bound paclitaxel), has also shown promise, presumably because they deplete tumor stroma, which leads to better delivery of gemcitabine to the tumor cells [11]. Beyond the use of gemcitabine alone or in combination with other agents, preliminary success has been achieved with chemotherapeutic combination regimen FOLFIRINOX (folinic acid, fluorouracil, irinotecan, and oxaliplatin), although toxicities associated with these treatments limit their utility in many patients [12]. Finally, studies are also ongoing to investigate the effectiveness of Hedgehog inhibitors in pancreatic cancer. Inhibition of this pathway has been proposed to target both the tumor stroma and the cancer stem cell population, although success has not yet been achieved in the clinic [13,14]. Clearly, much progress remains to be made. A summary of chemotherapeutic strategies for pancreatic cancer is shown in Table 1.
Table 1.
Current and potential future chemotherapeutic options for pancreatic ductal adenocarcinoma
| Agent | Patients targeted | Median overall survival | References |
|---|---|---|---|
| Mitomycin Ca | Patients with mutations in PALB2 | - | [61] |
| Olapariba | Patients with mutations in BRCA2 | - | [68,69] |
| Gemcitabine alone | All | 5.65-7.2 monthsb | [9,90-94] |
| Gemcitabine + cisplatin | All | 7.5 months | [91] |
| Gemcitabine + erlotinib | All | 6.2 months | [10] |
| Gemcitabine + capecitabine | All | 7.1-8.4 monthsb | [93,94] |
| Gemcitabine + docetaxel + capecitabine (GTX)a | All | - | [95] |
| Gemcitabine + Nab-paclitaxel | All | 12.2 months | [11] |
| Folinic acid + fluorouracil + irinotecan + oxaliplatin (FOLFIRINOX) | All | 11.1 months | [90] |
aThese regimens have shown promise based on preliminary data in pancreatic cancer or in clinical trials in other cancer types.
bIf more than one trial has been reported the range of median overall survivals is listed.
In recent years, advances in sequencing technologies have enabled the genetic and genomic events that underlie pancreatic carcinogenesis and progression to be deciphered in great detail. These efforts have greatly advanced our understanding of the key molecular events and mechanisms - for example, the driver genes characteristic of this tumor type and the core signaling pathways to which they correspond. We now also understand the timing of occurrence of these genetic events in pancreatic carcinogenesis and progression, and the implications of this information for targeted therapies in the setting of personalized medicine. Herein, we summarize these discoveries and their potential for improved clinical management of pancreatic cancer.
Pancreatic cancer genetics and genomics
There are four genes that are mutated at high frequency in pancreatic cancer: KRAS, CDKN2A, TP53, and SMAD4 (Table 2); these are referred to as 'driver' genes. The most common of these are genetic aberrations in KRAS (v-Ki-ras2 Kirsten rat sarcoma viral oncogene homolog) at codons 12, 13, and occasionally 61 [15,16]. KRAS encodes a GTPase that activates downstream effectors of receptor tyrosine kinases, such as the mitogen-activated protein kinase (MAPK) cascades [17]. Mutations in KRAS result in its constitutive activation. Inactivation of CDKN2A (p16, cyclin-dependent kinase inhibitor 2A gene) is also a common event in pancreatic cancer, occurring by intragenic mutation in association with allelic loss, homozygous deletion, or hypermethylation [18,19]. CDKN2A encodes a cyclin-dependent kinase inhibitor that controls the G1-S transition of the cell cycle; loss of CDKN2A removes this important cellular brake mechanism [20]. Alterations in both KRAS and CDKN2A have been detected at the earliest stages, in pancreatic cancer precursor lesions (called pancreatic intraepithelial neoplasia or PanIN) [21]. However, mutations in KRAS are predicted to precede those of CDKN2A because of a higher prevalence of KRAS mutations in early stage precursor lesions and the observation that most PanIN lesions containing CDKN2A inactivation also harbor a KRAS mutation (Figure 1). TP53 (encoding the tumor protein p53), a master regulator of cell stress responses, is a frequent mutational target in many solid tumors [22-24], and pancreatic cancer is no exception; mutations in TP53 occur in up to 75% of pancreatic cancers, most often by point mutation or small intragenic deletion [23,24]. Finally, SMAD4 (DPC4, SMAD family member 4 gene), encoding a transcription factor that mediates transforming growth factor-β (TGF-β) and bone morphogenetic protein (BMP) signaling, is affected by homozygous deletion or inactivating mutations and allelic loss in up to 55% of pancreatic cancer patients [25,26]. Unlike KRAS and CDKN2A, the TP53 and SMAD4 genes are mutated in late stage PanINs, typically PanIN-3 lesions [27,28] (Figure 1).
Table 2.
Sporadic and inherited genetic alterations in pancreatic ductal adenocarcinoma
| Genes | Full name | Known function(s) | Location | Effect‡ | Prevalence | S/I§ | Other cancers | Inherited syndrome | Est. rel. risk (if known) | References |
|---|---|---|---|---|---|---|---|---|---|---|
| None | - | None | None | 1 | ||||||
| KRAS | v-Ki-ras2 Kirsten rat sarcoma viral oncogene homolog | GDP/GTP binding proteins, proliferation, survival, others | 12p12.1 | Act | 90-95% | S | Bladder, breast, leukemia, and lung | Cardiofaciocutaneous syndrome | [15,16,96] | |
| CDKN2A (p16) | Cyclin-dependent kinase inhibitor 2A | Cell cycle | 9p21.3 | In | 90% | S/I | Melanoma | FAMMM | 20-34X¶ | [97,98] |
| TP53 | Tumor protein p53 | Cell cycle, apoptosis, DNA repair, others | 17p13.1 | In | 75% | S | Breast, colorectal, hepatocellular, others | Li-Fraumeni syndrome | [99] | |
| SMAD4/DPC4 | SMAD family member 4 | TGF-β signaling, BMP signaling, development, proliferation, others | 18q21.2 | In | 55% | S | Colorectal | Juvenile polyposis syndrome, Myhre syndrome | [26-28,100,101] | |
| TGFBR1,2 | Transforming growth factor, beta receptor 1, 2 | TGF-β signaling, development, proliferation, others | 9q22.33, 3p24.1 | In | 5-10% | S | Colorectal, esophageal | Loeys-Dietz syndrome | [29,102] | |
| ACVR1B | Activin A receptor, type IB | TGF-β signaling, development, proliferation, others | 12q13.13 | In | 5-10% | S | Unknown | Unknown | [30] | |
| MKK4 | Mitogen-activated protein kinase kinase 4 | Cell stress, JNK signaling | 17p12 | In | 5-10% | S | Unknown | Unknown | [103] | |
| MLL3 | Myeloid/lymphoid or mixed-lineage leukemia 3 | Chromatin remodeling, transcription | 7q36.1 | In | <10% | S | Breast, colorectal, and leukemia | [16,104] | ||
| ARID1A/B | AT-rich interaction domain-containing protein 1A/B | Chromatin remodeling, transcription | 1p36.11, 6q25.3 | In | <10% | S | Breast, ovary and liver | Coffin-Siris syndrome | [16,105,106] | |
| PBRM1 | Polybromo 1 | Chromatin remodeling, transcription | 3p21.1 | In | <10% | S | Kidney | [16,106] | ||
| SMARCA4 | Swi/Snf-related, matrix associated, actin-dependent regulator of chromatin, subfamily A, member 4 | Chromatin remodeling, transcription | 19p13.2 | In | <10% | S | Lung | Coffin-Siris syndrome, rhaboid tumor predisposition syndrome | [16,105-107] | |
| BRCA1, BRCA2 | Breast cancer 1, early onset, breast cancer 2, early onset | DNA repair, cell cycle, genome stability | 17q21.31, 13q13.1 | In | <10% | I | Breast, ovary and prostate | Breast cancer | 3.5-10X, 2X | [34,108,109] |
| PALB2, ATM, others | Partner and localizer of BRCA2, Ataxia telangiectasia mutated | DNA repair, cell cycle, genome stability | 7q34 | In | <10% | I | Unknown | Familial pancreatic cancer¥ | 32X (overall) | [31,32,35,39,40] |
| PRSS1 | Protease, serine, 1 (trypsin 1) | Cell metabolism and signaling | 19p13.3 | In | <10% | I | None | Familial pancreatitis | 50-80X | [110] |
| STK11/LKB1 | Serine/threonine kinase 11 | p53 signaling, DNA repair, apoptosis | 3p22.2, 2p21 | In | <10% | I | Breast, colorectal, gastroesophageal, and small bowel | Peutz-Jeghers syndrome | 132X | [33,111] |
| hMLH1, hMSH2, others | MutL Homolog 1, colon cancer, nonpolyposis type 2 (I), mutS homolog 2, colon cancer, nonpolyposis type 1 (I) | DNA mismatch repair | 9q22.32, 9p13.3 | In | <10% | I | Biliary tract, brain, colorectal, endometrial, ovarian, stomach, ureter and renal pelvis | HNPCC | [112,113] | |
| FANC-C, FANC-G | Fanconi anemia, complementation group C, Fanconi anemia, complementation group G | DNA stability and repair | 9q22, 9p13 | In | <10% | I | Unknown | Young-age-onset pancreatic cancer | [36-38] |
‡Effect of mutation: Act, activating; In, inactivating. §Commonly sporadic (S) or inherited (I). ¶This relative risk applies only to patients with germline CDKN2A mutations. ¥Familial pancreatic cancer is defined by three first degree relatives in these studies.
Figure 1.
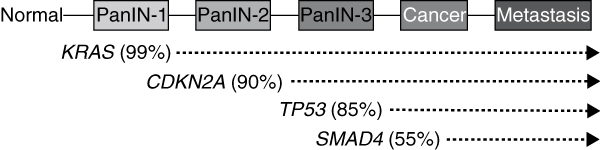
Progression model of pancreatic carcinogenesis. Pancreatic intraepithelial neoplasia (PanIN) is an established precursor lesion of infiltrating pancreatic ductal adenocarcinoma that involves the normal ductal epithelium of the pancreas. PanIN lesions develop from normal acinar cells in the pancreas, probably as the result of an activating KRAS mutation, leading to the formation of a PanIN-1 lesion characterized by a tall columnar epithelium lining the duct system but with little nuclear atypia. The development of inactivating mutations in CDKN2A coincides with the progression of a PanIN-1 to a PanIN-2 lesion, characterized by loss of polarity, pseudostratification, papillary formations, and nuclear atypia. Inactivating mutations of TP53 and SMAD4 are late events and most often detected in PanIN-3 stage lesions. PanIN-3 lesions are recognized by their complete lack of polarity, marked nuclear atypia, high nuclear/cytoplasmic ratio, and pseudopapillary formation. Mutations in additional genes may also occur during PanIN formation that are not illustrated in this example.
A variety of genes are also mutated at low frequency in sporadic pancreatic cancer, such as TGFBR1, TGFBR2, and ACVR1B, which encode ligand receptors in the TGF-β/activin signaling pathway, and the protein kinase MKK4 [29,30]. Additional low-frequency targets are germline variants associated with the familial aggregation of pancreatic cancer [31,32]. For example, germline mutations in the liver kinase B1 gene (LKB1) are associated with the development of hamartomatous polyps in association with Peutz-Jeghers syndrome, and patients with this syndrome have a >100-fold increased risk of developing pancreatic cancer in the context of intraductal papillary mucinous neoplasms [32]. However, LKB1 may also be inactivated in sporadic pancreatic cancer [33]. Inherited mutations in the BRCA2 gene (encoding breast cancer type 2 susceptibility protein, which is involved in DNA damage repair) are perhaps the best characterized of the germline variants [34,35]. In addition to the increased risk of developing breast or ovarian cancer, BRCA2 mutations are associated with a 3.5- to 10-fold increased risk of developing pancreatic cancer. Following the identification of BRCA2 mutations in familial pancreatic cancer, germline mutations in the Fanconi anemia genes FANCC, FANCG, and PALB2 (FANCN), whose protein product interacts with that of BRCA2, have also been implicated in familial pancreatic cancer [36-39]. Most recently, germline mutations in ATM (encoding the protein kinase Ataxia telangiectasia mutated) have been described in subsets of familial pancreatic cancer families [40]. Table 2 summarizes the somatic and germline alterations in known pancreatic cancer genes, their functions, and the relative risks that they confer.
Although the hallmark genetic changes contributing to pancreatic cancer have been well established, only recently has the pancreatic cancer genome been analyzed on a larger scale by whole-exome sequencing. In analyzing the exomes of 17 primary tumors and 7 metastases, Jones et al. [16] reaffirmed the known common genetic alterations in pancreatic cancer and revealed previously unrecognized alterations in genes that have a role in chromatin remodeling (ARID1A and MLL3). Moreover, although they demonstrated that there are several core pathways that are recurrently targeted in most pancreatic cancers, such as those that control cell division, cell death, adhesion, and various signaling pathways [16], the pathway components that were altered in any individual tumor varied widely (Figure 2). Most recently, Biankin et al. [41] performed whole-exome sequencing in combination with copy number analysis of 99 early stage (clinical stage I and II) infiltrating pancreatic cancers that yielded sequencing data with a high depth of coverage. A pathway-based analysis of this new comprehensive set of mutations confirmed the pancreatic cancer core pathways previously described by Jones et al. [16]. In addition, because of the larger sample size, Biankin et al. [41] could identify novel genetic targets in each core signaling pathway and also many alterations in genes encoding axon guidance factors that are typically expressed during embryogenesis [42]. The relevance of the human genomic data accrued by Biankin et al. [41] was also confirmed in a murine pancreatic cancer model based on transposon-mediated mutagenesis [43].
Figure 2.
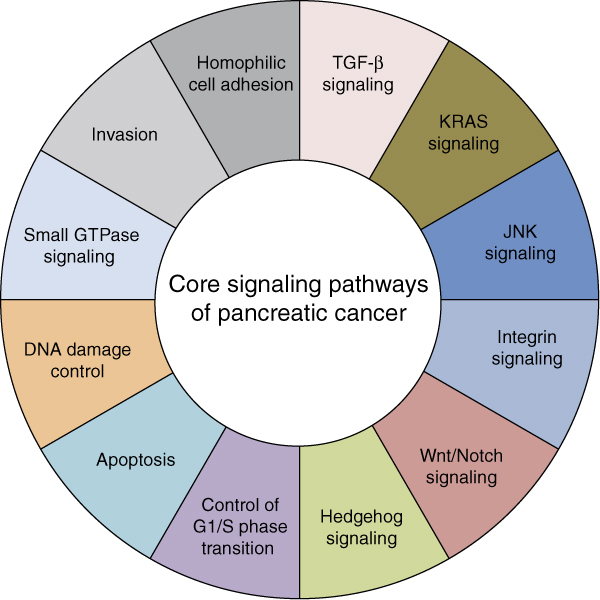
Core signaling pathways in pancreatic cancer. The pathways and processes whose component genes are genetically altered in most pancreatic cancers based on whole-exome sequencing are shown. Although some genes may correspond to a single pathway (for example, KRAS2 mutations and the KRAS signaling core pathway) others may have a role in more than one pathway (for example, TP53 mutations and the apoptosis, DNA damage, and JNK core signaling pathways). Therapeutic targeting of one or more of these pathways, rather than specific gene alterations that occur within a pathway, provides a new way of treating pancreatic cancer. TGF-β, transforming growth factor β.
The clinical significance of these findings is that there is extensive genetic heterogeneity among different patients' pancreatic cancers, partly explaining why many gene-based therapies will prove ineffective at targeting a genetic alteration that are present in only a small subset of carcinomas. By contrast, therapies that target core pathways, rather than the diverse genetic alterations that can occur within that pathway, may prove more effective, as such therapies would focus on the convergent phenotypes and not the diverse genotypes observed. Despite these sobering implications for therapy, the genetic heterogeneity observed in pancreatic cancer provides important additional information: the diverse somatic alterations can be used as evolutionary markers to reveal the life history of pancreatic cancer. Such studies are essential for understanding the contribution of genetic mutations to pancreatic carcinogenesis, progression, and metastasis.
The role of genetics and genomics in subclonal evolution
It has been over 150 years since Charles Darwin first described natural selection as a force in evolutionary change [44]. In 1976, Peter Nowell implicated evolutionary change in cancer when he hypothesized that variant subclones undergo stepwise selection in tumor progression [45]. We now consider cancer to be a genetic disease in which mutations accumulate over time leading to the eventual acquisition of advantageous 'hallmarks' or traits [46,47]. Moreover, large scale sequencing studies have revealed the various intragenic alterations, copy number variants and chromosomal rearrangements that characterize the many distinct types of cancer [48]. Although Darwin and Nowell provided the overall framework, the current challenge is to translate these concepts of genetic change and clonal evolution in cancer to our ability to diagnose and treat this disease.
Metastasis is a key feature of many aggressive cancers, including pancreatic cancer, and is caused by a variety of factors such as changes in gene expression of the tumor cell, the microenvironment, and angiogenesis [49,50]. However, although the genetic events associated with carcinogenesis are well delineated, the relevance of genetic events to these steps of tumor progression is unknown. To address this lack of knowledge for pancreatic cancer, Yachida et al. [51] reported the combinatorial effects of the four most commonly mutated genes in pancreatic cancer (KRAS, CDKN2A, TP53, and SMAD4) on patient outcome, with the hypothesis that the combination of somatic alterations in these four genes may be the predominant biological features of that neoplasm. The results supported this hypothesis. Patients whose carcinomas had at least three of these driver genes mutated showed a worse prognosis than those patients who had only one or two of these genes mutated in their carcinoma; patients whose carcinomas harbored at least three mutated genes also had high metastatic burden at autopsy. The authors [51] next evaluated the relationship of each gene specifically to patient outcome. Although no relationships were found for KRAS or CDKN2A, they noted that widely metastatic pancreatic cancer (characterized by hundreds to thousands of metastases present at rapid autopsy) typically arose from carcinomas with TP53 or SMAD4 mutations. Moreover, there were non-random patterns in which genetic alterations in TP53 and SMAD4 coexisted. For example, SMAD4 loss of function almost always occurred in association with genetic inactivation of TP53, whereas the converse was not true. This indicates that SMAD4 alterations are selected for in association with TP53 genetic alterations during PanIN progression. This also suggests that SMAD4 inactivation occurs later than TP53 inactivation in the genetic progression model of pancreatic carcinogenesis (Figure 1). The types of TP53 alterations in pancreatic cancer were also informative. SMAD4 loss most commonly occurred in association with inactivating TP53 missense mutations [51]. In contrast, TP53 inactivation in association with wild-type SMAD4 was highly enriched for nonsense, deletion, or frameshift mutations that abolish p53's DNA binding activity, suggesting that loss of SMAD4 during PanIN progression is selected for because of its cytostatic and apoptotic functions conferred by its own ability to bind DNA in association with SMAD2 or SMAD3 [51,52]. Collectively, these findings indicate that the genetic features of a primary carcinoma that accumulate during carcinogenesis strongly influence its metastatic propensity. Thus, the genetic features of a carcinoma that can be exploited for the purposes of early detection, for example TP53, might also provide information about the metastatic potential of that carcinoma.
The genetic alterations that underlie pancreatic cancer metastasis have also been explored on a more global level [53,54]. Yachida et al. [53] used whole-exome sequencing data of seven metastases to explore the timing and dynamics of genetic events in metastatic dissemination of pancreatic cancer. This approach revealed that mutations in a carcinoma could be classified into founders and progressors (Figure 3). Founder mutations are those that are present in all samples analyzed for a patient; from an evolutionary perspective, founder mutations are those clonal events that characterize the 'most recent common ancestor' of all cells in the neoplasm, or the parental clone that gave rise to it. These mutations were acquired during PanIN progression culminating in formation of the parental clone. By contrast, progressor mutations were those that occurred in only a subset of the samples analyzed for a patient. These sets of alterations were acquired later than founder mutations and thus highlight subclonal lineages that arose from the parental clone after the infiltrating carcinoma formed. Importantly, Campbell and colleagues [54] observed a similar pattern while exploring genomic instability and rearrangements in a set of 13 pancreatic cancer patients. Many genomic rearrangements were shared among all samples analyzed for a given patient, yet subsets of rearrangements were found in only a subset of samples or uniquely to a single metastasis. This study [54] also described several phylogenetic relationships between the primary and metastatic neoplasms within a patient and also evidence of organ-specific signatures indicating selection for and adaptation of subclones in the new environment.
Figure 3.
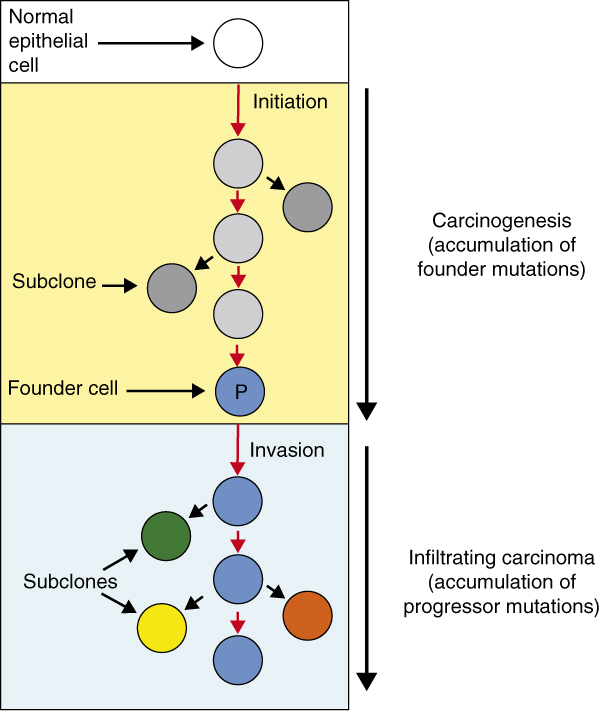
Model of pancreatic carcinogenesis and progression based on clonal evolution studies. Carcinogenesis begins with an initiating alteration in a normal epithelial cell progenitor that provides a selective advantage. Over time, waves of clonal expansion take place in association with the acquisition of mutations in genes such as CDKN2A, TP53, or SMAD4, corresponding to the genetic progression model of pancreatic intraepithelial neoplasia (PanIN). This clonal expansion is expected to generate more than one subclone within a PanIN, one of which will give rise to the founder cell that will eventually become the parental clone (P) of cells that initiate the infiltrating carcinoma. The time taken for a cell with an initiating alteration to accumulate all mutations eventually present in the founder cell that forms the parental clone of the neoplasm is estimated to be at least 12 years [45]. Additional waves of clonal expansion and accumulation of mutations continue to occur in cell lineages derived from the parental clone leading to the formation of numerous subclones and a genetically heterogeneous primary carcinoma.
There are at least two conclusions that can be made from genomic studies of pancreatic cancer progression. First, the time required for accumulation of all genetic alterations in a parental clone suggests a relatively long interval for potentially curative screening methods in this disease, in the order of a decade or longer [53] (Figure 3). Second, therapies that target subclonal genetic alterations (progressor events) will eventually lead to tumor recurrence, as subclones without those alterations will in theory be selected for by such therapies [55,56]. Therefore, successful approaches will ideally target genetic alterations in the parental clone of that pancreatic cancer (founder events) [53,54,57]. A recent study by Haeno and colleagues [58] using computational modeling further supports this by indicating that pancreatic cancer is commonly in an exponential growth phase at diagnosis. Thus, although it may take many years for a tumor to develop, very little time is needed for the cancer to have significant consequences and aggressive behavior once it is clinically evident. This exponential growth phase probably occurs after the formation of the parental clone, as subclones are continuously created and selected for and as they adapt to the dynamic microenvironment. Only by identifying and targeting the alterations representing the parental clone might we be able to cure this disease by targeted therapies. Approaches for targeting the tumor stroma or stem cell populations of pancreatic cancer also remain viable targets, as evidenced by prolonged survival in patients treated with agents that target them [11,13,14].
Using genomic information to guide treatment of pancreatic cancer
The ultimate goal of deciphering the genomic changes that occur in pancreatic cancer is to use the information gleaned from each individual patient to guide their treatment. Research in this area is still in progress, but there have been advances in both pancreatic and other cancer models that suggest promising directions.
Some success has been achieved in familial pancreatic cancer patients. As discussed earlier, subsets of familial cases of pancreatic cancer arise through inherited mutations in BRCA2, FANCC, FANCG, and PALB2 [34,36-39]. Cancers deficient in these genes are missing a key component of the double-strand break repair pathways necessary for error-free DNA repair, and treatment of cell lines or xenografts harboring defects in these genes with DNA crosslinking agents has proven a potent therapeutic option by exploiting these deficiencies [59-62]. It was also hypothesized that targeting an alternative DNA repair pathway - such as base-excision repair, in which PARP-1 (poly (ADP-ribose) polymerase 1) is a central player [63] - would lead to the accumulation of enough DNA damage to result in growth arrest or apoptosis of tumor cells. This was first shown to be true in cell lines (embryonic stem cells, Chinese hamster ovary cells, and MCF7 and MDA-MB-231 cell lines) that were isogenic for loss of either BRCA1 or BRCA2 [64-66]. A subsequent experiment revealed high efficacy of PARP inhibition in Capan-1 pancreatic cancer cells, which have a somatic mutation in BRCA2 [67]. Although these data are preliminary in pancreatic cancer so far, it is exciting to consider PARP inhibitors as treatment options for patients with germline or somatic BRCA2 or related pathway member mutations. Currently, olaparib, a PARP inhibitor, is undergoing clinical trials in breast and ovarian cancers for patients with BRCA1/2 mutations, with 40% response rates reported [68,69]. It remains to be seen whether PARP inhibition will also be a viable option for treating pancreatic cancer patients with germline or somatic mutations in these genes, and clinical trials to explore this possibility are currently underway [70]. However, because patients with mutations in familial pancreatic cancer genes, such as those described earlier, are relatively uncommon among patients newly diagnosed with this disease, agents that target pancreatic cancer based on more common somatic driver mutations outlined in Table 2 would be applicable to a greater proportion of patients.
One strategy for genomics-based therapeutics has been to induce synthetic lethality in cancer cells, in which certain cellular events, if present simultaneously, result in the death of the cell. In cancer, the goal is to discover agents that, in the presence of specific mutations, induce this phenomenon [71]. If possible, it seems that the ultimate target for pancreatic cancer treatment would be the KRAS protein itself, given that it is oncogenic, it is nearly ubiquitously mutated in pancreatic cancer, and it is a mutation found in the parental clone (and thus all cells) of the cancer [16,41,53]. However, targeting it has been an insurmountable hurdle so far. KRAS itself does not appear to be druggable, and a recent screen for synthetic lethality with KRAS activation proved ineffective in pancreatic cancer cell lines [72,73]. Nevertheless, in other model systems there have been genes identified whose inhibition is synthetically lethal with mutant KRAS, including TBK1 (involved in activation of NF-κB), STK33 (a serine/threonine kinase that activates S6K1), PLK1 (a serine/threonine kinase involved in mitosis), and members of the APC/C complex (anaphase-promoting complex) [74-76]. A proposed alternative would be a synthetic lethal screen of cells with CDKN2A inactivation, as this gene is also altered during carcinogenesis in the majority of pancreatic cancers, and is thus also harbored in the parental clone of the cancer [16,41,53].
Although limited success has been achieved in targeting KRAS directly, recent studies suggest that targeting its downstream effectors may prove more effective. A recent report from Collisson et al. [77] reported success in treating a model of pancreatic cancer with a combination of pathway inhibitors. Inhibition of the MEK-ERK MAPK signaling pathway was effective in pancreatic cancer cell lines and orthotopic tumors, but activation of AKT (protein kinase B) signaling was induced by this treatment. However, when the authors [77] treated cell lines with a combination of MEK and AKT inhibitors, a synergistic effect in killing the tumor cells was observed. Given that both of these pathways are activated by oncogenic KRAS, it is intriguing to consider targeting the downstream effectors of KRAS signaling as a bypass to the inability to target mutant KRAS itself. The concept of targeting downstream KRAS signaling is not a new one. Diep and colleagues [78] found synergistic effects of simultaneous EGFR and MEK inhibition in pancreatic cancer. This was especially evident in cells with wild-type KRAS, but not in those with KRAS mutations. However, certain cell lines with KRAS mutations were sensitive to MEK inhibition alone [78]. These data suggest that downstream inhibition of KRAS may be a promising option for pancreatic cancer patients.
Preliminary studies targeting somatic mutations in pancreatic cancer have shown some promise [79,80]. Synthetic lethality screens in cell lines isogenic for SMAD4 expression identified two novel compounds, UA62001 and UA62784, which selectively targeted SMAD4-negative cells [81,82]. Of particular interest, the effectiveness of UA62001 was shown to correlate well with overall TGF-β pathway mutational status in a panel of 11 commonly available pancreatic cancer cell lines [82]. More recently, Cui et al. [79] expanded the synthetic lethality concept by examining the efficacies of broadly acting drugs on 18 cell lines studied by Jones et al. [16] plus isogenic cell lines. Cui and colleagues [79] were able to identify a decreased sensitivity of cells with CDKN2A mutations to gemcitabine and mitomycin C, an increased sensitivity of cells with TP53 mutations to triptolide, a decreased sensitivity of cells with SMAD4 mutations to gemcitabine, and an increased sensitivity of cells with SMAD4 mutations to irinotecan and cisplatin. Although the differences in half maximal inhibitory concentration that were observed with these drugs were significant across these genotypes, the fold changes themselves were relatively small.
In an alternative approach demonstrating the utility of unbiased molecular profiling for identifying therapeutic vulnerabilities, von Hoff et al. [80] used immunohistochemistry, fluorescent in situ hybridization, and gene expression microarray analyses of a large series of refractory patient tumors, including pancreatic cancer, to identify differentially expressed genes that interact with a known therapeutic agent. Of 86 patients for which molecular profiling was performed, a molecular target was identified in 84 (98%) patients, 66 of whom were treated according to the results of the molecular profile. Of these 66 patients, 27% had a progression-free survival that was longer than expected from their times to disease progression while on previous failed regimens. This supports the use of therapies targeting specific molecular profiles identified in a patient's tumor tissues. More work will be required to validate these types of studies, but they represent important first steps in identifying genetic-based susceptibilities to therapies used in patients with pancreatic cancer.
Concluding remarks and future directions
The complexity of pancreatic cancer from its inception to metastatic colonization is now firmly established, leading to new insights into options for therapeutic targeting based on the genetic features of the neoplasm. For example, it is now evident that the genetic heterogeneity of a pancreatic cancer can be combined into a small number of pathways whose downstream effects can be targeted [16,41]. Moreover, genetic alterations in cancer can also be categorized by the timing of their development, and those that occur during carcinogenesis may be better targets for therapeutic development as they are contained within every cell of the neoplasm. Related to this, patients with inherited germline mutations in BRCA2 and related genes now have options for chemotherapeutic management that exploit their cancers' defects in DNA damage repair, for example mitomycin C or PARP inhibitors [61,68,69].
The elucidation of the pancreatic cancer genome has implications beyond that of treatment, such as for risk assessment or early detection. For example, sequencing the germline of any individual with a strong family history of pancreatic cancer could identify those with a genetic predisposition to the disease, thus prioritizing them for careful screening of their pancreas or other at-risk organs [8]. Moreover, given that most patients do not have a family history of pancreatic cancer, the development of technologies and biomarkers to detect pancreatic cancer while still in the curative stage is of utmost importance. Such a goal is in sight. For example, up to 3% of individuals have a pancreatic cyst that is detectable by computerized tomography [83], some of which are precursors to pancreatic cancer [84]. The distinction of precancerous cysts from those that do not require clinical intervention remains a challenge, but recent studies indicate this could be done simply by sequencing endoscopically obtained cyst fluid [85], thereby identifying patients who can be potentially cured by surgical management.
Although there is potential for the use of genetic and genomic information to guide pancreatic cancer diagnosis and treatment, how this will be done on a population level remains to be seen. For example, there are still no reliably sensitive or specific markers to diagnose pancreatic cancer in the curative stage [86,87]. However, considering exciting new data showing the ability to detect cancer DNA in circulating blood- or cell-based assays of patients with colorectal, breast, and gynecological malignancies [88,89], similar strategies could probably be applied to patients with pancreatic cancer. Such approaches may also help in identifying therapeutic targets. For example, in theory sequencing of circulating genomic DNA may be better than sequencing a single sample of a neoplasm, as the latter does not indicate the extent of its genetic heterogeneity following subclonal evolution. However, the extent to which circulating DNA or shed cells reflect the heterogeneous nature of a pancreatic cancer remains to be determined. Nonetheless, genome sequencing of pancreatic cancers and their metastases has provided invaluable insight into the biological features of this disease, and in the future it will no doubt help in identifying potential therapies.
Abbreviations
BRCA2: breast cancer 2; CDKN2A: cyclin-dependent kinase inhibitor 2A; FOLFIRINOX: folinic acid, fluorouracil, irinotecan, and oxaliplatin; KRAS: v-Ki-ras2 Kirsten rat sarcoma viral oncogene homolog; MAPK: mitogen-activated protein kinase; MEK: MAPK kinase; PanIN: pancreatic intraepithelial neoplasia; PARP: poly (ADP-ribose) polymerase; SMAD4: SMAD family member 4; TP53: tumor protein p53; TGF-β: transforming growth factor β.
Competing interests
The authors declare that they have no competing interests.
Contributor Information
Alvin Makohon-Moore, Email: amakoho1@jhmi.edu.
Jacqueline A Brosnan, Email: jbrosna1@jhmi.edu.
Christine A Iacobuzio-Donahue, Email: ciacobu@jhmi.edu.
Acknowledgements
This work was supported by NIH/NCI grants 140599, CA101955, CA62624 and CA121113, The Skip Viragh Pancreatic Cancer Center and The Sol Goldman Pancreatic Cancer Research Center.
References
- Ferlay J, Shin H-R, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;5:2893–2917. doi: 10.1002/ijc.25516. [DOI] [PubMed] [Google Scholar]
- Siegel R, Naishadham D, Jemal A. Cancer statistics, 2012. CA Cancer J Clin. 2012;5:10–29. doi: 10.3322/caac.20138. [DOI] [PubMed] [Google Scholar]
- Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011: the impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA Cancer J Clin. 2011;5:212–236. doi: 10.3322/caac.20121. [DOI] [PubMed] [Google Scholar]
- Goggins M. Markers of pancreatic cancer: working toward early detection. Clin Cancer Res. 2011;5:635–637. doi: 10.1158/1078-0432.CCR-10-3074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verslype C, Van Cutsem E, Dicato M, Cascinu S, Cunningham D, Diaz-Rubio E, Glimelius B, Haller D, Haustermans K, Heinemann V, Hoff P, Johnston PG, Kerr D, Labianca R, Louvet C, Minsky B, Moore M, Nordlinger B, Pedrazzoli S, Roth A, Rothenberg M, Rougier P, Schmoll H-J, Tabernero J, Tempero M, Van de Velde C, Van Laethem J-L, Zalcberg J. The management of pancreatic cancer. Current expert opinion and recommendations derived from the 8th World Congress on Gastrointestinal Cancer, Barcelona, 2006. Ann Oncol. 2007;5(Suppl 7):vii1–vii10. doi: 10.1093/annonc/mdm210. [DOI] [PubMed] [Google Scholar]
- Sohn TA, Yeo CJ, Cameron JL, Koniaris L, Kaushal S, Abrams RA, Sauter PIC, Coleman J, Hruban RH, Lillemoe KD. Resected adenocarcinoma of the pancreas - 616 patients: results, outcomes, and prognostic indicators. J Gastrointest Surg. 2000;5:567–579. doi: 10.1016/S1091-255X(00)80105-5. [DOI] [PubMed] [Google Scholar]
- Hidalgo M. Pancreatic cancer. N Engl J Med. 2010;5:1605–1617. doi: 10.1056/NEJMra0901557. [DOI] [PubMed] [Google Scholar]
- Bhutani MS, Thosani N, Suzuki R, Guha S. Pancreatic cancer screening: what we do and do not know. Clin Gastroenterol Hepatol. 2013. doi: 10.1016/j.cgh.2013.02.004. [DOI] [PubMed]
- Burris HA, Moore MJ, Andersen J, Green MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo A, Tarassoff P, Nelson R, Dorr FA, Stephens CD, Von Hoff DD. Improvements in survival and clinical benefit with gemcitabine as first-line therapy for patients with advanced pancreas cancer: a randomized trial. J Clin Oncol. 1997;5:2403–2413. doi: 10.1200/JCO.1997.15.6.2403. [DOI] [PubMed] [Google Scholar]
- Moore MJ, Goldstein D, Hamm J, Figer A, Hecht JR, Gallinger S, Au HJ, Murawa P, Walde D, Wolff RA, Campos D, Lim R, Ding K, Clark G, Voskoglou-Nomikos T, Ptasynski M, Parulekar W. Erlotinib plus gemcitabine compared with gemcitabine alone in patients with advanced pancreatic cancer: a phase III trial of the National Cancer Institute of Canada Clinical Trials Group. J Clin Oncol. 2007;5:1960–1966. doi: 10.1200/JCO.2006.07.9525. [DOI] [PubMed] [Google Scholar]
- Von Hoff DD, Ramanathan RK, Borad MJ, Laheru DA, Smith LS, Wood TE, Korn RL, Desai N, Trieu V, Iglesias JL, Zhang H, Soon-Shiong P, Shi T, Rajeshkumar NV, Maitra A, Hidalgo M. Gemcitabine plus nab-paclitaxel is an active regimen in patients with advanced pancreatic cancer: a phase I/II trial. J Clin Oncol. 2011;5:4548–4554. doi: 10.1200/JCO.2011.36.5742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conroy T, Desseigne F, Ychou M, Bouche O, Guimbaud R, Becouarn Y, Adenis A, Raoul J-L, Gourgou-Bourgade S, De la Douchardiere C, Bennouna J, Bachet J-B, Khemissa-Akouz F, Pere-Verge D, Delbaldo C, Assenat E, Chauggert B, Michel P, Montoto-Grillot C, Ducreux M. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N Engl J Med. 2011;5:768–769. doi: 10.1056/NEJMoa1011923. author reply 769. [DOI] [PubMed] [Google Scholar]
- Olive KP, Jacobetz MA, Davidson CJ, Gopinathan A, McIntyre D, Honess D, Madhu B, Goldgraben MA, Caldwell ME, Allard D, Frese KK, Denicola G, Feig C, Combs C, Winter SP, Ireland-Zecchini H, Reichelt S, Howat WJ, Chang A, Dhara M, Wang L, Rückert F, Grützmann R, Pilarsky C, Izeradjene K, Hingorani SR, Huang P, Davies SE, Plunkett W, Egorin M. et al. Inhibition of Hedgehog signaling enhances delivery of chemotherapy in a mouse model of pancreatic cancer. Science. 2009;5:1457–1461. doi: 10.1126/science.1171362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penchev VR, Rasheed ZA, Maitra A, Matsui W. Heterogeneity and targeting of pancreatic cancer stem cells. Clin Cancer Res. 2012;5:4277–4284. doi: 10.1158/1078-0432.CCR-11-3112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almoguera C, Shibata D, Forrester K, Martin J, Arnheim N, Perucho M. Most human carcinomas of the exocrine pancreas contain mutant c-K-ras genes. Cell. 1988;5:549–554. doi: 10.1016/0092-8674(88)90571-5. [DOI] [PubMed] [Google Scholar]
- Jones S, Zhang X, Parsons DW, Lin JC-H, Leary RJ, Angenendt P, Mankoo P, Carter H, Kamiyama H, Jimeno A, Hong S-M, Fu B, Lin M-T, Calhoun ES, Kamiyama M, Walter K, Nikolskaya T, Nikolsky Y, Hartigan J, Smith DR, Hidalgo M, Leach SD, Klein AP, Jaffee EM, Goggins M, Maitra A, Iacobuzio-Donahue C, Eshleman JR, Kern SE, Hruban RH. et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science. 2008;5:1801–1806. doi: 10.1126/science.1164368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jancík S, Drábek J, Radzioch D, Hajdúch M. Clinical relevance of KRAS in human cancers. Biomed Biotechnol. 2010;5:150960. doi: 10.1155/2010/150960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schutte M, Hruban RH, Geradts J, Maynard R, Hilgers W, Rabindran SK, Moskaluk CA, Hahn SA, Schwarte-waldhoff I, Schmiegel W, Baylin SB, Kern SE, Herman JG. Abrogation of the Rb/p16 tumor-suppressive pathway in virtually all pancreatic carcinomas. Cancer Res. 1997;5:3126–3130. [PubMed] [Google Scholar]
- Caldas C, Hahn SA, Da Costa LT, Redston MS, Schutte M, Seymour AB, Weinstein CL, Hruban RH, Yeo CJ, Kern SE. Frequent somatic mutations and homozygous deletions of the p16 (MTS1) gene in pancreatic adenocarcinoma. Nat Genet. 1994;5:27–32. doi: 10.1038/ng0994-27. [DOI] [PubMed] [Google Scholar]
- Liggett WH, Sidransky D. Role of the p16 tumor suppressor gene in cancer. J Clin Oncol. 1998;5:1197–1206. doi: 10.1200/JCO.1998.16.3.1197. [DOI] [PubMed] [Google Scholar]
- Moskaluk CA, Hruban RH, Kern SE. p16 and K-ras gene mutations in the intraductal precursors of human pancreatic adenocarcinoma. Cancer Res. 1997;5:2140–2143. [PubMed] [Google Scholar]
- Nigro JM, Baker SJ, Presinger AC, Jessup JM, Hostetter R, Cleary K, Bigner SH, Davidson N, Baylin S, Devilee P, Glover T, Collins FS, Weston A, Modali R, Harris CC, Vogelstein B. Mutations in the p53 gene occur in diverse human tumour types. Nature. 1989;5:705–708. doi: 10.1038/342705a0. [DOI] [PubMed] [Google Scholar]
- Barton C, Staddon S, Hughes C, Hall P, O'Sullivan C, Klöppel G, Theis B, Russell R, Neoptolemos J, Williamson R. Abnormalities of the p53 tumour suppressor gene in human pancreatic cancer. Br J Cancer. 1991;5:1076–1082. doi: 10.1038/bjc.1991.467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Redston MS, Caldas C, Seymour AB, Hruban RH, Costa L, Yeo CJ, Kern SE. p53 mutations in pancreatic carcinoma and evidence of common involvement of homocopolymer tracts in DNA microdeletions. Cancer Res. 1994;5:3025–3033. [PubMed] [Google Scholar]
- Heldin C-H, Moustakas A. Role of Smads in TGFβ signaling. Cell Tissue Res. 2012;5:21–36. doi: 10.1007/s00441-011-1190-x. [DOI] [PubMed] [Google Scholar]
- Hahn SA, Schutte M, Hoque AT, Moskaluk CA, Costa LT, Rozenblum E, Weinstein CL, Fischer A, Yeo CJ, Hruban RH, Kern SE. DPC4, a candidate tumor suppressor gene at human chromosome 18q21.1. Science. 1996;5:350–353. doi: 10.1126/science.271.5247.350. [DOI] [PubMed] [Google Scholar]
- Wilentz RE, Iacobuzio-Donahue CA, Argani P, Mccarthy DM, Parsons JL, Yeo CJ, Kern SE, Hruban RH. Loss of expression of Dpc4 in pancreatic intraepithelial neoplasia: evidence that DPC4 inactivation occurs late in neoplastic progression. Cancer Res. 2000;5:2002–2006. [PubMed] [Google Scholar]
- Lüttges J, Galehdari H, Bröcker V, Schwarte-Waldhoff I, Henne-Bruns D, Klöppel G, Schmiegel W, Hahn SA. Allelic loss is often the first hit in the biallelic inactivation of the p53 and DPC4 genes during pancreatic carcinogenesis. Am J Pathol. 2001;5:1677–1683. doi: 10.1016/S0002-9440(10)64123-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goggins M, Shekher M, Turnacioglu K, Coggins M, Yeo CJ, Hruban RH, Kern SE. Genetic alterations of the transforming growth factor β receptor genes in pancreatic and biliary adenocarcinomas. Cancer Res. 1998;5:5329–5332. [PubMed] [Google Scholar]
- Su GH, Bansal R, Murphy KM, Montgomery E, Yeo CJ, Hruban RH, Kern SE. ACVR1B (ALK4, activin receptor type 1B) gene mutations in pancreatic carcinoma. Proc Natl Acad Sci USA. 2001;5:3254–3257. doi: 10.1073/pnas.051484398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartsch DK, Gress TM, Langer P. Familial pancreatic cancer--current knowledge. Nat Rev Gastroenterol Hepatol. 2012;5:445–453. doi: 10.1038/nrgastro.2012.111. [DOI] [PubMed] [Google Scholar]
- Hruban RH, Canto MI, Goggins M, Schulick R, Klein AP. Update on familial pancreatic cancer. Adv Surg. 2010;5:293–311. doi: 10.1016/j.yasu.2010.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Su GH, Hruban RH, Bansal RK, Bova GS, Tang DJ, Shekher MC, Westerman AM, Entius MM, Goggins M, Yeo CJ, Kern SE. Germline and somatic mutations of the STK11/LKB1 Peutz-Jeghers gene in pancreatic and biliary cancers. Am J Pathol. 1999;5:1835–1840. doi: 10.1016/S0002-9440(10)65440-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goggins M, Schutte M, Lu J, Moskaluk CA, Weinstein CL, Petersen GM, Yeo CJ, Jackson CE, Lynch HT, Hruban RH, Kern SE. Germline BRCA2 gene mutations in patients with apparently sporadic pancreatic carcinomas. Cancer Res. 1996;5:5360–5364. [PubMed] [Google Scholar]
- Klein AP. Genetic susceptibility to pancreatic cancer. Mol Carcinog. 2012;5:14–24. doi: 10.1002/mc.20855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Heijden MS, Yeo CJ, Hruban RH, Kern SE. Fanconi anemia gene mutations in young-onset pancreatic cancer. Cancer Res. 2003;5:2585–2588. [PubMed] [Google Scholar]
- Rogers CD, Van der Heijden MS, Yeo CJ, Hruban RH, Kern SE, Goggins M. The genetics of FANCC and FANCG in familial pancreatic cancer. Cancer Biol Ther. 2004;5:167–169. doi: 10.4161/cbt.3.2.609. [DOI] [PubMed] [Google Scholar]
- Couch FJ, Johnson MR, Rabe K, Boardman L, Mcwilliams R. Germ line Fanconi anemia complementation group C mutations and pancreatic cancer. Cancer Res. 2005;5:383–386. [PubMed] [Google Scholar]
- Jones S, Hruban RH, Kamiyama M, Borges M, Zhang X, Parsons DW, Lin JC-H, Palmisano E, Brune K, Jaffee EM, Iacobuzio-Donahue CA, Maitra A, Parmigiani G, Kern SE, Velculescu VE, Kinzler KW, Vogelstein B, Eshleman JR, Goggins M, Klein AP. Exomic sequencing identifies PALB2 as a pancreatic cancer susceptibility gene. Science. 2009;5:217. doi: 10.1126/science.1171202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts NJ, Jiao Y, Yu J, Kopelovich L, Petersen GM, Bondy ML, Gallinger S, Schwartz AG, Syngal S, Cote ML, Axilbund J, Schulick R, Ali SZ, Eshleman JR, Velculescu VE, Goggins M, Vogelstein B, Papadopoulos N, Hruban RH, Kinzler KW, Klein AP. ATM mutations in patients with hereditary pancreatic cancer. Cancer Discov. 2012;5:41–46. doi: 10.1158/2159-8290.CD-11-0194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biankin AV, Waddell N, Kassahn KS, Gingras M-C, Muthuswamy LB, Johns AL, Miller DK, Wilson PJ, Patch A-M, Wu J, Chang DK, Cowley MJ, Gardiner BB, Song S, Harliwong I, Idrisoglu S, Nourse C, Nourbakhsh E, Manning S, Wani S, Gongora M, Pajic M, Scarlett CJ, Gill AJ, Pinho AV, Rooman I, Anderson M, Holmes O, Leonard C, Taylor D. et al. Pancreatic cancer genomes reveal aberrations in axon guidance pathway genes. Nature. 2012;5:399–405. doi: 10.1038/nature11547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ypsilanti AR, Zagar Y, Chédotal A. Moving away from the midline: new developments for Slit and Robo. Development. 2010;5:1939–1952. doi: 10.1242/dev.044511. [DOI] [PubMed] [Google Scholar]
- Mann KM, Ward JM, Yew CCK, Kovochich A, Dawson DW, Black MA, Brett BT, Sheetz TE, Dupuy AJ, Chang DK, Biankin AV, Waddell N, Kassahn KS, Grimmond SM, Rust AG, Adams DJ, Jenkins NA, Copeland NG. Sleeping Beauty mutagenesis reveals cooperating mutations and pathways in pancreatic adenocarcinoma. Proc Natl Acad Sci USA. 2012;5:5934–5941. doi: 10.1073/pnas.1202490109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ayala FJ. Darwin's greatest discovery: design without designer. Proc Natl Acad Sci USA. 2007;5(Suppl):8567–8573. doi: 10.1073/pnas.0701072104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowell PC. The clonal evolution of tumor cell populations. Science. 1976;5:23–28. doi: 10.1126/science.959840. [DOI] [PubMed] [Google Scholar]
- Fearon E, Vogelstein B. A genetic model for colorectal tumorigenesis. Cell. 1990;5:759–767. doi: 10.1016/0092-8674(90)90186-I. [DOI] [PubMed] [Google Scholar]
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;5:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- Stratton MR. Exploring the genomes of cancer cells: progress and promise. Science. 2011;5:1553–1558. doi: 10.1126/science.1204040. [DOI] [PubMed] [Google Scholar]
- Nguyen DX, Bos PD, Massagué J. Metastasis: from dissemination to organ-specific colonization. Nat Rev Cancer. 2009;5:274–284. doi: 10.1038/nrc2622. [DOI] [PubMed] [Google Scholar]
- Talmadge JE, Fidler IJ. The biology of cancer metastasis. Cancer Res. 2010;5:5649–5669. doi: 10.1158/0008-5472.CAN-10-1040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yachida S, White CM, Naito Y, Zhong Y, Brosnan JA, Macgregor-Das AM, Morgan RA, Saunders T, Laheru DA, Herman JM, Hruban RH, Klein AP, Jones S, Velculescu V, Wolfgang CL, Iacobuzio-Donahue CA. Clinical significance of the genetic landscape of pancreatic cancer and implications for identification of potential long-term survivors. Clin Cancer Res. 2012;5:6339–6347. doi: 10.1158/1078-0432.CCR-12-1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Massagué J. TGFβ in Cancer. Cell. 2008;5:215–30. doi: 10.1016/j.cell.2008.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yachida S, Jones S, Bozic I, Antal T, Leary R, Fu B, Kamiyama M, Hruban RH, Eshleman JR, Nowak MA, Velculescu VE, Kinzler KW, Vogelstein B, Iacobuzio-Donahue CA. Distant metastasis occurs late during the genetic evolution of pancreatic cancer. Nature. 2010;5:1114–1117. doi: 10.1038/nature09515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campbell PJ, Yachida S, Mudie LJ, Stephens PJ, Pleasance ED, Stebbings LA, Morsberger LA, Latimer C, McLaren S, Lin M-L, McBride DJ, Varela I, Nik-Zainal SA, Leroy C, Jia M, Menzies A, Butler AP, Teague JW, Griffin CA, Burton J, Swerdlow H, Quail MA, Stratton MR, Iacobuzio-Donahue C, Futreal PA. The patterns and dynamics of genomic instability in metastatic pancreatic cancer. Nature. 2010;5:1109–1113. doi: 10.1038/nature09460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz LA, Williams RT, Wu J, Kinde I, Hecht JR, Berlin J, Allen B, Bozic I, Reiter JG, Nowak MA, Kinzler KW, Oliner KS, Vogelstein B. The molecular evolution of acquired resistance to targeted EGFR blockade in colorectal cancers. Nature. 2012;5:537–540. doi: 10.1038/nature11219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misale S, Yaeger R, Hobor S, Scala E, Janakiraman M, Liska D, Valtorta E, Schiavo R, Buscarino M, Siravegna G, Bencardino K, Cercek A, Chen C-T, Veronese S, Zanon C, Sartore-Bianchi A, Gambacorta M, Gallicchio M, Vakiani E, Boscaro V, Medico E, Weiser M, Siena S, Di Nicolantonio F, Solit D, Bardelli A. Emergence of KRAS mutations and acquired resistance to anti-EGFR therapy in colorectal cancer. Nature. 2012;5:532–536. doi: 10.1038/nature11156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrett MT, Lenkiewicz E, Evers L, Holley T, Ruiz C, Bubendorf L, Sekulic A, Ramanathan RK, Von Hoff DD. Clonal evolution and therapeutic resistance in solid tumors. Front Pharmacol. 2013;5:2. doi: 10.3389/fphar.2013.00002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haeno H, Gonen M, Davis MB, Herman JM, Iacobuzio-Donahue CA, Michor F. Computational modeling of pancreatic cancer reveals kinetics of metastasis suggesting optimum treatment strategies. Cell. 2012;5:362–375. doi: 10.1016/j.cell.2011.11.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharan SK, Morimatsu M, Albrecht U, Lim D-S, Regel E, Dinh C, Sands A, Eichele G, Hasty P, Bradley A. Embryonic lethality and radiation hypersensitivity mediated by Rad51 in mice lacking Brca2. Nature. 1997;5:804–810. doi: 10.1038/386804a0. [DOI] [PubMed] [Google Scholar]
- Xia B, Sheng Q, Nakanishi K, Ohashi A, Wu J, Christ N, Liu X, Jasin M, Couch FJ, Livingston DM. Control of BRCA2 cellular and clinical functions by a nuclear partner, PALB2. Mol Cell. 2006;5:719–729. doi: 10.1016/j.molcel.2006.05.022. [DOI] [PubMed] [Google Scholar]
- Villarroel MC, Rajeshkumar NV, Garrido-Laguna I, De Jesus-Acosta A, Jones S, Maitra A, Hruban RH, Eshleman JR, Klein A, Laheru D, Donehower R, Hidalgo M. Personalizing cancer treatment in the age of global genomic analyses: PALB2 gene mutations and the response to DNA damaging agents in pancreatic cancer. Mol Cancer Ther. 2011;5:3–8. doi: 10.1158/1535-7163.MCT-10-0893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van der Heijden MS, Brody JR, Dezentje DA, Gallmeier E, Cunningham SC, Swartz MJ, DeMarzo AM, Offerhaus GJA, Isacoff WH, Hruban RH, Kern SE. In vivo therapeutic responses contingent on Fanconi anemia/BRCA2 status of the tumor. Clin Cancer Res. 2005;5:7508–7515. doi: 10.1158/1078-0432.CCR-05-1048. [DOI] [PubMed] [Google Scholar]
- James MR, Lehmann AR. Role of poly(adenosine diphosphate ribose) in deoxyribonucleic acid repair in human fibroblasts. Biochemistry. 1982;5:4007–4013. doi: 10.1021/bi00260a016. [DOI] [PubMed] [Google Scholar]
- Farmer H, McCabe N, Lord CJ, Tutt ANJ, Johnson DA, Richardson TB, Santarosa M, Dillon KJ, Hickson I, Knights C, Martin NMB, Jackson SP, Smith GCM, Ashworth A. Targeting the DNA repair defect in BRCA mutant cells as a therapeutic strategy. Nature. 2005;5:917–921. doi: 10.1038/nature03445. [DOI] [PubMed] [Google Scholar]
- Kraakman-van Der Zwet M, Overkamp WJI, Van Lange REE, Essers J, Van Duijn-goedhart A, Wiggers I, Swaminathan S, Van Buul PW, Errami A, Tan RTL, Jaspers GJ, Sharan SK, Kanaar R, Zdzienicka MZ. Brca2 (XRCC11) Deficiency results in radioresistant DNA synthesis and a higher frequency of spontaneous deletions. Mol Cell Biol. 2002;5:669–679. doi: 10.1128/MCB.22.2.669-679.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryant HE, Schultz N, Thomas HD, Parker KM, Flower D, Lopez E, Kyle S, Meuth M, Curtin NJ, Helleday T. Specific killing of BRCA2-deficient tumours with inhibitors of poly(ADP-ribose) polymerase. Nature. 2005;5:913–917. doi: 10.1038/nature03443. [DOI] [PubMed] [Google Scholar]
- Mccabe N, Lord CJ, Tutt ANJ, Martin NMB, Smith GCM, Ashworth A. BRCA2-deficient CAPAN-1 cells are extremely sensitive to the inhibition of poly (ADP-ribose) polymerase. Cancer Biol Ther. 2005;5:934–936. doi: 10.4161/cbt.4.9.2141. [DOI] [PubMed] [Google Scholar]
- Tutt A, Robson M, Garber JE, Domchek SM, Audeh MW, Weitzel JN, Friedlander M, Arun B, Loman N, Schmutzler RK, Wardley A, Mitchell G, Earl H, Wickens M, Carmichael J. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and advanced breast cancer: a proof-of-concept trial. Lancet. 2010;5:235–244. doi: 10.1016/S0140-6736(10)60892-6. [DOI] [PubMed] [Google Scholar]
- Audeh MW, Carmichael J, Penson RT, Friedlander M, Powell B, Bell-McGuinn KM, Scott C, Weitzel JN, Oaknin A, Loman N, Lu K, Schmutzler RK, Matulonis U, Wickens M, Tutt A. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patients with BRCA1 or BRCA2 mutations and recurrent ovarian cancer: a proof-of-concept trial. Lancet. 2010;5:245–251. doi: 10.1016/S0140-6736(10)60893-8. [DOI] [PubMed] [Google Scholar]
- ClinicalTrials.gov. http://www.clinicaltrials.gov
- Nijman SMB. Synthetic lethality: general principles, utility and detection using genetic screens in human cells. FEBS Lett. 2011;5:1–6. doi: 10.1016/j.febslet.2010.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcotte R, Brown KR, Suarez F, Sayad A, Karamboulas K, Krzyzanowski PM, Sircoulomb F, Medrano M, Fedyshyn Y, Koh JLY, Van Dyk D, Fedyshyn B, Luhova M, Brito GC, Vizeacoumar FJ, Vizeacoumar FS, Datti A, Kasimer D, Buzina A, Mero P, Misquitta C, Normand J, Haider M, Ketela T, Wrana JL, Rottapel R, Neel BG, Moffat J. Essential gene profiles in breast, pancreatic, and ovarian cancer cells. Cancer Discov. 2012;5:172–189. doi: 10.1158/2159-8290.CD-11-0224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ward AF, Braun BS, Shannon KM. Targeting oncogenic Ras signaling in hematologic malignancies. Blood. 2012;5:3397–3406. doi: 10.1182/blood-2012-05-378596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scholl C, Fröhling S, Dunn IF, Schinzel AC, Barbie DA, Kim SY, Silver SJ, Tamayo P, Wadlow RC, Ramaswamy S, Döhner K, Bullinger L, Sandy P, Boehm JS, Root DE, Jacks T, Hahn WC, Gilliland DG. Synthetic lethal interaction between oncogenic KRAS dependency and STK33 suppression in human cancer cells. Cell. 2009;5:821–834. doi: 10.1016/j.cell.2009.03.017. [DOI] [PubMed] [Google Scholar]
- Barbie DA, Tamayo P, Boehm JS, Kim SY, Moody SE, Dunn IF, Schinzel AC, Sandy P, Meylan E, Scholl C, Fröhling S, Chan EM, Sos ML, Michel K, Mermel C, Silver SJ, Weir BA, Reiling JH, Sheng Q, Gupta PB, Wadlow RC, Le H, Hoersch S, Wittner BS, Ramaswamy S, Livingston DM, Sabatini DM, Meyerson M, Thomas RK, Lander ES. et al. Systematic RNA interference reveals that oncogenic KRAS-driven cancers require TBK1. Nature. 2009;5:108–112. doi: 10.1038/nature08460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo J, Emanuele MJ, Li D, Creighton CJ, Schlabach MR, Westbrook TF, Wong K-K, Elledge SJ. A genome-wide RNAi screen identifies multiple synthetic lethal interactions with the Ras oncogene. Cell. 2009;5:835–848. doi: 10.1016/j.cell.2009.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collisson EA, Trejo CL, Silva JM, Gu S, Korkola JE, Heiser LM, Charles R-P, Rabinovich BA, Hann B, Dankort D, Spellman PT, Phillips WA, Gray JW, McMahon M. A central role for RAF→MEK→ERK signaling in the genesis of pancreatic ductal adenocarcinoma. Cancer Discov. 2012;5:685–693. doi: 10.1158/2159-8290.CD-11-0347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diep CH, Munoz RM, Choudhary A, Von Hoff DD, Han H. Synergistic effect between erlotinib and MEK inhibitors in KRAS wild-type human pancreatic cancer cells. Clin Cancer Res. 2011;5:2744–2756. doi: 10.1158/1078-0432.CCR-10-2214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui Y, Brosnan JA, Blackford AL, Sur S, Hruban RH, Kinzler KW, Vogelstein B, Maitra A, Diaz LA, Iacobuzio-Donahue CA, Eshleman JR. Genetically defined subsets of human pancreatic cancer demonstrate unique in vitro chemosensitivity. Clin Cancer Res. 2012;5:6519–6530. doi: 10.1158/1078-0432.CCR-12-0827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- von Hoff DD, Stephenson JJ, Rosen P, Loesch DM, Borad MJ, Anthony S, Jameson G, Brown S, Cantafio N, Richards DA, Fitch TR, Wasserman E, Fernandez C, Green S, Sutherland W, Bittner M, Alarcon A, Mallery D, Penny R. Pilot study using molecular profiling of patients' tumors to find potential targets and select treatments for their refractory cancers. J Clin Oncol. 2010;5:4877–4883. doi: 10.1200/JCO.2009.26.5983. [DOI] [PubMed] [Google Scholar]
- Wang H, Stephens B, Von Hoff DD, Han H. Identification and characterization of a novel anticancer agent with selectivity against deleted in pancreatic cancer locus 4 (DPC4)-deficient pancreatic and colon cancer cells. Pancreas. 2009;5:551–557. doi: 10.1097/MPA.0b013e31819d7415. [DOI] [PubMed] [Google Scholar]
- Wang H, Han H, Von Hoff DD. Identification of an agent selectively targeting DPC4 (deleted in pancreatic cancer locus 4)-deficient pancreatic cancer cells. Cancer Res. 2006;5:9722–9730. doi: 10.1158/0008-5472.CAN-05-4602. [DOI] [PubMed] [Google Scholar]
- Laffan TA, Horton KM, Klein AP, Berlanstein B, Siegelman SS, Kawamoto S, Johnson PT, Fishman EK, Hruban RH. Prevalence of unsuspected pancreatic cysts on MDCT. Am J Roentgenol. 2008;5:802–807. doi: 10.2214/AJR.07.3340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matthaei H, Schulick RD, Hruban RH, Maitra A. Cystic precursors to invasive pancreatic cancer. Nat Rev Gastroenterol Hepatol. 2011;5:141–150. doi: 10.1038/nrgastro.2011.2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J, Jiao Y, Dal Molin M, Maitra A, De Wilde RF, Wood LD, Eshleman JR, Goggins MG, Wolfgang CL, Canto MI, Schulick RD, Edil BH, Choti MA, Adsay V, Klimstra DS, Offerhaus GJA, Klein AP, Kopelovich L, Carter H, Karchin R, Allen PJ, Schmidt CM, Naito Y, Diaz LA, Kinzler KW, Papadopoulos N, Hruban RH, Vogelstein B. Whole-exome sequencing of neoplastic cysts of the pancreas reveals recurrent mutations in components of ubiquitin-dependent pathways. Proc Natl Acad Sci USA. 2011;5:21188–21193. doi: 10.1073/pnas.1118046108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondo N, Murakami Y, Uemura K, Hayashidani Y, Sudo T, Hashimoto Y, Nakashima A, Sakabe R, Shigemoto N, Kato Y, Ohge H, Sueda T. Prognostic impact of perioperative serum CA 19-9 levels in patients with resectable pancreatic cancer. Ann Surg Oncol. 2010;5:2321–2329. doi: 10.1245/s10434-010-1033-0. [DOI] [PubMed] [Google Scholar]
- Barton JG, Bois JP, Sarr MG, Wood CM, Qin R, Thomsen KM, Kendrick ML, Farnell MB. Predictive and prognostic value of CA 19-9 in resected pancreatic adenocarcinoma. J Gastrointest Surg. 2009;5:2050–2058. doi: 10.1007/s11605-009-0849-z. [DOI] [PubMed] [Google Scholar]
- Leary RJ, Sausen M, Kinde I, Papadopoulos N, Carpten JD, Craig D, O'Shaughnessy J, Kinzler KW, Parmigiani G, Vogelstein B, Diaz LA, Velculescu VE. Detection of chromosomal alterations in the circulation of cancer patients with whole-genome sequencing. Sci Transl Med. 2012;5:162ra154. doi: 10.1126/scitranslmed.3004742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinde I, Bettegowda C, Wang Y, Wu J, Agrawal N, Shih I-M, Kurman R, Dao F, Levine DA, Giuntoli R, Roden R, Eshleman JR, Carvalho JP, Marie SKN, Papadopoulos N, Kinzler KW, Vogelstein B, Diaz LA. Evaluation of DNA from the papanicolaou test to detect ovarian and endometrial cancers. Sci Transl Med. 2013;5:167ra4. doi: 10.1126/scitranslmed.3004952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conroy T, Desseigne F, Ychou M, Bouche O, Guimbaud R, Becouarn Y, Adenis A, Raoul J-L, Gourgou-Bourgade S, De la Douchardiere C, Bennouna J, Bachet J-B, Khemissa-Akouz F, Pere-Verge D, Delbaldo C, Assenat E, Chauggert B, Michel P, Montoto-Grillot C, Ducreux M. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N Engl J Med. 2011;5:768–769. doi: 10.1056/NEJMoa1011923. [DOI] [PubMed] [Google Scholar]
- Heinemann V, Quietzsch D, Gieseler F, Gonnermann M, Schönekäs H, Rost A, Neuhaus H, Haag C, Clemens M, Heinrich B, Vehling-Kaiser U, Fuchs M, Fleckenstein D, Gesierich W, Uthgenannt D, Einsele H, Holstege A, Hinke A, Schalhorn A, Wilkowski R. Randomized phase III trial of gemcitabine plus cisplatin compared with gemcitabine alone in advanced pancreatic cancer. J Clin Oncol. 2006;5:3946–3952. doi: 10.1200/JCO.2005.05.1490. [DOI] [PubMed] [Google Scholar]
- Stathopoulos GP, Syrigos K, Aravantinos G, Polyzos A, Papakotoulas P, Fountzilas G, Potamianou A, Ziras N, Boukovinas J, Varthalitis J, Androulakis N, Kotsakis A, Samonis G, Georgoulias V. A multicenter phase III trial comparing irinotecan-gemcitabine (IG) with gemcitabine (G) monotherapy as first-line treatment in patients with locally advanced or metastatic pancreatic cancer. Br J Cancer. 2006;5:587–592. doi: 10.1038/sj.bjc.6603301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrmann R, Bodoky G, Ruhstaller T, Glimelius B, Bajetta E, Schüller J, Saletti P, Bauer J, Figer A, Pestalozzi B, Köhne C-H, Mingrone W, Stemmer SM, Tàmas K, Kornek GV, Koeberle D, Cina S, Bernhard J, Dietrich D, Scheithauer W. Gemcitabine plus capecitabine compared with gemcitabine alone in advanced pancreatic cancer: a randomized, multicenter, phase III trial of the Swiss Group for Clin Cancer Res and the Central European Cooperative Oncology Group. J Clin Oncol. 2007;5:2212–2217. doi: 10.1200/JCO.2006.09.0886. [DOI] [PubMed] [Google Scholar]
- Cunningham D, Chau I, Stocken DD, Valle JW, Smith D, Steward W, Harper PG, Dunn J, Tudur-Smith C, West J, Falk S, Crellin A, Adab F, Thompson J, Leonard P, Ostrowski J, Eatock M, Scheithauer W, Herrmann R, Neoptolemos JP. Phase III randomized comparison of gemcitabine versus gemcitabine plus capecitabine in patients with advanced pancreatic cancer. J Clin Oncol. 2009;5:5513–5518. doi: 10.1200/JCO.2009.24.2446. [DOI] [PubMed] [Google Scholar]
- De Jesus-Acosta A, Oliver GR, Blackford A, Kinsman K, Flores EI, Wilfong LS, Zheng L, Donehower RC, Cosgrove D, Laheru D, Le DT, Chung K, Diaz LA. A multicenter analysis of GTX chemotherapy in patients with locally advanced and metastatic pancreatic adenocarcinoma. Cancer Chemother Pharmacol. 2012;5:415–424. doi: 10.1007/s00280-011-1704-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niihori T, Aoki Y, Narumi Y, Neri G, Cavé H, Verloes A, Okamoto N, Hennekam RCM, Gillessen-Kaesbach G, Wieczorek D, Kavamura MI, Kurosawa K, Ohashi H, Wilson L, Heron D, Bonneau D, Corona G, Kaname T, Naritomi K, Baumann C, Matsumoto N, Kato K, Kure S, Matsubara Y. Germline KRAS and BRAF mutations in cardio-facio-cutaneous syndrome. Nat Genet. 2006;5:294–296. doi: 10.1038/ng1749. [DOI] [PubMed] [Google Scholar]
- Bartsch DK, Sina-Frey M, Lang S, Wild A, Gerdes B, Barth P, Kress R, Grützmann R, Colombo-Benkmann M, Ziegler A, Hahn SA, Rothmund M, Rieder H. CDKN2A germline mutations in familial pancreatic cancer. Ann Surg. 2002;5:730–737. doi: 10.1097/00000658-200212000-00005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker JF, Florell SR, Alexander A, DiSario JA, Shami PJ, Leachman SA. Pancreatic carcinoma surveillance in patients with familial melanoma. Arch Dermatol. 2003;5:1019–1025. doi: 10.1001/archderm.139.8.1019. [DOI] [PubMed] [Google Scholar]
- Varley JM. Germline TP53 mutations and Li-Fraumeni syndrome. Hum Mutat. 2003;5:313–320. doi: 10.1002/humu.10185. [DOI] [PubMed] [Google Scholar]
- Andrabi S, Bekheirnia MR, Robbins-Furman P, Lewis RA, Prior TW, Potocki L. SMAD4 mutation segregating in a family with juvenile polyposis, aortopathy, and mitral valve dysfunction. Am J Med Genet. 2011;5:1165–1169. doi: 10.1002/ajmg.a.33968. [DOI] [PubMed] [Google Scholar]
- Caputo V, Cianetti L, Niceta M, Carta C, Ciolfi A, Bocchinfuso G, Carrani E, Dentici ML, Biamino E, Belligni E, Garavelli L, Boccone L, Melis D, Andria G, Gelb BD, Stella L, Silengo M, Dallapiccola B, Tartaglia M. A restricted spectrum of mutations in the SMAD4 tumor-suppressor gene underlies Myhre syndrome. Am J Hum Genet. 2012;5:161–169. doi: 10.1016/j.ajhg.2011.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loeys BL, Schwarze U, Holm T, Callewaert BL, Thomas GH, Pannu H, De Backer JF, Oswald GL, Symoens S, Manouvrier S, Roberts AE, Faravelli F, Greco MA, Pyeritz RE, Milewicz DM, Coucke PJ, Cameron DE, Braverman AC, Byers PH, De Paepe AM, Dietz HC. Aneurysm syndromes caused by mutations in the TGF-beta receptor. N Engl J Med. 2006;5:788–798. doi: 10.1056/NEJMoa055695. [DOI] [PubMed] [Google Scholar]
- Xin W, Yun KJ, Ricci F, Zahurak M, Qiu W, Su GH, Yeo CJ, Hruban RH, Kern SE, Iacobuzio-Donahue CA. MAP2K4/MKK4 expression in pancreatic cancer: genetic validation of immunohistochemistry and relationship to disease course. Clin Cancer Res. 2004;5:8516–8520. doi: 10.1158/1078-0432.CCR-04-0885. [DOI] [PubMed] [Google Scholar]
- Balakrishnan A, Bleeker FE, Lamba S, Rodolfo M, Daniotti M, Scarpa A, Van Tilborg AA, Leenstra S, Zanon C, Bardelli A. Novel somatic and germline mutations in cancer candidate genes in glioblastoma, melanoma, and pancreatic carcinoma. Cancer Res. 2007;5:3545–3550. doi: 10.1158/0008-5472.CAN-07-0065. [DOI] [PubMed] [Google Scholar]
- Tsurusaki Y, Okamoto N, Ohashi H, Kosho T, Imai Y, Hibi-Ko Y, Kaname T, Naritomi K, Kawame H, Wakui K, Fukushima Y, Homma T, Kato M, Hiraki Y, Yamagata T, Yano S, Mizuno S, Sakazume S, Ishii T, Nagai T, Shiina M, Ogata K, Ohta T, Niikawa N, Miyatake S, Okada I, Mizuguchi T, Doi H, Saitsu H, Miyake N. et al. Mutations affecting components of the SWI/SNF complex cause Coffin-Siris syndrome. Nat Genet. 2012;5:376–378. doi: 10.1038/ng.2219. [DOI] [PubMed] [Google Scholar]
- Shain AH, Giacomini CP, Matsukuma K, Karikari CA, Bashyam MD, Hidalgo M, Maitra A, Pollack JR. Convergent structural alterations define SWItch/Sucrose NonFermentable (SWI/SNF) chromatin remodeler as a central tumor suppressive complex in pancreatic cancer. Proc Natl Acad Sci USA. 2011;5:E252–E259. doi: 10.1073/pnas.1114817109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneppenheim R, Frühwald MC, Gesk S, Hasselblatt M, Jeibmann A, Kordes U, Kreuz M, Leuschner I, Martin Subero JI, Obser T, Oyen F, Vater I, Siebert R. Germline nonsense mutation and somatic inactivation of SMARCA4/BRG1 in a family with rhabdoid tumor predisposition syndrome. Am J Hum Genet. 2010;5:279–284. doi: 10.1016/j.ajhg.2010.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozcelik H, Schmocker B, Di Nicola N, Shi X-H, Langer B, Moore M, Taylor BR, Narod SA, Darlington G, Andrulis IL, Gallinger S, Redston M. Germline BRCA2 6174delT mutations in Ashkenazi Jewish pancreatic cancer patients. Nat Genet. 1997;5:17–18. doi: 10.1038/ng0597-17. [DOI] [PubMed] [Google Scholar]
- Lynch HT, Deters CA, Snyder CL, Lynch JF, Villeneuve P, Silberstein J, Martin H, Narod SA, Brand RE. BRCA1 and pancreatic cancer: pedigree findings and their causal relationships. Cancer Genet Cytogenet. 2005;5:119–125. doi: 10.1016/j.cancergencyto.2004.01.032. [DOI] [PubMed] [Google Scholar]
- Lowenfels AB, Eugene P, Elitsur Y, Gates LK, Whitcomb DC. Hereditary pancreatitis and the risk of pancreatic cancer. J Natl Cancer Inst. 1997;5:442–446. doi: 10.1093/jnci/89.6.442. [DOI] [PubMed] [Google Scholar]
- Giardiello FM, Brensinger JD, Tersmette AC, Goodman SN, Petersen GM, Booker SV, Cruz-Correa M, Offerhaus JA. Very high risk of cancer in familial Peutz-Jeghers syndrome. Gastroenterology. 2000;5:1447–1453. doi: 10.1053/gast.2000.20228. [DOI] [PubMed] [Google Scholar]
- Goggins M, Offerhaus GJ, Hilgers W, Griffin CA, Shekher M, Tang D, Sohn TA, Yeo CJ, Kern SE, Hruban RH. Pancreatic adenocarcinomas with DNA replication errors (RER+) are associated with wild-type K-ras and characteristic histopathology. Poor differentiation, a syncytial growth pattern, and pushing borders suggest RER+. Am J Pathol. 1998;5:1501–1507. [PMC free article] [PubMed] [Google Scholar]
- Yamamoto H, Itoh F, Nakamura H. Genetic and clinical features of human pancreatic ductal adenocarcinomas with widespread microsatellite instability. Cancer Res. 2001;5:3139–3144. [PubMed] [Google Scholar]