

Abstract
AIM: To evaluate the effects of different types of dietary fats on the hepatic lipid content and oxidative stress parameters in rat liver with experimental non-alcoholic fatty liver disease (NAFLD).
METHODS: A total of 32 Sprague-Dawley rats were randomly divided into five groups. The rats in the control group (n = 8) were on chow diet (Group 1), rats (n = 6) on methionine choline-deficient diet (MCDD) (Group 2), rats (n = 6) on MCDD enriched with olive oil (Group 3), rats (n = 6) on MCDD with fish oil (Group 4) and rats (n = 6) on MCDD with butter fat (Group 5). After 2 mo, blood and liver sections were examined for lipids composition and oxidative stress parameters.
RESULTS: The liver weight/rat weight ratio increased in all treatment groups as compared with the control group. Severe fatty liver was seen in MCDD + fish oil and in MCDD + butter fat groups, but not in MCDD and MCDD + olive oil groups. The increase in hepatic triglycerides (TG) levels was blunted by 30% in MCDD + olive oil group (0.59 ± 0.09) compared with MCDD group (0.85 ± 0.04, p < 0.004), by 37% compared with MCDD + fish oil group (0.95 ± 0.07, p < 0.001), and by 33% compared with MCDD + butter group (0.09 ± 0.1, p < 0.01). The increase in serum TG was lowered by 10% in MCDD + olive oil group (0.9 ± 0.07) compared with MCDD group (1.05 ± 0.06). Hepatic cholesterol increased by 15-fold in MCDD group [(0.08 ± 0.02, this increment was blunted by 21% in MCDD + fish oil group (0.09 ± 0.02)]. In comparison with the control group, ratio of long-chain polyunsaturated fatty acids omega-6/omega-3 increased in MCDD + olive oil, MCDD + fish oil and MCDD + butter fat groups by 345-, 30- and 397-fold, respectively. In comparison to MCDD group (1.58 ± 0.08), hepatic MDA contents in MCDD + olive oil (3.3 ± 0.6), MCDD + fish oil (3.0 ± 0.4), and MCDD + butter group (2.9 ± 0.36) were increased by 108%, 91% and 87%, respectively (p < 0.004). Hepatic paraoxonase activity decreased significantly in all treatment groups, mostly with MCDD + olive oil group (-68%).
CONCLUSION: Olive oil decreases the accumulation of triglyceride in the liver of rats with NAFLD, but does not provide the greatest antioxidant activity.
Keywords: Fatty liver, Non-alcoholic steatohepatitis, Dietary fat, Fatty acids, Methionine choline-deficient diet, Insulin resistance, Olive oil, Oxidative stress, Paraoxonase
INTRODUCTION
Non-alcoholic steatohepatitis (NASH) pathogenesis includes insulin resistance, increased exposure of the hepatocytes to TNF-α and increased oxidative stress[1]. The development of hepatic steatosis results from an imbalance in the rate of entry, synthesis, or clearance of fat from the liver. More specifically, the increased influx of free fatty acids to the liver, reduced free fatty acid β-oxidation, a reduced hepatic secretion of triglyceride-rich lipoproteins and increased lipid peroxidation may be altered[1].
Hepatic triglycerides are derived from different sources. First, from endogenous fatty acids which are synthesized in the liver. Second, from dietary triglycerides that enter the liver via the chylomicron remnant pathway and finally from free fatty acids that are released from adipose tissues during periods of fasting. An impaired postprandial triglyceride response has been reported in patients with NASH[2]. This may promote fatty liver in several ways, such as increasing hepatic uptake of triglyceride-rich lipoproteins and their remnants in the postprandial period[2,3]. The contribution of hepatic lipogenesis to triglyceride secretion was 3 times higher in the patients as compared with the controls while that of non-esterified fatty acids (NEFA) re-esterification was reduced by 52%. Thus, enhanced lipogenesis appears as a prominent abnormality of hepatic fatty metabolism in the subjects with NASH[4].
Several studies have emphasized the importance of dietary composition in the treatment of fatty liver. However, the optimal diet for prevention of NASH is unknown. NASH patients had significantly higher overnight fasting blood concentration of total and free fatty acids than controls, and higher total saturated and monounsaturated levels of lipid fractions, mainly due to increase of palmitate, palmitoleate and oleate acids[5,6]. Low-calorie diet for 2 wk in Sprague-Dawley rats exerted no significant effects on either serum lipid disorders or hepatic inflammatory changes[7]. Dietary omega-3 and omega-6 polyunsaturated fatty acids (PUFA), present in fish oil, can regulate hepatic lipogenesis by reducing sterol-regulatory element-binding protein-1 in the liver[8]. Moreover, PUFA administration decreased fatty acids, such as oleate (C18:1 n-9), palmitate (C16:0) and palmitoleate (C16:1 n-7)[9]. Diet of NASH patients who were free of hyperlipidemia, diabetes and obesity was richer in saturated fat and poorer in PUFA[10]. PUFA may act as “fuel partitioners”, directing fatty acids away from triglyceride storage and toward oxidation[11]. A recent clinical study that compared 19 patients who had non-alcoholic fatty liver disease (NAFLD) with 11 normal control subjects revealed that patients with NAFLD had a higher ratio of long-chain polyunsaturated fatty acids omega-6/omega-3 in their livers than did control subjects[6]. When dietary monounsaturated fatty acids, mainly oleic acid present in olive oil, replace high-carbohydrate diets, plasma triglycerides concentration decrease, both in patients without diabetes and in those with type 2 diabetes[12]. These findings provide the rational for undertaking the experiment to test the hypothesis that supplementation of different oils to methionine choline-deficient diet could attenuate fatty liver changes in a novel model of hepatic steatosis.
The aim of the current study was to evaluate the effects of different types of dietary fats (monosaturated fatty acids present in olive oil; polyunsaturated fatty acids present in fish oil, and saturated fatty acids present in butter) on the hepatic lipid content and oxidative stress in rat liver with experimental NAFLD.
MATERIALS AND METHODS
Animals and protocol
Methionine choline-deficient diet (MCDD) is the most widely used model for NASH in rats[13]. The development of steatosis is caused by impaired very low-density lipoprotein (VLDL) secretion. Unlike animals fed a choline-sufficient diet which developed only steatosis but not inflammation and fibrosis, animals fed an MCDD diet showed fibrosing steatohepatitis[13]. Thirty-two Sprague-Dawley rats were randomly divided into five groups. The animals in the control group (n = 8) were on chow diet (Group 1). In group 2, there were six rats on methionine choline-deficient diet (MCDD). In group 3, there were six rats on MCDD supplemented with olive oil (0.45 mg/g rat weight). In group 4, six rats were on MCDD with fish oil (0.45 mg/g rat weight) and in group 5, 6 rats were on MCDD with butter-fat (0.45 mg/g rat weight). In all treatment groups, the MCDD diet was supplemented simply with the different oils and not as a semi-purified diet. The dietary fat was monitored daily and was supplied by the local pharmacy of the Sieff Hospital, Safed, Israel, and not by commercial sources. After two months, the rats were generally anesthetized and blood was sampled by heart puncture. Rats were sacrificed and the liver sections were examined. All animals were treated humanely and in accordance with the guidelines of the institution. The composition of the experimental diets and the levels of lipid fractions are shown in Table 1.
Table 1.
Lipid composition of dietary fats (g/100 g total lipids)
| Lipid | Olive oil | Fish oil | Butter |
| Energy (kcal) | 884 | 902 | 717 |
| Fatty acid, total saturated | 13.8 | 29.89 | 51.36 |
| 16:0 palmitic | 11.29 | 16.64 | 21.69 |
| Fatty acid, total monounsaturated | 72.96 | 33.84 | 21.02 |
| 18:1 undifferentiated | 71.26 | 14.75 | 19.96 |
| Fatty acid, total polyunsaturated | 10.52 | 31.86 | 3.04 |
| 18:2 undifferentiated | 9.76 | 2.01 | 2.72 |
Biochemistry
Determination of hepatic lipids composition: Rat liver lipid was extracted using the homogenate according to Folch et al[14]. Triglyceride concentration was determined according to the method of Gottfried and Rosenberg[15]. Total cholesterol concentration was determined as previously described[16]. Total lipid concentrations were determined using sulphophosphovaniline[17]. Phospholipids concentrations were determined by lipid phosphorus assay[18]. Phospholipids were separated into component species by thin layer chromatography (TLC) using silica gel G plates and chloroform: methanol: water (65:25:4, v/v) as the solvent system. The position of individual phospholipids was determined using the respective standards. The results were reported as percentage of total phospholipids content[19].
Determination of hepatic pro-oxidant and anti-oxidants: Paraoxonase activity was measured according to the method described elsewhere[20] using 1.0 mmol/L phenyl acetate as substrate. α-tocopherol was estimated spectrophotometrically[21]. Hepatic maleic dialdehyde (MDA) was estimated spectrophotometrically using thiobarbituric acid assay[22].
Fatty acids composition: Rat liver cytosol was prepared by differential centrifugation. Approximately 0.5 g of the liver was homogenized in 5 mL of ice-cold 50 mmol/L phosphate buffer solution (pH 7.4). The liver homogenate was centrifuged for 10 min at 5000 r/min at 4°C, and the pellet was discarded. The supernatant was further centrifuged for 30 min at 8000 r/min at 4°C. Fatty acid methyl esters were analyzed by gas chromatography using palmitic acid as the marker of fatty acids[23].
Histology
The specimens were fixed in buffered formalin and embedded in paraffin. Hematoxylin and eosin (H&E) sections, periodic acid Schiff with diastase stain for inflammatory grading as well as Mason’s trichrome and reticulin stains were reviewed for fibrosis and architectural changes. The necroinflammatory grade and stage of fibrosis were assessed according the method of Brunt
et al[24]. Steatosis was graded by semi-quantitative analysis as follows: mild = 5%-30%; moderate = 30%-60%; and severe = more than 60% of hepatocytes affected. Each biopsy was analyzed and graded by the same pathologist who was blinded to the rats group.
Statistical analysis
Results were expressed as mean ± SD. Analysis of variance was used to compare multiple group means, followed by the Newman-Keuls test to determine statistical significance between two groups. When the data were not normally distributed, the Kruskal-Wallis test was performed to compare multiple group means, followed by Mann-Whitney test. Correlation analysis was performed using Spearman rank correlation. The statistical comparisons were performed using the unit values rather than percentages. P < 0.05 was considered statistically significant. The statistical analysis was performed using the Winstat program for windows (Kalmia, MA).
RESULTS
Liver weight/rat weight ratio
Rat weights significantly increased in the control group by 49% and decreased in MCDD, MCDD + olive oil, MCDD + fish oil, and MCDD + butter group by 35%, 40%, 40%, and 37%, respectively (P < 0.05). Rat liver weights decreased relatively to the control group in MCDD, MCDD + olive oil, MCDD + fish oil, and MCDD + butter group by 22%, 25%, 34%, and 25%, respectively (P < 0.05, Table 2). Rat liver weight: rat weight ratios were markedly increased in MCDD, MCDD + olive oil, MCDD + fish oil, and MCDD + butter group by 54%, 54%, 38%, and 54%, respectively (P < 0.05).
Table 2.
Effect of MCDD alone or enriched with different fats on rat body or liver weight (mean ± SD, n = 6)
| Group | Diet | Baseline rats weight (g) | Final rats weight (g) | Final liver weight (g) | Liver weight/rat weight ratio |
| 1 | Control | 246 ± 6 | 367 ± 3 | 8.8 ± 0.3 | 0.023 |
| 2 | MCDD | 249 ± 6 | 184 ± 8 | 6.8 ± 0.6 | 0.037ª |
| 3 | MCDD + olive oil | 247 ± 6 | 177 ± 7 | 6.6 ± 0.6 | 0.037ª |
| 4 | MCDD + fish oil | 246 ± 7 | 176 ± 6 | 5.8 ± 0.6 | 0.033ª |
| 5 | MCDD + butter | 242 ± 5 | 178 ± 6 | 6.6 ± 0.5 | 0.037ª |
ªP < 0.05 vs control group.
Extent of fatty infiltration
There was a significant increase in the extent of fatty liver in MCDD, MCDD + olive oil, MCDD + fish oil, and MCDD + butter groups as compared to the control group (P < 0.001). While no severe fatty (> 60%) infiltration was observed in MCDD + olive oil group, severe hepatic fatty infiltration was observed in 33% of rats in MCDD + fish oil group and in 83% of rats in MCDD + butter group (P < 0.01, Figure 1A). Mild to moderate fatty infiltration was similar in MCDD and MCDD + olive oil groups but significantly lower in MCDD + fish oil (67% of rats) and MCDD +butter group (17% of rats) (P < 0.01, Figure 1B). No inflammation was detected in the livers of the control group. Mild inflammatory changes were observed in all rats in MCDD, and MCDD + olive oil groups. While 66% of the rats in MCDD + butter group were without hepatic inflammatory changes, 50% of the rats in MCDD + fish oil group had moderate inflammatory changes (Table 3).
Figure 1.
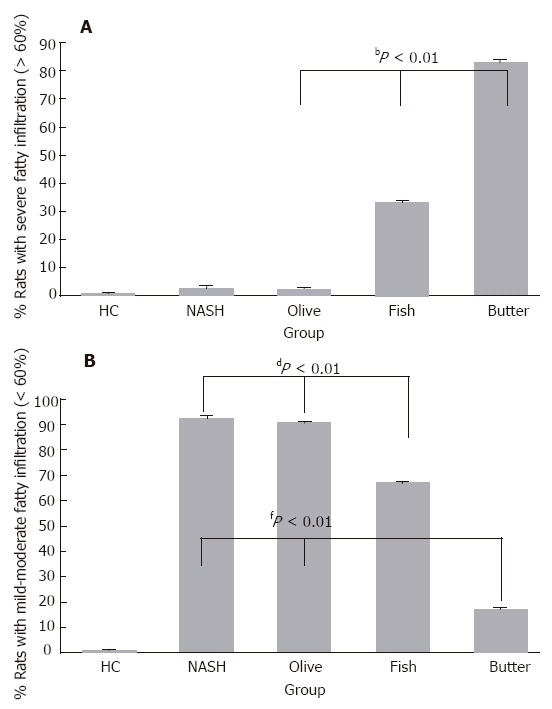
Extent of hepatic fatty infiltration (A: severe; B: mild) in the different treatment groups. bP < 0.01, comparison among olive oil, fish oil and butter fat groups; dP < 0.01, comparison among fish oil, NASH and olive oil groups; fP < 0.01, comparison among butter fat, NASH and olive oil groups.
Table 3.
Effect of MCDD alone or enriched with different fats on hepatic fat infiltration and inflammation
| Group | Diet |
Fatty liver |
Inflammation |
|||
| < 60% (%) | > 60% (%) | Absent (%) | Mild (%) | Moderate (%) | ||
| 1 | Control (n = 8) | 0 | 0 | 0 | 0 | 0 |
| 2 | MCDD (n = 5) | 100ª | 0 | 0 | 100ª | 0 |
| 3 | MCDD + olive oil (n = 6) | 100ª | 0 | 0 | 100ª | 0 |
| 4 | MCDD + fish oil (n = 6) | 67ª | 33.3ª | 17 | 33.3ª | 50ac |
| 5 | MCDD + butter (n = 6) | 17 | 83ª | 66ac | 17 | 17 |
ªP < 0.05 vs control group; cP < 0.05 vs group 3.
Hepatic lipids
Hepatic triglyceride content (mol/g liver) increased by 43-, 30-, 48- and 45-fold in MCDD (0.85 ± 0.04), MCDD + olive oil (0.59 ± 0.09), MCDD + fish oil (0.95 ± 0.07), and MCDD + butter (0.09 ± 0.1) groups, respectively, as compared to the control group (0.024 ± 0.007, P < 0.001). Most importantly, this increase was blunted by 30% in MCDD + olive oil group compared with MCDD group only (P < 0.004), by 37% in MCDD + olive oil group compared with MCDD + fish oil group (P < 0.001), and by 33% in MCDD + olive oil group compared with MCDD + butter group (P < 0.001, Figure 2). Hepatic cholesterol content increased by 15-fold in MCDD group (0.08 ± 0.02) compared with the control group (0.03 ± 0.01). In MCDD + fish oil group, this increment was blunted by 21% compared with MCDD group. There was a significant increase in lipid content in the liver of MCDD (1.9 ± 0.14), MCDD + olive oil (2.06 ± 0.4), MCDD + fish oil (1.9 ± 0.35), and MCDD + butter (1.98 ± 0.39) groups by 6.8-, 7.1-, 6.6- and 6.8-fold, respectively, as compared to the control group (0.28 ± 0.03, P < 0.01). Phospholipids content in the liver decreased in MCDD group (0.21 ± 0.04) by 2.6-fold compared with the control group (0.55 ± 0.04, P < 0.05). All phospholipids fractions (lysolecithin, sphingomyelin, lecithin, phosphatidyl inositol and phosphatidyl ethanolamine) were reduced in the liver of all treatment groups.
Figure 2.
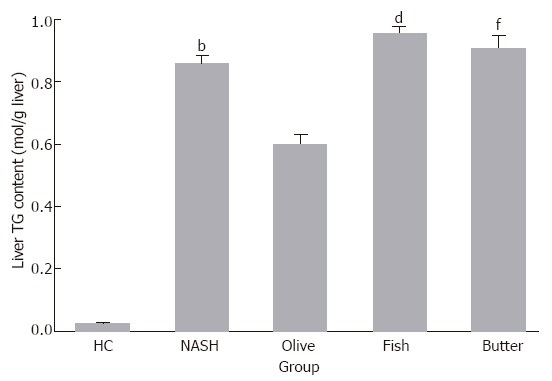
Hepatic triglycerides content (mol/g liver) in the different treatment groups. bP < 0.004, dP < 0.001, fP < 0.001 vs olive oil group.
Plasma lipids
Serum triglyceride levels (mmol/L) increased significantly in MCDD (1.05 ± 0.06), MCDD + olive oil (0.9 ± 0.07), MCDD + fish oil (1.02 ± 0.08), and MCDD + butter (0.97 ± 0.4) groups by 98%, 79%, 94%, and 85%, respectively, as compared to the control group (0.53 ± 0.09), (P < 0.05). This increment was significantly lowered (by -10%) in MCDD + olive oil group as compared with MCDD group (P < 0.02). Serum cholesterol levels (mmol/L) were not significantly different among the control (1.2 ± 0.37), MCDD (1.0 ± 0.1), and MCDD + olive oil (0.87 ± 0.2) groups, but increased by 33% and 22% in MCDD + fish oil (1.4 ± 0.2), and MCDD + butter (1.27 ± 0.2) groups, respectively, when compared with MCDD group. Plasma lipid content (3.4 ± 1.0) did not change in any treatment group. Phospholipids levels in serum decreased in MCDD group (0.63 ± 0.1) by 5.4-fold compared with the control group (3.4 ± 0.4), but no significant difference in phospholipids level was observed between among the treatment groups. All phospholipids fractions (lysolecithin, sphingomyelin, lecithin, phosphatidyl inositol and phosphatidyl ethanolamine) were reduced in plasma.
Hepatic pro-oxidant and anti-oxidants
MDA hepatic content (mol/g liver) increased significantly in MCDD (1.58 ± 0.8), MCDD + olive oil (3.3 ± 0.6), MCDD + fish oil (3.0 ± 0.4), and MCDD + butter (2.9 ± 0.36) groups in comparison to the control group (0.2 ± 0.1) by 7.4-, 15.3-, 14 and 13.7-fold, respectively (P < 0.05). Moreover, MDA hepatic contents in MCDD + olive oil, MCDD + fish oil, and MCDD + butter groups were higher in comparison to MCDD group by 108%, 91% and 87%, respectively (P < 0.004). Paraoxonase activity decreased significantly in the livers of all treatment groups. The most prominent decrease in paraoxonase activity was observed in MCDD + olive oil group (1.36 ± 0.28, -68%, P < 0.01, Figure 3). α-tocopherol content in the liver (mg/g liver) decreased in MCDD + olive oil group (0.26 ± 0.09) by 63% compared with the control group (0.7 ± 0.05, P < 0.01), by 61% in comparison with MCDD + fish oil (0.66 ± 0.1) group (P < 0.003) and by 64% compared with MCDD + butter group (0.7 ± 0.09, P < 0.002). However, no significant differences were found among the other groups.
Figure 3.
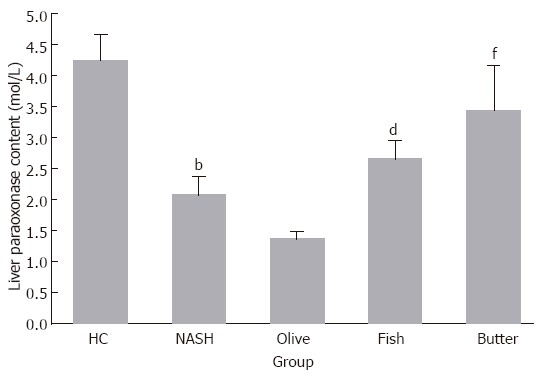
Hepatic paraoxonase contents (mol/L) in the different treatment groups. bP < 0.01, dP < 0.01, fP < 0.01 vs olive oil group.
Plasma pro-oxidant and anti-oxidants
There was no obvious difference between the control group (8.8 ± 1.6) and MCDD group (7.5 ± 1.5) in plasma concentrations of MDA. MCDD + olive oil (11.9 ± 3.2), MCDD + fish oil (11.3 ± 5.1) and MCDD + butter (10.1 ± 6.3) groups had significantly higher plasma concentration of MDA in comparison with MCDD group by 59%, 51% and 35%, respectively (P < 0.05). Plasma α-tocopherol level decreased in MCDD group (2.5 ± 0.87) by 57% in comparison with the control group (5.7 ± 0.78, P < 0.001). Plasma α-tocopherol levels were increased by 70% in MCDD + olive oil group (4.2 ± 0.5, P = 0.003), by 87.3% in MCDD + fish oil group (4.6 ± 0.56) and by 45% in MCDD + butter group (3.6 ± 0.5) as comparison to MCDD group (P < 0.05).
Hepatic composition of fatty acids
MCCD diet did not increase palmitic acid percentage in the livers of the rats, but did increase significantly the oleic acid percentage by 4.2-fold. MCDD enrichment by olive oil further increased the oleic acid percentage by 25% (P < 0.05). Palmitoleic acid had small contribution to fatty acid liver content and increased in all other treated rat groups on MCDD by 4-7-fold. Fish oil did not alter either the palmitic or palmitoleic acid percentages in the livers of MCDD + fish oil group as compared with MCDD group (Table 4).
Table 4.
Effect of MCDD alone or enriched with different fats on fatty acids percentage in the liver
| Fatty acids components | Control | MCDD | MCDD + olive oil | MCDD + fish oil | MCDD + butter |
1P |
2P |
| (Friedman test) | |||||||
| C14:0 Myristic | 0.2 ± 0.2 | 0.5 ± 0.1 | 0.3 ± 0.2 | 0.4 ± 0.2 | 0.4 ± 0.2 | 0.003 | 0.002 |
| C16:0 Palmitic | 19 ± 1 | 17 ± 1.1 | 15.3 ± 1.4ac | 15.5 ± 1.1 | 15.7 ± 2.3 | 0.001 | 0.006 |
| C16:1 Palmitoleic | 0.1 ± 0.2 | 0.7 ± 0.1 | 0.4 ± 0.2 | 0.7 ± 0.1 | 0.4 ± 0.2 | 0.007 | 0.02 |
| C18:0 Stearic | 21.8 ± 1.6 | 5.8 ± 0.6 | 5.7 ± 0.7ac | 6.2 ± 0.5 | 6.1 ± 0.9 | 0.002 | 0.01 |
| C18:1n9t Elaidic | 2.1 ± 0.1 | 1.5 ± 1.3 | 22.8 ± 1.3 | 17.8 ± 1.1 | 21.3 ± 1.1 | 0.001 | 0.006 |
| C18:1n9c Oleic | 4.9 ± 0.8 | 20.7 ± 0.7 | 25.9 ± 3.3ac | 2.0 ± 0.1 | 5.3 ± 8.7 | 0.001 | 0.006 |
| C18:2n6c Linolelaidic | 18.6 ± 1.8 | 32.4 ± 0.7 | 30.5 ± 1.7ac | 29.5 ± 2.1 | 31.5 ± 2.1 | 0.001 | 0.006 |
| C18:3n3 Linolenic | 0.5 ± 0.3 | 0.3 ± 0.1 | 0.2 ± 0.2c | 0.2 ± 0.2 | 0.3 ± 0.2 | 0.007 | 0.02 |
| C23:0 Tricosanoic | 22.0 ± 2.4 | 10.0 ± 1.1 | 7.8 ± 2.1ac | 0 | 0 | 0.003 | 0.02 |
| C20:4n6 Arachidonic | 0.1 ± 0.2 | 0 | 9.2 ± 0.6ac | 6.5 ± 0.6 | 9.7 ± 1.2 | 0.002 | 0.006 |
| C22:6n3 Docosahexaenoic | 4.5 ± 0.9 | 1.3 ± 0.5 | 1.2 ± 0.4 | 9.0 ± 1.2 | 1.1 ± 0.6 | 0.006 | 0.01 |
| C20:5n3 Eicosapentaenoic | 0 | 0.1 ± 0.1 | 0 | 0.9 ± 0.1 | 0 | 0.003 | 0.002 |
1Comparison between MCDD and treatment groups; 2comparison between the three treatment groups.
P < 0.02 vs MCDD + fish oil;
P < 0.02 vs MCDD + butter group.
Long-chain polyunsaturated fatty acids (PUFA) n6:n3 ratio increased in MCDD + olive oil, MCDD + fish oil and MCDD + butter fat groups by 345-, 30- and 397-fold, respectively in comparison with the control group. MCDD enriched with olive or fish oil or butter fat increased arachidonic acid percentages in the rat livers. This increment was 29% less in MCDD + fish oil group when compared with MCDD + olive oil (P < 0.05). C20:4n6/C18:2n6 ratios increased in MCDD + olive fat, MCDD + fish oil and MCDD + butter fat groups by 56-, 41- and 57-fold, respectively, in comparison with the control group (Table 4). C22:6n3 percentage in the rat livers decreased by 3.5-fold in MCDD group in comparison with the control group. This decrease was also noted in the rat groups on MCDD enriched with olive oil or with butter fats. On the contrary, C22:6n3 percentage in the liver increased by 2-fold in MCDD + fish oil group in comparison with the control group (Table 4). Enrichment of MCDD by olive oil, fish oil and butter fat increased elaidic acid percentage by 15.2-, 11.9- and 14.2-fold, respectively, compared to MCDD group. Fish oil blunted elaidic acid percentage increase in the rat livers by 22% in comparison with olive oil. C18:1n9c/C18:0 ratios increased in MCDD, MCDD + olive oil, MCDD + fish oil and MCDD + butter fat groups by 16-, 20-, 1-, and 4-fold, respectively, when compared to the control group (Table 4). Correlations between the levels of various fats, triglycerides, cholesterol, phospholipids, saturated and unsaturated fatty acids in the MCDD + olive oil group and the oxidant-antioxidant status are shown in Table 5. A strong correlation was noted between hepatic MDA levels and hepatic cholesterol (r = -0.8), phospholipids (r = -0.6), and palmitic acid (r = 0.6), as well as between hepatic alpha-tocopherol and linolelaidic acid (r = 0.97), elaidic acid (r = 0.9), oleic acid (r = -0.7), and arachidonic acid (r = -0.7). Hepatic paroxonase correlated well with tricosanoic acid (r = 0.9), stearic acid (r = -0.9), oleic acid (r = -0.8), and arachidonic acid (r = -0.7) (Table 5).
Table 5.
Correlations between hepatic lipid fractions and parameters of oxidant-antioxidant status in the MCDD + olive oil group
| Parameter | Malonyl dialdehyde (MDA) | Alpha-tocopherol | Paraoxonase |
| Triglycerides | r = -0.4 | r = 0.45 | r = -0.004 |
| Cholesterol | r = -0.8 | r = 0.05 | r = 0.3 |
| Phospholipids | r = -0.6 | r = -0.4 | r = -0.6 |
| Lipids | r = -0.2 | r = -0.4 | r = -0.2 |
| Palmitic | r = 0.6 | r = 0.1 | r = 0.1 |
| Stearic | r = -0.5 | r = -0.4 | r = -0.9 |
| Elaidic | r = 0.1 | r = 0.9 | r = 0.6 |
| Oleic | r = -0.2 | r = -0.7 | r = -0.8 |
| Linolelaidic | r = 0.2 | r = 0.97 | r = 0.66 |
| Tricosanoic | r = 0.5 | r = 0.4 | r = 0.9 |
| Arachidonic | r = -0.3 | r = -0.7 | r = -0.7 |
DISCUSSION
The results of this study clearly indicate that rats on MCDD diet supplemented with olive oil, but not fish oil or butter fat, store less triglycerides in their liver and prevent the occurrence of dietary-induced severe hepatic steatosis. Moreover, the results indicate also that olive oil diet provides the least antioxidant capability, whereas the fish oil and butter fat diets behave similarly with regard to antioxidant activities. As expected, these rats lost about 40% of body weight[25]. This weight loss is not in accordance with the clinical picture of NASH found in humans and may then results from the loss of subcutaneous fat[26]. Hepatic insulin resistance in the MCDD model of fatty liver is most likely caused by CYP2E1-induced oxidative stress and by elevated serum triglyceride levels[1]. In the current study, serum triglyceride levels were higher in all groups on MCDD (alone or enriched with fat) than in the control group, a finding which is also in accordance with the clinical NASH profile. Triglyceride content in the livers of the MCDD + olive oil group was lower than other groups on MCDD or MCDD enriched with saturated or polyunsaturated fatty acids. This may have been due to the inhibition of hepatic triacylglycerol synthesis[27], and stimulation of hepatic peroxisomal beta-oxidation produced by the olive oil diet[28]. Decrease in lipoprotein lipase and triacylglycerol lipase in the liver after olive and fish oil consumption has also been described and this may be an adaptive response to the low concentration of substrates (triacylglycerol) for these enzymes. The low levels of triglycerides in the liver after olive oil consumption may also be related to the reduced levels of plasma triglycerides found in our study. Moreover, monounsaturated fatty acids, present in olive oil, may improve insulin resistance, and enhance the release of triglyceride from the liver. This also decreases leak of free fatty acids from peripheral adipose tissue back to the liver and may explain the difference between the olive oil and butter groups.
In animal studies, saturated fat, present in butter, significantly increased insulin resistance, whereas long- and short-chain omega-3 fatty acids improved it, and the effects of monounsaturated and omega-6 polyunsaturated fatty acids ranged somewhere in between the two. In humans, shifting from a diet rich in saturated to one rich in monounsaturated fatty acid improved insulin sensitivity in healthy people[28]. Saturated fat [(myristic acid (14:0) and di-homo-γ-linolenic acid (18:3 n-6)] and the percentage of body fat had the strongest correlation with fasting insulin[29]. In normal subjects, consuming diet enriched with 10% elaidic (18:1 trans) acid induced insulin resistance acutely. In the present study, polyunsaturated fatty acids, present in fish oil failed to blunt the triglyceride accumulation in the liver. Fifty-nine percent of fatty acids stored in the liver of NASH patients arises from serum non-esterified fatty acids (NEFA), 26% from de novo lipogenesis and 15% from the diet[30]. Thus, the contribution of dietary olive oil to the fatty acid content in the liver appears small, suggesting the presence of other mechanisms preventing the accumulation of fatty acids in the liver under diet enrichment with olive oil. Eicosapentaenoic fatty acid administration decreased hepatic triglyceride content in ob/ob mice[31]. Polyunsaturated fatty acids can shift the energy balance from storage to consumption. Polyunsaturated fatty acids, particularly those of the n-3 family, play pivotal roles as “fuel partitioners” in that they direct fatty acids away from triglyceride storage and toward oxidation and they enhance glucose flux to glycogen[31]. In the present study, the MCDD model of NASH did not show the protective effect of polyunsaturated fatty acids on triglyceride accumulation in the liver. Whether this is due to enhanced hepatic lipogenesis or decreased β-oxidation remains unclear.
The rates of oxidation for the same amount of fatty acids differed among healthy persons, e.g., more than 37% of the dose of lauric acid (12:0) was oxidized compared with less than 15% of the dose of stearic acid (18:0)[32]. In dietary fat-enriched diets, the increased free fatty acids influx into the hepatocytes exceeds the liver capacity to assemble or/and export triglyceride-rich VLDL particles, causing hepatic steatosis. This increases the substrate for lipid peroxidation in the liver[33-35]. Polyunsaturated fatty acids are prone to spontaneous peroxidation, thus forming lipid hydroperoxides and reactive aldehydes[33,34]. At the same time, free fatty acid content in the hepatocytes up-regulates PPAR-α. This enhances mitochondrial long-chain fatty acid β-oxidation, initiation of omega-oxidation of fatty acids in the microsomes[36,37] and peroxisomal β-oxidation[38] resulting in generation of reactive oxygen species accumulation[39]. MCDD diet increased oxidative stress in the liver, but not in the plasma. Fat enrichment with olive oil, fish oil or butter fat added more oxidative stress in the liver and in the plasma. This increased oxidative stress in the liver did not impoverish α-tocopherol hepatic content in the rat groups fed diets enriched with fish oil or with butter. This, presumably, is due to the high content of α-tocopherol in these two food resources. Hepatic paraoxonase activity decreased in all rat groups on MCDD. This decreased activity may be the result of hepatic injury secondary to fat accumulation and increased oxidative stress. Plasma paraoxonase 1 activity is decreased in chronic liver disease in correlation with the degree of hepatic dysfunction[40]. Paraoxonase gene expression has been observed only in the liver[41]. This decrease in paraoxonase activity was more pronounced in the rats fed MCDD enriched with olive oil. The oxidative stress in the liver of this group was also higher than other groups on MCDD. The reasons for that are unclear. It appears that changes in the liver fatty acid composition, due mainly to n-3 lipids, may increase the activity of some anti-oxidant enzymes. A potential mechanism for the induction of hepatic anti-oxidants following butter feeding might be an increase in the expression of their genes; induction of the expression of anti-oxidant enzymes has been reported in circumstances where an increase in free radicals is produced, such as ageing[42].
Analysis of the fatty acid composition in the rat liver showed that MCDD increased palmitoleic and oleic acids percentage in the liver, but not palmitic acid. NASH patients showed increased overnight fasting serum levels of palmitic, palmitoleic and oleic acids[5], suggesting that the source of these fatty acids in the liver is from blood circulation and not from gastrointestinal tract. Hepatic long-chain PUFA n6/n3 ratios increased in MCDD + olive oil, MCDD + fish oil and MCDD + butter fat groups in comparison with the control group. Increased long-chain PUFA n-6/n-3 ratio in the liver of NASH patients may favor lipid synthesis over oxidation and secretion, thereby leading to steatosis[6]. This ratio increased less in MCDD + fish oil group, yet the MCDD + olive oil group had a lower fat content in the liver, making this ratio less effective in determining fatty acid content in the fatty liver in the MCDD rat model.
MCDD alone or enriched with olive, butter or fish fat increased arachidonic acid percentage in the rat livers, a possible marker of increased inflammatory state in the liver. Fish oil increased arachidonic acid less than olive or butter-supplemented groups, because PUFA can confer anti-inflammatory effect.
In the MCDD model, there was no reduction in C20:2n6/C18:2n6 ratio. In a previous study, NASH patients showed depletion of long-chain PUFA of the n-6 and n-3 series in the liver triacylglycerols[6]. In the present study, C22:6n3 was depleted in MCDD, MCDD + olive oil and MCDD + butter fat, but not in MCDD + fish oil due to dietary addition to the MCDD. MCDD enriched with olive oil, fish oil and butter fat increased hepatic elaidic acid percentage. Elaidic acid leads to defective PUFA desaturation by inhibiting desaturase and subsequently to depletion of hepatic long-chain PUFA[6], but in the present study, C22:6n3 was depleted in MCDD group without a change in hepatic content percentage of elaidic acid. C22:6n3 depletion in MCDD, MCDD + olive oil and MCDD + butter fat may be due to inadequate intake of precursors, such as 18:3 n-3 and a higher peroxidation of long-chain PUFA due to oxidative stress[6].
Although the MCDD model of fatty liver is the most commonly used in experimental studies, one limitation of our study is that MCDD model is not the ideal model for studying insulin resistance in humans and an extensive number of additional experiments would be required to be performed in order to derive definitive data in insulin-resistant models[26]. Another concern is that the rats fed MCDD for nine weeks did not develop the same phenotype as previous reports of MCDD-induced NASH in mice (as short as 10 d of MCDD). The phenotype described here largely consisted of marked steatosis with mild inflammation and mild fibrosis[43].
Increased MDA level, and reduced serum paraoxonase 1 (PON1) activity reflect increased oxidative damage in rats with steatohepatitis. Reactive oxygen species, which are formed due to increased microsomal, peroxisomal and mitochondrial oxidation, trigger lipid peroxidation; this in turn causes cell death and release of MDA[1]. It also has been shown that TNF-α down-regulates mRNA expression of paraoxonase in Hep G2 cells[44]. This cytokine-mediated reduction of paraoxonase production by the liver might be responsible for the decreased paraoxonase activity in steatohepatitis. On the other hand, decreased paraoxonase activity could be accepted as another evidence of increased lipid peroxidation, since it was shown that a decrease in liver microsomal paraoxonase activity is an early biochemical change related to lipid peroxidation and liver injury observed in the rats with CCl4-induced cirrhosis[45].
In conclusion, olive oil decreases the accumulation of triglycerides in the liver of rats with NASH, thereby suggesting that olive oil should be included in the diet of patients with NASH and NAFLD.
Footnotes
S- Editor Wang GP L- Editor Kumar M E- Editor Lu W
References
- 1.Day CP. Pathogenesis of steatohepatitis. Best Pract Res Clin Gastroenterol. 2002;16:663–678. doi: 10.1053/bega.2002.0333. [DOI] [PubMed] [Google Scholar]
- 2.Cassader M, Gambino R, Musso G, Depetris N, Mecca F, Cavallo-Perin P, Pacini G, Rizzetto M, Pagano G. Postprandial triglyceride-rich lipoprotein metabolism and insulin sensitivity in nonalcoholic steatohepatitis patients. Lipids. 2001;36:1117–1124. doi: 10.1007/s11745-001-0822-5. [DOI] [PubMed] [Google Scholar]
- 3.Cooper AD. Hepatic uptake of chylomicron remnants. J Lipid Res. 1997;38:2173–2192. [PubMed] [Google Scholar]
- 4.Diraison F, Moulin P, Beylot M. Contribution of hepatic de novo lipogenesis and reesterification of plasma non esterified fatty acids to plasma triglyceride synthesis during non-alcoholic fatty liver disease. Diabetes Metab. 2003;29:478–485. doi: 10.1016/s1262-3636(07)70061-7. [DOI] [PubMed] [Google Scholar]
- 5.de Almeida IT, Cortez-Pinto H, Fidalgo G, Rodrigues D, Camilo ME. Plasma total and free fatty acids composition in human non-alcoholic steatohepatitis. Clin Nutr. 2002;21:219–223. doi: 10.1054/clnu.2001.0529. [DOI] [PubMed] [Google Scholar]
- 6.Araya J, Rodrigo R, Videla LA, Thielemann L, Orellana M, Pettinelli P, Poniachik J. Increase in long-chain polyunsaturated fatty acid n - 6/n - 3 ratio in relation to hepatic steatosis in patients with non-alcoholic fatty liver disease. Clin Sci (Lond) 2004;106:635–643. doi: 10.1042/CS20030326. [DOI] [PubMed] [Google Scholar]
- 7.Fan JG, Zhong L, Xu ZJ, Tia LY, Ding XD, Li MS, Wang GL. Effects of low-calorie diet on steatohepatitis in rats with obesity and hyperlipidemia. World J Gastroenterol. 2003;9:2045–2049. doi: 10.3748/wjg.v9.i9.2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Clarke SD, Jump DB. Dietary polyunsaturated fatty acid regulation of gene transcription. Annu Rev Nutr. 1994;14:83–98. doi: 10.1146/annurev.nu.14.070194.000503. [DOI] [PubMed] [Google Scholar]
- 9.Sekiya M, Yahagi N, Matsuzaka T, Najima Y, Nakakuki M, Nagai R, Ishibashi S, Osuga J, Yamada N, Shimano H. Polyunsaturated fatty acids ameliorate hepatic steatosis in obese mice by SREBP-1 suppression. Hepatology. 2003;38:1529–1539. doi: 10.1016/j.hep.2003.09.028. [DOI] [PubMed] [Google Scholar]
- 10.Musso G, Gambino R, De Michieli F, Cassader M, Rizzetto M, Durazzo M, Fagà E, Silli B, Pagano G. Dietary habits and their relations to insulin resistance and postprandial lipemia in nonalcoholic steatohepatitis. Hepatology. 2003;37:909–916. doi: 10.1053/jhep.2003.50132. [DOI] [PubMed] [Google Scholar]
- 11.Kliewer SA, Sundseth SS, Jones SA, Brown PJ, Wisely GB, Koble CS, Devchand P, Wahli W, Willson TM, Lenhard JM, et al. Fatty acids and eicosanoids regulate gene expression through direct interactions with peroxisome proliferator-activated receptors alpha and gamma. Proc Natl Acad Sci USA. 1997;94:4318–4323. doi: 10.1073/pnas.94.9.4318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Grundy SM. Comparison of monounsaturated fatty acids and carbohydrates for lowering plasma cholesterol. N Engl J Med. 1986;314:745–748. doi: 10.1056/NEJM198603203141204. [DOI] [PubMed] [Google Scholar]
- 13.Koteish A, Diehl AM. Animal models of steatosis. Semin Liver Dis. 2001;21:89–104. doi: 10.1055/s-2001-12932. [DOI] [PubMed] [Google Scholar]
- 14.Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 1957;226:497–509. [PubMed] [Google Scholar]
- 15.Gottfried SP, Rosenberg B. Improved manual spectrophotometric procedure for determination of serum triglycerides. Clin Chem. 1973;19:1077–1078. [PubMed] [Google Scholar]
- 16.Taylor RP, Broccoli AV, Grisham CM. Enzymatic and colorimetric determination of total serum cholesterol. An undergraduate biochemistry laboratory experiment. J Chem Educ. 1978;55:63–64. doi: 10.1021/ed055p63. [DOI] [PubMed] [Google Scholar]
- 17.Barnes H, Blackstock J. Estimation of lipids in marine animals and tissues: detailed investigation of the sulfophosphovanillin method for 'total' lipid. J exp Mar Biol Ecol. 1973;12:103–118. [Google Scholar]
- 18.Bartlett GR. Phosphorus assay in column chromatography. J Biol Chem. 1959;234:466–468. [PubMed] [Google Scholar]
- 19.Kupke IR, Zeugner S. Quantitative high-performance thin-layer chromatography of lipids in plasma and liver homogenates after direct application of 0.5-microliter samples to the silica-gel layer. J Chromatogr. 1978;146:261–271. doi: 10.1016/s0378-4347(00)81892-7. [DOI] [PubMed] [Google Scholar]
- 20.Gan KN, Smolen A, Eckerson HW, La Du BN. Purification of human serum paraoxonase/arylesterase. Evidence for one esterase catalyzing both activities. Drug Metab Dispos. 1991;19:100–106. [PubMed] [Google Scholar]
- 21.Gowenlock HA, McMurray JR. Vitamins. In: Gowenlock HA, McMurray JR, McLauchlan DM, editors. Varley's Practical Clinical Biochemistry. 6th ed. London: Heinmann Medical Books; 1988. pp. 894–930. [Google Scholar]
- 22.Yagi K. Lipid peroxides and human diseases. Chem Phys Lipids. 1987;45:337–351. doi: 10.1016/0009-3084(87)90071-5. [DOI] [PubMed] [Google Scholar]
- 23.Parks EJ, Krauss RM, Christiansen MP, Neese RA, Hellerstein MK. Effects of a low-fat, high-carbohydrate diet on VLDL-triglyceride assembly, production, and clearance. J Clin Invest. 1999;104:1087–1096. doi: 10.1172/JCI6572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Brunt EM, Janney CG, Di Bisceglie AM, Neuschwander-Tetri BA, Bacon BR. Nonalcoholic steatohepatitis: a proposal for grading and staging the histological lesions. Am J Gastroenterol. 1999;94:2467–2474. doi: 10.1111/j.1572-0241.1999.01377.x. [DOI] [PubMed] [Google Scholar]
- 25.Farrell G. Animal models of steatohepatitis. In: Farrell G, George J, de la M Hall P, McCullough AJ, editors. Fatty liver disease: NASH and related disorders. Massachusetts: Blackwell; 2005. pp. 91–108. [Google Scholar]
- 26.Rinella ME, Green RM. The methionine-choline deficient dietary model of steatohepatitis does not exhibit insulin resistance. J Hepatol. 2004;40:47–51. doi: 10.1016/j.jhep.2003.09.020. [DOI] [PubMed] [Google Scholar]
- 27.Wong SH, Nestel PJ, Trimble RP, Storer GB, Illman RJ, Topping DL. The adaptive effects of dietary fish and safflower oil on lipid and lipoprotein metabolism in perfused rat liver. Biochim Biophys Acta. 1984;792:103–109. doi: 10.1016/0005-2760(84)90209-1. [DOI] [PubMed] [Google Scholar]
- 28.Rivellese AA, De Natale C, Lilli S. Type of dietary fat and insulin resistance. Ann N Y Acad Sci. 2002;967:329–335. doi: 10.1111/j.1749-6632.2002.tb04288.x. [DOI] [PubMed] [Google Scholar]
- 29.Lovejoy JC, Champagne CM, Smith SR, DeLany JP, Bray GA, Lefevre M, Denkins YM, Rood JC. Relationship of dietary fat and serum cholesterol ester and phospholipid fatty acids to markers of insulin resistance in men and women with a range of glucose tolerance. Metabolism. 2001;50:86–92. doi: 10.1053/meta.2001.19440. [DOI] [PubMed] [Google Scholar]
- 30.Donnelly KL, Smith CI, Schwarzenberg SJ, Jessurun J, Boldt MD, Parks EJ. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J Clin Invest. 2005;115:1343–1351. doi: 10.1172/JCI23621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Clarke SD. Nonalcoholic steatosis and steatohepatitis. I. Molecular mechanism for polyunsaturated fatty acid regulation of gene transcription. Am J Physiol Gastrointest Liver Physiol. 2001;281:G865–G869. doi: 10.1152/ajpgi.2001.281.4.G865. [DOI] [PubMed] [Google Scholar]
- 32.DeLany JP, Windhauser MM, Champagne CM, Bray GA. Differential oxidation of individual dietary fatty acids in humans. Am J Clin Nutr. 2000;72:905–911. doi: 10.1093/ajcn/72.4.905. [DOI] [PubMed] [Google Scholar]
- 33.Mylonas C, Kouretas D. Lipid peroxidation and tissue damage. In Vivo. 1999;13:295–309. [PubMed] [Google Scholar]
- 34.Jaeschke H, Gores GJ, Cederbaum AI, Hinson JA, Pessayre D, Lemasters JJ. Mechanisms of hepatotoxicity. Toxicol Sci. 2002;65:166–176. doi: 10.1093/toxsci/65.2.166. [DOI] [PubMed] [Google Scholar]
- 35.Tang DG, La E, Kern J, Kehrer JP. Fatty acid oxidation and signaling in apoptosis. Biol Chem. 2002;383:425–442. doi: 10.1515/BC.2002.046. [DOI] [PubMed] [Google Scholar]
- 36.Mortensen PB. Formation and degradation of dicarboxylic acids in relation to alterations in fatty acid oxidation in rats. Biochim Biophys Acta. 1992;1124:71–79. doi: 10.1016/0005-2760(92)90128-i. [DOI] [PubMed] [Google Scholar]
- 37.Kaikaus RM, Chan WK, Lysenko N, Ray R, Ortiz de Montellano PR, Bass NM. Induction of peroxisomal fatty acid beta-oxidation and liver fatty acid-binding protein by peroxisome proliferators. Mediation via the cytochrome P-450IVA1 omega-hydroxylase pathway. J Biol Chem. 1993;268:9593–9603. [PubMed] [Google Scholar]
- 38.Kliewer SA, Xu HE, Lambert MH, Willson TM. Peroxisome proliferator-activated receptors: from genes to physiology. Recent Prog Horm Res. 2001;56:239–263. doi: 10.1210/rp.56.1.239. [DOI] [PubMed] [Google Scholar]
- 39.Bocher V, Pineda-Torra I, Fruchart JC, Staels B. PPARs: transcription factors controlling lipid and lipoprotein metabolism. Ann N Y Acad Sci. 2002;967:7–18. doi: 10.1111/j.1749-6632.2002.tb04258.x. [DOI] [PubMed] [Google Scholar]
- 40.Ferré N, Camps J, Prats E, Vilella E, Paul A, Figuera L, Joven J. Serum paraoxonase activity: a new additional test for the improved evaluation of chronic liver damage. Clin Chem. 2002;48:261–268. [PubMed] [Google Scholar]
- 41.Leviev I, Negro F, James RW. Two alleles of the human paraoxonase gene produce different amounts of mRNA. An explanation for differences in serum concentrations of paraoxonase associated with the (Leu-Met54) polymorphism. Arterioscler Thromb Vasc Biol. 1997;17:2935–2939. doi: 10.1161/01.atv.17.11.2935. [DOI] [PubMed] [Google Scholar]
- 42.de Haan JB, Newman JD, Kola I. Cu/Zn superoxide dismutase mRNA and enzyme activity, and susceptibility to lipid peroxidation, increases with aging in murine brains. Brain Res Mol Brain Res. 1992;13:179–187. doi: 10.1016/0169-328x(92)90025-7. [DOI] [PubMed] [Google Scholar]
- 43.George J, Pera N, Phung N, Leclercq I, Yun Hou J, Farrell G. Lipid peroxidation, stellate cell activation and hepatic fibrogenesis in a rat model of chronic steatohepatitis. J Hepatol. 2003;39:756–764. doi: 10.1016/s0168-8278(03)00376-3. [DOI] [PubMed] [Google Scholar]
- 44.Kumon Y, Nakauchi Y, Suehiro T, Shiinoki T, Tanimoto N, Inoue M, Nakamura T, Hashimoto K, Sipe JD. Proinflammatory cytokines but not acute phase serum amyloid A or C-reactive protein, downregulate paraoxonase 1 (PON1) expression by HepG2 cells. Amyloid. 2002;9:160–164. doi: 10.3109/13506120209114817. [DOI] [PubMed] [Google Scholar]
- 45.Ferré N, Camps J, Cabré M, Paul A, Joven J. Hepatic paraoxonase activity alterations and free radical production in rats with experimental cirrhosis. Metabolism. 2001;50:997–1000. doi: 10.1053/meta.2001.25589. [DOI] [PubMed] [Google Scholar]