

Abstract
Background.
Antiretroviral therapy has reduced the incidence of adverse events and early mortality in HIV-infected persons. Despite these benefits, important comorbidities that increase with age (eg, diabetes, cardiovascular disease, cancer, liver disease, and neurocognitive impairment) are more prevalent in HIV-infected persons than in HIV-uninfected persons at every age, and geriatric syndromes such as falls and frailty occur earlier in HIV-infected persons. This raises a critical research question: Does HIV accelerate aging through pathways and mechanisms common to the aging process or is HIV simply an additional risk factor for a wide number of chronic conditions, thus accentuating aging?
Methods.
Extensive literature review.
Results.
The purpose of this review is to briefly outline the evidence that age-related clinical syndromes are exacerbated by HIV, examine the ways in which HIV is similar, and dissimilar from natural aging, and assess the validity of HIV as a model of premature aging. Specific biomarkers of aging are limited in HIV-infected hosts and impacted by antiretroviral therapy, and a high rate of modifiable life style confounders (eg, smoking, substance abuse, alcohol) and coinfections (eg, hepatitis) in HIV-infected participants.
Conclusions.
There is a need for validated biomarkers of aging in the context of HIV. Despite these differences, welldesigned studies of HIV-infected participants are likely to provide new opportunities to better understand the mechanisms that lead to aging and age-related diseases.
Key Words: HIV, AIDS, Multimorbidity, Immunosenescence, Biomarkers of aging
Major reductions in HIV-associated mortality have occurred as a result of the widespread use of antiretroviral therapy (ART). Median life expectancy for HIV-infected individuals treated with ART increased from 55 years of age in 1996 to 70 years by 2005, and about half of the HIV-infected persons in the United States, will be >50 years of age by 2015 (1,2). Even in sub-Saharan Africa 14.3% of HIV-infected adults are older than 50 years of age (3), and that number is expected to triple within the next 30 years in South Africa (4) with similar trends in Asia (5).
These life expectancy gains are largely due to the prevention of AIDS-related events after ART administration, but they are also due to a decrease in diseases and conditions that are not generally considered to be “AIDS-related” (6,7). These chronic illnesses, termed HIV-associated non-AIDS conditions, are frequently chronic conditions that are linked with advancing age and chronic inflammation (cardiovascular disease, cancers, liver disease, renal disease, and neurocognitive decline). Accumulated comorbidities in multiple systems lead to geriatric syndromes—multimorbidity, polypharmacy, frailty, and even falls an average of 15–20 years earlier than in HIV-uninfected participants (6–19).
A critical research question is whether HIV is accelerating aging itself through pathways and mechanisms common to the aging process (19,20), or, alternatively, HIV may simply be an additional risk factor for a wide number of chronic conditions thus accentuating the prevalence of disease at every age (Figure 1) (7,19). Although this may seem to be a semantic difference, it is critical to defining approaches that mitigate the sequelae of aging with HIV infection, and perhaps aging in uninfected persons.
Figure 1.
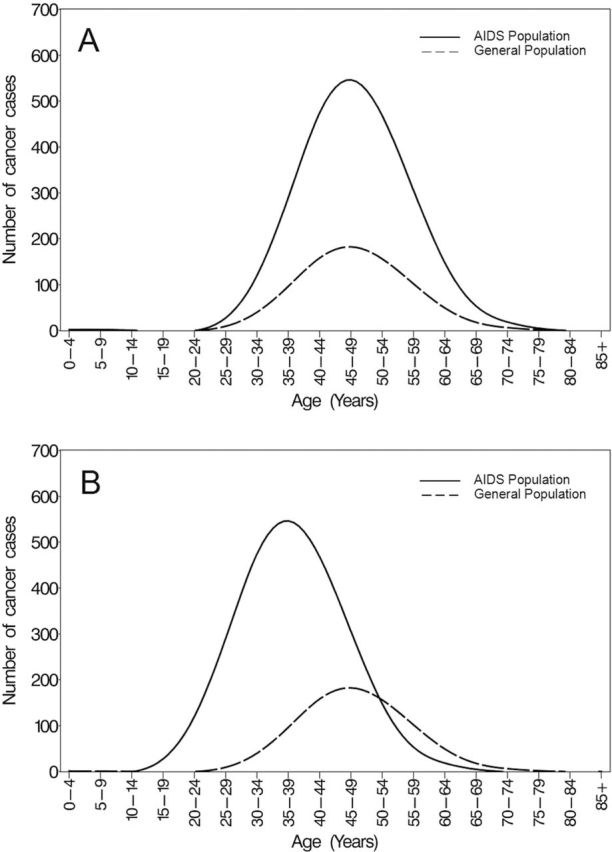
Hypothetical age-at-diagnosis distributions of cancer in the AIDS and general populations. (A) Accentuated: cancer occurs at the same ages but more often among HIV-infected participants than among HIV-uninfected comparators. (B) Accelerated and accentuated: cancer occurs earlier among HIV-infected participants compared with HIV-uninfected comparators and there are more cancer events. Reprinted from reference (38) with permission.
To resolve this question, it will be important to account for differential exposure to risk factors (eg, smoking, alcohol, non-HIV sexually transmitted diseases) between HIV-infected and HIV-uninfected populations that are likely to result in residual confounding when assessing associations of HIV with increased risk of age-related illness. This situation is further complicated by concern that toxicities of ART, not HIV itself, may contribute to aging and age-related illness. However, in the Strategies for Management of Antiretroviral Therapy study that examined an aggressive treatment strategy versus a drug-conserving strategy, deaths from HIV-associated non-AIDS conditions were much more common in the drug conservation group including higher rates of major cardiovascular, renal, and liver disease (10–12), suggesting uncontrolled HIV is more detrimental than ART in this regard. Further, many age-related illnesses are driven by HIV itself, though drug toxicity may play a role in specific organ systems. Because it is unethical to withhold ART once CD4 counts fall below specific thresholds, it is nearly impossible to separate HIV infection from ART duration and potential toxicities except when explicit adverse events or mechanisms have been linked to individual drugs. Further insight regarding morbidity due to HIV itself versus the drugs used to treat HIV may be provided by ongoing studies (19,21–24).
The purpose of this review is to examine the ways in which HIV is similar, and dissimilar, from natural aging (Table 1), and to assess the validity of HIV as a model of premature aging. We have divided the text into three parts: (i) a very brief review of selected clinical illnesses and age-related syndromes in HIV-infected populations; (ii) common pathways thought to lead to chronic illness (eg, immune activation and senescence, inflammation, hypercoagulation, and body composition) both in those with and without HIV, and (iii) proposal of novel biomarkers of aging (BoA) minimally affected by HIV viral replication or ART that may constitute new biomarkers to measure aging in complex populations such as those with HIV.
Table 1.
Similarities and Dissimilarities Between HIV Disease and Normal Aging
| Similarities Between HIV Infection and Normal Aging | Differences Between HIV Infection and Normal Aging |
|---|---|
| Syndromes | |
| Frailty phenotype associated with obesity | Increased substance abuse, smoking, mental health issues, and social isolation in HIV |
| Cardiovascular pathology–vascular stiffness/calcifications, diastolic dysfunction | HIV increases the risk of many cancers, but does not increase the risk of: the two most influenced by age—breast and prostate cancer |
| Multimorbidity and polypharmacy | |
| Bone loss and fragility fractures | |
| Neurocognitive decline—prevalent mild cognitive impairment and increased dementia | |
| Prevalent chronic renal insufficiency | |
| Mechanisms | |
| Thymic involution—loss of naive T-cell production | HIV infection typically absent in aging—HIV replication not a source of immune activation or inflammation in aging |
| Immune senescent phenotype—accumulation of end-stage (short telomeres, CD28−) effector T cells with senescence-associated secretory phenotype | Antiretroviral therapy confounds aging in the presence of HIV |
| Prevalent low-level inflammation | |
| Sarcopenia–muscle loss/fatty infiltration in muscle, reduced power and contractile force | |
Age-Related Conditions in HIV-Infected Persons
The hypothesis of premature aging in HIV has received criticism primarily due to limitations in characterization of participants, in particular, the possibility of differential exposure to potential risk factors between HIV-infected and HIV-uninfected populations (20,25,26). The differential exposures to risk factors between HIV-infected and HIV-uninfected populations and residual confounding could result in an apparent increased risk of age-related outcomes. Previous studies have also compared HIV-infected and uninfected populations from different cohorts and do not always take into consideration comorbid conditions that may influence outcome measures. Comparing HIV-infected with uninfected persons who have a lower burden of non-HIV–related conditions makes it difficult to determine whether elevated biomarkers are driven by HIV infection or by the increased comorbidity burden among the HIV-infected persons (27).
A full review of the increased incidence of specific clinical illnesses in those with HIV is beyond the scope of this review, but a few examples are provided for context. For example, cardiovascular risk factors and rates of acute coronary syndromes are markedly increased in HIV-infected versus age-matched control participants (28), and coronary artery “age” assessed by coronary artery calcium score is accelerated on average by about 15 years (29). Higher levels of C-reactive protein, interleukin-6 (IL-6), and D-dimer have been shown to be significantly associated with an increased risk of all-cause mortality in HIV-infected individuals not on ART, and much of this is cardiovascular mortality (12). Specific ART drugs also may be causally associated with early heart disease, even after controlling for age and traditional cardiovascular risk factors (11). Further, lipodystrophy, a syndrome of metabolic abnormalities (altered body fat, hyperlipidemia, and insulin resistance) is common in HIV-infected patients receiving ART (30). The redistribution of fat mass and progression to metabolic syndrome (12/100 patient-years) typically occurs within 3 years after the initiation of ART (31) when weight gain is greatest, substantial increasing cardiovascular disease risk. Enhanced cardiovascular aging is not limited to coronary artery disease. Left ventricular diastolic dysfunction and increased vascular stiffness (32–34) are more common in HIV-infected participants versus uninfected, age-matched controls even after controlling for hypertension and other risk factors.
Other organ system illnesses are also more prevalent in those with HIV including kidney disease (35), low bone mineral density and fractures (36), and a number of non-AIDS–defining cancers (37). Interestingly, although cancer is more common in those with HIV, it may be only minimally “accelerated” with regard to age at diagnosis (38). It is important to note that some age-related cancers are not increased in those with HIV; there is no increased risk of breast cancer in women or prostate cancer in men (38).
Geriatric Syndromes
Multimorbidity is common, occurring in up to 65% of U.S. HIV-infected participants, occurs at an earlier age than in HIV-uninfected participants, and obesity is associated with a higher likelihood of multimorbidity (9). The best-validated index of multimorbidity in HIV patients, the veterans aging cohort study index, integrates HIV-specific parameters and evidence of impaired end-organ reserve (39), and is predictive of the risk of death, hospitalization, intensive care unit hospitalization, frailty, and markers of inflammation, coagulation, and monocyte activation (27).
Frailty is also prevalent in HIV-infected patients. The most widely published definition of frailty in HIV-infected participants is the Fried frailty phenotype (40) generally described as the presence of at least three of the following five criteria: unintentional weight loss; exhaustion; low level of physical activity; slow motor performance; and weakness (7,41). Early in the HIV epidemic, frailty was more common in those with advanced HIV disease, wasting and a low body mass index, but with the widespread use of ART, more recent studies suggest frailty in HIV is now associated with central adiposity, sarcopenia, and the density of muscle fat (42) mirroring non-HIV–infected seniors. Frail HIV participants have significantly higher serum IL-6 levels and significantly lower hemoglobin and/or hematocrit than non-frail participants (43).
As in HIV-uninfected participants, age is a major predictor of cognitive impairment in HIV-infected participants. In the pre-ART era of HIV, dementia was three times (19%) more prevalent among patients older than 75 years compared with patients younger than 35 years (6%) (44). Older individuals with viral replication within the central nervous system (detectable HIV in cerebrospinal fluid) have twice the prevalence of neuropsychological impairment and there is an interaction of viral burden with age (45). Advanced AIDS patients on ART typically experience stable or improved cognition (performance on neuropsychological tests 3–5 y after initiating ART), but remain more impaired than the general population (46).
Immunological and Inflammatory Changes in HIV Infection—Parallels With Aging
Inflammation and activation of coagulation pathways are central to the pathophysiology leading to morbidity and mortality in HIV as demonstrated by data from the Strategies for Management of Antiretroviral Therapy study, which showed that IL-6 and D-dimer were strongly related to non-AIDS–defining comorbidities and all-cause mortality in patients on ART (12). Although CD4+ T-cell count and HIV plasma viral load are routinely used to monitor clinical response to ART, ongoing immune activation (ie, expression of activation markers on the surface of T cells) more accurately predicts HIV disease progression to AIDS and death (47,48). Immune activation is a hallmark of chronic HIV infection and may be mediated by several mechanisms: increased pathogen burden as a result of impaired immunity, chronic viral replication of HIV and other viruses (eg, cytomegalovirus [CMV], hepatitis viruses), and microbial translocation of bacterial products across damaged mucosa (eg, in the gut). These factors may also drive chronic immune activation in HIV-uninfected seniors (49).
Immune activation (ie, T-cell activation) seems to be of particular importance in ART-naive patients, and in those with undetectable plasma HIV RNA levels in the absence of ART (aka “elite controllers”) (50,51). However, in ART-treated patients it appears that inflammation (ie, macrophage activation and cytokine secretion) may play a more important role in the development of morbidity and mortality. Additionally, various antivirals have been shown to induce inflammatory signals and may contribute to inflammation (52). These relationships are further explored in the following paragraphs.
T-Cell Activation and Senescence in Chronic HIV Infection
Many of the T-cell abnormalities associated with aging are similar to those observed in untreated HIV infection (7,49,53) (Table 1). Immunosenescence is collectively defined as the functional limitation of immunity resulting from age-associated changes in a variety of cells related to both innate and adaptive immunity, and an imbalance between the two arms (54).
The immune system in the elderly participants is characterized by progressive enrichment of terminally differentiated T cells (primarily CD8+ effector cells), a reduction in T-cell renewal manifested by a decline in naive T cells and a lower ratio of CD4+ to CD8+ cells, increased T-cell activation, and increased levels of several inflammatory markers. This translates into a state of immune senescence characterized by low-level, constitutive inflammation, but an inability to rapidly mount adequate immune activation upon challenge (either by a vaccine or infecting organism) (49). The expansion of terminally differentiated T cells is likely due to chronic, repetitive stimulation, particularly by endogenous viruses that establish chronic infection (eg, CMV, HIV, hepatitis viruses). Such cells can be identified by the loss of cell surface CD28 expression and gain of CD57 expression (reviewed in reference 55). CD28 is a costimulatory molecule on T cells; binding to its ligand, CD80, on antigen-presenting cells leads to IL-2 production, telomerase activation, and “survival cytokine signals” for nearby T cells. Loss of CD28 occurs on CD4+ cells and is associated with reduced T-cell–driven B-cell proliferation and antibody production. However, CD28− cells rarely comprise >10% of all CD4+ T cells even in far advanced age. In contrast, CD28− cells accumulate to tremendous levels in the CD8+ T-cell compartment, and in extreme old age, these CD8+/CD28− T cells can account for >70% of all circulating T cells (56). Furthermore, elderly CMV-seropositive adults have lower CD4/CD8 ratios, lower naive T-cell numbers, and higher frequencies of the CD8+CD28− phenotype compared with CMV-seronegative adults (57). Loss of CD28 on T cells is a key indicator of immune senescence (49), and CD8+/CD28− T-cell percentage is highly correlated with impaired vaccine responses (58). Further, CD28−/CD8+ T cells acquire a “senescence-associated secretory phenotype” (59,60) in which excessive, constitutive secretion of proinflammatory cytokines occurs. Although any cell type can acquire the senescence-associated secretory phenotype, senescent T cells are able to produce extremely high levels of cytokine (61,62).
Continuous stimulation of the immune system by HIV, and perhaps other viral coinfections (CMV, hepatitis C) greatly accelerates the progression of immune changes to that of an immune senescent phenotype. Depletion of T-cell precursors also leads to a progressive loss of the naive T cells, further promoting an imbalance of T-cell phenotypes similar to that observed in the elderly participants (49). ART-treated patients with a fully suppressed viral load (median age 56 years, <50 HIV-1 RNA copies/mL, median CD4 count 724 cells/mm3) have frequencies of senescent CD8+ T cells (CD57+CD28− phenotype) similar to HIV-negative individuals decades older (median age 88 years) (63). There is also a strong association between CMV seropositivity and an increased number of CD28− CD4 or CD8 T cells (64,65)—see section on CMV below.
Immunosenescence is associated with clinical outcomes in HIV infection. In a large cohort of HIV-infected adults initiating ART, a low pre-ART CD4 nadir was associated with higher percentages of CD28−CD4+ and CD28−CD8+ cells. Effective ART reduced the proportion of CD28− T cells. However, levels remained abnormally high and closer to levels in older HIV-uninfected individuals (66). Another study suggested that untreated HIV-1 infection and progression is associated with premature aging of memory CD8+ and CD4+ T cells and reduced levels of naive CD4+ T cells (67). T-cell activation and senescence also predict subclinical carotid artery disease in HIV-infected women. Compared with HIV-uninfected women, frequencies of CD4+CD38+HLA-DR+, CD8+CD38+HLA-DR+, and CD8+CD28−CD57+ T cells were higher among HIV-infected women, including those who achieved viral suppression while receiving ART. Among HIV-infected patients, higher frequencies of T cells expressing markers of activation (CD38, HLA-DR) and senescence (presence of CD57, absence of CD28) were associated with greater arterial wall thickness (68). Markers of T-cell activation (CD4+CD38+HLA-DR+) are also associated with increased vascular stiffness among HIV-infected women (69).
Microbial Translocation as a Model of Immune Activation
Microbial translocation of bacterial products across damaged mucosal surfaces, primarily in the gut, may contribute to immune activation. Gastrointestinal tract mucosa is a major site of T-cell destruction in the first few weeks after HIV infection, and profound CD4+ T-cell depletion is present at all stages of HIV disease (70). This leads to poor epithelial surface integrity, facilitating the translocation of microbial products such as lipopolysaccharides across the “leaky” mucosa into the systemic circulation (71). This early loss of gut mucosal integrity provides a source for continuous activation of innate immune cells such as monocytes/macrophages and dendritic cells. Binding of various microbial products such as lipopolysaccharides to Toll-like receptors on these cells and its related signaling molecule CD14 induces the release of proinflammatory cytokines and further activation of the immune system (72).
Age-related increases in innate immune biomarkers such as CXCL10 and soluble CD163 may occur independently of senescent changes in CD8+ T lymphocytes in those with HIV (73). HIV infection in women confers the equivalent of a 10–14 year increase in the levels of innate immune aging markers (73). Young HIV-infected men exhibit age-related changes to the monocyte phenotype and levels of innate immune activation that resemble those observed in HIV-uninfected individuals aged approximately 30 years older (74). Higher plasma levels of soluble CD14 and progression of atherosclerosis have been observed in HIV-infected adults (75), and as a predictor of the development of hypertension (76). Soluble CD14 has also been demonstrated to independently predict all-cause mortality in HIV-infected individuals (77). Interestingly, HIV-1 elite controllers spontaneously maintain suppressed levels of viremia, but exhibit significant persistent T-cell activation and inflammation (50). Soluble CD163 was markedly increased in elite controllers compared with ART-treated patients and HIV-negative controls. The prevalence of atherosclerosis was also increased in elite controllers compared with uninfected control participants (78). This demonstrates that despite excellent virologic and immunologic control and no confounding ART, HIV infection confers significant innate immune activation and coronary atherosclerosis risk.
CMV Coinfection: An Important Additive Factor in Immune Activation?
CMV coinfection is very common, 90%–100%, in HIV-infected populations (79) and chronic CMV infection is associated with accelerated immunosenescence and mortality among the healthy elderly populations suggesting a major role for this virus in aging (80). Long-term successfully treated HIV-infected individuals have high levels of CMV-specific effector cells, similar to that observed in the elderly participants, but occurring about 40 years earlier (81). The possible role of CMV in cardiovascular risk has also been investigated, and HIV-infected women with increased CMV immunoglobulin G levels were more likely to have carotid artery stiffness compared with uninfected controls (82). In a population comprising both genders, HIV-infected participants had thicker carotid intima-media thickness compared with controls. While HIV patients also had higher T-cell activation, high-sensitivity C-reactive protein levels, and CMV-specific T-cell responses, only CMV-specific T-cell responses were independently associated with intima-media thickness (83), suggesting that CMV-specific T-cell response may be the driving force linking HIV and carotid intima-media thickness. CMV and HIV may influence immunological aging in an additive manner, however, it has also been suggested that CMV may be the “smoking gun” in immunosenescence among persons coinfected with both pathogens, increasing the rate of immunological aging and the development of chronic diseases of inflammatory etiology, especially among those receiving ART (84). Thus, CMV or HIV may be codrivers of immunosenescence, or it may be that CMV accentuates or accelerates this mechanism. Possible mechanisms for the very high frequencies of CMV-specific T cells in antiretroviral-treated HIV disease include subclinical CMV replication and/or a dysregulated and heightened immunologic response to a normal level of CMV replication (82,84).
Biomarkers of Immune Dysfunction in HIV
In HIV infection, the heightened inflammation and immune dysfunction related to accelerated aging has stimulated assessment of several relevant biomarkers including C-reactive protein, IL-6, and D-dimer and their associations with age-related morbidity and HIV infection. These biomarkers of immune dysfunction have been reviewed (85). Developing interventions to decrease persistent inflammation in treated HIV infection is fast evolving as a priority within HIV research agenda (19,86). Cellular markers of immune activation as well as soluble inflammatory markers have been used as surrogate markers of disease and clinical endpoints in several studies (Supplementary Table 1).
BoA in HIV Infection
BoA have not been widely used to despite the hypothesis of premature or accelerated aging in HIV. Only two validated BoA that satisfy the majority of the criteria for a BoA as proposed by Baker and Sprott (87) have been broadly examined in those with HIV, telomere length (TL), and CDKN2A (p16ink4a) expression. CD8 T cells that lose expression of CD28 have shorter TL compared with other T- or B-cell subsets (88). CD28−CD8+ T cells from HIV-infected patients have significantly shorter TL compared with age-matched controls, with TL in the HIV-infected group similar to those found in elderly participants (89). However, TL is affected by psychosocial confounders, genetics, and potentially antiretroviral drugs (90), as human telomerase is a reverse transcriptase sharing homology with the HIV reverse transcriptase (91). Expression levels of the cell cycle regulator CDKN2A (which codes for the protein p16ink4a) may represent a more robust BoA (92). Increasing levels of CDKN2A messenger RNA occur with age and decreasing function of solid organs and peripheral blood leukocytes (93–95), and accumulation is accelerated in HIV patients. However, this relationship may be confounded in HIV infection by other mechanisms that lead to CDKN2A expression including smoking, and other factors (93,96). TL is shorter in HIV-infected individuals compared with age- and gender-matched uninfected counterparts, and similarly CDKN2A expression is increased in HIV-infected individuals (97).
Novel Ocular BoA: Potential Utility in HIV Infection
IL-6, D-dimer, and other inflammatory/coagulation markers are robustly influenced by comorbidity and HIV itself, and TL and CDKN2A expression are both primarily examined in peripheral blood T cells—obviously affected by HIV itself—novel biomarkers are required to evaluate biologic aging in those with HIV (98). Ocular markers may provide novel BoA because specific anatomic and functional parameters of the eye change with increasing chronological age (99). The eye is relatively easy to examine compared with other organ systems and there are few other locations in the body where cells, vasculature, and neurological tissues are directly and readily visible. A description of parameters that may be most informative as aging biomarkers is summarized in Table 2.
Table 2.
Ocular Parameters, Methods of Measurement, and the Impact of Aging
| Anatomical Site | Parameter | Method of Measurement | Age-Related Changes |
|---|---|---|---|
| Cornea | Corneal endothelium | Specular microscopy | Reduced cell count |
| Lens | Lens opacity | Subjective grading systems | Opacification at different sites in the lens (cataract) |
| Pentacam—lens densitometry | |||
| Retina | Retinal nerve fiber layer (RNFL) | Optical coherence tomography | Thinner RNFL |
| Retinal vessel caliber | Semiautomated retinal analysis software | Reduced diameter of arterioles and venules | |
| Retinal ganglion cells and photoreceptors | |||
| Confocal scanning ophthalmoscopy | Reduction in cone photoreceptor density | ||
| Visual function | Visual acuity | Visual acuity chart (ETDRS) | All reduced |
| Contrast sensitivity | Pelli–Robson chart | ||
| Color vision | Farnsworth–Munsell 100 hue test |
Note. ETDRS = Early treatment diabetic retinopathy study.
Conclusions
Does HIV accelerate or accentuate aging? The answer is probably organ and disease/condition specific. For many processes, there appears to be a pattern of accelerated aging. This is most clear in the immune system where T-cell TL and CDKN2A expression, accumulation of CD28−CD8+ T cells, reduced naive T-cell generation, and evidence on ongoing immune activation strongly suggest accelerated immune senescence. Clinically, it is also clear that the development of specific geriatric syndromes such as multimorbidity, frailty, and polypharmacy are hastened in those with HIV. In specific end-organ diseases, it is less clear, but many illnesses appear to be accentuated rather than accelerated (Figure 2). Cardiovascular disease, diabetes, and several other conditions are more prevalent at all ages in those with HIV, suggesting there is an extra “hit” by HIV and/or ART—that is, accentuated aging. Novel biomarkers in organs relatively protected from the effects of the HIV virus itself, concomitant exposures to additional stressors, and/or ART drug toxicity, such as the ocular measures proposed above, may prove to be useful, accurate BoA in those with HIV.
Figure 2.
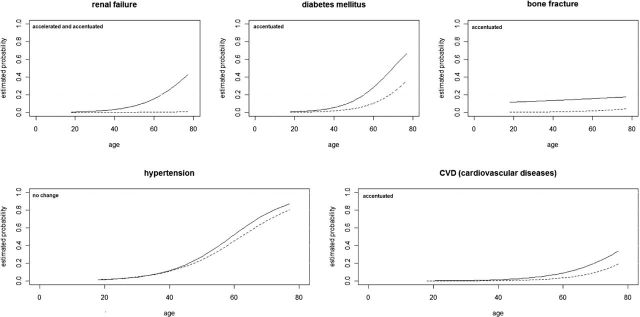
Comparative risk of hypertension, diabetes mellitus, renal failure, cardiovascular disease, and fracture, by age, among HIV-infected participants versus control participants. Modified from reference (6).
Supplementary Material
Supplementary material can be found at: http://biomedgerontology.oxfordjournals.org/.
Funding
A.L.L. is funded by Chicago Developmental Center for AIDS Research P30 AI 082151 and AI 068636. K.P.H. is funded by P30 AG021332.
References
- 1. Antiretroviral Therapy Cohort C. Life expectancy of individuals on combination antiretroviral therapy in high-income countries: a collaborative analysis of 14 cohort studies. Lancet. 2008;372:293–299 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. CDC. HIV/AIDS among persons aged 50 or over. http://www.cdc.gov/hiv/surveillance/resources/reports/2005report/ 2010
- 3. Negin J, Cumming RG. HIV infection in older adults in sub-Saharan Africa: extrapolating prevalence from existing data. Bull World Health Organ. 2010;88:847–853. 10.2471/BLT.10.076349 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Hontelez JA, Lurie MN, Newell ML, et al. Ageing with HIV in South Africa. AIDS. 2011;25:1665–1667. 10.1097/QAD.0b013e32834982ea [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Liu H, Lin X, Xu Y, Chen S, Shi J, Morisky D. Emerging HIV epidemic among older adults in Nanning, China. AIDS Patient Care STDS. 2012;26:565–567. 10.1089/apc.2012.0227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Guaraldi G, Orlando G, Zona S, et al. Premature age-related comorbidities among HIV-infected persons compared with the general population. Clin Infect Dis. 2011;53:1120–1126. 10.1093/cid/cir627 [DOI] [PubMed] [Google Scholar]
- 7. Effros RB, Fletcher CV, Gebo K, et al. Aging and infectious diseases: workshop on HIV infection and aging: what is known and future research directions. Clin Infect Dis. 2008;47:542–553. 10.1086/590150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Goulet JL, Fultz SL, Rimland D, et al. Aging and infectious diseases: do patterns of comorbidity vary by HIV status, age, and HIV severity? Clin Infect Dis. 2007;45:1593–1601. 10.1086/523577 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Kim DJ, Westfall AO, Chamot E, et al. Multimorbidity patterns in HIV-infected patients: the role of obesity in chronic disease clustering. J Acquir Immune Defic Syndr. 2012;61:600–605. 10.1097/QAI.0b013e31827303d5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. El-Sadr WM Lundgren JD Neaton JD et al. CD4+ count-guided interruption of antiretroviral treatment. N Engl J Med. 2006;355:2283–2296 [DOI] [PubMed] [Google Scholar]
- 11. Deeks SG, Phillips AN. HIV infection, antiretroviral treatment, ageing, and non-AIDS related morbidity. BMJ. 2009;338:a3172. 0.1136/bmj.a3172 [DOI] [PubMed] [Google Scholar]
- 12. Kuller LH, Tracy R, Belloso W, et al. ; INSIGHT SMART Study Group. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 2008;5:e203. 10.1371/journal.pmed.0050203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Deeks SG. Immune dysfunction, inflammation, and accelerated aging in patients on antiretroviral therapy. Top HIV Med. 2009;17:118–123 [PubMed] [Google Scholar]
- 14. Rickerts V, Brodt H, Staszewski S, Stille W. Incidence of myocardial infarctions in HIV-infected patients between 1983 and 1998: the Frankfurt HIV-cohort study. Eur J Med Res. 2000;5:329–333 [PubMed] [Google Scholar]
- 15. Salter ML, Lau B, Go VF, Mehta SH, Kirk GD. HIV infection, immune suppression, and uncontrolled viremia are associated with increased multimorbidity among aging injection drug users. Clin Infect Dis. 2011;53:1256–1264. 10.1093/cid/cir673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Hasse B, Ledergerber B, Furrer H, et al. ; Swiss HIV Cohort Study. Morbidity and aging in HIV-infected persons: the Swiss HIV cohort study. Clin Infect Dis. 2011;53:1130–1139. 10.1093/cid/cir626 [DOI] [PubMed] [Google Scholar]
- 17. Desquilbet L, Jacobson LP, Fried LP, et al. ; Multicenter AIDS Cohort Study. HIV-1 infection is associated with an earlier occurrence of a phenotype related to frailty. J Gerontol A Biol Sci Med Sci. 2007;62:1279–1286 [DOI] [PubMed] [Google Scholar]
- 18. Erlandson KM, Allshouse AA, Jankowski CM, et al. Risk factors for falls in HIV-infected persons. J Acquir Immune Defic Syndr. 2012;61:484–489. 10.1097/QAI.0b013e3182716e38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. High KP, Brennan-Ing M, Clifford DB, et al. ; OAR Working Group on HIV and Aging. HIV and aging: state of knowledge and areas of critical need for research. A report to the NIH Office of AIDS Research by the HIV and Aging Working Group. J Acquir Immune Defic Syndr. 2012;60(suppl 1):S1–18. 10.1097/QAI.0b013e31825a3668 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Martin J, Volberding P. HIV and premature aging: a field still in its infancy. Ann Intern Med. 2010;153:477–479. 10.7326/0003-4819-153-7-201010050-00013 [DOI] [PubMed] [Google Scholar]
- 21. Laprise C, Baril JG, Dufresne S, Trottier H. Association between tenofovir exposure and reduced kidney function in a cohort of HIV-positive patients: results from 10 years of follow-up. Clin Infect Dis. 2013;56:567–575. 10.1093/cid/cis937 [DOI] [PubMed] [Google Scholar]
- 22. Scherzer R, Estrella M, Li Y, et al. Association of tenofovir exposure with kidney disease risk in HIV infection. AIDS. 2012;26:867–875. 10.1097/QAD.0b013e328351f68f [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Hall AM, Hendry BM, Nitsch D, Connolly JO. Tenofovir-associated kidney toxicity in HIV-infected patients: a review of the evidence. Am J Kidney Dis. 2011;57:773–780. 10.1053/j.ajkd.2011.01.022 [DOI] [PubMed] [Google Scholar]
- 24. Leeansyah E, Cameron PU, Solomon A, et al. Inhibition of telomerase activity by human immunodeficiency virus (HIV) nucleos(t)ide reverse transcriptase inhibitors: a potential factor contributing to HIV-associated accelerated aging. J Infect Dis. 2013;207:1157–1165. 10.1093/infdis/jit006 [DOI] [PubMed] [Google Scholar]
- 25. Fisher M, Cooper V. HIV and ageing: premature ageing or premature conclusions? Curr Opin Infect Dis. 2012;25:1–3. 10.1097/QCO.0b013e32834f14fa [DOI] [PubMed] [Google Scholar]
- 26. Capeau J. Premature aging and premature age-related comorbidities in HIV-infected patients: facts and hypotheses. Clin Infect Dis. 2011;53:1127–1129. 10.1093/cid/cir628 [DOI] [PubMed] [Google Scholar]
- 27. Armah KA, McGinnis K, Baker J, et al. HIV status, burden of comorbid disease, and biomarkers of inflammation, altered coagulation, and monocyte activation. Clin Infect Dis. 2012;55:126–136. 10.1093/cid/cis406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Triant VA, Lee H, Hadigan C, Grinspoon SK. Increased acute myocardial infarction rates and cardiovascular risk factors among patients with human immunodeficiency virus disease. J Clin Endocrinol Metab. 2007;92:2506–2512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Guaraldi G, Zona S, Alexopoulos N, et al. Coronary aging in HIV-infected patients. Clin Infect Dis. 2009;49:1756–1762. 10.1086/648080 [DOI] [PubMed] [Google Scholar]
- 30. Martinez E, Mocroft A, García-Viejo MA, et al. Risk of lipodystrophy in HIV-1-infected patients treated with protease inhibitors: a prospective cohort study. Lancet. 2001;357:592–598 [DOI] [PubMed] [Google Scholar]
- 31. Wand H, Calmy A, Carey DL, et al. ; INITIO Trial International Coordinating Committee. Metabolic syndrome, cardiovascular disease and type 2 diabetes mellitus after initiation of antiretroviral therapy in HIV infection. AIDS. 2007;21:2445–2453 [DOI] [PubMed] [Google Scholar]
- 32. Cerrato E, D’Ascenzo F, Biondi-Zoccai G, et al. Cardiac dysfunction in pauci symptomatic human immunodeficiency virus patients: a meta-analysis in the highly active antiretroviral therapy era. Eur Heart J. 2013;34:1432–1436. 10.1093/eurheartj/ehs471 [DOI] [PubMed] [Google Scholar]
- 33. Papita A, Albu A, Fodor D, Itu C, Cârstina D. Arterial stiffness and carotid intima-media thickness in HIV infected patients. Med Ultrason. 2011;13:127–134 [PubMed] [Google Scholar]
- 34. Seaberg EC, Benning L, Sharrett AR, et al. Association between human immunodeficiency virus infection and stiffness of the common carotid artery. Stroke. 2010;41:2163–2170. 10.1161/STROKEAHA.110.583856 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Islam FM, Wu J, Jansson J, Wilson DP. Relative risk of renal disease among people living with HIV: a systematic review and meta-analysis. BMC Public Health. 2012;12:234. 10.1186/1471-2458-12-234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Cazanave C, Dupon M, Lavignolle-Aurillac V, et al. ; Groupe d’Epidémiologie Clinique du SIDA en Aquitaine. Reduced bone mineral density in HIV-infected patients: prevalence and associated factors. AIDS. 2008;22:395–402. 10.1097/QAD.0b013e3282f423dd [DOI] [PubMed] [Google Scholar]
- 37. Burgi A, Brodine S, Wegner S, et al. Incidence and risk factors for the occurrence of non-AIDS-defining cancers among human immunodeficiency virus-infected individuals. Cancer. 2005;104:1505–1511 [DOI] [PubMed] [Google Scholar]
- 38. Shiels MS, Pfeiffer RM, Engels EA. Age at cancer diagnosis among persons with AIDS in the United States. Ann Intern Med. 2010;153:452–460. 10.7326/0003-4819-153-7-201010050- 00008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Justice AC, Freiberg MS, Tracy R, et al. ; VACS Project Team. Does an index composed of clinical data reflect effects of inflammation, coagulation, and monocyte activation on mortality among those aging with HIV? Clin Infect Dis. 2012;54:984–994. 10.1093/cid/cir989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Fried LP, Tangen CM, Walston J, et al. ; Cardiovascular Health Study Collaborative Research Group. Frailty in older adults: evidence for a phenotype. J Gerontol A Biol Sci Med Sci. 2001;56:M146–M156 [DOI] [PubMed] [Google Scholar]
- 41. Bhavan KP, Kampalath VN, Overton ET. The aging of the HIV epidemic. Curr HIV/AIDS Rep. 2008;5:150–158 [DOI] [PubMed] [Google Scholar]
- 42. Shah K, Hilton TN, Myers L, Pinto JF, Luque AE, Hall WJ. A new frailty syndrome: central obesity and frailty in older adults with the human immunodeficiency virus. J Am Geriatr Soc. 2012;60:545–549. 10.1111/j.1532-5415.2011.03819.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Leng S, Chaves P, Koenig K, Walston J. Serum interleukin-6 and hemoglobin as physiological correlates in the geriatric syndrome of frailty: a pilot study. J Am Geriatr Soc. 2002;50:1268–1271 [DOI] [PubMed] [Google Scholar]
- 44. Janssen RS, Nwanyanwu OC, Selik RM, Stehr-Green JK. Epidemiology of human immunodeficiency virus encephalopathy in the United States. Neurology. 1992;42:1472–1476 [DOI] [PubMed] [Google Scholar]
- 45. Cherner M, Ellis RJ, Lazzaretto D, et al. ; HIV Neurobehavioral Research Center Group. Effects of HIV-1 infection and aging on neurobehavioral functioning: preliminary findings. AIDS. 2004;18(suppl 1):S27–S34 [PubMed] [Google Scholar]
- 46. McCutchan JA, Wu JW, Robertson K, et al. HIV suppression by HAART preserves cognitive function in advanced, immune-reconstituted AIDS patients. AIDS. 2007;21:1109–1117 [DOI] [PubMed] [Google Scholar]
- 47. Giorgi JV, Hultin LE, McKeating JA, et al. Shorter survival in advanced human immunodeficiency virus type 1 infection is more closely associated with T lymphocyte activation than with plasma virus burden or virus chemokine coreceptor usage. J Infect Dis. 1999;179: 859–870 [DOI] [PubMed] [Google Scholar]
- 48. Rodríguez B, Sethi AK, Cheruvu VK, et al. Predictive value of plasma HIV RNA level on rate of CD4 T-cell decline in untreated HIV infection. JAMA. 2006;296:1498–1506 [DOI] [PubMed] [Google Scholar]
- 49. Desai S, Landay A. Early immune senescence in HIV disease. Curr HIV/AIDS Rep. 2010;7:4–10. 10.1007/s11904-009-0038-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Hunt PW, Brenchley J, Sinclair E, et al. Relationship between T cell activation and CD4+ T cell count in HIV-seropositive individuals with undetectable plasma HIV RNA levels in the absence of therapy. J Infect Dis. 2008;197:126–133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Brenchley JM, Paiardini M, Knox KS, et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood. 2008;112:2826–2835. 10.1182/blood-2008-05-159301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Lagathu C, Eustace B, Prot M, et al. Some HIV antiretrovirals increase oxidative stress and alter chemokine, cytokine or adiponectin production in human adipocytes and macrophages. Antivir Ther. 2007;12:489–500 [PubMed] [Google Scholar]
- 53. Appay V, Almeida JR, Sauce D, Autran B, Papagno L. Accelerated immune senescence and HIV-1 infection. Exp Gerontol. 2007; 42:432–437 [DOI] [PubMed] [Google Scholar]
- 54. Pawelec G, Akbar A, Beverley P, et al. Immunosenescence and Cytomegalovirus: where do we stand after a decade? Immun Ageing. 2010;7:13. 10.1186/1742-4933-7-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. High KP, Akbar AN, Nikolich-Zugich J. Translational research in immune senescence: assessing the relevance of current models. Semin Immunol. 2012;24:373–382. 10.1016/j.smim.2012.04.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Czesnikiewicz-Guzik M, Lee WW, Cui D, et al. T cell subset-specific susceptibility to aging. Clin Immunol. 2008;127:107–118. 10.1016/j.clim.2007.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Vescovini R, Biasini C, Fagnoni FF, et al. Massive load of functional effector CD4+ and CD8+ T cells against cytomegalovirus in very old subjects. J Immunol. 2007;179:4283–4291 [DOI] [PubMed] [Google Scholar]
- 58. Xie D, McElhaney JE. Lower GrB+ CD62Lhigh CD8 TCM effector lymphocyte response to influenza virus in older adults is associated with increased CD28null CD8 T lymphocytes. Mech Ageing Dev. 2007;128:392–400 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Coppé JP, Desprez PY, Krtolica A, Campisi J. The senescence-associated secretory phenotype: the dark side of tumor suppression. Annu Rev Pathol. 2010;5:99–118. 10.1146/annurev-pathol-121808- 102144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Coppé JP, Rodier F, Patil CK, Freund A, Desprez PY, Campisi J. Tumor suppressor and aging biomarker p16(INK4a) induces cellular senescence without the associated inflammatory secretory phenotype. J Biol Chem. 2011;286:36396–36403. 10.1074/jbc.M111.257071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Onyema OO, Njemini R, Bautmans I, Renmans W, De Waele M, Mets T. Cellular aging and senescence characteristics of human T-lymphocytes. Biogerontology. 2012;13:169–181. 10.1007/s10522-011-9366-z [DOI] [PubMed] [Google Scholar]
- 62. Macaulay R, Akbar AN, Henson SM. The role of the T cell in age-related inflammation. Age (Dordr). 2013;35:563–572. 10.1007/s11357-012-9381-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Desai SR, Usuga X, Martinson J. Immune senescence, activation, and abnormal T cell homeostasis despite effective HAART, a hallmark of early aging in HIV disease. Presented at: 16th Conference on Retroviruses and Opportunistic Infections; 2009; February 8–11, 2009; Montreal, Canada [Google Scholar]
- 64. Khan N, Shariff N, Cobbold M, et al. Cytomegalovirus seropositivity drives the CD8 T cell repertoire toward greater clonality in healthy elderly individuals. J Immunol. 2002;169:1984–1992 [DOI] [PubMed] [Google Scholar]
- 65. Looney RJ, Falsey A, Campbell D, et al. Role of cytomegalovirus in the T cell changes seen in elderly individuals. Clin Immunol. 1999;90:213–219 [DOI] [PubMed] [Google Scholar]
- 66. Tassiopoulos K, Landay A, Collier AC, et al. CD28-negative CD4+ and CD8+ T cells in antiretroviral therapy-naive HIV-infected adults enrolled in adult clinical trials group studies. J Infect Dis. 2012;205:1730–1738. 10.1093/infdis/jis260 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Cao W, Jamieson BD, Hultin LE, Hultin PM, Effros RB, Detels R. Premature aging of T cells is associated with faster HIV-1 disease progression. J Acquir Immune Defic Syndr. 2009;50:137–147. 10.1097/QAI.0b013e3181926c28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Kaplan RC, Sinclair E, Landay AL, et al. T cell activation and senescence predict subclinical carotid artery disease in HIV-infected women. J Infect Dis. 2011;203:452–463. 10.1093/infdis/jiq071 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Kaplan RC, Sinclair E, Landay AL, et al. T cell activation predicts carotid artery stiffness among HIV-infected women. Atherosclerosis. 2011;217:207–213. 10.1016/j.atherosclerosis.2011.03.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Brenchley JM, Schacker TW, Ruff LE, et al. CD4+ T cell depletion during all stages of HIV disease occurs predominantly in the gastrointestinal tract. J Exp Med. 2004;200:749–759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Brenchley JM, Price DA, Schacker TW, et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med. 2006;12:1365–1371. 10.1016/j.tim.2012.09.001 [DOI] [PubMed] [Google Scholar]
- 72. Ipp H, Zemlin A. The paradox of the immune response in HIV infection: when inflammation becomes harmful. Clin Chim Acta. 2013;416:96–99. 10.1016/j.cca.2012.11.025 [DOI] [PubMed] [Google Scholar]
- 73. Martin GE, Gouillou M, Hearps AC, et al. Age-associated changes in monocyte and innate immune activation markers occur more rapidly in HIV infected women. PLoS One. 2013;8:e55279. 10.1371/journal.pone.0055279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Hearps AC, Maisa A, Cheng WJ, et al. HIV infection induces age-related changes to monocytes and innate immune activation in young men that persist despite combination antiretroviral therapy. AIDS. 2012;26:843–853. 10.1097/QAD.0b013e328351f756 [DOI] [PubMed] [Google Scholar]
- 75. Kelesidis T, Yang OO, Kendall MA, Hodis HN, Currier JS. Biomarkers of microbial translocation and macrophage activation are associated with progression of atherosclerosis in HIV infection: ACTG NWCS 332/A5078 study. Presented at: 19th Conference on Retroviruses and Opportunistic Infections; March 5–8, 2012; Seattle, WA [Google Scholar]
- 76. Manner I Kvale D Baekken M et al. Microbial translocation independently predicts future hypertension in HIV-infected individuals. Presented at: 19th Conference on Retroviruses and Opportunistic Infections; 2012; March 5–8, 2012; Seattle, WA [Google Scholar]
- 77. Sandler NG, Wand H, Roque A, et al. ; INSIGHT SMART Study Group. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J Infect Dis. 2011;203:780–790. 10.1093/infdis/jiq118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Pereyra F, Lo J, Triant VA, et al. Increased coronary atherosclerosis and immune activation in HIV-1 elite controllers. AIDS. 2012;26:2409–2412. 10.1097/QAD.0b013e32835a9950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Lang DJ, Kovacs AA, Zaia JA, et al. Seroepidemiologic studies of cytomegalovirus and Epstein-Barr virus infections in relation to human immunodeficiency virus type 1 infection in selected recipient populations. Transfusion Safety Study Group. J Acquir Immune Defic Syndr. 1989;2:540–549 [PubMed] [Google Scholar]
- 80. Alonso Arias R, Moro-García MA, Echeverría A, Solano-Jaurrieta JJ, Suárez-García FM, López-Larrea C. Intensity of the humoral response to cytomegalovirus is associated with the phenotypic and functional status of the immune system. J Virol. 2013;87:4486–4495. 10.1128/JVI.02425-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Naeger DM, Martin JN, Sinclair E, et al. Cytomegalovirus-specific T cells persist at very high levels during long-term antiretroviral treatment of HIV disease. PLoS One. 2010;5:e8886. 10.1371/journal.pone.0008886 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Parrinello CM, Sinclair E, Landay AL, et al. Cytomegalovirus immunoglobulin G antibody is associated with subclinical carotid artery disease among HIV-infected women. J Infect Dis. 2012;205:1788–1796 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Hsue PY, Hunt PW, Sinclair E, et al. Increased carotid intima-media thickness in HIV patients is associated with increased cytomegalovirus-specific T-cell responses. AIDS. 2006;20:2275–2283 [DOI] [PubMed] [Google Scholar]
- 84. Aiello AE, Simanek AM. Cytomegalovirus and immunological aging: the real driver of HIV and heart disease? J Infect Dis. 2012;205:1772–1774. 10.1093/infdis/jis288 [DOI] [PubMed] [Google Scholar]
- 85. Nixon DE, Landay AL. Biomarkers of immune dysfunction in HIV. Curr Opin HIV AIDS. 2010;5:498–503. 10.1097/COH.0b013e32833ed6f4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Deeks SG, Verdin E, McCune JM. Immunosenescence and HIV. Curr Opin Immunol. 2012;24:501–506. 10.1016/j.coi.2012.05.004 [DOI] [PubMed] [Google Scholar]
- 87. Baker GT, 3rd, Sprott RL. Biomarkers of aging. Exp Gerontol. 1988;23:223–239 [DOI] [PubMed] [Google Scholar]
- 88. Lin J, Epel E, Cheon J, et al. Analyses and comparisons of telomerase activity and telomere length in human T and B cells: insights for epidemiology of telomere maintenance. J Immunol Methods. 2010;352:71–80. 10.1016/j.jim.2009.09.012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Effros RB, Allsopp R, Chiu CP, et al. Shortened telomeres in the expanded CD28-CD8+ cell subset in HIV disease implicate replicative senescence in HIV pathogenesis. AIDS. 1996;10:F17–F22 [DOI] [PubMed] [Google Scholar]
- 90. Yamaguchi T, Takayama Y, Saito M, Ishikawa F, Saneyoshi M. Telomerase-inhibitory effects of the triphosphate derivatives of some biologically active nucleosides. Nucleic Acids Res Suppl. 2001:211–212 [DOI] [PubMed] [Google Scholar]
- 91. Peng Y, Mian IS, Lue NF. Analysis of telomerase processivity: mechanistic similarity to HIV-1 reverse transcriptase and role in telomere maintenance. Mol Cell. 2001;7:1201–1211 [DOI] [PubMed] [Google Scholar]
- 92. Shiels PG. Improving precision in investigating aging: why telomeres can cause problems. J Gerontol A Biol Sci Med Sci. 2010;65:789–791. 10.1093/gerona/glq095 [DOI] [PubMed] [Google Scholar]
- 93. Liu Y, Sanoff HK, Cho H, et al. Expression of p16(INK4a) in peripheral blood T-cells is a biomarker of human aging. Aging Cell. 2009;8:439–448. 10.1111/j.1474-9726.2009.00489.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94. Krishnamurthy J, Torrice C, Ramsey MR, et al. Ink4a/Arf expression is a biomarker of aging. J Clin Invest. 2004;114:1299–1307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95. Koppelstaetter C, Schratzberger G, Perco P, et al. Markers of cellular senescence in zero hour biopsies predict outcome in renal transplantation. Aging Cell. 2008;7:491–497. 10.1111/j.1474-9726. 2008.00398.x [DOI] [PubMed] [Google Scholar]
- 96. Nelson JA, Krishnamurthy J, Menezes P, et al. Expression of p16(INK4a) as a biomarker of T-cell aging in HIV-infected patients prior to and during antiretroviral therapy. Aging Cell. 2012;11:916–918. 10.1111/j.1474-9726.2012.00856.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Pathai S, Lawn SD, Gilbert CE, et al. Accelerated biological ageing in HIV-infected individuals in South Africa: a case-control study. AIDS. 2013;27:2375–2384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Lundgren JD, Baxter J, Deeks SG, Lane HC. Biomarkers in HIV disease. Curr Opin HIV AIDS. 2010;5:459–462. 10.1097/COH.0b013e32833f2ed6 [DOI] [PubMed] [Google Scholar]
- 99. Pathai S, Shiels PG, Lawn SD, Cook C, Gilbert C. The eye as a model of ageing in translational research–molecular, epigenetic and clinical aspects. Ageing Res Rev. 2013;12:490–508. 10.1016/j.arr.2012.11.002 [DOI] [PubMed] [Google Scholar]