

Abstract
Background
Human epidermal γδ T cells are known to play crucial roles in the defense and homeostasis of the skin. However, their precise mechanism of action in skin inflammation remains less clear.
Objective
In this study, we analyzed the cytokine expression profile of human epidermal γδ T cells and compared it to that of peripheral blood γδ T cells to investigate the specific activity of epidermal γδ T cells in modulating skin inflammation.
Methods
We isolated γδ T cells from epidermal tissue or peripheral blood obtained from healthy volunteers. Isolated γδ T cells were stimulated using immobilized anti-CD3 antibody and interleukin-2 plus phytohaemagglutinin, and were then analyzed using a cytokine array kit.
Results
Both epidermal and peripheral blood γδ T cells produced comparable levels of granulocyte-macrophage colony-stimulating factor, I-309, interferon-γ, macrophage migration inhibitory factor, macrophage inflammatory protein-1α, and chemokine (C-C) ligand 5. The epidermal γδ T cells produced significantly higher levels of interleukin-4, -8, -13, and macrophage inflammatory protein-1β than the peripheral blood γδ T cells did. Notably, the epidermal γδ T cells produced several hundred-fold higher levels of interleukin-13 than interleukin-4.
Conclusion
These results suggest that the epidermal γδ T cells have a stronger potential to participate in the Th2-type response than the peripheral blood γδ T cells do. Furthermore, epidermal γδ T cells might play an important role in the pathogenesis of Th2-dominant skin diseases because of their active production of interleukin-13.
Keywords: Epidermis, Interleukin-4, Interleukin-13, γδ T cells
INTRODUCTION
The skin, composed of epidermis, dermis, and subcutaneous tissue, is one of the most active organs in the human body. The epidermis, the outermost barrier tissue, is composed of keratinocytes, melanocytes, and immune cells such as Langerhans cells and γδ T cells1. Epidermal cells actively communicate with each other through a variety of soluble signaling factors (growth factors, cytokines, chemokines, and inflammatory mediators) and through direct contact mediated by cell surface molecules. γδ T cells are the first T cells that emigrate from the thymus, and many of these γδ T cells take residence in epithelial tissues, including the skin, intestine, lung, and reproductive tract2. Epidermis-resident γδ T cells are derived from fetal thymic precursor cells and move to skin before birth3. Evidence from multiple laboratories indicates that epidermal γδ T cells play specialized roles in the maintenance of epithelial homeostasis, wound healing, tumor surveillance, infection, and inflammation1,2,3,4,5,6,7,8. These γδ T cells have been shown to express interleukin (IL)-2, IL-3, granulocyte-macrophage colony stimulating factor (GM-CSF), interferon-γ (IFNγ), tumor necrosis factor-α, chemokine (C-C) ligand (CCL)-3 (macrophage inflammatory protein [MIP]-1α), CCL-4 (MIP-1β), CCL-5 (RANTES), and chemokine (C motif) ligand 12,5,6. Similar to chemokines, these cytokines are detected at low levels under resting conditions, but are rapidly upregulated upon stimulation. The ability of epidermal γδ T cells to produce numerous cytokines and chemokines suggests that these molecules are key players in the immune responses stimulated by the epidermal γδ T cells.
Epidermal γδ T cells are significantly different from those of the peripheral blood in terms of ontogeny, tissue tropism, and antigen receptor diversity6,9. In humans, most of the peripheral blood γδ T cells express the Vδ2 gene segment10. Peripheral γδ T cells recognize, expand, and release cytokines in response to non-peptide antigens, which are mostly of microbial origin. In contrast, most γδ T cells in epithelial tissues express the Vδ1 gene segment and respond to poorly defined self-antigens expressed by stressed cells3,11,12,13,14. In addition, epidermal γδ T cells primarily possess tissue-specific T cell receptors (TCRs) with limited or no diversity compared to the diverse TCRs expressed by γδ T cells found in the peripheral lymphoid organs and blood13. However, it is unclear whether human epidermal γδ T cells exert any functions different from those of peripheral γδ T cells resulting from their differential cytokine production profiles.
In the present study, we compared the cytokine profiles of human epidermal and peripheral blood γδ T cells to investigate the differential activities of epidermal γδ T cells, which affect their neighboring epidermal cells.
MATERIALS AND METHODS
Human skin specimens and blood samples
Skin and peripheral blood samples were collected from human volunteers after informed consent had been obtained according to the approval of the Institutional Review Board at Seoul National University Hospital (IRB No. H-1012-054-344).
Preparation of human epidermal γδ T cells
Whole skin specimens larger than 3×5 cm2 were obtained from three healthy volunteers undergoing elective breast surgery. Skin specimens were incubated in RPMI media containing 2.4 U/ml dispase II (Roche Applied Science, Indianapolis, IN, USA) overnight at 4℃. Epidermal sheets were separated from the dermis by using forceps, and then cultured in complete media (RPMI 1640 media supplemented with 2 mM L-glutamine, 100 U/ml penicillin, 100 mg/ml streptomycin, 10% heat-inactivated fetal calf serum, 100 mM nonessential amino acids, 25 mM 4-(2-Hydroxyethyl)piperazine-1-ethanesulfonic acid (HEPES), 1 mM sodium pyruvate, and 50 mM 2-b-mercaptoethanol in the presence of 50 U/ml IL-2 at 37℃ in a 5% CO2 incubator for 3 to 4 days. Epidermal sheets were then gently agitated to isolate the epidermal cells. Harvested epidermal cell suspensions were enriched for T cells by using Histopaque 1077 (Sigma-Aldrich, St. Louis, MO, USA) gradient centrifugation at 400×g for 30 minutes at room temperature. Epidermal γδ T cells were selected from epidermal mononuclear cell suspensions by using the TCRγδ+ T Cell Isolation Kit (Miltenyi Biotec, Bergisch Gladbach, Germany) and a magnetic cell separator, according to the manufacturer's instructions.
Preparation of human peripheral blood γδ T cells
Human peripheral blood mononuclear cells were isolated from peripheral blood, collected from the antecubital veins of three healthy volunteers, by performing Histopaque 1077 gradient centrifugation at 400×g for 30 minutes at room temperature. Peripheral γδ T cells were selected using the TCRγδ+ T Cell Isolation Kit and a magnetic cell separator, according to the manufacturer's instructions.
Flow cytometry
The purity of γδ T cells was analyzed by flow cytometry using fluorescein isothiocyanate-conjugated anti-human CD3 monoclonal antibodies (mAbs; BD Biosciences, Bergisch Gladbach, Germany), APC-conjugated anti-human TCRγδ complex mAbs (BD Biosciences, San Jose, CA, USA), or the corresponding fluorescently conjugated isotype-matched control Abs. Two-color flow cytometry was performed using the BD FACSCalibur Flow Cytometer, and the results were analyzed using the Cell Quest software (BD Biosciences, USA).
Cytokine array
Epidermal and peripheral blood γδ T cells were stimulated using immobilized 10 mg/ml anti-human CD3 mAb (BD Biosciences, USA) and 100 U/ml IL-2 plus 500 ng/ml phytohaemagglutinin for 36 hours. The supernatants of the cultures were analyzed using the Proteome Profiler Human Cytokine Array Kit, Panel A (R&D systems, Minneapolis, MN, USA), according to the manufacturer's instructions. The cytokine blot intensities on the array membranes were measured using the ChemiDoc gel documentation system (Bio-Rad, Hercules, CA, USA). The relative intensities of the blots were calculated using the following formula: 100×(intensity of sample-intensity of negative control)/(intensity of positive control-intensity of negative control). Data are presented as means±standard deviation.
Enzyme-linked immunosorbent assays
IL-4 and IL-13 levels were measured in the culture supernatants by using the OptEIA™ human IL-4 enzyme-linked immunosorbent assay (ELISA) set and human IL-13 ELISA set (BD Biosciences, USA), respectively, according to the manufacturer's instructions.
Statistical analysis
Data were analyzed using the Student's t-test to determine the significant differences between the cytokine levels in epidermal γδ T cells versus those in peripheral γδ T cells. SPSS ver. 15.0.1. software (SPSS Inc., Chicago, IL, USA) was used for statistical analyses. A p-value of <0.05 was considered statistically significant.
RESULTS
Flow cytometric analysis of the purified human epidermal and peripheral blood γδ T cells
The purity of CD3+ TCR γδ+ cells isolated from the epidermis was 98.1% (Fig. 1A) and 90.6% from the peripheral blood (Fig. 1B). The results confirmed that the purities of γδ T cells were sufficient for them to be used in the cytokine array experiments.
Fig. 1.

Flow cytometric analysis of the purified epidermal and peripheral blood γδ T cells. Purified γδ T cells obtained from human epidermal tissue (A) and from the peripheral blood (B) of healthy volunteers were stained using a fluorescein-conjugated anti-CD3 monoclonal antibody (mAb) and APC-conjugated anti-γδ T cell receptor mAb or the corresponding fluorescently conjugated isotype-matched control Abs. Three independent experiments were performed for epidermal cells and peripheral blood cells respectively, and the representative results are presented. FITC: fluorescein isothiocyanate.
Comparison of the cytokine expression profiles of epidermal and peripheral blood γδ T cells
The blot intensities on the cytokine array membranes from activated epidermal (Fig. 2A~C) and activated peripheral blood γδ T cells (Fig. 2D~F) were measured as described in the Materials and Methods. A summary of these data is presented in Fig. 2G (epidermal γδ T cells) and Fig. 2H (peripheral blood γδ T cells). These data showed that both epidermal and peripheral blood γδ T cells produced comparable levels of GM-CSF, I-309 (CCL-1), IFNγ, macrophage migration inhibitory factor, MIP-1α (CCL-3), and RANTES (CCL-5). In addition, we found that epidermal γδ T cells produced significantly higher levels of IL-4, IL-8, IL-13, and MIP-1β than peripheral blood γδ T cells did (p<0.05).
Fig. 2.

Comparison of cytokine expression profiles of epidermal and peripheral blood γδ T cells. Epidermal γδ T cells were isolated from the epidermes of three healthy volunteers (A~C) by using the method described in the Materials and Methods section. Peripheral blood γδ T cells were isolated from the peripheral blood of three healthy volunteers (D~F) by using conventional Histopaque 1077 gradient centrifugation, followed by magnetic-activated cell sorting. Epidermal and peripheral blood γδ T cells were stimulated using immobilized anti-CD3 antibody and IL-2 plus phytohaemagglutinin. The culture supernatants were analyzed after 36 hours by using the Human Cytokine Array Panel A. (A~F) cytokine array membranes. (G, H) The blot intensities on the array membranes were measured using a gel documentation system. Data are presented as the mean±standard deviation. Statistical analyses were performed using the Student's t-test. C5a: complement component 5a, G-CSF: granulocyte colony-stimulating factor, GM-CSF: granulocyte-macrophage colony stimulating factor, GROα: growth-related oncogene α, sICAM-1: soluble intercellular adhesion molecule-1, IFNγ: interferon-γ, IL: interleukin, IP-10: interferon gamma-induced protein 10, I-TAC: interferon-inducible T cell α chemoattractant, MCP-1: monocyte chemotactic protein 1, MIP: macrophage inflammatory protein, RANTES: regulated on activation, normal T cell expressed and secreted, SDF-1: stromal cell-derived factor 1, TNF-α: tumor necrosis factor α, sTREM-1: soluble triggering receptor expressed on myeloid cells 1. *p<0.05, epidermal vs. peripheral blood γδ T cells.
Quantitative analyses of interleukin-4 and interleukin-13 production in epidermal and peripheral blood γδ T cells
The ELISA results presented in Fig. 3 confirmed that epidermal γδ T cells produced significantly higher levels of IL-4 and IL-13 than peripheral blood γδ T cells did. At the same time, epidermal γδ T cells produced IL-13 at levels several hundred-fold higher than the levels of IL-4 produced.
Fig. 3.
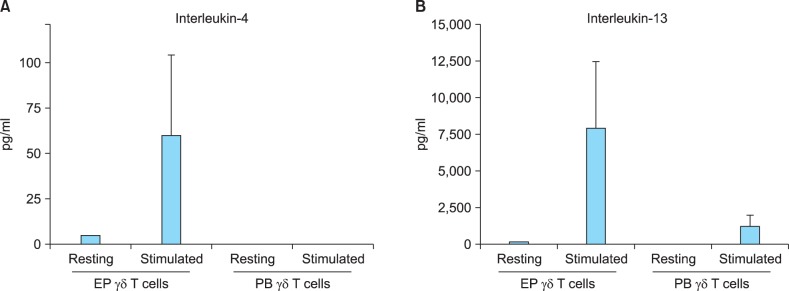
Quantitative analyses of interleukin-4 and interleukin-13 production in epidermal γδ T cells and peripheral blood γδ T cells. Epidermal and peripheral blood γδ T cells were each isolated from three healthy volunteers. Cells were stimulated using immobilized anti-CD3 antibody and interleukin-2 plusphytohaemagglutinin, or cultured without any stimulation. The culture supernatants were analyzed after 36 hours by using ELISAs. Data are presented as the mean±standard deviation of three independent experiments. Statistical analyses were performed using the Student's t-test. EP: epidermal, PB: peripheral blood. *p<0.05, epidermal vs. peripheral γδ T cells.
DISCUSSION
In this study, we analyzed and compared the cytokine expression profiles of human epidermal and peripheral blood γδ T cells to determine the specific activity of epidermal γδ T cells in modulating skin immune responses.
Although activated epidermal γδ T cells are a rich source of several cytokines and chemokines, the transcripts of IL-1α, IL-1β, IL-6, IL-7, IL-10, and IL-13 are in low abundance, and intracellular cytokine staining fails to detect IL-4, IL-5, and IL-1015,16. Thus, it was unexpected that we found that activated human epidermal γδ T cells produced significant levels of both Th1-type cytokines (IFNγ) and Th2-type cytokines (IL-4, IL-5, and IL-13) at the protein level (Fig. 2A). Next, we checked the purity of the cells to confirm that IL-4 and IL-13 were produced by γδ T cells and not by other cells contaminating the cultures. Our flow cytometry results showed that 98.1% and 90.6% of epidermal and peripheral blood cells, respectively, were CD3+TCRγδ+ (Fig. 1).
Interestingly, our results showed that activated epidermal γδ T cells produced significantly higher levels of IL-4, IL-8, IL-13, and MIP-1β than peripheral blood γδ T cells (Fig. 2). IL-8 and MIP-1β are well-known chemotactic factors that are produced from epithelial cells as well as from lymphocytes. In this study, we focused on the predominant cytokines produced by T cells, IL-4 and IL-13, which are typical Th2-type cytokines17. These cytokines affect a variety of cell types including T cells, B cells, natural killer cells, mast cells, monocytes/macrophages, endothelial cells, epithelial cells, dendritic cells, fibroblasts and keratinocytes18,19. In the skin, IL-4 and IL-13 are known to play multiple roles in diseases such as atopic dermatitis (AD) and vitiligo20,21. The concentration of IL-13 is reported to be abnormally high in the serum of AD and vitiligo patients22,23,24,25, and the acute skin lesions of AD patients contain increased numbers of cells expressing IL-4, IL-5, and IL-13 mRNA26,27,28. IL-4 might be important during the initial phase of allergic responses and in the priming and development of Th2 cells, whereas IL-13 plays a more critical role in immunoglobulin E induction and chronic AD pathogenesis29. Our quantitative ELISA data confirmed that epidermal γδ T cells produced markedly higher levels of IL-4 and IL-13 than peripheral blood γδ T cells, and the amount of IL-13 produced by the cells was several hundred-fold higher than the levels of IL-4 produced (Fig. 3). In summary, our results suggest that epidermis-resident γδ T cells have stronger potential to participate in Th2-type responses than γδ T cells existing in the periphery. These data suggest that epidermal γδ T cells play an important role in the pathogenesis of Th2-dominant skin diseases through the active production of IL-13.
ACKNOWLEDGMENT
This study was supported by a grant of the Korean Health Technology R&D Project, Ministry of Health & Welfare, Republic of Korea (Grant No. HN10C0009) and under a research agreement with AmorePacific Corporation, Republic of Korea.
References
- 1.Komori HK, Meehan TF, Havran WL. Epithelial and mucosal gamma delta T cells. Curr Opin Immunol. 2006;18:534–538. doi: 10.1016/j.coi.2006.06.001. [DOI] [PubMed] [Google Scholar]
- 2.Toulon A, Breton L, Taylor KR, Tenenhaus M, Bhavsar D, Lanigan C, et al. A role for human skin-resident T cells in wound healing. J Exp Med. 2009;206:743–750. doi: 10.1084/jem.20081787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Havran WL, Chien YH, Allison JP. Recognition of self antigens by skin-derived T cells with invariant gamma delta antigen receptors. Science. 1991;252:1430–1432. doi: 10.1126/science.1828619. [DOI] [PubMed] [Google Scholar]
- 4.Ferrarini M, Ferrero E, Dagna L, Poggi A, Zocchi MR. Human gammadelta T cells: a nonredundant system in the immune-surveillance against cancer. Trends Immunol. 2002;23:14–18. doi: 10.1016/s1471-4906(01)02110-x. [DOI] [PubMed] [Google Scholar]
- 5.Ebert LM, Meuter S, Moser B. Homing and function of human skin gammadelta T cells and NK cells: relevance for tumor surveillance. J Immunol. 2006;176:4331–4336. doi: 10.4049/jimmunol.176.7.4331. [DOI] [PubMed] [Google Scholar]
- 6.Macleod AS, Havran WL. Functions of skin-resident γδ T cells. Cell Mol Life Sci. 2011;68:2399–2408. doi: 10.1007/s00018-011-0702-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jameson J, Havran WL. Skin gammadelta T-cell functions in homeostasis and wound healing. Immunol Rev. 2007;215:114–122. doi: 10.1111/j.1600-065X.2006.00483.x. [DOI] [PubMed] [Google Scholar]
- 8.Nestle FO, Di Meglio P, Qin JZ, Nickoloff BJ. Skin immune sentinels in health and disease. Nat Rev Immunol. 2009;9:679–691. doi: 10.1038/nri2622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Girardi M. Immunosurveillance and immunoregulation by gammadelta T cells. J Invest Dermatol. 2006;126:25–31. doi: 10.1038/sj.jid.5700003. [DOI] [PubMed] [Google Scholar]
- 10.Urban EM, Chapoval AI, Pauza CD. Repertoire development and the control of cytotoxic/effector function in human gammadelta T cells. Clin Dev Immunol. 2010;2010:732893. doi: 10.1155/2010/732893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hayday AC. [gamma][delta] cells: a right time and a right place for a conserved third way of protection. Annu Rev Immunol. 2000;18:975–1026. doi: 10.1146/annurev.immunol.18.1.975. [DOI] [PubMed] [Google Scholar]
- 12.Carding SR, Egan PJ. Gammadelta T cells: functional plasticity and heterogeneity. Nat Rev Immunol. 2002;2:336–345. doi: 10.1038/nri797. [DOI] [PubMed] [Google Scholar]
- 13.Groh V, Steinle A, Bauer S, Spies T. Recognition of stress-induced MHC molecules by intestinal epithelial gammadelta T cells. Science. 1998;279:1737–1740. doi: 10.1126/science.279.5357.1737. [DOI] [PubMed] [Google Scholar]
- 14.Ferrarini M, Ferrero E, Dagna L, Poggi A, Zocchi MR. Human gammadelta T cells: a nonredundant system in the immune-surveillance against cancer. Trends Immunol. 2002;23:14–18. doi: 10.1016/s1471-4906(01)02110-x. [DOI] [PubMed] [Google Scholar]
- 15.Matsue H, Cruz PD, Jr, Bergstresser PR, Takashima A. Profiles of cytokine mRNA expressed by dendritic epidermal T cells in mice. J Invest Dermatol. 1993;101:537–542. doi: 10.1111/1523-1747.ep12365917. [DOI] [PubMed] [Google Scholar]
- 16.Boismenu R, Feng L, Xia YY, Chang JC, Havran WL. Chemokine expression by intraepithelial gamma delta T cells. Implications for the recruitment of inflammatory cells to damaged epithelia. J Immunol. 1996;157:985–992. [PubMed] [Google Scholar]
- 17.Romagnani S. Regulation of the T cell response. Clin Exp Allergy. 2006;36:1357–1366. doi: 10.1111/j.1365-2222.2006.02606.x. [DOI] [PubMed] [Google Scholar]
- 18.Taub D. Current Protocols in Immunology. New York: John Wiley & Sons, Inc.; 2004. Cytokine, growth factor, and chemokine ligand database; pp. 6.29.1–6.29.89. [DOI] [PubMed] [Google Scholar]
- 19.Yang Y, Yoo HM, Choi I, Pyun KH, Byun SM, Ha H. Interleukin 4-induced proliferation in normal human keratinocytes is associated with c-myc gene expression and inhibited by genistein. J Invest Dermatol. 1996;107:367–372. doi: 10.1111/1523-1747.ep12363346. [DOI] [PubMed] [Google Scholar]
- 20.Heinzmann A, Mao XQ, Akaiwa M, Kreomer RT, Gao PS, Ohshima K, et al. Genetic variants of IL-13 signalling and human asthma and atopy. Hum Mol Genet. 2000;9:549–559. doi: 10.1093/hmg/9.4.549. [DOI] [PubMed] [Google Scholar]
- 21.Oh MH, Oh SY, Yu J, Myers AC, Leonard WJ, Liu YJ, et al. IL-13 induces skin fibrosis in atopic dermatitis by thymic stromal lymphopoietin. J Immunol. 2011;186:7232–7242. doi: 10.4049/jimmunol.1100504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Novak N, Kruse S, Kraft S, Geiger E, Klüken H, Fimmers R, et al. Dichotomic nature of atopic dermatitis reflected by combined analysis of monocyte immunophenotyping and single nucleotide polymorphisms of the interleukin-4/interleukin-13 receptor gene: the dichotomy of extrinsic and intrinsic atopic dermatitis. J Invest Dermatol. 2002;119:870–875. doi: 10.1046/j.1523-1747.2002.00191.x. [DOI] [PubMed] [Google Scholar]
- 23.Hamid Q, Naseer T, Minshall EM, Song YL, Boguniewicz M, Leung DY. In vivo expression of IL-12 and IL-13 in atopic dermatitis. J Allergy Clin Immunol. 1996;98:225–231. doi: 10.1016/s0091-6749(96)70246-4. [DOI] [PubMed] [Google Scholar]
- 24.Tazawa T, Sugiura H, Sugiura Y, Uehara M. Relative importance of IL-4 and IL-13 in lesional skin of atopic dermatitis. Arch Dermatol Res. 2004;295:459–464. doi: 10.1007/s00403-004-0455-6. [DOI] [PubMed] [Google Scholar]
- 25.Wańkowicz-Kalińska A, van den Wijngaard RM, Tigges BJ, Westerhof W, Ogg GS, Cerundolo V, et al. Immunopolarization of CD4+ and CD8+ T cells to Type-1-like is associated with melanocyte loss in human vitiligo. Lab Invest. 2003;83:683–695. doi: 10.1097/01.lab.0000069521.42488.1b. [DOI] [PubMed] [Google Scholar]
- 26.Hamid Q, Boguniewicz M, Leung DY. Differential in situ cytokine gene expression in acute versus chronic atopic dermatitis. J Clin Invest. 1994;94:870–876. doi: 10.1172/JCI117408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hamid Q, Naseer T, Minshall EM, Song YL, Boguniewicz M, Leung DY. In vivo expression of IL-12 and IL-13 in atopic dermatitis. J Allergy Clin Immunol. 1996;98:225–231. doi: 10.1016/s0091-6749(96)70246-4. [DOI] [PubMed] [Google Scholar]
- 28.Novak N, Kraft S, Bieber T. Unraveling the mission of FcepsilonRI on antigen-presenting cells. J Allergy Clin Immunol. 2003;111:38–44. doi: 10.1067/mai.2003.2. [DOI] [PubMed] [Google Scholar]
- 29.Akdis M, Akdis CA, Weigl L, Disch R, Blaser K. Skin-homing, CLA+ memory T cells are activated in atopic dermatitis and regulate IgE by an IL-13-dominated cytokine pattern: IgG4 counter-regulation by CLA-memory T cells. J Immunol. 1997;159:4611–4619. [PubMed] [Google Scholar]