

Abstract
The class Thermoplasmata harbors huge uncultured archaeal lineages at the order level, so-called Groups E2 and E3. A novel archaeon Kjm51a affiliated with Group E2 was enriched from anaerobic sludge in the present study. Clone library analysis of the archaeal 16S rRNA and mcrA genes confirmed a unique archaeal population in the enrichment culture. The 16S rRNA gene-based phylogeny revealed that the enriched archaeon Kjm51a formed a distinct cluster within Group E2 in the class Thermoplasmata together with Methanomassiliicoccus luminyensis B10T and environmental clone sequences derived from anaerobic digesters, bovine rumen, and landfill leachate. Archaeon Kjm51a showed 87.7% 16S rRNA gene sequence identity to the closest cultured species, M. luminyensis B10T, indicating that archaeon Kjm51a might be phylogenetically novel at least at the genus level. In fluorescence in situ hybridization analysis, archaeon Kjm51a was observed as coccoid cells completely corresponding to the archaeal cells detected, although bacterial rod cells still coexisted. The growth of archaeon Kjm51a was dependent on the presence of methanol and yeast extract, and hydrogen and methane were produced in the enrichment culture. The addition of 2-bromo ethanesulfonate to the enrichment culture completely inhibited methane production and increased hydrogen concentration, which suggested that archaeon Kjm51a is a methanol-reducing hydrogenotrophic methanogen. Taken together, we propose the provisional taxonomic assignment, named Candidatus Methanogranum caenicola, for the enriched archaeon Kjm51a belonging to Group E2. We also propose to place the methanogenic lineage of the class Thermoplasmata in a novel order, Methanomassiliicoccales ord. nov.
Keywords: Methanogranum caenicola, methanogen, Thermoplasmata, rice cluster III, anaerobic digested sludge
Until recently, the class Thermoplasmata had consisted of mainly acidophilic, aerobic, mesophilic to thermophilic, and sulfur-reducing archaea such as genera Acidiplasma (15), Ferroplasma (14), Picrophilus (47), Thermoplasma (4), Thermogymnomonas (28), and Candidatus Acidulipro-fundum boonei (50). Archaeal members of those genera mainly inhabit extreme environments such as acidic and solfataric fields. On the other hand, culture-independent approaches have retrieved a diverse array of environmental clones belonging to the class Thermoplasmata from ordinary environments, and many of these clones form huge uncultured archaeal lineages at the order level, so-called Groups E2 and E3 (6, 33, 39). Groups E2 and E3 consist of sublineages such as Marine group II, deep-sea hydrothermal vent Euryarchaeotic group 1 and 2 (DHVE1 and DHVE2), and rice cluster III (RC-III), which is derived from the alimentary canal (12, 19, 54), anaerobic digester (13), contaminated aquifer (8), deep-sea hydrothermal vent (40, 55), marine plankton (5, 7), and rice field soil (3, 17, 30). More recently, a uniformly shaped pure culture B10T, given the name Methanomassiliicoccus luminyensis, was isolated from human feces, and revealed to be a methanol-reducing, mesophilic, slightly alkaliphilic methanogen belonging to the class Thermoplasmata (9). These findings suggest that the class Thermoplasmata is a phenotypically versatile taxon; however, very little is known about the phylogenetic diversity and ecological distribution of methanogens in the class Thermoplasmata.
In our previous study, members of RC-III within Group E2 in the Thermoplasmata as well as Methanoculleus, Methanosarcina, and Methanothermobacter species have been detected from methanogenic bioreactors (1, 21, 43, 44, 46). To obtain cultures of those methanogens, we conducted enrichment cultures from methanogenic digester sludge and eventually succeeded in enriching a novel methanogen belonging to Group E2 in class Thermoplasmata. Thus, this paper deals with the phylogenetic characterization of the enriched methanogen in Thermoplasmata and the provisional characterization of the phenotypes.
Materials and Methods
Sampling
The anaerobic sludge was collected from a methanogenic packed-bed reactor at Kajima Technical Research Institute on 16th December 2004. The reactor, which was packed with carbon fiber textile as supporting media (43–45), had been properly operated at 55°C and was stably producing methane gas from garbage slurry as feedstock. The garbage slurry was prepared from kitchen waste from the company cafeteria. It was diluted with an equal amount of water after removing non-biodegradable materials and then pulverized using a homogenizer. The physicochemical properties of the slurry were as follows: pH 5.2; chemical oxygen demand (COD), approx. 203 g L−1; and volatile suspended solids (VSS), approx. 104 g L−1.
Enrichment from the sludge
The basal medium was used with or without 0.01% (w/v) yeast extract (Becton Dickinson, Franklin Lakes, NJ, USA), designated YB and B media, respectively, in this study. Basal medium was composed of (L−1): 0.54 g NH4Cl, 0.14 g KH2PO4, 0.20 g MgCl2·6H2O, 0.15 g CaCl2·2H2O, 2.5 g NaHCO3, and 1.0 mL trace element solution (58) containing 4.0 mg Na2WO4·H2O and eliminating NaCl. Prior to inoculation, the pH of the medium was adjusted to 7.0 with 6 N HCl, dissolved oxygen was removed by flushing with N2:CO2 (4:1, v/v), and 10 mL vitamin solution (L−1) (60) and 10 mL sterile stock solution of Na2S/cysteine-HCl solution (each 50.0 g L−1) (26) were added. H2:CO2 (4:1, v/v; approx. 150 kPa), formate, acetate, or methanol (all at 10 mM) was added to the basal medium as the sole substrate. For enrichment, 0.5 mL anaerobic sludge was inoculated into 20 mL of each medium and incubated at 30°C for a week. A stable enrichment culture was obtained after three cultivations in MYB medium, YB medium supplied with methanol. The enrichment culture was maintained in MYB medium by consecutive transfer monthly.
Preparation of DNA, PCR amplification, and DNA sequencing
The genomic DNA was extracted from the enrichment culture and purified as described previously (49). The archaeal and bacterial 16S rRNA genes were amplified by PCR using the following primers: A10F (5′-TCYGGTTGATCCYGCCRG-3′) and A1400R (5′-ACGGGCGGTGTGTGCAAG-3′) for the domain Archaea, U27F and U1492R (25) for the domain Bacteria. The PCR mixture (50 μL) contained 1×PCR buffer, 3.5 mM MgCl2, 10 mM deoxynucleoside triphosphates (dNTPs), 1.25 U AmpliTaq Gold (each from Applied Biosystems, Foster City, CA, USA), and 0.4 μM of each forward and reverse primer. Approximately 100 ng genomic DNA was used as a template under the following cycling conditions: initial AmpliTaq Gold activation at 95°C for 9 min, followed by 30 cycles of denaturation at 95°C for 30 s, annealing at 56°C for 30 s, extension at 72°C for 1.5 min, and a final extension step at 72°C for 5 min. The mcrA gene encoding the alpha-subunit of methyl-coenzyme M reductase was also partially amplified by PCR with primers MR1mod and ME2mod (35) under almost the same PCR conditions except for its cycle number (40 cycles) and time of extension step (1 min) in the cycle. The PCR product was purified using the QIAquick PCR purification kit (Qiagen, Hilden, Germany), and sequenced using the BigDye terminator v3.1 cycle sequencing kit with a 3130xl genetic analyzer (both from Applied Biosystems).
Clone library
The purified archaeal 16S rRNA and mcrA genes were cloned with a pT7Blue T-vector kit (Novagen, Madison, WI, USA). The clonal DNAs were amplified from randomly selected recombinants by direct PCR with M13 primers, and then used as templates for sequencing. A universal primer 907r (56) and T7 promoter primers were used for sequencing the cloned 16S rRNA and mcrA genes, respectively. The obtained sequences of all the 16S rRNA gene clones (~690 bp) and the mcrA gene clones (~475 bp) were compared with those in the GenBank database using the BLAST program (NCBI-BLAST, www.ncbi.nlm.nih.gov/BLAST), and aligned using the CLUSTAL_X program. Sequence identity of 99% was used as the cut-off value for grouping the sequences into different operational taxonomic units (OTUs).
Phylogenetic analyses
Almost full-length 16S rRNA gene and partial mcrA gene sequences were determined for phylogenetic analysis. The following primers were used for sequencing the PCR product of the archaeal 16S rRNA gene: A10F, Kjm700F (5′-TGGGGTAGGGGTAAA ATCCT-3′), Kjm1000F (5′-ACTCACCAGGGGAGACTGTT-3′), A500R (5′-GTGTTACCGCGGCKGCTGG-3′), Kjm700R (5′-GTG GTCCTTCTAGGATTACA-3′), and A1400R, that of the bacterial 16S rRNA gene: U520F (5′-GTGCCAGCAGCCGCGG-3′) and U1492R, and that of mcrA gene: MR1mod and ME2mod. Sequences were compared using the BLAST program with those available in the DDBJ/EMBL/GenBank databases. Phylogenetic analyses were carried out using the 16S rRNA gene sequence and deduced amino acid sequence of the mcrA gene. The 16S rRNA gene sequences were aligned with an ARB data set using ARB software (32). According to the previously described method (27), thirty-five reference sequences of the phylogenetically related archaea and environmental clones were selected as authentic sequences located in the class Thermoplasmata. The data set of deduced McrA amino acid sequences was aligned using the CLUSTAL_X program. Phylogenetic trees were constructed by the neighbor-joining (NJ) method with the CLUSTAL_X program (42, 57) and the maximum-likelihood (ML) method with MORPHY software version 2.3b3 (10, 20). In addition, the posterior probabilities of branching points were estimated by Bayesian inference using MrBayes 3.1 (23, 41).
Fluorescence in situ hybridization
The enriched archaeon Kjm51a grew on the aforementioned MYB medium for 8 d. The harvested cells were fixed in 4% paraformaldehyde at 4°C for 2 h and stored in 99% ethanol–phosphate-buffered saline (1:1). The fixed cells were incubated in hybridization buffer (0.9 M NaCl, 0.01% sodium dodecyl sulfate, 20 mM Tris-HCl, pH 7.2, and an appropriate amount of formamide) containing fluorescently labeled probes (0.5 pmol μL−1). After incubation at 46°C for 10 h, the buffer was replaced with washing solution (378 mM NaCl, 0.01% sodium dodecyl sulfate, 20 mM Tris-HCl, pH 7.2, and 5 mM EDTA). The sample was incubated at 46°C for 20 min. and then stained with 1 μg mL−1 of 4′,6-diamidino-2-phenylindole (DAPI). The sample obtained was observed under a confocal laser scanning microscope (LSM710; Carl Zeiss Microscopy, Tokyo, Japan). A specific oligonucleotide probe targeting the 16S rRNA gene of the enriched archaeon Kjm51a (RC281r2, 5′-AAGGCCCATACCCGTCATC-3′) was designed using the Probe Design tool of the ARB software package (32). The overall Gibbs free energy of this probe and target sequence calculated with the mathFISH web server was −9.2 kcal mol−1 (62). The probes were labeled with fluorescent dye, Alexa Fluor 555 (Japan Bio Services, Saitama, Japan). Two domain-specific probes were also used: EUB338 labeled with Alexa Fluor 647 for detection of almost all bacteria (2), and ARC915 labeled with Alexa Fluor 488 for detection of almost all archaea (53). The stringency of hybridization was adjusted by adding formamide to the hybridization buffer (15% [v/v] for all the probes used in this study). More than 8,000 DAPI-stained cells were counted to determine the ratio of ARC915-hybridized cells to EUB338-hybridized cells.
Physiological characteristics
Growth conditions were determined using MYB medium. Aerobic and microaerobic conditions were prepared by the substitution of air and the addition of 2% (v/v) oxygen, respectively, with filtration through a 0.2 μm-pore membrane filter. Prior to inoculation, acetate, lactate, or pyruvate (all at 10 mM) were added as carbon sources instead of yeast extract. 2-Bromo ethanesulfonate (BES, final concentration 20 mM) was added as the inhibitor of methane production. Then, 0.2 mL of the preculture of the enrichment was inoculated into 20 mL fresh medium containing each substrate. The culture was incubated at 30°C for two weeks. After the transfer twice, hydrogen and methane concentrations in the headspaces of serum bottles were determined with a gas chromatograph (GC-14A; Shimadzu, Kyoto, Japan) equipped with a thermal conductivity detector and a porapack Type Q 80–100, mesh 80–100 (Waters, Tokyo, Japan). The analysis conditions were as follows; column temperature, 60°C; injector temperature, 80°C; and detector temperature, 100°C; current, 80 mA; carrier gas, N2.
Accession numbers
The 16S rRNA gene and mcrA gene sequences of the enriched archaeon Kjm51a have been deposited in the DDBJ/EMBL/NCBI, and GenBank nucleotide sequence databases under accession numbers AB749767 and AB749768, respectively.
Results
Methanogenic enrichment cultures from anaerobic sludge
A methanogenic enrichment culture was obtained from anaerobic digester using MYB medium containing methanol and yeast extract. The archaeal population in the culture was analyzed using archaeal 16S rRNA gene- and mcrA gene-specific primers. A total of 113 and 61 clones were obtained, respectively. A sole phylotype was obtained in both clone libraries, i.e., all the cloned 16S rRNA gene and mcrA gene sequences in the two libraries were almost identical to the sequence identities of 99.4–100% and 99.5–100%, respectively.
An almost full-length 16S rRNA gene sequence (1,309 bp) was determined for a novel archaeon, designated phylotype Kjm51a, in the enrichment culture. In the phylogenetic trees of the 16S rRNA gene sequences constructed using NJ, ML, and Bayesian methods, the enriched archaeon Kjm51a was placed into an uncultured archaeal lineage, Group E2, in the class Thermoplasmata (6) (Fig. 1). The topologies of the trees generated by the three phylogenetic analysis methods were almost identical, and were supported by high bootstrap values (99–100%). Archaeon Kjm51a was a member of RC-III, a sublineage within Group E2, and showed the highest sequence similarities (91.3–96.2%) to the environmental clones derived from anaerobic digesters, bovine rumen, and landfill leachate (13, 22, 54, 61). The nearest cultivated neighbor of archaeon Kjm51a was Methanomassiliicoccus luminyensis B10T with 87.7% sequence identity. A partial mcrA gene sequence (1,109 bases) was also determined for the enriched archaeon Kjm51a. The NJ tree constructed using the McrA amino acid sequence deduced from the mcrA gene sequence demonstrated that archaeon Kjm51a formed a monophyletic cluster together with M. luminyensis B10T and environmental clone sequences derived from anaerobic bioreactor and bovine rumen (Fig. 2), and that the cluster was apparently distinct from the four known methanogenic lineages, the classes Methanobacteria, ‘Methanomicrobia’, Methanococci, and Methanopyri. The McrA amino acid sequence of the enriched archaeon Kjm51a showed 76.0% identity with that of the closest species, M. luminyensis B10T.
Fig. 1.

Phylogenetic affiliation of the enriched archaeon Kjm51a within Group E2 in the class Thermoplasmata on the basis of the 16S rRNA gene sequences. The tree was constructed using the neighbor-joining method. Solid circles at branching nodes indicate supporting probabilities above 95% by all the phylogenetic analysis methods (NJ, ML, and Bayesian), and open circles indicate probabilities above 85% by two or more analyses. Bar, 0.02 substitutions per nucleotide position.
Fig. 2.

Deduced McrA amino acid sequence-based phylogeny showing the phylogenetic relationships among the enriched archaeon Kjm51a, its related archaeon and environmental clones, and other known methanogens. Solid circles at branching nodes indicate supporting probabilities above 95%, and open circles indicate probabilities above 80%. Bar, 0.02 substitutions per nucleotide position.
Coccoid- and rod-shaped cells were observed under the microscope. The cocci and rods were identified as archaeal and bacterial cells, respectively, by fluorescence in situ hybridization with archaeal and bacterial probes (Fig. 3A and B). A ratio of archaeal cells to total cells was at least 3.5±1.4% in the enrichment culture. Cocci were hybridized with a Kjm51a-specific probe, but rods were not (Fig. 3C). Cells hybridized with the Kjm51a-specific probe completely corresponded to those with the archaeal probe (Fig. 3B and D). The bacterial rods in the enrichment culture were provisionally identified as Clostridium celerecrescens (sequence identity: 99.9%, X71848) by bacterial 16S rRNA gene sequence analysis.
Fig. 3.

Epifluorescence micrographs of in situ hybridization of the enrichment archaeon Kjm51a grown on MYB medium for a week. The same microscopic field is shown after hybridization with a Kjm51a-specific probe (red), an archaeal probe ARC915 (green), and a bacterial probe EUB338 (blue). A, blue color; B, green color; C, merge of red, green and blue colors; D, red color. Bars, 10 μm.
Physiological property of the enriched archaeon Kjm51a
The enriched archaeon Kjm51a was strictly anaerobic and was capable of growing in MYB medium under a N2/CO2 (4:1 [v/v]) atmosphere, but could not grow under microaerobic or aerobic conditions. Both methanol and yeast extract were required for the growth of archaeon Kjm51a. Acetate, lactate, and pyruvate were not utilized as carbon sources instead of yeast extract. Metabolic products of the enrichment culture with methanol and yeast extract were hydrogen and methane, and that with yeast extract and without methanol was hydrogen (Fig. 4). Growth of the enriched archaeon Kjm51a in the presence of methanol and its methane production were completely inhibited by the addition of BES. The amount of hydrogen in the presence of BES was approximately three times higher than that in the absence of BES.
Fig. 4.
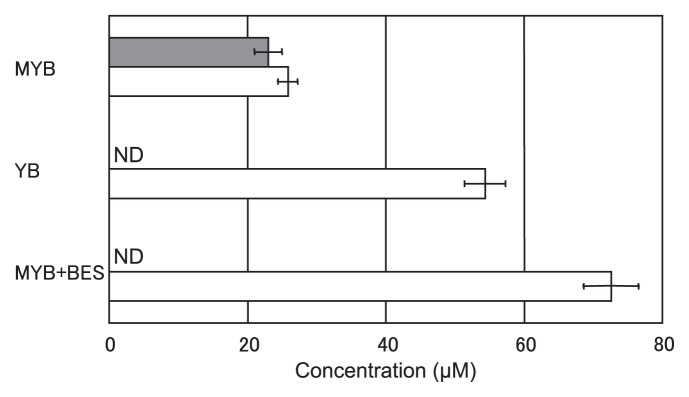
Methane and hydrogen production in the enrichment culture obtained from anaerobic sludge. Filled bars, methane; open squares, hydrogen. Data points and bars are the means and standard deviations, respectively (n=3). Abbreviations: MYB, MYB medium supplied with methanol in YB medium; YB, YB medium; MYB+BES, MYB medium supplied with BES; ND, not detected.
Discussion
The novel archaeon Kjm51a was successfully enriched from anaerobic sludge using MYB medium containing methanol and yeast extract. The archaeon was not yet purified in this study because C. celerecrescens was dominantly isolated in the presence of yeast extract, although we made a great effort to isolate the archaeon using agar plate culture and dilution-extinction culture. Clone library analysis demonstrated that cloned 16S rRNA and mcrA gene sequences obtained from the enrichment culture were almost identical in each. Furthermore, FISH analysis also showed that coccal cells hybridized with a Kjm51a-specific probe completely corresponded to those with an archaeal probe. These findings strongly support the archaeal purity of the enriched archaeon Kjm51a in MYB medium, although bacterial cells still coexisted.
The enriched archaeon Kjm51a was a strictly anaerobic and chemoheterotrophic cocci showing growth and methane production in the presence of methanol, the inhibition of methane production by BES, a well-known inhibitor of methanogenesis (18). Hydrogen production was also observed in the absence of methanol, which indicated that the coexisting bacterium, C. celerecrescens, produced hydrogen by its fermentation (38). Inhibition of methane production by BES resulted in increased hydrogen production. These physiological properties suggest that the enriched archaeon Kjm51a might be a methanol-reducing hydrogenotrophic methanogen.
As reported for Methanosphaera stadtmanae (34), Methanomicrococcus blatticola (51), and Methanosarcina barkeri strain Fusaro (36), the enriched archaeon Kjm51a is likely to produce methane by the hydrogen-dependent reduction of methanol through the following reaction: H2+ CH3OH → CH4+H2O (11, 29, 52, 59). Methanomassiliicoccus luminyensis, a recently isolated methanogen from human feces, belonging to Group E2, also produced methane from methanol in the presence of hydrogen (9). The genome of M. luminyensis likely encodes only a partial methanogenesis pathway (16). Most recently, archaeon MpT1 in Group E2 was enriched from termite guts as a methanogen, which converted methanol to methane (39). Methanol may be a common substrate for methanogenesis in Group E2. Schink and Zeikus reported that heterotrophic microbes anaerobically produced methanol as a major end product from pectin, which is a component of plant tissue (48). Biodegradation of plants occurs in a wide variety of environments, such as the rumen, rice field soil and anaerobic digester treating garbage, and Group E2 methanogens may contribute to carbon flux.
The enriched archaeon Kjm51a is the first culture representative derived from an anaerobic methanogenic digester in Group E2. In the phylogenetic trees constructed using 16S rRNA gene sequences, the archaeon Kjm51a and M. luminyensis B10T were completely separated in Group E2 with their low sequence identity (87.7%), which was sufficiently low to classify them into different genera. The enriched archaeon Kjm51a and M. luminyensis B10T were affiliated with RC-III, one of the sublineages in Group E2. RC-III was clearly and completely separated from the validly described order Thermoplasmatales. Its monophyletic lineage was strongly supported by the probability scores (>99%) calculated using all the phylogenetic analysis methods. The 16S rRNA gene sequence of the enriched archaeon Kjm51a and M. luminyensis B10T had similarities of only 77.1 to 80.3% with those of the known archaeal species in the order Thermoplasmatales. These similarities are lower than the 85% similarity that is generally used as a cut-off value for distinguishing lineages at the phylum, as suggested by Hugenholtz et al. (24). Therefore, RC-III composed of the enriched culture Kjm51a and M. luminyensis B10T is a distinct order level lineage in the class Thermoplasmata. Previously, Kemnitz et al. (30) reported that the RC-III archaea might heterotrophically grow using peptides, based on their enrichment culture experiment; however, taken together with the recent study (6), our findings clearly indicated that RC-III is a novel methanogenic lineage.
In conclusion, an archaeal representative enriched from the anaerobic methanogenic digester is a novel methanogen belonging to RC-III within Group E2 in the class Thermoplasmata. According to the recommendations of Murray and Stackebrandt (37), we propose the provisional taxonomic assignment of Candidatus Methanogranum caenicola for the enriched archaeon Kjm51a. Most recently, the order Methanoplasmatales was provisionally proposed for the deep-branching lineage accommodating M. luminyensis and the enriched archaea MpT1 and MpM2 (39); however, this lineage should be proposed as the Methanomassiliicoccales on the basis of Rule 47a of the Bacteriological Code to avoid bacteriological confusion (31). Consequently, we propose to rename the order ‘Methanoplasmatales’ as Methanomassiliicoccales for the sublineage accommodating M. luminyensis B10T and the enriched archaeon Kjm51a as described below. The proposal of this novel order follows the description of the new family Methanomassiliicoccaceae. To purify the enriched archaeon Kjm51a and understand its ecological role in the methanogenic environment, further study via enrichment culture will be necessary.
Description of Candidatus Methanogranum caenicola
Methanogranum caenicola (Me.tha.no.gra’num. cae.ni. co’la. N.L. n. methanum [from French n. méth(yle) and chemical suffix -ane], methane; N.L. pref. methano-, pertaining to methane; L. neut. n. granum, grain, kernel; N.L. neut. n. Methanogranum, a methane-producing grain: L. n. caenum, mud, sludge; L. suff. -cola [from L. n. incola], inhabitant, dweller; N.L. n. caenicola, an inhabitant of sludge).
Strictly anaerobic, chemoheterotrophic. Cells form cocci occurring as single cells. Produce methane dependent on hydrogen and methanol. Represent a distinct phylogenetic lineage in the class Thermoplasmata based on 16S rRNA gene sequence analysis. Enriched from an anaerobic sludge in a methanogenic digester.
Description of Methanomassiliicoccaceae fam. nov
Methanomassiliicoccaceae (Me.tha.no.mas.si.li.i.coc.ca’ce.ae. N.L. neut. n. Methanomassiliicoccus type genus of the family; -aceae ending to denote a family; N.L. fem. pl. n. Methanomassiliicoccaceae family of the genus Methanomassiliicoccus).
The family Methanomassiliicoccaceae is defined on the basis of a phylogenetic tree constructed by phylogenetic analysis of the 16S rRNA gene sequence of a single cultivated representative, of the enriched culture, and of environmental clone sequences derived mainly from the alimentary canal, anaerobic digester, landfill leachate, and rice field soil. The type genus is Methanomassiliicoccus.
Description of Methanomassiliicoccales ord. nov
Methanomassiliicoccales (Me.tha.no.mas.si.li.i.coc.cal’es. N.L. neut. n. Methanomassiliicoccus type genus of the order; -ales ending to denote an order; N.L. fem. pl. n. Methanomassiliicoccales order of the genus Methanomassiliicoccus).
The description is the same as that for the family Methanomassiliicoccaceae. The type genus is Methanomassiliicoccus.
Acknowledgements
The authors thank Dr. Hiroyuki Imachi (Japan Agency for Marine-Earth Science & Technology) for helpful discussions on several aspects of the paper.
References
- 1.Akuzawa M, Hori T, Haruta S, Ueno Y, Ishii M, Igarashi Y. Distinctive responses of metabolically active microbiota to acidification in a thermophilic anaerobic digester. Microb Ecol. 2011;61:595–605. doi: 10.1007/s00248-010-9788-1. [DOI] [PubMed] [Google Scholar]
- 2.Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, Stahl DA. Combination of 16S rRNA-targeted oligo-nucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl Environ Microbiol. 1990;56:1919–1925. doi: 10.1128/aem.56.6.1919-1925.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chin K-J, Thomas L, Conrad R. Effect of temperature on structure and function of the methanogenic archaeal community in an anoxic rice field soil. Appl Environ Microbiol. 1999;65:2341–2349. doi: 10.1128/aem.65.6.2341-2349.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Darland G, Brock TD, Samsonoff W, Conti SF. A thermophilic, acidophilic mycoplasma isolated from a coal refuse pile. Science. 1970;170:1416–1418. doi: 10.1126/science.170.3965.1416. [DOI] [PubMed] [Google Scholar]
- 5.DeLong EF. Archaea in coastal marine environments. Proc Natl Acad Sci USA. 1992;89:5685–5689. doi: 10.1073/pnas.89.12.5685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.DeLong EF, Pace NR. Environmental diversity of bacteria and archaea. Syst Biol. 2001;50:470–478. [PubMed] [Google Scholar]
- 7.DeLong EF, Wu KY, Prezelin BB, Jovine RV. High abundance of Archaea in Antarctic marine picoplankton. Nature. 1994;371:695–697. doi: 10.1038/371695a0. [DOI] [PubMed] [Google Scholar]
- 8.Dojka MA, Hugenholtz P, Haack SK, Pace NR. Microbial diversity in a hydrocarbon- and chlorinated-solvent-contaminated aquifer undergoing intrinsic bioremediation. Appl Environ Microbiol. 1998;64:3869–3877. doi: 10.1128/aem.64.10.3869-3877.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dridi B, Fardeau M-L, Ollivier B, Raoult D, Drancourt M. Methanomassiliicoccus luminyensis gen. nov., sp. nov., a methanogenic archaeon isolated from human faeces. Int J Syst Evol Microbiol. 2012;62:1902–1907. doi: 10.1099/ijs.0.033712-0. [DOI] [PubMed] [Google Scholar]
- 10.Felsenstein J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol. 1981;17:368–376. doi: 10.1007/BF01734359. [DOI] [PubMed] [Google Scholar]
- 11.Fricke WF, Seedorf H, Henne A, Krüer M, Liesegang H, Hedderich R, Gottschalk G, Thauer RK. The genome sequence of Methanosphaera stadtmanae reveals why this human intestinal archaeon is restricted to methanol and H2 for methane gormation and ATP synthesis. J Bacteriol. 2006;188:642–658. doi: 10.1128/JB.188.2.642-658.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Friedrich MW, Schmitt-Wagner D, Lueders T, Brune A. Axial differences in community structure of Crenarchaeota and Euryarchaeota in the highly compartmentalized gut of the soil-feeding termite Cubitermes orthognathus. Appl Environ Microbiol. 2001;67:4880–4890. doi: 10.1128/AEM.67.10.4880-4890.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Godon J-J, Zumstein E, Dabert P, Habouzit F, Moletta R. Molecular microbial diversity of an anaerobic digestor as determined by small-subunit rDNA sequence analysis. Appl Environ Microbiol. 1997;63:2802–2813. doi: 10.1128/aem.63.7.2802-2813.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Golyshina OV, Pivovarova TA, Karavaiko GI, et al. Ferroplasma acidiphilum gen. nov., sp. nov., an acidophilic, autotrophic, ferrous-iron-oxidizing, cell-wall-lacking, mesophilic member of the Ferroplasmaceae fam. nov., comprising a distinct lineage of the Archaea. Int J Syst Evol Microbiol. 2000;50:997–1006. doi: 10.1099/00207713-50-3-997. [DOI] [PubMed] [Google Scholar]
- 15.Golyshina OV, Yakimov MM, Lünsdorf H, Ferrer M, Nimtz M, Timmis KN, Wray V, Tindall BJ, Golyshin PN. Acidiplasma aeolicum gen. nov., sp. nov., a euryarchaeon of the family Ferroplasmaceae isolated from a hydrothermal pool, and transfer of Ferroplasma cupricumulans to Acidiplasma cupricumulans comb. nov. Int J Syst Evol Microbiol. 2009;59:2815–2823. doi: 10.1099/ijs.0.009639-0. [DOI] [PubMed] [Google Scholar]
- 16.Gorlas A, Robert C, Gimenez G, Drancourt M, Raoult D. Complete genome sequence of Methanomassiliicoccus luminyensis, the largest genome of a human-associated Archaea species. J Bacteriol. 2012;194:4745. doi: 10.1128/JB.00956-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Großkpof R, Stubner S, Liesack W. Novel euryarchaeotal lineages detected on rice roots and in the anoxic bulk soil of flooded rice microcosms. Appl Environ Microbiol. 1998;63:4983–4989. doi: 10.1128/aem.64.12.4983-4989.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gunsalus RP, Romesser JA, Wolfe RS. Preparation of coenzyme M analogues and their activity in the methyl coenzyme M reductase system of Methanobacterium thermoautotrophicum. Biochemistry. 1978;17:2374–2377. doi: 10.1021/bi00605a019. [DOI] [PubMed] [Google Scholar]
- 19.Hara K, Shinzato N, Seo M, Oshima T, Yamagishi A. Phylogenetic analysis of symbiotic archaea living in the gut of xylophagous cockroaches. Microb Environ. 2002;17:185–190. [Google Scholar]
- 20.Hasegawa M, Kishino H, Yano TA. Dating of the human ape splitting by a molecular clock of mitochondrial-DNA. J Mol Evol. 1985;22:160–174. doi: 10.1007/BF02101694. [DOI] [PubMed] [Google Scholar]
- 21.Hori T, Haruta S, Ueno Y, Ishii M, Igarashi Y. Dynamic transition of a methanogenic population in response to the concentration of volatile fatty acids in a thermophilic anaerobic digester. Appl Environ Microbiol. 2006;72:1623–1630. doi: 10.1128/AEM.72.2.1623-1630.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Huang LN, Chen YQ, Zhou H, Luo S, Lan CY, Qu LH. Characterization of methanogenic Archaea in the leachate of a closed municipal solid waste landfill. FEMS Microbiol Ecol. 2003;46:171–177. doi: 10.1016/S0168-6496(03)00218-6. [DOI] [PubMed] [Google Scholar]
- 23.Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001;17:754–755. doi: 10.1093/bioinformatics/17.8.754. [DOI] [PubMed] [Google Scholar]
- 24.Hugenholtz P, Pitulle C, Hershberger KL, Pace NR. Novel division level bacterial diversity in a Yellowstone hot spring. J Bacteriol. 1998;180:366–376. doi: 10.1128/jb.180.2.366-376.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Iino T, Mori K, Tanaka K, Suzuki K, Harayama S. Oscillibacter valericigenes gen. nov., sp. nov., a valerate-producing anaerobic bacterium isolated from the alimentary canal of a Japanese corbicula clam. Int J Syst Evol Microbiol. 2007;57:1840–1845. doi: 10.1099/ijs.0.64717-0. [DOI] [PubMed] [Google Scholar]
- 26.Iino T, Suzuki K, Harayama S. Lacticigenium naphtae gen. nov., sp. nov., a novel halotolerant and motile lactic acid bacterium isolated from crude oil. Int J Syst Evol Microbiol. 2009;59:775–780. doi: 10.1099/ijs.0.003293-0. [DOI] [PubMed] [Google Scholar]
- 27.Iino T, Mori K, Uchino Y, Nakagawa T, Harayama S, Suzuki K. Ignavibacterium album gen. nov., sp. nov., a moderately thermophilic anaerobic bacterium isolated from microbial mats at a terrestrial hot spring and proposal of Ignavibacteria classis nov., for a novel lineage at the periphery of green sulfur bacteria. Int J Syst Evol Microbiol. 2010a;60:1376–1382. doi: 10.1099/ijs.0.012484-0. [DOI] [PubMed] [Google Scholar]
- 28.Itoh T, Yoshikawa N, Takashina T. Thermogymnomonas acidicola gen. nov., sp. nov., a novel thermoacidophilic, cell wall-less archaeon in the order Thermoplasmatales, isolated from a solfataric soil in Hakone, Japan. Int J Syst Evol Microbiol. 2007;57:2557–2561. doi: 10.1099/ijs.0.65203-0. [DOI] [PubMed] [Google Scholar]
- 29.Keltjens JT, van der Drift C. Electron transfer reactions in methanogens. FEMS Microbiol Rev. 1986;39:259–303. [Google Scholar]
- 30.Kemnitz D, Kolb S, Conrad R. Phenotypic characterization of Rice Cluster III archaea without prior isolation by applying quantitative polymerase chain reaction to an enrichment culture. Environ Microbiol. 2005;7:553–565. doi: 10.1111/j.1462-2920.2005.00723.x. [DOI] [PubMed] [Google Scholar]
- 31.Lapage SP, Sneath PHA, Lessel EF, Skerman VBD, Seeliger HPR, Clark WA, editors. International Code of Nomenclature of Bacteria, 1990 Revision. Published for the International Union of Microbiological Societies by the American Society for Microbiology; Washington, D.C: 1992. PHA Sneath editor for 1992 edition. [Google Scholar]
- 32.Ludwig W, Strunk O, Westram R, et al. ARB: a software environment for sequence data. Nucleic Acids Res. 2004;32:1363–1371. doi: 10.1093/nar/gkh293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mihajlovski A, Alric M, Brugére JF. A putative new order of methanogenic Archaea inhabiting the human gut, as revealed by molecular analyses of the mcrA gene. Res Microbiol. 2008;159:516–521. doi: 10.1016/j.resmic.2008.06.007. [DOI] [PubMed] [Google Scholar]
- 34.Miller TL, Wollin MJ. Methanosphaera stadtmaniae gen. nov., sp. nov.: a species that forms methane by reducing methanol with hydrogen. Arch Microbiol. 1985;141:116–122. doi: 10.1007/BF00423270. [DOI] [PubMed] [Google Scholar]
- 35.Mori K, Iino T, Suzuki K, Yamaguchi K, Kamagata Y. Aceticlastic and NaCl-requiring methanogen “Methanosaeta pelagica” sp. nov., isolated from marine tidal flat sediment. Appl Environ Microbiol. 2012;78:3416–3423. doi: 10.1128/AEM.07484-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Müller V, Blaut M, Gottschalk G. Utilization of methanol plus hydrogen by Methanosarcina barkeri for methanogenesis and growth. Appl Environ Microbiol. 1986;52:269–274. doi: 10.1128/aem.52.2.269-274.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Murray RGE, Stackebrandt E. Taxonomic Note: Implementation of the provisional status Candidatus for incompletely described procaryotes. Int J Syst Bacteriol. 1995;45:186–187. doi: 10.1099/00207713-45-1-186. [DOI] [PubMed] [Google Scholar]
- 38.Palop MLL, Valles S, Piñaga F, Flors A. Isolation and characterization of an anaerobic, cellulolytic bacterium, Clostridium celerecrescens sp. nov. Int J Syst Bacteriol. 1989;39:68–71. [Google Scholar]
- 39.Paul K, Nonoh JO, Mikulski L, Brune A. “Methanoplasmatales,” thermoplasmatales-related archaea in termite guts and other environments, are the seventh order of methanogens. Appl Environ Microbiol. 2012;78:8245–8253. doi: 10.1128/AEM.02193-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Reysenbach AL, Longnecker K, Kirshtein J. Novel bacterial and archaeal lineages from an in situ growth chamber deployed at a Mid-Atlantic Ridge hydrothermal vent. Appl Environ Microbiol. 2000;66:3798–3806. doi: 10.1128/aem.66.9.3798-3806.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–1574. doi: 10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- 42.Saitou N, Nei M. A neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–425. doi: 10.1093/oxfordjournals.molbev.a040454. [DOI] [PubMed] [Google Scholar]
- 43.Sasaki K, Haruta S, Ueno Y, Ishii M, Igarashi Y. Archaeal population on supporting material in methanogenic packed-bed reactor. J Biosci Bioeng. 2006a;102:244–246. doi: 10.1263/jbb.102.244. [DOI] [PubMed] [Google Scholar]
- 44.Sasaki K, Haruta S, Tatara M, Yamazawa A, Ueno Y, Ishii M, Igarashi Y. Microbial community in methanogenic packed-bed reactor successfully operating at short hydraulic retention time. J Biosci Bioeng. 2006b;101:271–273. doi: 10.1263/jbb.101.271. [DOI] [PubMed] [Google Scholar]
- 45.Sasaki K, Haruta S, Ueno Y, Ishii M, Igarashi Y. Microbial population in the biomass adhering to supporting material in a packed-bed reactor degrading organic solid waste. Appl Microbiol Biotechnol. 2007;75:941–952. doi: 10.1007/s00253-007-0888-x. [DOI] [PubMed] [Google Scholar]
- 46.Sasaki D, Hori T, Haruta S, Ueno Y, Ishii M, Igarashi Y. Methanogenic pathway and community structure in a thermophilic anaerobic digestion process of organic solid waste. J Biosci Bioeng. 2011;111:41–46. doi: 10.1016/j.jbiosc.2010.08.011. [DOI] [PubMed] [Google Scholar]
- 47.Schleper C, Pühler G, Klenk HP, Zillig W. Picrophilus oshimae and Picrophilus torridus fam. nov., gen. nov., sp. nov., two species of hyperacidophilic, thermophilic, heterotrophic, aerobic archaea. Int J Syst Bacteriol. 1996;46:814–816. [Google Scholar]
- 48.Schink B, Zeikus JG. Microbial methanol formation: a major end product of pectin metabolism. Curr Microbiol. 1980;4:387–389. [Google Scholar]
- 49.Schmidt TM, DeLong EF, Pace NR. Analysis of a marine picoplankton community by 16S rRNA gene cloning and sequencing. J Bacteriol. 1991;173:4371–4378. doi: 10.1128/jb.173.14.4371-4378.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shouten S, Baas M, Hopmans EC, Reysenbach A-L, Damsé JSS. Tetraether membrane lipids of Candidatus “Aciduliprofundum boonei”, a cultivated obligate thermoacidophilic euryarchaeote from deep-sea hydrothermal vents. Extremophiles. 2008;12:119–124. doi: 10.1007/s00792-007-0111-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sprenger WW, van Belzen MC, Rosenberg J, Hackstein JHP, Keltjens JT. Methanomicrococcus blatticola gen. nov., sp. nov., a methanol- and methylamine-reducing methanogen from the hindgut of the cockroach Periplaneta Americana. Int J Syst Evol Microbiol. 2000;50:1989–1999. doi: 10.1099/00207713-50-6-1989. [DOI] [PubMed] [Google Scholar]
- 52.Sprenger WW, Hackstein JHP, Keltjens JT. The energy metabolism of Methanomicrococcus blatticola: physiological and biochemical aspects. Antonie van Leeuwenhoek. 2005;87:289–299. doi: 10.1007/s10482-004-5941-5. [DOI] [PubMed] [Google Scholar]
- 53.Stahl DA, Amann R. Development and application of nucleic acid probes. In: Stackbrandt E, Goodfellow M, editors. Nucleic Acid Techniques in Bacterial Systematics. John Wiley & Sons; New York: 1991. pp. 205–248. [Google Scholar]
- 54.Tajima K, Nagamine T, Matsui H, Nakamura M, Aminov RI. Phylogenetic analysis of archaeal 16S rRNA libraries from the rumen suggests the existence of a novel group of archaea not associated with known methanogens. FEMS Microbiol Lett. 2001;200:67–72. doi: 10.1111/j.1574-6968.2001.tb10694.x. [DOI] [PubMed] [Google Scholar]
- 55.Takai K, Horikoshi K. Genetic diversity of archaea in deep-sea hydrothermal vent environments. Genetics. 1999;152:1285–1297. doi: 10.1093/genetics/152.4.1285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tamaki H, Sekiguchi Y, Hanada S, Nakamura K, Nomura N, Matsumura M, Kamagata Y. Comparative analysis of bacterial diversity in freshwater sediment of a shallow eutrophic lake by molecular and improved cultivation-based techniques. Appl Environ Microbiol. 2005;71:2162–2169. doi: 10.1128/AEM.71.4.2162-2169.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The Clustal_X Windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tool. Nucleic Acids Res. 1997;24:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Touzel JP, Albagnac G. Isolation and characterization of Methanococcus mazei strain MC3. FEMS Microbiol Lett. 1983;16:241–245. [Google Scholar]
- 59.van de Wijngaard WMH, Creemers J, Vogels GD, van der Drift C. Methanogenic pathways in Methanosphaera stadtmanae. FEMS Microbiol Lett. 1991;80:207–212. doi: 10.1016/0378-1097(91)90596-3. [DOI] [PubMed] [Google Scholar]
- 60.Wolin EA, Wolin MJ, Wolfe RS. Formation of methane by bacterial extracts. J Biol Chem. 1963;238:2882–2886. [PubMed] [Google Scholar]
- 61.Xia Y, Masse DI, McAllister TA, Kong Y, Seviour R, Beaulieu C. Identity and diversity of archaeal communities during anaerobic co-digestion of chicken feathers and other animal wastes. Bioresour Technol. 2012;110:111–119. doi: 10.1016/j.biortech.2012.01.107. [DOI] [PubMed] [Google Scholar]
- 62.Yulmaz JS, Noguera DR. Mechanistic approach to the problem of hybridization efficiency in fluorescent in situ hybridization. Appl Environ Microbiol. 2004;70:7126–7139. doi: 10.1128/AEM.70.12.7126-7139.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]