

Abstract
Most models of dystonia pathophysiology predict alterations of activity in the basal ganglia thalamocortical motor circuit. The globus pallidus interna (GPi) shows bursting and oscillatory neuronal discharge in both human dystonia and in animal models, but it is not clear which intrinsic basal ganglia pathways are implicated in this abnormal output. The subthalamic nucleus (STN) receives prominent excitatory input directly from cortical areas implicated in dystonia pathogenesis and inhibitory input from the external globus pallidus. The goal of this study was to elucidate the role of the STN in dystonia by analyzing STN neuronal discharge in patients with idiopathic dystonia. Data were collected in awake patients undergoing microelectrode recording for implantation of STN deep brain stimulation electrodes. We recorded 62 STN neurons in 9 patients with primary dystonia. As a comparison group, we recorded 143 STN neurons in 20 patients with Parkinson's disease (PD). Single-unit activity was discriminated off-line by principal component analysis and evaluated with respect to discharge rate, bursting, and oscillatory activity. The mean STN discharge rate in dystonia patients was 26.3 Hz (SD 13.6), which was lower than that in the PD patients (35.6 Hz, SD 15.2), but higher than published values for subjects without basal ganglia dysfunction. Oscillatory activity was found in both disorders, with a higher proportion of units oscillating in the beta range in PD. Bursting discharge was a prominent feature of both dystonia and PD, whereas sensory receptive fields were expanded in PD compared with dystonia. The STN firing characteristics, in conjunction with those previously published for GPi, suggest that bursting and oscillatory discharge in basal ganglia output may be transmitted via pathways involving the STN and provide a pathophysiologic rationale for STN as a surgical target in dystonia.
INTRODUCTION
Dystonia is a movement disorder defined by sustained muscle contractions, resulting in involuntary twisting, repetitive movements, and abnormal postures (Fahn et al. 1998). Several lines of evidence implicate the basal ganglia (BG) as key structures in the pathophysiology of dystonia (Chiken et al. 2008; Dang et al. 2005; Marsden et al. 1985; Perlmutter et al. 1997a,b; Playford et al. 1993). Early models of dystonia pathophysiology focused on changes in the relative contributions of the “direct” and “indirect” intrinsic BG pathways (Mink 1996, 2003; Sanger 2003; Vitek 2002; Vitek et al. 1999). In human dystonia patients undergoing microelectrode-guided pallidal surgery in the awake state, globus pallidus interna (GPi) neurons show bursting discharge and 2- to 30-Hz oscillations that are also seen in Parkinson's disease (PD) (Hutchison et al. 2003; Merello et al. 2004; Starr et al. 2005; Tang et al. 2007; Vitek et al. 1999). The GPi, however, reflects the final stage of BG processing. Thus GPi recording cannot distinguish the relative contributions of intrinsic BG pathways to BG output.
Several lines of evidence prompt a reexamination of the role of subthalamic nucleus (STN) in dystonia. First, the STN in primates receives substantial direct cortical projections, via the “hyperdirect” pathway (Nambu et al. 2002), from wide areas of the frontal lobe (Kitai and Deniau 1981; Monakow et al. 1978; Nambu et al. 1996, 1997). Cortical areas projecting to the STN have been implicated in dystonia pathophysiology (Carbon and Eidelberg 2009; Draganski et al. 2003; Garraux et al. 2004; Ikoma et al. 1996; Quartarone et al. 2003, 2008; Toro et al. 2000). Second, the STN has strong reciprocal connections with the globus pallidus externus (GPe), which in turn is modulated by D2 receptor-positive striatal cells. Human functional imaging has demonstrated abnormal striatal D2 binding in patients with idiopathic focal and segmental dystonia using [123I]iodobenzamide, [123I]epidepride, and [123I]-beta-CIT SPECT (Hierholzer et al. 1994; Naumann et al. 1998) and [18F]spiperone positron emission tomography (PET) imaging (Perlmutter et al. 1997a), as well as in asymptomatic carriers of the DYT1 dystonia mutation using [11C]raclopride PET imaging (Asanuma et al. 2005). Studies of nonhuman primates (NHPs) have found decreased striatal D2-like specific binding sites during the temporary dystonic phase following treatment with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) (Perlmutter et al. 1997b). These findings suggest that striatal abnormalities in dystonia may be transmitted to the GPe–STN network (Sani et al. 2009).
Single-unit physiology in the STN has been studied in detail in Parkinson's disease (Hutchison et al. 1998; Levy et al. 2000; Lozano et al. 2000; Magarinos-Ascone et al. 2000; Magnin et al. 2000; Rodriguez-Oroz et al. 2001; Romanelli et al. 2004; Theodosopoulos et al. 2003), but only one study to date has reported single-unit recordings from the STN in patients with dystonia (Zhuang et al. 2004). However, this study did not include a nondystonic comparison group and did not analyze oscillatory activity or bursting behavior quantitatively.
Expanded somatosensory receptive fields in the STN are a well-documented feature of the parkinsonian state (Rodriguez-Oroz et al. 2001; Vitek et al. 1998). GPi recording in dystonia animal models (Chiken et al. 2008) and in some human subjects (Lenz et al. 1998; Magarinos-Ascone et al. 2008; Vitek et al. 1999) suggests that the same is true for dystonia, but no studies of STN physiology in humans have addressed this. Here, we study STN single-unit discharge in awake patients with primary dystonia and PD. We tested the hypothesis that spontaneous and movement-related discharge rates, bursting discharge, oscillatory activity, and sensory receptive fields distinguish dystonia from PD and used these data to reassess current models of the contribution of STN to the pathophysiology of dystonia and PD.
METHODS
Patient population
Single-unit recordings in the STN were obtained from patients with primary dystonia undergoing physiologic mapping for placement of STN deep brain stimulation (DBS) electrodes. All of the dystonia patients had significant disability, failed medical therapy, and received prior botulinum toxin injections, but no longer obtained significant relief. All subjects provided written informed consent to participate in a protocol approved by the Institutional Review Board and all research-related activities were performed in compliance with national legislation and the World Medical Association Declaration of Helsinki. A subset of subjects simultaneously participated in a clinical study of STN DBS for adult-onset primary dystonia (Ostrem et al. 2008). Presurgical clinical evaluations were obtained by movement disorders neurologists (JLO, LES) in the month prior to surgery and the severity was quantified according to two standard clinical rating scales: 1) the Burke–Fahn–Marsden Dystonia Rating Scale (BFMDRS) (Burke et al. 1985) and 2) the Toronto Western Spasmodic Torticollis Rating Scale (TWSTRS) (Comella et al. 1997).
As a comparison group, patients with Parkinson's disease undergoing microelectrode-guided stereotactic surgery for the placement of STN DBS electrodes were studied. All patients in this group were responsive to levodopa (3,4-dihydroxy-l-phenylalanine) and had either levodopa-induced dyskinesias, motor fluctuations, or medically intractable tremor. The severity of disease was assessed prior to surgery according to the Hoehn and Yahr Parkinson's disease staging system. Antiparkinsonian medications were withheld for ≥12 h before the surgery and all PD patients displayed overt parkinsonian symptoms without dyskinesias during the procedure. PD patients with off-period dystonia were excluded from study.
Surgical procedures and data collection
The methods of microelectrode-guided stereotactic surgery for the implantation of DBS electrodes in the STN were similar to those previously described (Starr 2002; Starr et al. 2002). Single-unit recordings were obtained using glass-coated platinum/iridium microelectrodes with impedance 0.4–1.0 MΩ (Microprobe, Gaithersburg, MD; or FHC, Bowdoin, ME). Signals were band-pass filtered (300 Hz to 5 kHz), amplified, played on an audio monitor, displayed on an oscilloscope, and digitized (20-kHz sampling rate) using the Guideline System 3000 or 4000 (FHC) or the Microguide system (Alpha Omega, Nazareth Illit, Israel). Microelectrodes were advanced into the brain using a motorized or manual microdrive (FHC, Alpha Omega, or Elekta, Stockholm). In a typical surgical case, one to two microelectrode penetrations were made serially through the STN on each side, separated by 2–3 mm. Cells were recorded at approximately every 300–800 μm along each trajectory. For well-isolated cells, spontaneous neuronal activity was collected for a minimum of 20 s.
Neurons were screened for movement-related activity based on audible changes in action potential discharge evoked by passive (investigator-initiated) contralateral limb movements (i.e., shoulder, elbow, wrist, hip, knee, and ankle joints). Proprioceptive responsiveness of a neuron in relation to movements of one or more joints was determined by concurrence between the examiner and one or more operating room staff based on audiovisual assessments of the response.
When single-cell isolation could be maintained, neurons that showed a reproducible response to passive movement in the initial multijoint screening were selected for quantitative investigation, focusing only on movement of the joint that produced the maximal modulation of discharge (i.e., a transient increase or decrease in action potential firing that is time-locked with movement) on the initial screen. A triaxial accelerometer (FHC) was strapped to the limb distal to the joint being assessed to indicate the timing of movement onset. Each passive movement (alternating between flexion and extension or internal and external rotation) was followed by 3–5 s of immobility and repeated 6–18 times. Some neurons were also studied while subjects performed voluntary flexion and extension of the identified joint, in alternating fashion, at a rhythm of approximately one cycle/s. Subjects were prompted verbally to begin making the train of movements, but no external cues or commands were provided during their execution.
Prior to recording all patients were sedated with propofol for the surgical incision and skull opening. Propofol was stopped ≥30 min prior to neuronal recording, which is sufficient to wash out its known effect on STN single-unit discharge (Raz et al. 2008). All patients were awake and alert for physiologic mapping and were asked to remain as still as possible with eyes open during periods of neuronal recording.
Data analysis
Digitized spike trains were imported into off-line spike-sorting software (Plexon, Dallas, TX) for discrimination of single populations of action potentials by principal components analysis. The software provided a two-dimensional representation of neuronal waveforms, allowing visual separation of distinct waveform morphologies. This software generated a record of spike times (subsequently reduced to millisecond accuracy) for each action potential waveform detected. The spike times were used to calculate discharge rate, bursting, and oscillatory activity in the 0- to 200-Hz range (see following text). Analyses were performed using Matlab software (The MathWorks, Natick, MA). Neuronal data were included in this study only if action potentials could be discriminated with a high degree of certainty, as indicated by the presence of a clear refractory period in the interspike interval (ISI) histogram (>3 ms, as illustrated in Fig. 1), and if the spontaneous activity of the neuron was recorded for >20 s. Neurons whose action potential morphology varied greatly in synchrony with the cardiac cycle were excluded. In addition, only cells recorded in the dorsal 3 mm of the STN were analyzed, based on prior studies indicating that movement-related activity is generally restricted to the dorsal 3 mm (Rodriguez-Oroz et al. 2001; Romanelli et al. 2004; Theodosopoulos et al. 2003).
Fig. 1.

Examples of single-unit activity from a typical subthalamic nucleus (STN) neuron in a patient with cranial-cervical dystonia (left column) and a patient with the akinetic-rigid subtype of Parkinson's disease (PD, right column). A: neuronal recordings. A 2-s interval is shown. B: interspike interval (ISI) histograms, bin size of 1 ms. Inset: expanded timescale demonstrating the absence of ISIs of <3-ms duration, consistent with the neuronal refractory period. C: raster diagrams showing bursting discharge. Bursts as defined by the Poisson “surprise” method (surprise value >5) are labeled with a black bar above spikes that constitute a burst. Note the higher proportion of bursts per total number of spikes shown in the dystonia neuron (0.40 vs. 0.26 in the PD neuron). Consecutive rows (3 s of data per row) from bottom to top represent continuous 36-s recordings. D: autocorrelograms. The right autocorrelogram shows oscillatory activity of about 11 Hz. The unit on the left was not found to have significant oscillations.
Bursting
For the quantification of bursting discharge, three methods for burst detection that have been previously described were used: 1) The “burst index” (Hutchison et al. 2003), 2) the “L” statistic (Goldberg et al. 2002; Kaneoke and Vitek 1996), and 3) the Poisson “surprise” method (Legendy and Salcman 1985; Wichmann et al. 1999). The burst index is defined as the mean ISI divided by the modal value. To calculate the “L” statistic, the spike train is rebinned into segments of duration equal to the ISI. “L” is defined as the number of distinct values taken on by the rebinned spike train. In the “surprise” method, bursts in the spike train were defined as segments of data with a Poisson surprise value of >5. The minimum number of spikes that can constitute a burst in this method was two. The resulting data were tabulated as the proportion of ISIs within bursts compared with the total number of ISIs in the entire data stream.
Oscillatory activity
Oscillations in the spike train at 0–200 Hz were evaluated using the “global spike shuffling” method (Rivlin-Etzion et al. 2006) to eliminate the artifactual autocorrelations that arise from the neuronal refractory period. Spike shuffling is a process in which the order of ISIs is randomized. Spike time stamps were converted to a data stream consisting of 1-ms bins in which the occurrence or absence of a spike was represented by 1 or 0, respectively, in that bin (Halliday et al. 1995). A 2,048-point fast Fourier transform with Hanning window was used, resulting in a spectral resolution of 0.5 Hz. Similar analysis was performed on “control” data in which ISIs had been randomly shuffled 100 times (Rivlin-Etzion et al. 2006). Twenty shuffles have been reported to yield a good estimate of the “control” spectrum; we increased the number of shuffles fivefold to ensure the accuracy of the estimates. Statistically significant peaks in the spike train data (after normalization with the spectrum of the shuffled data) were determined by using the 300- to 500-Hz part of the spectrum as the control segment and its SD was used as a measure of random fluctuations in the spectrum. Each frequency point between 0 and 200 Hz was then checked for deviation from the expected power, at a significance level of P < 0.01, after correction for multiple (400) comparisons.
The spike-shuffling method for detecting oscillatory activity would not detect first-order oscillations, which can arise when neuronal discharge is very regular with a narrow range of ISIs (Rivlin-Etzion et al. 2006). In such cases, the oscillation frequency is equal to the mean discharge rate. To ensure that no such first-order oscillations were missed, oscillations analysis was also performed without normalizing by the shuffled spectrum and significant oscillations at frequencies at or near the neuron's mean firing frequency were noted.
Quantitative analysis of perimovement discharge
Methods for quantitative analysis of perimovement discharge were as described by Chang et al. (2007). Neuronal data were quantitatively analyzed for passive movement responses only if at least six movement trials in each direction were successfully completed. The onsets of investigator-imposed passive joint movements were detected by manual inspection of individual accelerometer traces. The time of initial deflection in acceleration from baseline was taken as the onset of movement. For each cell tested, there were only two directions about a single joint. For flexion/extension pairs, movement occurred in the same plane but in opposite directions. For ball-and-socket joints (hip and shoulder), care was taken by the examiner to ensure that flexion and extension movements took place in the same plane given the larger degree of freedom at these joints.
Peristimulus mean spike density functions (SDFs; i.e., 1-ms resolution spike delta functions smoothed with a 50-ms sigma Gaussian) (Szucs 1998) were averaged across trials aligned with movement onsets. Statistically significant increases or decreases in neuronal activity were determined by comparison with the mean and variance of the SDF in a prestimulus baseline period. With movement onset defined as time 0, the prestimulus baseline period was from 1.5 to 0.5 s and the analysis period lasted 1,500 ms from −0.5 to 1.0 s. A significant response was defined as a deviation from the mean baseline firing rate that exceeded 3SD of the baseline for 50 ms. The beginning of the response corresponds to the first millisecond of the SDF, the value of which falls outside of the 3SD threshold. This threshold provided an omnibus sensitivity of P < 0.05 after correcting for multiple comparisons (actual P = 0.05/30 comparisons), where the number of independent comparisons was estimated from the duration of the analysis period divided by the degree of smoothing (i.e., 1,500 ms/50 ms). For each neuronal response that was significant, the type of response (i.e., increase, decrease, or combinations of both) was identified by visual inspection of the SDF and the duration of significant response for each neuron was calculated.
Confirmation of electrode location
STN lead placement was verified by postoperative magnetic resonance imaging (MRI) using a transmit–receive headcoil in accordance with the device manufacturer recommendations. The MR images were transferred to an image-processing station (Framelink version 4.1; Medtronic) for analysis. Images were computationally reformatted into standard anatomic planes orthogonal to the intercommissural line and midsagittal plane. Lateral (x), vertical (z), and anterior–posterior (y) coordinates of the distal tip of the DBS lead were measured with respect to the midpoint of the line between the anterior commissure (AC) and the posterior commissure (PC).
Statistical analysis
The distributions of discharge rates, measures of bursting, and lead locations were checked for normality using the Kolmogorov–Smirnov test (“kstest” command from the Matlab statistics toolbox; The MathWorks). Since they were not normally distributed, comparisons between disease states were performed using the Wilcoxon rank-sum test for unpaired data. Paired comparisons of discharge parameters between rest and movement were performed using the Wilcoxon sign-rank test. The chi-squared (χ2) test was used for categorical data (proportions of cells with oscillatory activity).
RESULTS
Patient characteristics
Nine patients with adult or late juvenile-onset dystonia (five males, four females) were included in the study. All dystonia patients had primary dystonia. Three patients had only cervical involvement, whereas the remaining patients had cranial dystonia (i.e., eyes, mouth, or speech/swallow involvement) in addition to prominent cervical dystonia. Three of the six patients with cranial-cervical dystonia also had dystonia involving one or both arms. Two patients also had truncal involvement. There were no patients with generalized dystonia. At the time of surgery, the mean age of the dystonia patients was 46.7 yr (SD 14.1) and the mean duration of symptoms was 11.0 yr (SD 9.8). Further details of clinical symptoms and rating scores are provided in Table 1.
Table 1.
Clinical characteristics of patients with primary dystonia
| Subject | Dystonia Type | Gender | Age at Surgery, yr | Duration of Symptoms, yr | Body Parts Involved (BFMDRS Score >0) | Baseline BFMDRS Movement Score | Baseline TWSTRS Score (Total) | Preoperative Medications (Total mg/day) |
|---|---|---|---|---|---|---|---|---|
| 1 | Seg | F | 45 | 2 | M, S, A (both), N, T | 30 | 60 | Intrathecal baclofen (545 mcg), oxycodone (15) |
| 2 | Seg | F | 47 | 2 | E, M, S, N | 20 | 41 | Diazepam (10), baclofen (40), levetiracetam (1,000) |
| 3 | Foc | F | 51 | 10 | N | 4 | 53 | Oxycodone ER (190), escitalopram (20), clonazepam (2), tizanidine (100), nortriptyline (25), gabapentin (1,800), amlodipine/benazepril (10/20) |
| 4 | Seg | M | 23 | 11 | M, S, N, A (right) | 32 | 56.75 | None |
| 5 | Seg | M | 48 | 7 | N, T | 20 | 53.5 | Clonazepam (1.5), morphine sr (90), acetaminophen/ hydrocodone (1,500/15) |
| 6 | Seg | M | 61 | 7 | E, M, S, N | 22.5 | 61.25 | Morphine sr (90), sertraline (200) |
| 7 | Seg | M | 34 | 17 | M, S, N, A (both) | 40 | 53.5 | Trihexyphenidyl (6), clonazepam (3), topiramate (25) |
| 8 | Foc | F | 40 | 9 | N | 8 | 64 | Clonazepam (1), lexapro (40), tramadol (150), acetaminophen/hydrocodone (3,000/30) |
| 9 | Seg | M | 71 | 34 | E, M, N | 13 | 58.5 | Lorazepam (9) |
| Mean (SD) | 46.7 (14.1) | 11.0 (9.8) | 21.1 (11.7) | 55.7 (6.7) |
A, arm; BFMDRS, Burke–Fahn–Marsden Dystonia Rating Scale; E, eyes; Foc, focal; Seg, segmental; M, mouth; N, neck; S, speech/swallow; T, trunk; TWSTRS, Toronto Western Torticollis Rating Scale.
As a comparison group, 20 patients with PD (17 males; 3 females) were studied. Thirteen of 20 were of the akinetic-rigid subtype and suffered from motor fluctuations or dyskinesias, whereas 7 had tremor as the major component of their disease. All patients were off of antiparkinsonian medications for >12 h prior to surgery and experienced their typical off-medication symptoms during the procedure. No PD patient had off-period dystonia. The mean age at the time of surgery was 60.3 yr (SD 9.0), the mean duration of symptoms was 10.4 yr (SD 3.8), and all patients had a Hoehn and Yahr score of at least Stage 2 (mean 2.6, SD 0.6).
Spontaneous discharge parameters: rate and bursting
In the “resting state” (no attempted voluntary movement), we recorded 62 STN neurons from dystonia patients and 143 STN neurons from PD patients. All recorded cells were considered to be within the motor territory of the nucleus based on the detection of movement-related changes in neuronal discharge in the region of the recording and the fact that recording was restricted to the dorsal 3 mm of the nucleus (Rodriguez-Oroz et al. 2001; Romanelli et al. 2004; Theodosopoulos et al. 2003). (In all, 26% of these dystonia neurons and 39% of these PD neurons showed discharge modulation in response to passive movement and these units form a subset of all neurons screened for responses to passive movement as described further in the following text). Mean recording duration was 33.2 s (SD 13.6, range 10.6–75.0) in dystonia patients and 39.0 s (SD 15.5, range 2.7–118.6) in PD patients. Firing characteristics of a typical dystonia and PD STN neuron are shown in Fig. 1. Group statistics for neuronal firing rates and bursting analysis are provided in Table 2. STN neuronal discharge rate was significantly lower in dystonia patients (26.3 Hz, SD 13.6, P = < 0.001) than that in PD patients (35.6 Hz, SD 15.2), as shown in Fig. 2. Bursting was a prominent feature in both conditions, but STN neurons in patients with dystonia were significantly more bursty than those in PD patients by two of three measures of burstiness (see Table 2). These distinctions between dystonia and PD were also present when neurons from PD patients were subgrouped into akinetic-rigid versus tremor-dominant subtypes, as shown in Table 3. There were no significant differences in spontaneous discharge parameters between the two subtypes of PD. Subgroup analysis of dystonia patients showed no significant differences in spontaneous discharge parameters between focal cervical and segmental dystonia patient groups.
Table 2.
Electrophysiological characteristics of STN neurons in dystonia and Parkinson's disease (PD) at rest
| Factor | Dystonia | Parkinson's Disease (All Units) | P |
|---|---|---|---|
| Number of neurons | 62 | 143 | |
| Mean firing rate, Hz | 26.30 (13.6) | 35.60 (15.2) | <0.001* |
| Burst index | 7.20 (4.0) | 4.70 (2.6) | <0.001* |
| Proportion of discharges in bursts† | 0.26 (0.14) | 0.19 (0.14) | <0.001* |
| L-statistic | 7.10 (1.5) | 6.70 (1.7) | 0.07 |
| Proportion of cells with significant oscillations | |||
| 0–200 Hz | 32.3% | 39.9% | 0.35 |
| 3–30 Hz | 6.5% | 20.3% | 0.01* |
| 13–30 Hz | 0.0% | 6.3% | 0.03* |
Values are means and SD (in parentheses).
Denotes statistically significant difference between dystonia and PD by Wilcoxon rank-sum test or chi-squared test.
As determined using the Poisson “surprise” method.
Fig. 2.

Histograms of spontaneous STN discharge rates in PD (black bars) and dystonia (gray bars). Bin size is 10 Hz. x-axis labels indicate the bin midpoint (i.e., 5 refers to 0–9.99, 15 refers to 10–19.99, 25 to 20–20.99, etc.). Bars are slightly offset for visual clarity. STN neuronal discharge rate was significantly lower in dystonia patients than that in PD patients (P < 0.001).
Table 3.
Electrophysiological characteristics of STN neurons in dystonia and Parkinson's disease (PD) subtypes
| Factor | Dystonia | Parkinson's Disease (Akinetic–Rigid) | P | Parkinson's Disease (Tremor Dominant) | P |
|---|---|---|---|---|---|
| Number of neurons | 62 | 93 | 50 | ||
| Mean firing rate, Hz | 26.30 (13.6) | 35.00 (15.2) | <0.001* | 36.70 (15.1) | <0.001* |
| Burst index | 7.20 (4.0) | 4.70 (2.5) | <0.001* | 4.70 (2.7) | <0.001* |
| Proportion of discharges in bursts† | 0.26 (0.14) | 0.19 (0.13) | 0.001* | 0.20 (0.51) | 0.02* |
| L-statistic | 7.10 (1.5) | 6.70 (1.7) | 0.05 | 6.80 (1.7) | 0.28 |
| Proportion of cells with significant oscillations | |||||
| 0–200 Hz | 32.3% | 39.8% | 0.64 | 40.0% | 0.64 |
| 3–30 Hz | 6.5% | 20.4% | 0.05 | 20.0% | 0.05 |
| 13–30 Hz | 0.0% | 6.5% | 0.09 | 6.0% | 0.09 |
Values are means and SD (in parentheses).
Denotes statistically significant difference between dystonia and PD subtype identified in the preceding column by Wilcoxon rank-sum test or chi-squared test.
As determined using the Poisson “surprise” method.
Oscillatory activity
Results of oscillation analysis are shown in Tables 2 and 3, as well as in Fig. 3. STN oscillatory activity in the 0- to 200-Hz range was a prominent feature of both PD (40% of recorded units showed oscillatory activity) and dystonia (32% with oscillatory activity). In all, 10.5% of PD neurons and 6.5% of dystonia neurons oscillated at more than one frequency. Where multiple oscillation frequencies were detected in a single neuron, they were not in a harmonic relationship. There was a greater incidence of oscillatory activity in low-frequency bands (3–30 Hz) in PD than that in dystonia (20.3 vs. 6.5%, P = 0.01). Oscillatory activity in the beta band (13–30 Hz) was found only in PD (6.3 vs. 0%, P = 0.03). When PD patients were subdivided into tremor-dominant versus akinetic-rigid subtypes based on clinical features, there were no significant differences between oscillatory activity in dystonia versus PD patients of either subtype. Subgroup analysis of dystonia patients did not reveal significant differences in oscillatory activity between focal cervical and segmental dystonia patients. We also looked for first-order oscillations that could occur near the mean firing frequency in neurons with very regular firing patterns, by performing the analysis on spike trains without normalization by the power spectrum of the shuffled spike train (see methods). There were no units in any group oscillating at or very near the mean firing frequency, showing that all oscillatory activity detected reflected second- or higher-order properties of the spike train.
Fig. 3.

A: distribution of frequencies of significant oscillations in spontaneous STN activity in dystonia (top graph) and PD (bottom graph). Bins are left-inclusive, except for the first bin, which starts at 0.4 Hz since the lower limit of oscillatory activity was 0.49 Hz. (i.e., the bins represent 0.4–2.99, 3–7.99, 8–12.99, 13–29.99, 30–59.99, 60–99.99, and 100–199.99 Hz). Several cells oscillated at more than one frequency (see results). There was a greater incidence of oscillatory activity in low-frequency bands (3–30 Hz) in PD than that in dystonia (P = 0.01). Oscillatory activity in the beta band (13–30 Hz) was found only in PD (P = 0.03). B, top: the power spectra of spike trains generated by the global spike shuffling method between 0 and 30 Hz for a patient with the akinetic-rigid subtype of PD. This is the same PD cell displayed in Fig. 1. There is a single significant peak at 11 Hz. Middle: the power spectra between 0 and 30 Hz for a patient with cranial-cervical dystonia. There is a single significant peak at 3.9 Hz. Bottom: the power spectra between 0 and 115 Hz for a patient with akinetic-rigid PD. There are 2 significant peaks at 3.4 and 90.3 Hz.
Neuronal responses to passive movement
Details of somatosensory responses found during the initial screen are shown in Table 4. During the initial intraoperative screen of responses to passive movement based on audiovisual monitoring, 100 STN neurons in 9 dystonia patients and 180 STN neurons in 17 PD patients were tested. A greater proportion of passive joint responses was found in PD than that in dystonia (64 vs. 46%, χ2 = 0.002). Of those units that were responsive to passive movement, 33% were multijoint responsive in PD versus 15% in dystonia (χ2 = 0.02). Of these multijoint responsive cells, 4 of 38 in PD and one of 7 in dystonia were responsive to both upper and lower limb movements. Thirty-four of 46 movement-responsive cells in dystonia patients involved passive movement responses in limbs that did not have clinical dystonia based on their preoperative BFMDRS limb scores. Differences in somatosensory responses between dystonia and PD were maintained when PD patients were divided into akinetic-rigid and tremor-dominant PD subtypes. There was no difference in the proportion of cells responsive to passive movement between akinetic-rigid and tremor-dominant PD subtypes (67 and 57%, respectively; χ2 = 0.17).
Table 4.
Responses to passive movement on initial intraoperative screen of contralateral joint movements
| Factor | Parkinson's Disease | Dystonia | P* |
|---|---|---|---|
| Number of cells tested | 180 | 100 | |
| Movement responsive | 64% | 46% | 0.002 |
| Multijoint responsive† | 33% | 15% | 0.02 |
Chi-squared test.
Proportion of movement-responsive cells with responses to multiple joints.
Responses to passive movement were quantitated using perievent spike density functions in 6 cells in three dystonia patients and 12 cells in four PD patients. Examples of various response patterns are shown in Fig. 4. Neuronal response to only one movement phase was more common in dystonia than that in PD (χ2 = 0.03). However, there were no significant differences in the proportion of movement responses that were positive (increased firing rate), negative (decreased firing rate), or biphasic (increased followed by decreased firing rate) between PD and dystonia (χ2 = 0.16), as detailed in Table 5. The mean duration of significant response was 0.43 s (SD 0.28) for dystonia and 0.37 s (SD 0.23) for PD.
Fig. 4.
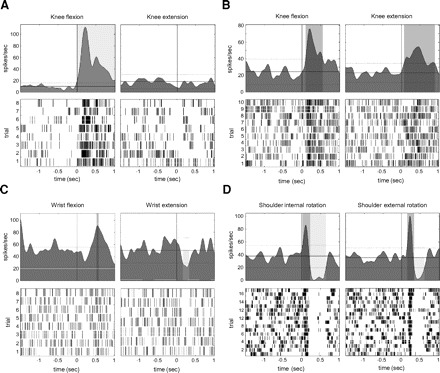
Representative STN neuronal responses to passive limb movement in dystonia and PD. For each example, neuronal responses for reciprocal movements are shown in left and right columns. For each neuron, top panel shows a mean spike density function centered at the initial deflection in limb acceleration (time = 0 s). The dotted line shows the thresholds for significant changes in mean discharge rate. The bottom panel shows a raster display of neuronal responses to each individual trial. A: neuronal response to passive knee movement in a dystonia patient: flexion (left) evoked a brisk increase response, but extension (right) resulted in no appreciable change in firing. B: neuronal response to passive knee movement in a patient with PD: flexion (left) and extension (right) both resulted in firing increases. C: neuronal response to passive wrist movement in a patient with PD: flexion (left) resulted in increased firing and extension (right) evoked a decrease in firing. D: neuronal response to passive shoulder movement in a patient with PD: both external rotation (left) and internal rotation (right) evoked a brisk biphasic response of increased neuronal firing followed by decreased firing.
Table 5.
Quantitative analysis of passive movement–related responses

*Sample includes four cells from two patients with focal cervical dystonia and two cells from a patient with cranial-cervical dystonia. No cells from patients with arm involvement were included. †Chi-squared test. ‡Response proportion calculated using both movement phases. §Wilcoxon rank-sum test.
Movement-related discharge
Nine cells in dystonia and 19 cells in PD were recorded both at rest and during repetitive, voluntary flexion/extension movements of a contralateral limb joint, using the joint that was found to modulate discharge rate during the passive movement screening. Voluntary movement produced a nonsignificant increase in discharge rate in both disease states (Table 6). During movement, in this small sample size, there was a trend toward increased mean STN discharge rate in PD compared with dystonia.
Table 6.
Electrophysiological characteristics of STN neurons with subjects at rest and with voluntary movement
| Dystonia |
PD |
||||||
|---|---|---|---|---|---|---|---|
| Factor | Rest | Movement | P | Rest | Movement | P | PD versus Dystonia (During Active Movement) P |
| Number of neurons | 9 | 9 | 19 | 19 | |||
| Mean firing rate | 23.70 (6.3) | 26.60 (13.3) | 0.5 | 31.70 (13.5) | 35.20 (15.6) | 0.15 | 0.10 |
| Burst index | 7.20 (1.8) | 7.70 (3.9) | 1.0 | 6.80 (2.7) | 5.60 (1.9) | 0.03* | 0.20 |
| Proportion of spikes in bursts | 0.20 (0.08) | 0.29 (0.11) | 1.0 | 0.30 (0.14) | 0.26 (0.13) | 0.26 | 0.70 |
| L-statistic | 7.00 (0.9) | 7.20 (1.4) | 1.0 | 7.70 (1.5) | 7.10 (1.4) | 0.06 | 0.90 |
Values are means and SD in parentheses. Only neurons studied both at rest and during voluntary movement are included in this comparison.
Denotes significant difference between rest and voluntary movement by Wilcoxon paired-sample test.
Confirmation of electrode location
All microelectrode recordings were made within 2 mm of the final DBS electrode trajectory. Thus DBS electrode locations provided an anatomic confirmation of the general region in which cells were recorded and controlled for any systematic difference in the targeted region in dystonia versus PD patients. All DBS leads traversed the dorsolateral STN based on their AC–PC coordinates, as mapped to standard brain atlases (Schaltenbrand and Walker 1982; Talairach et al. 1967). There was no significant difference between lead tip locations in dystonia and PD with respect to the midcommissural point, as measured by postoperative MRI (X [mean]: 11.0 mm [SD 1.3] vs. 11.1 mm [SD 1.1]; Y: 3.5 mm [SD 0.9] vs. 4.1 mm [SD 1.7]; Z: 7.2 mm [SD 1.2] vs. 5.9 mm [SD 2.0]). Methods for confirming electrode location do not allow unambiguous verification of recording location, due to the possibility of misregistration between microelectrode recording and DBS lead placement and the intrinsic MRI error.
DISCUSSION
We compared STN single-unit activity in patients with primary dystonia and Parkinson's disease and found that: 1) mean STN discharge rate in dystonia patients (26.3 Hz, SD 13.6) was lower than that in PD patients (35.6 Hz, SD 15.2); 2) bursting discharge was prevalent in both conditions but even more common in dystonia than that in PD; 3) single-unit oscillatory activity was a prominent feature in both dystonia and PD; 4) beta-range oscillatory activity was more prominent in PD than that in dystonia; 5) dystonia patients had more restricted somatosensory receptive fields in STN neurons, compared with PD; 6) subgroup analysis of both PD (akinetic-rigid vs. tremor-dominant PD) and dystonia (focal cervical vs. segmental dystonia) revealed no significant differences in STN discharge rate, bursting, oscillatory activity, or receptive field size between disease subtypes.
One previous study reported single-unit recordings from the STN in patients with dystonia (Zhuang et al. 2004). The STN mean discharge rate in this mixed population of primary and secondary dystonia patients was found to be 31.5 Hz. Unlike our study, this study did not include a nondystonic comparison group and did not quantitatively analyze oscillations and bursting behavior.
STN discharge rates distinguish different movement disorders
Spontaneous STN discharge rates in the awake state have been measured in several disease states in humans, in the normal state in nonhuman primates (NHPs), and in MPTP-induced parkinsonism in NHPs. Although we do not have normal control data from humans, Steigerwald et al. (2008) recently studied STN neuronal discharge in patients with essential tremor (a disorder thought not to involve the basal ganglia; Lorenz and Deuschl 2007) and found the mean firing rate to be 19.3 Hz. Similarly, most measures of spontaneous discharge rates reported for the STN motor territory in awake normal macaques are close to 20 Hz (Bergman et al. 1994; DeLong et al. 1985; Georgopoulos et al. 1983; Wichmann and Soares 2006). In contrast, STN discharge rates in the off-medication parkinsonian state in humans and MPTP-treated NHPs are usually reported as >30 Hz (Hutchison et al. 1998; Magarinos-Ascone et al. 2000; Magnin et al. 2000; Rodriguez-Oroz et al. 2001; Steigerwald et al. 2008; Theodosopoulos et al. 2003). In this context, our findings suggest that the spontaneous STN discharge rate in dystonia (26.3 Hz) is likely to be higher than normal, although less elevated than in the parkinsonian state. Rodent studies suggest that STN firing rate and pattern are determined in part by a driving influence of the corticosubthalamic pathway and this influence is modulated by dopamine (Bevan et al. 2007; Magill et al. 2001). An additional strong influence on STN firing is the GPe, which in turn is inhibited by D2 receptor-positive striatal cells. Functional imaging studies show reduced D2 receptor activation in a variety of dystonic conditions (Asanuma et al. 2005; Naumann et al. 1998; Perlmutter et al. 1997a), which could suppress GPe discharge and elevate STN discharge. However, a profound elevation in STN discharge rate (>30 Hz) appears to be uniquely associated with PD.
Bursting discharge in the STN
Bursting discharge in the basal ganglia nuclei has been considered to be characteristic of the parkinsonian state, based on an increase in bursting in nonhuman primate and rodent models of PD compared with the normal state (Bergman et al. 1994; Hassani et al. 1996). Pallidal recordings in humans show that primary dystonia is similarly associated with bursting (Starr et al. 2005; Tang et al. 2007; Vitek et al. 1999). This has been confirmed in rodent models of dystonia (Chiken et al. 2008; Gernert et al. 2002). Here, we show that in the STN, bursting discharge is not only present in dystonia, but it is in fact even more prominent than that in PD. This suggests that bursting discharge in basal ganglia output in dystonia is transmitted via the STN. The increased burstiness of STN in dystonia compared with that in PD is consistent with the recent finding that pauses in GPe activity are more prominent in primary dystonia than those in PD (Sani et al. 2009). Since the GPe provides tonic inhibition to the STN, a pause in GPe activity should result in a burst in STN activity. The ultimate effect of basal ganglia bursts on cortical activity is uncertain. Bursts in the activity of basal ganglia inhibitory output neurons may actually facilitate thalamocortical activation and the excessive motor outflow of dystonia by a process of disinhibition (Person and Perkel 2005). Transient reduction in basal ganglia output (such as occurs between bursts) could also paradoxically decrease cortical excitability, as demonstrated in a rodent model of epilepsy (Paz et al. 2007).
Oscillatory activity in the STN
Basal ganglia oscillations have been extensively characterized in the parkinsonian state. Three- to 30-Hz oscillations are present at many points in the basal ganglia–thalamocortical network, in both single-unit and local field potential (LFP) recordings (Hammond et al. 2007). In particular, a pathological role of oscillatory activity in the beta band (13–30 Hz) in the parkinsonian state has been suggested (Brown 2003). In the STN of normal nonhuman primates, very few STN single neurons show oscillatory activity, whereas 27% of neurons show oscillatory activity in MPTP-treated parkinsonian monkeys (Bergman et al. 1994; Wichmann et al. 1994). Induction of parkinsonism in rodents also increases STN single-unit oscillations (Mallet et al. 2008). Although we do not have normal data from humans, patients with a movement disorder not involving the basal ganglia (essential tremor) have fewer 3- to 30-Hz oscillations than those in PD patients (Steigerwald et al. 2008). Thus it seems likely that the oscillatory activity shown here in 32% of STN single units in dystonia patients is a manifestation of the disease state. Since this is a similar proportion as seen in GPi neurons in dystonia patients (Starr et al. 2005), it is also plausible that excitatory inputs from the STN contribute directly to pallidal oscillations. Of note, a mathematical model of dystonia pathophysiology predicts oscillatory activity in structures contributing to the indirect striatopallidal pathway (Sanger 2003).
GPi oscillation frequencies in LFP recordings have been shown to distinguish PD and dystonia, with predominant beta-band oscillations in PD, in contrast to predominant theta–alpha-band oscillations in dystonia (Silberstein et al. 2003). Consistent with this, we found a higher proportion of beta oscillations in spontaneous single-unit STN discharge in PD. Single-unit recording and LFP recording represent different aspects of neuronal activity, since the latter reflects synchronized activity across many neuronal elements and also includes subthreshold synaptic potentials (Chen et al. 2006). Some studies have shown coherence of single neurons with the LFP (Goldberg et al. 2004; Weinberger et al. 2006), whereas others have not (Heimer et al. 2002). Moran et al. (2008) suggested that there may be two distinct neuronal populations with oscillatory activity in PD—tremor frequency (3–7 Hz) and high-frequency (8–20 Hz) neurons—that can be distinguished by their degree of synchronized activity with surrounding neurons.
Responses to passive movement
Widened receptive fields of basal ganglia neurons are a well-documented characteristic of the parkinsonian state (Bergman et al. 1994; Boraud et al. 2000; Rodriguez-Oroz et al. 2001; Vitek et al. 1998) and have been shown in a model of DYT1 dystonia (Chiken et al. 2008). Here, somatosensory receptive fields were widened in PD compared with dystonia. Of note, most passive movement responses in dystonia were tested in limbs that did not clinically have dystonia; thus it is possible that widened receptive fields might be more prominent in patients with generalized dystonia. Most of the multijoint movement-responsive cells in dystonia patients, however, involved passive movement responses in limbs that did not have clinical dystonia. Thus some alteration in STN receptive field properties could represent a dystonic endophenotype (Quartarone et al. 2003, 2008). Quantitative study of neuronal responses to passive movement at a single joint showed that response to only one movement phase was more common in dystonia than that in PD, whereas direction and duration of response did not distinguish PD from dystonia. Responses to passive joint manipulation in our study were similar to those previously described in the GPi in dystonic patients (Chang et al. 2007). Almost all responses to passive movement were increases in discharge, in both PD and dystonia.
Implications for current models of basal ganglia function
Early models of the functional organization of the basal ganglia (Albin et al. 1989; DeLong 1990) emphasize two distinct intrinsic pathways linking the basal ganglia input (striatum) to the basal ganglia outputs (GPi and SNr), which require a balance for normal motor function. Vitek (2002) extended this concept to primary dystonia, proposing that striatal cells originating both the direct and indirect pathways are overactive. This is in contrast to parkinsonism, in which only the striatal cells originating the indirect pathway are overactive. Our findings are consistent with Vitek's model, but indicate that the extent of indirect pathway activation is less than that present in PD. Furthermore, it is now recognized that the corticosubthalamic pathway has a prominent excitatory influence on STN activity (Bevan et al. 2007; Magill et al. 2001); thus our findings would also be consistent with hyperactivity in this pathway in dystonia.
Mink (1996, 2003) proposed a “center–surround” organization of the basal ganglia intrinsic pathways. In the normal state, the inhibitory direct pathway or “center” selects a desired motor program via focal disinhibition of GPi ouput, whereas the divergent excitatory STN–GPi activity provides a broad “surround” inhibition of unwanted motor programs. Nambu and colleagues modified this model to emphasize the “hyperdirect” cortex–STN–GPi pathway as the major source of surround inhibition (Nambu 2004; Nambu et al. 2002). When applied to dystonia, this model predicts that deficient surround inhibition via the STN, or excessive center excitation, results in incomplete suppression of motor pattern generators (Mink 2003). Our results do suggest a relative decrease in surround inhibition in dystonia compared with PD. However, our finding of relatively high spontaneous STN discharge rate in dystonia does not support an absolute decrease in surround inhibition. Further, our limited data on movement-related activity produced no evidence for a movement-related decrease in surround inhibition, since voluntary movement tended to produce an increase in STN firing in both dystonia and PD. Thus the center–surround framework, if applicable to dystonia, would need to emphasize an expanded “center” rather than a deficient “surround” or would need to invoke bursting discharge rather than discharge rate as the mechanism for excessive thalamocortical activations in “surround” channels (Person and Perkel 2005).
Implications for surgical target choice
At most centers performing movement disorders surgery, the STN is the preferred target for PD, whereas GPi is preferred for dystonia. This distinction is largely based on empiric findings rather than physiological principles. It has been hypothesized that the therapeutic effect of DBS may be related to the attenuation of abnormal firing patterns in the basal ganglia–thalamocortical circuit. Here, we show that the major features of pallidal discharge in dystonia that were documented in prior studies are also found in the STN. This provides a physiological basis for the STN as an alternative therapeutic DBS target for the treatment of dystonia. Several cases of STN DBS for the treatment dystonia have been reported in the literature, with beneficial effects in most cases of cervical (Chou et al. 2005; Ostrem et al. 2008), segmental (Kleiner-Fisman et al. 2007; Ostrem et al. 2008), primary generalized (Detante et al. 2004; Kleiner-Fisman et al. 2007; Novak et al. 2008; Sun et al. 2007), and tardive dystonia (Sun et al. 2007; Zhang et al. 2006), whereas outcomes in secondary dystonia patients have not been favorable (Detante et al. 2004; Zhang et al. 2006). Recently, interest in exploring alternative targets for treatment of cervical and segmental dystonia has been spurred by the report that prolonged GPi DBS may cause reversible stimulation-induced bradykinesia in previously nondystonic limbs that cannot be attributed to spread of stimulation to surrounding corticospinal fibers (Berman et al. 2009; Hubl et al. 2009; Ostrem et al. 2007; Zauber et al. 2008).
Pitfalls in interpretation
The present study has some limitations. First, isolation of single units in the human STN is challenging due to high neuronal density and periodic loss of unit isolation in synchrony with the cardiac cycle. As a result, the number of stable, well-isolated units recorded from each study subject was small. Also, the dystonia patients in this study had focal or segmental rather than generalized dystonia. The differences between PD and dystonia found here might have been accentuated in generalized dystonia patients. Nonetheless, even patients with focal dystonia (Garraux et al. 2004; Peller et al. 2006; Quartarone et al. 2008) or nonmanifesting DYT1 gene carriers (Asanuma et al. 2005; Carbon et al. 2008; Eidelberg et al. 1998; Ghilardi et al. 2003; Trost et al. 2002) have global network abnormalities. Additionally, in GPi, similar oscillatory activities are seen in generalized and cervical dystonia patients (Liu et al. 2008).
Conclusions
Spontaneous STN single-unit discharge in humans with primary dystonia shows many of the same features as in PD: prominent bursting and oscillatory discharge. However, oscillatory activity in the beta range is greater in PD than that in dystonia, supporting a current hypothesis that increased beta-band oscillations play a role in the pathophysiology of PD. Although sensory receptive fields are widened in PD compared with those in dystonia, the direction and duration of responses to passive movements are similar in the two conditions. Our findings provide a physiological basis for surgical intervention in STN in primary dystonia.
REFERENCES
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci 12: 366–375, 1989 [DOI] [PubMed] [Google Scholar]
- Asanuma K, Ma Y, Okulski J, Dhawan V, Chaly T, Carbon M, Bressman SB, Eidelberg D. Decreased striatal D2 receptor binding in non-manifesting carriers of the DYT1 dystonia mutation. Neurology 64: 347–349, 2005 [DOI] [PubMed] [Google Scholar]
- Bergman H, Wichmann T, Karmon B, DeLong MR. The primate subthalamic nucleus. II. Neuronal activity in the MPTP model of parkinsonism. J Neurophysiol 72: 507–520, 1994 [DOI] [PubMed] [Google Scholar]
- Berman BD, Starr PA, Marks WJ, Jr, Ostrem JL. Induction of bradykinesia with pallidal deep brain stimulation in patients with cranial-cervical dystonia. Stereotact Funct Neurosurg 87: 37–44, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bevan MD, Hallworth NE, Baufreton J. GABAergic control of the subthalamic nucleus. Prog Brain Res 160: 173–188, 2007 [DOI] [PubMed] [Google Scholar]
- Boraud T, Bezard E, Bioulac B, Gross CE. Ratio of inhibited-to-activated pallidal neurons decreases dramatically during passive limb movement in the MPTP-treated monkey. J Neurophysiol 83: 1760–1763, 2000 [DOI] [PubMed] [Google Scholar]
- Brown P. Oscillatory nature of human basal ganglia activity: relationship to the pathophysiology of Parkinson's disease. Mov Disord 18: 357–363, 2003 [DOI] [PubMed] [Google Scholar]
- Burke RE, Fahn S, Marsden CD, Bressman SB, Moskowitz C, Friedman J. Validity and reliability of a rating scale for the primary torsion dystonias. Neurology 35: 73–77, 1985 [DOI] [PubMed] [Google Scholar]
- Carbon M, Eidelberg D. Abnormal structure-function relationships in hereditary dystonia. Neuroscience 164: 220–229, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carbon M, Ghilardi MF, Argyelan M, Dhawan V, Bressman SB, Eidelberg D. Increased cerebellar activation during sequence learning in DYT1 carriers: an equiperformance study. Brain 131: 146–154, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang EF, Turner RS, Ostrem JL, Davis VR, Starr PA. Neuronal responses to passive movement in the globus pallidus internus in primary dystonia. J Neurophysiol 98: 3696–3707, 2007 [DOI] [PubMed] [Google Scholar]
- Chen CC, Kuhn AA, Trottenberg T, Kupsch A, Schneider GH, Brown P. Neuronal activity in globus pallidus interna can be synchronized to local field potential activity over 3–12 Hz in patients with dystonia. Exp Neurol 202: 480–486, 2006 [DOI] [PubMed] [Google Scholar]
- Chiken S, Shashidharan P, Nambu A. Cortically evoked long-lasting inhibition of pallidal neurons in a transgenic mouse model of dystonia. J Neurosci 28: 13967–13977, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chou KL, Hurtig HI, Jaggi JL, Baltuch GH. Bilateral subthalamic nucleus deep brain stimulation in a patient with cervical dystonia and essential tremor. Mov Disord 20: 377–380, 2005 [DOI] [PubMed] [Google Scholar]
- Comella CL, Stebbins GT, Goetz CG, Chmura TA, Bressman SB, Lang AE. Teaching tape for the motor section of the Toronto Western Spasmodic Torticollis Scale. Mov Disord 12: 570–575, 1997 [DOI] [PubMed] [Google Scholar]
- Dang MT, Yokoi F, McNaught KS, Jengelley TA, Jackson T, Li J, Li Y. Generation and characterization of Dyt1 DeltaGAG knock-in mouse as a model for early-onset dystonia. Exp Neurol 196: 452–463, 2005 [DOI] [PubMed] [Google Scholar]
- DeLong MR. Primate models of movement disorders of basal ganglia origin. Trends Neurosci 13: 281–285, 1990 [DOI] [PubMed] [Google Scholar]
- DeLong MR, Crutcher MD, Georgopoulos AP. Primate globus pallidus and subthalamic nucleus: functional organization. J Neurophysiol 53: 530–543, 1985 [DOI] [PubMed] [Google Scholar]
- Detante O, Vercueil L, Krack P, Chabardes S, Benabid AL, Pollak P. Off-period dystonia in Parkinson's disease but not generalized dystonia is improved by high-frequency stimulation of the subthalamic nucleus. Adv Neurol 94: 309–314, 2004 [PubMed] [Google Scholar]
- Draganski B, Thun-Hohenstein C, Bogdahn U, Winkler J, May A. “Motor circuit” gray matter changes in idiopathic cervical dystonia. Neurology 61: 1228–1231, 2003 [DOI] [PubMed] [Google Scholar]
- Eidelberg D, Moeller JR, Antonini A, Kazumata K, Nakamura T, Dhawan V, Spetsieris P, deLeon D, Bressman SB, Fahn S. Functional brain networks in DYT1 dystonia. Ann Neurol 44: 303–312, 1998 [DOI] [PubMed] [Google Scholar]
- Fahn S, Bressman SB, Marsden CD. Classification of dystonia. Adv Neurol 78: 1–10, 1998 [PubMed] [Google Scholar]
- Garraux G, Bauer A, Hanakawa T, Wu T, Kansaku K, Hallett M. Changes in brain anatomy in focal hand dystonia. Ann Neurol 55: 736–739, 2004 [DOI] [PubMed] [Google Scholar]
- Georgopoulos AP, DeLong MR, Crutcher MD. Relations between parameters of step-tracking movements and single cell discharge in the globus pallidus and subthalamic nucleus of the behaving monkey. J Neurosci 3: 1586–1598, 1983 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gernert M, Bennay M, Fedrowitz M, Rehders JH, Richter A. Altered discharge pattern of basal ganglia output neurons in an animal model of idiopathic dystonia. J Neurosci 22: 7244–7253, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghilardi MF, Carbon M, Silvestri G, Dhawan V, Tagliati M, Bressman S, Ghez C, Eidelberg D. Impaired sequence learning in carriers of the DYT1 dystonia mutation. Ann Neurol 54: 102–109, 2003 [DOI] [PubMed] [Google Scholar]
- Goldberg JA, Boraud T, Maraton S, Haber SN, Vaadia E, Bergman H. Enhanced synchrony among primary motor cortex neurons in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine primate model of Parkinson's disease. J Neurosci 22: 4639–4653, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JA, Rokni U, Boraud T, Vaadia E, Bergman H. Spike synchronization in the cortex/basal-ganglia networks of parkinsonian primates reflects global dynamics of the local field potentials. J Neurosci 24: 6003–6010, 2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halliday DM, Rosenberg JR, Amjad AM, Breeze P, Conway BA, Farmer SF. A framework for the analysis of mixed time series/point process data: theory and application to the study of physiological tremor, single motor unit discharges and electromyograms. Prog Biophys Mol Biol 64: 237–278, 1995 [DOI] [PubMed] [Google Scholar]
- Hammond C, Bergman H, Brown P. Pathological synchronization in Parkinson's disease: networks, models and treatments. Trends Neurosci 30: 357–364, 2007 [DOI] [PubMed] [Google Scholar]
- Hassani OK, Mouroux M, Feger J. Increased subthalamic neuronal activity after nigral dopaminergic lesion independent of disinhibition via the globus pallidus. Neuroscience 72: 105–115, 1996 [DOI] [PubMed] [Google Scholar]
- Heimer G, Bar-Gad I, Goldberg JA, Bergman H. Dopamine replacement therapy reverses abnormal synchronization of pallidal neurons in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine primate model of parkinsonism. J Neurosci 22: 7850–7855, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hierholzer J, Cordes M, Schelosky L, Richter W, Keske U, Venz S, Semmler W, Poewe W, Felix R. Dopamine D2 receptor imaging with iodine-123-iodobenzamide SPECT in idiopathic rotational torticollis. J Nucl Med 35: 1921–1927, 1994 [PubMed] [Google Scholar]
- Hubl J, Kupsch A, Kuhn AA. Frequency-specific effects of deep brain stimulation on hand motor function in patients with cervical dystonia. Mov Disord 24, Suppl. 1: S88, 2009 [Google Scholar]
- Hutchison WD, Allan RJ, Opitz H, Levy R, Dostrovsky JO, Lang AE, Lozano AM. Neurophysiological identification of the subthalamic nucleus in surgery for Parkinson's disease. Ann Neurol 44: 622–628, 1998 [DOI] [PubMed] [Google Scholar]
- Hutchison WD, Lang AE, Dostrovsky JO, Lozano AM. Pallidal neuronal activity: implications for models of dystonia. Ann Neurol 53: 480–488, 2003 [DOI] [PubMed] [Google Scholar]
- Ikoma K, Samii A, Mercuri B, Wassermann EM, Hallett M. Abnormal cortical motor excitability in dystonia. Neurology 46: 1371–1376, 1996 [DOI] [PubMed] [Google Scholar]
- Kaneoke Y, Vitek JL. Burst and oscillation as disparate neuronal properties. J Neurosci Methods 68: 211–223, 1996 [DOI] [PubMed] [Google Scholar]
- Kitai ST, Deniau JM. Cortical inputs to the subthalamus: intracellular analysis. Brain Res 214: 411–415, 1981 [DOI] [PubMed] [Google Scholar]
- Kleiner-Fisman G, Liang GS, Moberg PJ, Ruocco AC, Hurtig HI, Baltuch GH, Jaggi JL, Stern MB. Subthalamic nucleus deep brain stimulation for severe idiopathic dystonia: impact on severity, neuropsychological status, and quality of life. J Neurosurg 107: 29–36, 2007 [DOI] [PubMed] [Google Scholar]
- Legendy CR, Salcman M. Bursts and recurrences of bursts in the spike trains of spontaneously active striate cortex neurons. J Neurophysiol 53: 926–939, 1985 [DOI] [PubMed] [Google Scholar]
- Lenz FA, Suarez JI, Metman LV, Reich SG, Karp BI, Hallett M, Rowland LH, Dougherty PM. Pallidal activity during dystonia: somatosensory reorganisation and changes with severity. J Neurol Neurosurg Psychiatry 65: 767–770, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Levy R, Hutchison WD, Lozano AM, Dostrovsky JO. High-frequency synchronization of neuronal activity in the subthalamic nucleus of parkinsonian patients with limb tremor. J Neurosci 20: 7766–7775, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu X, Wang S, Yianni J, Nandi D, Bain PG, Gregory R, Stein JF, Aziz TZ. The sensory and motor representation of synchronized oscillations in the globus pallidus in patients with primary dystonia. Brain 131: 1562–1573, 2008 [DOI] [PubMed] [Google Scholar]
- Lorenz D, Deuschl G. Update on pathogenesis and treatment of essential tremor. Curr Opin Neurol 20: 447–452, 2007 [DOI] [PubMed] [Google Scholar]
- Lozano AM, Lang AE, Levy R, Hutchison W, Dostrovsky J. Neuronal recordings in Parkinson's disease patients with dyskinesias induced by apomorphine. Ann Neurol 47: S141–S146, 2000 [PubMed] [Google Scholar]
- Magarinos-Ascone CM, Figueiras-Mendez R, Riva-Meana C, Cordoba-Fernandez A. Subthalamic neuron activity related to tremor and movement in Parkinson's disease. Eur J Neurosci 12: 2597–2607, 2000 [DOI] [PubMed] [Google Scholar]
- Magarinos-Ascone CM, Regidor I, Gomez-Galan M, Cabanes-Martinez L, Figueiras-Mendez R. Deep brain stimulation in the globus pallidus to treat dystonia: electrophysiological characteristics and 2 years' follow-up in 10 patients. Neuroscience 152: 558–571, 2008 [DOI] [PubMed] [Google Scholar]
- Magill PJ, Bolam JP, Bevan MD. Dopamine regulates the impact of the cerebral cortex on the subthalamic nucleus-globus pallidus network. Neuroscience 106: 313–330, 2001 [DOI] [PubMed] [Google Scholar]
- Magnin M, Morel A, Jeanmonod D. Single-unit analysis of the pallidum, thalamus and subthalamic nucleus in parkinsonian patients. Neuroscience 96: 549–564, 2000 [DOI] [PubMed] [Google Scholar]
- Mallet N, Pogosyan A, Sharott A, Csicsvari J, Bolam JP, Brown P, Magill PJ. Disrupted dopamine transmission and the emergence of exaggerated beta oscillations in subthalamic nucleus and cerebral cortex. J Neurosci 28: 4795–4806, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marsden CD, Obeso JA, Zarranz JJ, Lang AE. The anatomical basis of symptomatic hemidystonia. Brain 108: 463–483, 1985 [DOI] [PubMed] [Google Scholar]
- Merello M, Cerquetti D, Cammarota A, Tenca E, Artes C, Antico J, Leiguarda R. Neuronal globus pallidus activity in patients with generalised dystonia. Mov Disord 19: 548–554, 2004 [DOI] [PubMed] [Google Scholar]
- Mink JW. The basal ganglia: focused selection and inhibition of competing motor programs. Prog Neurobiol 50: 381–425, 1996 [DOI] [PubMed] [Google Scholar]
- Mink JW. The basal ganglia and involuntary movements: impaired inhibition of competing motor patterns. Arch Neurol 60: 1365–1368, 2003 [DOI] [PubMed] [Google Scholar]
- Monakow KH, Akert K, Kunzle H. Projections of the precentral motor cortex and other cortical areas of the frontal lobe to the subthalamic nucleus in the monkey. Exp Brain Res 33: 395–403, 1978 [DOI] [PubMed] [Google Scholar]
- Moran A, Bergman H, Israel Z, Bar-Gad I. Subthalamic nucleus functional organization revealed by parkinsonian neuronal oscillations and synchrony. Brain 131: 3395–3409, 2008 [DOI] [PubMed] [Google Scholar]
- Nambu A. A new dynamic model of the cortico-basal ganglia loop. Prog Brain Res 143: 461–466, 2004 [DOI] [PubMed] [Google Scholar]
- Nambu A, Takada M, Inase M, Tokuno H. Dual somatotopical representations in the primate subthalamic nucleus: evidence for ordered but reversed body-map transformations from the primary motor cortex and the supplementary motor area. J Neurosci 16: 2671–2683, 1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nambu A, Tokuno H, Inase M, Takada M. Corticosubthalamic input zones from forelimb representations of the dorsal and ventral divisions of the premotor cortex in the macaque monkey: comparison with the input zones from the primary motor cortex and the supplementary motor area. Neurosci Lett 239: 13–16, 1997 [DOI] [PubMed] [Google Scholar]
- Nambu A, Tokuno H, Takada M. Functional significance of the cortico-subthalamo-pallidal “hyperdirect” pathway. Neurosci Res 43: 111–117, 2002 [DOI] [PubMed] [Google Scholar]
- Naumann M, Pirker W, Reiners K, Lange KW, Becker G, Brucke T. Imaging the pre- and postsynaptic side of striatal dopaminergic synapses in idiopathic cervical dystonia: a SPECT study using [123I] epidepride and [123I] beta-CIT. Mov Disord 13: 319–323, 1998 [DOI] [PubMed] [Google Scholar]
- Novak KE, Nenonene EK, Bernstein LP, Vergenz S, Cozzens JW, Rezak M. Successful bilateral subthalamic nucleus stimulation for segmental dystonia after unilateral pallidotomy. Stereotact Funct Neurosurg 86: 80–86, 2008 [DOI] [PubMed] [Google Scholar]
- Ostrem JL, Marks WJ, Jr, Volz MM, Heath SL, Starr PA. Pallidal deep brain stimulation in patients with cranial-cervical dystonia (Meige syndrome). Mov Disord 22: 1885–1891, 2007 [DOI] [PubMed] [Google Scholar]
- Ostrem JL, Volz MM, Starr PA. Deep brain stimulation (DBS) of the subthalamic nucleus (STN) in patients with segmental and focal cervical dystonia. Mov Disord 23: S380, 2008 [Google Scholar]
- Paz JT, Chavez M, Saillet S, Deniau JM, Charpier S. Activity of ventral medial thalamic neurons during absence seizures and modulation of cortical paroxysms by the nigrothalamic pathway. J Neurosci 27: 929–941, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peller M, Zeuner KE, Munchau A, Quartarone A, Weiss M, Knutzen A, Hallett M, Deuschl G, Siebner HR. The basal ganglia are hyperactive during the discrimination of tactile stimuli in writer's cramp. Brain 129: 2697–2708, 2006 [DOI] [PubMed] [Google Scholar]
- Perlmutter JS, Stambuk MK, Markham J, Black KJ, McGee-Minnich L, Jankovic J, Moerlein SM. Decreased [18F]spiperone binding in putamen in idiopathic focal dystonia. J Neurosci 17: 843–850, 1997a [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perlmutter JS, Tempel LW, Black KJ, Parkinson D, Todd RD. MPTP induces dystonia and parkinsonism. Clues to the pathophysiology of dystonia. Neurology 49: 1432–1438, 1997b [DOI] [PubMed] [Google Scholar]
- Person AL, Perkel DJ. Unitary IPSPs drive precise thalamic spiking in a circuit required for learning. Neuron 46: 129–140, 2005 [DOI] [PubMed] [Google Scholar]
- Playford ED, Fletcher NA, Sawle GV, Marsden CD, Brooks DJ. Striatal [18F]dopa uptake in familial idiopathic dystonia. Brain 116: 1191–1199, 1993 [DOI] [PubMed] [Google Scholar]
- Quartarone A, Bagnato S, Rizzo V, Siebner HR, Dattola V, Scalfari A, Morgante F, Battaglia F, Romano M, Girlanda P. Abnormal associative plasticity of the human motor cortex in writer's cramp. Brain 126: 2586–2596, 2003 [DOI] [PubMed] [Google Scholar]
- Quartarone A, Morgante F, Sant'angelo A, Rizzo V, Bagnato S, Terranova C, Siebner HR, Berardelli A, Girlanda P. Abnormal plasticity of sensorimotor circuits extends beyond the affected body part in focal dystonia. J Neurol Neurosurg Psychiatry 79: 985–990, 2008 [DOI] [PubMed] [Google Scholar]
- Raz A, Bergman H, Eimerl D, Israel Z. Propofol induced changes in the neuronal activity of subthalamic nucleus neurons. Mov Disord 23: S117, 2008 [Google Scholar]
- Rivlin-Etzion M, Ritov Y, Heimer G, Bergman H, Bar-Gad I. Local shuffling of spike trains boosts the accuracy of spike train spectral analysis. J Neurophysiol 95: 3245–3256, 2006 [DOI] [PubMed] [Google Scholar]
- Rodriguez-Oroz MC, Rodriguez M, Guridi J, Mewes K, Chockkman V, Vitek J, DeLong MR, Obeso JA. The subthalamic nucleus in Parkinson's disease: somatotopic organization and physiological characteristics. Brain 124: 1777–1790, 2001 [DOI] [PubMed] [Google Scholar]
- Romanelli P, Heit G, Hill BC, Kraus A, Hastie T, Bronte-Stewart HM. Microelectrode recording revealing a somatotopic body map in the subthalamic nucleus in humans with Parkinson disease. J Neurosurg 100: 611–618, 2004 [DOI] [PubMed] [Google Scholar]
- Sanger TD. Childhood onset generalised dystonia can be modelled by increased gain in the indirect basal ganglia pathway. J Neurol Neurosurg Psychiatry 74: 1509–1515, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sani S, Ostrem JL, Shimamoto S, Levesque N, Starr PA. Single unit “pauser” characteristics of the globus pallidus pars externa distinguish primary dystonia from secondary dystonia and Parkinson's disease. Exp Neurol 216: 295–299, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaltenbrand G, Walker AE. Stereotaxy of the Human Brain: Anatomical, Physiological and Clinical Applications. New York: Thieme-Stratton, 1982, p. xiv [Google Scholar]
- Silberstein P, Kuhn AA, Kupsch A, Trottenberg T, Krauss JK, Wohrle JC, Mazzone P, Insola A, Di Lazzaro V, Oliviero A, Aziz T, Brown P. Patterning of globus pallidus local field potentials differs between Parkinson's disease and dystonia. Brain 126: 2597–2608, 2003 [DOI] [PubMed] [Google Scholar]
- Starr PA. Placement of deep brain stimulators into the subthalamic nucleus or globus pallidus internus: technical approach. Stereotact Funct Neurosurg 79: 118–145, 2002 [DOI] [PubMed] [Google Scholar]
- Starr PA, Christine CW, Theodosopoulos PV, Lindsey N, Byrd D, Mosley A, Marks WJ., Jr Implantation of deep brain stimulators into the subthalamic nucleus: technical approach and magnetic resonance imaging-verified lead locations. J Neurosurg 97: 370–387, 2002 [DOI] [PubMed] [Google Scholar]
- Starr PA, Rau GM, Davis V, Marks WJ, Jr, Ostrem JL, Simmons D, Lindsey N, Turner RS. Spontaneous pallidal neuronal activity in human dystonia: comparison with Parkinson's disease and normal macaque. J Neurophysiol 93: 3165–3176, 2005 [DOI] [PubMed] [Google Scholar]
- Steigerwald F, Potter M, Herzog J, Pinsker M, Kopper F, Mehdorn H, Deuschl G, Volkmann J. Neuronal activity of the human subthalamic nucleus in the parkinsonian and nonparkinsonian state. J Neurophysiol 100: 2515–2524, 2008 [DOI] [PubMed] [Google Scholar]
- Sun B, Chen S, Zhan S, Le W, Krahl SE. Subthalamic nucleus stimulation for primary dystonia and tardive dystonia. Acta Neurochir (Wien) 97: 207–214, 2007 [DOI] [PubMed] [Google Scholar]
- Szucs A. Applications of the spike density function in analysis of neuronal firing patterns. J Neurosci Methods 81: 159–167, 1998 [DOI] [PubMed] [Google Scholar]
- Talairach J, Szikla G, Tournoux P. Atlas d'Anatomie Stéréotaxique du Télencéphale: Etudes Anatomo-Radiologiques (Atlas of Stereotaxic Anatomy of the Telencephalon: Anatomo-Radiological Studies). Paris: Masson et Cie, 1967 [Google Scholar]
- Tang JK, Moro E, Mahant N, Hutchison WD, Lang AE, Lozano AM, Dostrovsky JO. Neuronal firing rates and patterns in the globus pallidus internus of patients with cervical dystonia differ from those with Parkinson's disease. J Neurophysiol 98: 720–729, 2007 [DOI] [PubMed] [Google Scholar]
- Theodosopoulos PV, Marks WJ, Jr, Christine C, Starr PA. Locations of movement-related cells in the human subthalamic nucleus in Parkinson's disease. Mov Disord 18: 791–798, 2003 [DOI] [PubMed] [Google Scholar]
- Toro C, Deuschl G, Hallett M. Movement-related electroencephalographic desynchronization in patients with hand cramps: evidence for motor cortical involvement in focal dystonia. Ann Neurol 47: 456–461, 2000 [PubMed] [Google Scholar]
- Trost M, Carbon M, Edwards C, Ma Y, Raymond D, Mentis MJ, Moeller JR, Bressman SB, Eidelberg D. Primary dystonia: is abnormal functional brain architecture linked to genotype? Ann Neurol 52: 853–856, 2002 [DOI] [PubMed] [Google Scholar]
- Vitek JL. Pathophysiology of dystonia: a neuronal model. Mov Disord 17, Suppl. 3: S49–S62, 2002 [DOI] [PubMed] [Google Scholar]
- Vitek JL, Bakay RA, Hashimoto T, Kaneoke Y, Mewes K, Zhang JY, Rye D, Starr P, Baron M, Turner R, DeLong MR. Microelectrode-guided pallidotomy: technical approach and its application in medically intractable Parkinson's disease. J Neurosurg 88: 1027–1043, 1998 [DOI] [PubMed] [Google Scholar]
- Vitek JL, Chockkan V, Zhang JY, Kaneoke Y, Evatt M, DeLong MR, Triche S, Mewes K, Hashimoto T, Bakay RA. Neuronal activity in the basal ganglia in patients with generalized dystonia and hemiballismus. Ann Neurol 46: 22–35, 1999 [DOI] [PubMed] [Google Scholar]
- Weinberger M, Mahant N, Hutchison WD, Lozano AM, Moro E, Hodaie M, Lang AE, Dostrovsky JO. Beta oscillatory activity in the subthalamic nucleus and its relation to dopaminergic response in Parkinson's disease. J Neurophysiol 96: 3248–3256, 2006 [DOI] [PubMed] [Google Scholar]
- Wichmann T, Bergman H, DeLong MR. The primate subthalamic nucleus. III. Changes in motor behavior and neuronal activity in the internal pallidum induced by subthalamic inactivation in the MPTP model of parkinsonism. J Neurophysiol 72: 521–530, 1994 [DOI] [PubMed] [Google Scholar]
- Wichmann T, Bergman H, Starr PA, Subramanian T, Watts RL, DeLong MR. Comparison of MPTP-induced changes in spontaneous neuronal discharge in the internal pallidal segment and in the substantia nigra pars reticulata in primates. Exp Brain Res 125: 397–409, 1999 [DOI] [PubMed] [Google Scholar]
- Wichmann T, Soares J. Neuronal firing before and after burst discharges in the monkey basal ganglia is predictably patterned in the normal state and altered in parkinsonism. J Neurophysiol 95: 2120–2133, 2006 [DOI] [PubMed] [Google Scholar]
- Zauber SE, Watson N, Comella CL, Bakay RA, Metman LV. Stimulation-induced parkinsonism after posteroventral deep brain stimulation of the globus pallidus internus for craniocervical dystonia. J Neurosurg 110: 229–233, 2009 [DOI] [PubMed] [Google Scholar]
- Zhang JG, Zhang K, Wang ZC, Ge M, Ma Y. Deep brain stimulation in the treatment of secondary dystonia. Chin Med J 119: 2069–2074, 2006 [PubMed] [Google Scholar]
- Zhuang P, Li Y, Hallett M. Neuronal activity in the basal ganglia and thalamus in patients with dystonia. Clin Neurophysiol 115: 2542–2557, 2004 [DOI] [PubMed] [Google Scholar]