

The conventional pulmonary circulatory route begins with the pulmonary artery that travels in parallel with the airway, dividing with the airway, until finally reaching the capillary bed within the acinus (4; Fig. 1A). The capillary bed consists of vessels 7 to 10 μm in diameter, never exceeding 13 μm even under very high, non-physiological perfusion pressures (8). The conventional veins then collect blood from capillaries, combining to form progressively larger vessels. Despite this traditional view of the pulmonary vascular circuit, there is substantial anatomic evidence of large-diameter arteriovenous anastomoses in the lung that bypass the traditional blood flow circuit (Fig. 1B).
Fig. 1.
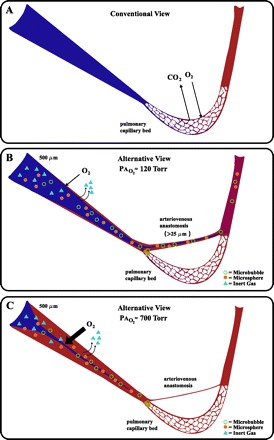
Conventional and alternative views of the pulmonary circulation and gas exchange during exercise. A) Conventional view with the primary gas exchange site restricted to capillary bed. B: alternative view under normoxic conditions (PaO2 ≈ 120 Torr), note precapillary O2 exchange is minimal, but sufficient for the excretion of low solubility multiple inert gas elimination technique (MIGET) gases while precapillary arteriovenous anastomoses allow for deoxygenated blood to bypass capillary beds and allow for some large diameter bubbles and microspheres to escape entrapment by the capillary bed. C: alternative view under hyperoxic conditions (PaO2 ≈ 700 Torr), note precapillary O2 exchange is maximal with elimination of MIGET gases while precapillary arteriovenous anastomoses are closed preventing or reducing arteriovenous blood flow and large diameter bubbles and/or microsphere escape.
A shunt can be defined as “a vascular passage by which blood is diverted from its usual or normal path (arteriovenous)(1).” Arteriovenous anastomoses (i.e., shunts) were first described 129 years ago (19) and these pathways allow for arterial blood to bypass the capillary beds and join up with postcapillary venous blood. Large diameter intrapulmonary arteriovenous pathways (or shunts) are known to exist in many species including humans (25, 27), dogs (16), cats (17), and rabbits (17). A critique of previous anatomic work is that the methods used were not physiological. Recently we documented intrapulmonary arteriovenous pathways using 50 and 25 μm solid microspheres in healthy human, baboon, and dog lungs, which were isolated, ventilated, and perfused at physiological pressures (14, 22). These studies established the patency and functional diameter of some of these intrapulmonary arteriovenous shunt vessels under conditions that more closely replicate physiological conditions.
Using all anatomic based approaches, there is a significant amount of evidence that intrapulmonary arteriovenous shunting during exercise is indeed real. In healthy humans we have demonstrated transpulmonary passage of saline contrast bubbles during submaximal through maximal exercise, but not during upright normoxic rest (6, 12, 13, 23, 24). With the use of saline contrast echocardiography, intrapulmonary shunt is defined as the presence of saline contrast bubbles in the left heart three or more cardiac cycles after appearance of contrast bubbles in the right heart (6, 9, 12, 23). Because saline contrast bubbles small enough to travel through even the largest pulmonary capillaries (<13 μm) have a life span less than three cardiac cycles (even at maximal exercise), transpulmonary passage of these bubbles must occur via large diameter intrapulmonary arteriovenous shunt pathways (2, 15, 18, 28, 29). Of note, saline contrast bubbles can be forced through the normal pulmonary microcirculation using a firmly wedged pulmonary artery catheter with a perfusion pressure of 300 Torr. However, these extreme pulmonary driving pressures do not occur in healthy exercising humans making this an unlikely explanation for the transpulmonary passage of saline contrast bubbles (15).
Consistent with the human, intrapulmonary arteriovenous shunting occurs in dogs. Intravenously injected 25 μm microspheres were found in the tissue and arterial blood of the systemic circulation during exercise but not at rest (22). Dogs were confirmed not to have intracardiac shunts and with an established diameter of 25 μm, these microspheres bypassed the pulmonary capillaries via arteriovenous vessels at least 25 μm in diameter.
Arteriovenous vessels would divert deoxygenated blood away from pulmonary capillaries. If a significant amount of cardiac output was diverted through these pathways when mixed venous partial pressure of oxygen is reduced, such as during exercise, then pulmonary gas exchange as evaluated by the alveolar to arterial oxygen difference (AaDO2) would be impaired. With the use of the Bergman equation, only a 2% shunt of cardiac output would be required to increase AaDO2 during exercise (11). Indeed, a 1.4 ± 0.8% shunt has been calculated in exercising dogs (22) and exercise-induced intrapulmonary arteriovenous shunting is correlated to AaDO2 in healthy humans (23), suggesting these vessels may play an important role in pulmonary gas exchange impairment during exercise.
Based on the amount of morphological and functional anatomic-based data supporting the existence of inducible intrapulmonary shunts, it may be somewhat surprising that work using the 100% O2 technique or the multiple inert gas elimination technique (MIGET) has not detected these pathways in healthy humans during exercise(see Ref. 5 for complete list of references), suggesting that shunts are imaginary. However, this discrepancy may be explained by precapillary gas exchange and the vasomotor effect of O2 on the pulmonary circulation, both of which are critically dependent on concentration gradient and physical properties of the gas (Fig. 1, B and C).
Conhaim and Staub (3) demonstrated precapillary O2 exchange in rapidly frozen cat lungs. In these studies, oxyhemoglobin saturation in 500 μm pulmonary arteries from lungs ventilated with room air were as high as 77% at the perimeter of the blood vessel, while blood at the core of the vessel was as low as 47% saturated with oxygen (3). The size of the perimeter of the blood vessel becoming oxygenated increased from 62 μm in normoxia to 401 μm in lungs ventilated with 100% O2. The authors calculated that in normoxia, mixed venous blood may be as much as 15% oxygenated by the time it reaches the alveolar capillary, while blood would be fully oxygenated before reaching the capillary when breathing 100% O2. Importantly, precapillary gas exchange of both O2 and N2 have also been demonstrated in humans (10, 20). These studies demonstrated that precapillary O2 exchange occurs in normoxia, with a greater O2 exchange occurring in larger vessels with an increased fraction of inspired oxygen. Accordingly, in subjects breathing 100% O2 during exercise, O2 exchange would occur proximal to the intrapulmonary arteriovenous pathways(3, 10, 20), and thus these vessels would not be “seen” as true shunt, as the calculated venous admixture (Qs/Qt) would be minimal (Fig. 1C).
Furthermore, a fundamental assumption of the 100% O2 technique is that the elevated level of inspired oxygen does not have an effect on the pulmonary microcirculation. This does not appear correct, as we recently demonstrated that exercise-induced intrapulmonary arteriovenous shunting can be eliminated in subjects within 2 min of breathing 100% O2 (13). These findings raise a concern for the use of the 100% O2 technique as a valid method for assessing exercise-induced arteriovenous shunt in normoxia and may explain why venous admixture decreases from 3.5% to 0.5% of cardiac output when subjects breathe 100% O2 during exercise (26).
With respect to the MIGET, even a small degree of precapillary gas exchange (i.e., restricted to the perimeter blood of a 500 μm vessel) would allow elimination of low-soluble inert gas within the arteries/arterioles. Therefore, if low-solubility gases are exiting the blood within the pulmonary artery upstream of the capillary beds, then these inert gases would never even reach smaller functional arteriovenous shunt vessels (>25 to 50 μm), and thus these anatomical shunts would appear imaginary to those using the MIGET. In addition, intrapulmonary arteriovenous pathways themselves may participate in limited gas exchange restricted to their perimeter blood, which would allow some deoxygenated core blood to bypass the pulmonary capillary bed in normoxia, but not be recorded as true mixed venous shunt for the same reasons detailed above (7, 21).
More than 100 years of anatomic data document large diameter arteriovenous pathways in the lung. Recent work has simply demonstrated that these vessels are not always open but become functional under specific conditions, such as during exercise. Is exercise-induced intrapulmonary shunting real? When using anatomic-based techniques (microbubbles and microspheres) they are indeed real.
GRANTS
This work was funded by AHA Scientist Development Grant (to A. T. Lovering), AHA Grant-In-Aid 0550176Z (to M. W. Eldridge) and Canadian Institutes of Health Research New Investigator Award (to M. K. Stickland).
REFERENCES
- 1.Webster's Third New International Dictionary of the English Language, Unabridged. Springfield, MA: Merriam-Webster's International, 1993
- 2.Butler BD, Hills BA. The lung as a filter for microbubbles. J Appl Physiol 47: 537–543, 1979 [DOI] [PubMed] [Google Scholar]
- 3.Conhaim RL, Staub NC. Reflection spectrophotometric measurement of O2 uptake in pulmonary arterioles of cats. J Appl Physiol 48: 848–856, 1980 [DOI] [PubMed] [Google Scholar]
- 4.De Mello DE, Reid LM. Arteries and veins. In: The Lung Scientific Foundations, edited by Crystal RG, West JB, Weibel ER, Barnes PJ. Philadelphia: Lippincott-Raven, 1997, p. 1117–1127.
- 5.Dempsey JA, Wagner PD. Exercise-induced arterial hypoxemia. J Appl Physiol 87: 1997–2006, 1999 [DOI] [PubMed] [Google Scholar]
- 6.Eldridge MW, Dempsey JA, Haverkamp HC, Lovering AT, Hokanson JS. Exercise-induced intrapulmonary arteriovenous shunting in healthy humans. J Appl Physiol 97: 797–805, 2004 [DOI] [PubMed] [Google Scholar]
- 7.Genovesi MG, Tierney DF, Taplin GV, Eisenberg H. An intravenous radionuclide method to evaluate hypoxemia caused by abnormal alveolar vessels. Limitation of conventional techniques. Am Rev Respir Dis 114: 59–65, 1976 [DOI] [PubMed] [Google Scholar]
- 8.Glazier JB, Hughes JM, Maloney JE, West JB. Measurements of capillary dimensions and blood volume in rapidly frozen lungs. J Appl Physiol 26: 65–76, 1969 [DOI] [PubMed] [Google Scholar]
- 9.Gudavalli A, Kalaria VG, Chen X, Schwarz KQ. Intrapulmonary arteriovenous shunt: diagnosis by saline contrast bubbles in the pulmonary veins. J Am Soc Echocardiogr 15: 1012–1014, 2002 [DOI] [PubMed] [Google Scholar]
- 10.Jameson AG. Diffusion of gases from alveolus to precapillary arteries. Science 139: 826–828, 1963 [DOI] [PubMed] [Google Scholar]
- 11.Lovering AT, Haverkamp HC, Eldridge MW. Responses and limitations of the respiratory system to exercise. Clin Chest Med 26: 439–457, 2005 [DOI] [PubMed] [Google Scholar]
- 12.Lovering AT, Romer LM, Haverkamp HC, Pegelow DF, Hokanson JS, Eldridge MW. Intrapulmonary shunting and pulmonary gas exchange during normoxic and hypoxic exercise in healthy humans. J Appl Physiol 104: 1418–1425, 2008 [DOI] [PubMed] [Google Scholar]
- 13.Lovering AT, Stickland MK, Amann M, Murphy JC, O'Brien MJ, Hokanson JS, Eldridge MW. Hyperoxia prevents exercise-induced intrapulmonary arteriovenous shunt in healthy humans. J Physiol 586: 4559–4565, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lovering AT, Stickland MK, Kelso AJ, Eldridge MW. Direct demonstration of 25- and 50-μm arteriovenous pathways in healthy human and baboon lungs. Am J Physiol Heart Circ Physiol 292: H1777–H1781, 2007 [DOI] [PubMed] [Google Scholar]
- 15.Meltzer RS, Sartorius OE, Lancee CT, Serruys PW, Verdouw PD, Essed CE, Roelandt J. Transmission of ultrasonic contrast through the lungs. Ultrasound Med Biol 7: 377–384, 1981 [DOI] [PubMed] [Google Scholar]
- 16.Niden AH, Aviado DM Jr. Effects of pulmonary embolism on the pulmonary circulation with special reference to arteriovenous shunts in the lung. Circ Res 4: 67–73, 1956 [DOI] [PubMed] [Google Scholar]
- 17.Prinzmetal M, Ornitz ME, Simkin B, Bergman HC. Arterio-venous anastomoses in liver, spleen, and lungs. Am J Physiol 152: 48–52, 1948 [DOI] [PubMed] [Google Scholar]
- 18.Roelandt J. Contrast echocardiography. Ultrasound Med Biol 8: 471–492, 1982 [DOI] [PubMed] [Google Scholar]
- 19.Sappey PC. Traité Dánatomie Descriptive. Paris: V. A. Delahaye, 1879
- 20.Sobol BJ, Bottex G, Emirgil C, Gissen H. Gaseous diffusion from alveoli to pulmonary vessels of considerable size. Circ Res 13: 71–79, 1963 [DOI] [PubMed] [Google Scholar]
- 21.Stickland MK, Lovering AT. Exercise-induced intrapulmonary arteriovenous shunting and pulmonary gas exchange. Exerc Sport Sci Rev 34: 99–106, 2006 [DOI] [PubMed] [Google Scholar]
- 22.Stickland MK, Lovering AT, Eldridge MW. Exercise-induced arteriovenous intrapulmonary shunting in dogs. Am J Respir Crit Care Med 176: 300–305, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Stickland MK, Welsh RC, Haykowsky MJ, Petersen SR, Anderson WD, Taylor DA, Bouffard M, Jones RL. Intra-pulmonary shunt and pulmonary gas exchange during exercise in humans. J Physiol 561: 321–329, 2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stickland MK, Welsh RC, Haykowsky MJ, Petersen SR, Anderson WD, Taylor DA, Bouffard M, Jones RL. Effect of acute increases in pulmonary vascular pressures on exercise pulmonary gas exchange. J Appl Physiol 100: 1910–1917, 2006 [DOI] [PubMed] [Google Scholar]
- 25.Tobin CE. Arteriovenous shunts in the peripheral pulmonary circulation in the human lung. Thorax 21: 197–204, 1966 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vogiatzis I, Zakynthinos S, Boushel R, Athanasopoulos D, Guenette JA, Wagner H, Roussos C, Wagner PD. The contribution of intrapulmonary shunts to the alveolar-to-arterial oxygen difference during exercise is very small. J Physiol 586: 2381–2391, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.von Hayek H. The Human Lung. [Translated from Die Menschliche Lunge by V. E. Krahl]. New York and London: Hafner, 1960
- 28.Yang WJ, Echigo R, Wotton DR, Hwang JB. Experimental studies of the dissolution of gas bubbles in whole blood and plasma. I. Stationary bubbles. J Biomech 4: 275–281, 1971 [DOI] [PubMed] [Google Scholar]
- 29.Yang WJ, Echigo R, Wotton DR, Hwang JB. Experimental studies of the dissolution of gas bubbles in whole blood and plasma. II. Moving bubbles or liquids. J Biomech 4: 283–288, 1971 [DOI] [PubMed] [Google Scholar]