

Abstract
The mallard (Anas platyrhynchos) is a focal species in game management, epidemiology and ornithology, but comparably little research has focused on the ecology of the migration seasons. We studied habitat use, time-budgets, home-range sizes, habitat selection, and movements based on spatial data collected with GPS devices attached to wild mallards trapped at an autumn stopover site in the Northwest European flyway. Sixteen individuals (13 males, 3 females) were followed for 15–38 days in October to December 2010. Forty-nine percent (SD = 8.4%) of the ducks' total time, and 85% of the day-time (SD = 28.3%), was spent at sheltered reefs and bays on the coast. Two ducks used ponds, rather than coast, as day-roosts instead. Mallards spent most of the night (76% of total time, SD = 15.8%) on wetlands, mainly on alvar steppe, or in various flooded areas (e.g. coastal meadows). Crop fields with maize were also selectively utilized. Movements between roosting and foraging areas mainly took place at dawn and dusk, and the home-ranges observed in our study are among the largest ever documented for mallards (mean = 6,859 ha; SD = 5,872 ha). This study provides insights into relatively unknown aspects of mallard ecology. The fact that autumn-staging migratory mallards have a well-developed diel activity pattern tightly linked to the use of specific habitats has implications for wetland management, hunting and conservation, as well as for the epidemiology of diseases shared between wildlife and domestic animals.
Introduction
The mallard (Anas platyrhynchos) is the most numerous and widespread dabbling duck in the world and an important quarry species in the northern hemisphere. It is often used as a model species for research on ecological processes [1], harvest management [2], and disease [3]–[5]. In Europe, Asia and North America, most mallard populations are migratory and depend on suitable stopover sites along the route to complete their annual migrations [6], [7]. However, stopover ecology is one of the least studied aspects of avian migration [8], and even for a well-studied species like the mallard, little has been published about habitat selection, time-budgets, foraging behaviour, and diet selection during migration. Although food resources of the widely sympatric Eurasian teal (Anas crecca crecca) have been investigated at the flyway level [9], [10], detailed data on an individual basis are very scarce for migrating dabbling ducks. Moreover, in an extensive review of North American dabbling ducks, Callicutt et al. (2011) concluded that there were no studies of food use at the scale of appropriate daily home-range sizes [11].
Active migration requires energy, which birds build up as fat reserves before departing [12], and most migrating birds depend on refuelling along the route [13], [14]. The overall speed of migration is therefore determined by spatial availability of stopover sites and refuelling rates within these sites [12], [13], [15], [16]. Based on analyses of oesophagus and gut contents, mallards generally shift from an invertebrate-dominated diet during spring and summer to a largely granivorous diet during autumn and winter [17]–[22]. Apart from seeds and various water plants, mallards feed on agricultural crops at this time of the year. Accordingly, ducks may aggregate in major grain-growing regions in autumn and winter [16], sometimes causing extensive crop damage [23]. Diet selection depends on food availability, and crop choice by farmers could influence stopover behaviours and spatial dynamics in ducks [24].
Mallards use separate habitats for roosting and foraging, at least in winter [25]–[28]. In general terms, the daily home-range is thought to primarily depend on the spatial distribution of food resources [29], but it may also be affected by predation and hunting pressure, disturbance, and inter- and intraspecific population densities [30]. Hence, home-range sizes vary among regions [31], but also between seasons. For practical reasons, most studies on dabbling ducks have been carried out in daylight [32]. This is an obvious shortcoming with respect to foraging behaviour, as mallards forage mainly at night for a large part of the year [26], [33], [34].
Modern technology provides new opportunities to locate birds without visually following them. By utilizing small, portable GPS devices, birds can be tracked and highly accurate spatial data can be collected remotely. In the present study, we equipped autumn-staging mallards in SE Sweden with local read-out GPS transmitters and followed their movements in a coastal agricultural landscape. The aim was to better understand patterns of habitat use, local movements, home-range sizes, and habitat selection of individual mallards at an important stopover site in the Northwest European flyway [35].
Methods
Ethics statement
This research was carried out following strict protocols during all steps that included contacts with the birds under study. Ethical approval for trapping, sampling, and keeping birds was obtained from the Swedish Animal Research Ethics Board (“Linköpings djurförsöksetiska nämnd”, reference number 46–09). Birds were handled for the minimum amount of time possible. There was no evidence that any individuals died, were hampered, or otherwise negatively affected as a direct result of having a GPS tag attached.
Study area
Öland is a Swedish island in the southern Baltic Sea. It is approximately 140 km from N to S and 18 km E to W at the widest. Öland is a major stopover site for dabbling ducks during autumn, due to a rich food supply and its location between breeding and wintering areas [36]–[38]. Most of the coast consists of shallow, stony beaches, many clad with seaweed beds. Reefs and wave-protected bays are rare. The southern part of Öland is dominated by farmland, pastures and alvar steppe. The alvar is a limestone-rich steppe habitat which, in Sweden, is limited mainly to the Baltic Sea islands of Öland and Gotland. It is an open and flat habitat with little or no soil. The scrubby vegetation is dominated by grass and juniper, and is often grazed by cattle. There are only a few permanent, small wetlands on the southern part of the island, but due to rains the alvar habitat contains several ephemeral wetlands in late autumn.
Trapping
Ottenby Bird Observatory (56°12′N, 16°24′E) uses a stationary trap specifically designed for catching ducks for banding and epidemiological studies (e.g. sampling for avian influenza viruses). The trap is oblong with several funnel-shaped entrances on one side, through which ducks are attracted by bait grain. The trap is cleared daily, at which time ducks that have entered it are herded into a separate section where they are caught, placed in individual cardboard boxes, and taken to a nearby field lab (see [5] for further details). Sampling for this study started on 25 October 2010, during the peak of mallard autumn migration.
GPS devices
After being banded and measured, 40 wild mallards (24 males and 16 females, all first calendar year birds), caught 25–27 October, were equipped with a GPS device and then released at the trap. The devices were of the type “Bird 2AA2” (e-obs Digital Telemetry, Grünwald, Germany) with the maximum measurement 90×30×16 mm, and were equipped with a 70 mm long antenna, angled backwards. Total device weight was 51 grams (approximately 5% of mallard body mass). The device was attached with a harness (see [39] for further details) and a 4 mm thick neoprene pad glued to the bottom of the device. Each device was set to record one data point every 15 minutes, including information about time, location (usually accurate to within 10 meters), and speed over ground. The expected battery life was estimated to be three weeks minimum; some of the devices were still functioning after almost seven weeks.
Fieldwork
The birds were tracked on a daily basis using portable download equipment with receiver. Under ideal conditions, standing at the top of the lighthouse at the southern tip of Öland, 40 m above ground level, data could be downloaded from a distance of 4 km. This was done every day. From the ground, however, downloading data was sometimes difficult even within a few hundred metres. To ensure that we collected data from all tagged ducks remaining in the study area, we almost daily flew over southern Öland with a light airplane on which a receiver was mounted. On three flying occasions, we also searched the northern part of the island and the mainland coast, without finding any tagged individuals. On days with unsuitable weather for airplane flights (>50% of days), 25–30 km of the east coast, as well as a bay on the west coast of the island where birds had previously been located, were covered by car and foot. After 17 November, the airplane was used only once (8 December) due to poor weather. Instead, efforts on the ground were intensified. We were able to download data from all but two devices, yielding a gross sample from 38 birds. Field work was terminated after 47 days. At that time, at least one duck was still staging on southeast Öland.
Analysed data
We excluded birds that showed movement patterns that suggested they were habituated to the duck trap, as these birds' movements were predominately driven by the predictable food source at the trap. Some mallards frequented the Ottenby area and the duck trap during the first week after the tracking device was attached, then abruptly and simultaneously left for another day-roost and started visiting crop fields and/or inland wetlands. Ducks were considered to behave naturally (i.e. not showing trap-dependence) 24 hours after visiting the trap area (<20 metres from the trap), if never returning thereafter. This left us with data from 16 ducks (13 males, 3 females). For these individuals, data on natural behaviour (as described above) were available for on average 24 days (range: 15–38). Some GPS fixes were missing, but in general data could be considered continuous, as average time lag between consecutive locations was only slightly more than the programmed 15 minutes (Table 1). GPS data for the 16 mallards were analysed with respect to habitat use (data points attributed to each habitat), habitat preference, home-range size, and distance of movements. Data points assumed to be from ducks in flight (i.e. if speed was >4 m/s) were excluded. The mean number of data points per duck was 2,180 (min = 1,330; max = 3,366; SD = 646). All the GPS data is stored at Movebank (www.movebank.org), and accessible under the project name “Mallard Waldenstrom Ottenby Sweden” (Movebank ID 3109235).
Table 1. Average time lag (in minutes) between consecutive fixes for 16 mallards studied on southern Öland, Sweden, October – December 2010.
| Individual | Average time lag between consecutive fixes (min) | Standard error |
| 1425 | 15.7 | 13.7 |
| 1426 | 15.4 | 2.5 |
| 1427 | 15.5 | 2.9 |
| 1429 | 15.3 | 2.5 |
| 1440 | 15.3 | 2.4 |
| 1441 | 15.1 | 1.5 |
| 1445 | 15.3 | 2.1 |
| 1451 | 15.3 | 2.3 |
| 1452 | 15.4 | 2.9 |
| 1456 | 15.4 | 2.9 |
| 1457 | 15.4 | 2.8 |
| 1459 | 15.3 | 2.2 |
| 1460 | 15.4 | 2.6 |
| 1461 | 15.3 | 2.5 |
| 1462 | 15.7 | 12.6 |
| 1463 | 15.4 | 2.7 |
| Mean all | 15.4 | 3.83 |
| StDev | 0.14 | 3.66 |
Habitat definitions
A habitat category map was obtained from the Kalmar County Administrative Board. Each data point was assigned a habitat type in order to calculate individual habitat use and habitat selection. The habitat map was rasterized into 10 m square pixels using ArcGis 10 (ESRI 2011) and visualized with the package SMDtools for R software (R Development Core Team 2010). We considered two main habitat categories, dry and wet, further subdivided into more specific groups. Wet habitats were coast/sea, alvar wetland (generally small, 0.01–0.5 ha wetlands situated in the alvar steppe landscape), ordinary wetland (i.e. non-alvar wetlands), flooded area (field or alvar in connection to stream or ditch/canal), wet areas on coastal meadows (often grazed) and pond (man-made). Dry habitats consisted mainly of different types of cropland; pasture, maize, rapeseed, wheat, barley, rye, beans, peas, strawberries and triticale (Figure 1), but also included a few other habitats, such as farm backyard and golf course.
Figure 1. Spatial distribution of mallard GPS fixes (white dots, all individuals pooled) on southern Öland, Sweden, October – December 2010.

Fixes from flying birds (speed >4 m/s) are excluded.
Home-ranges and distances between consecutive locations
We investigated the spatial activity of each individual per day (24-hour-period) using two quantitative measures: the 100% Minimum Convex Polygons (hereafter MCP) and the distances between consecutive locations (hereafter Dcl). Days (24 h) were divided into four periods based on expected differences in behavior: dawn, defined as the period from first light to sunrise (close to two hours duration); day, defined as the period from sunrise to sunset (7–9 hours); dusk, defined as the period from sunset to last light (close to two hours duration); and night, defined as the period from last light to first light (11–13 hours). MCP is classically used for estimating home-range size [40], although kernel estimators [41] and LoCoH (Local Convex Hull; local kernel associated with a local convex polygon) [42] give better estimates of home-range size by excluding unexplored areas within home-ranges. However, we preferred MCP home-range estimates as they better describe an individual's degree of exploration.
We investigated whether the ducks adopted different searching and exploring activities during dawn, day, dusk, and night, using linear mixed models with individual fitted as a random term to account for daily repeated responses of MCP and Dcl for a given individual. Since the number of GPS fixes varied among periods of the day, we tested for a period effect on the MCP while controlling for the number of fixes, using the model MCP = nloc+per, where nloc is a continuous explanatory variable giving the number of fixes for each period, and per is the four-level factor period. The response variables MCP and Dcl were transformed using the Box-Cox transformation[43] to fit normality assumptions. The effect of period was evaluated with the Akaike Information Criterion (AIC) [44] by comparing the models with and without a period effect. All analyses were carried out using the package lme4 for the R software 2.14.
Habitat selection
Following the approach of Manly et al. [45], we investigated patterns of habitat use using selection ratios (w). Habitat selection can be defined as preference for a specific environment [46]–[48]. The preference for a given habitat j, given by its selection ratio wj, is calculated as the number of times a Resource Unit (RU = patch or pixel) within the habitat j is used, divided by the available number of RUs for that given habitat. Under the hypothesis of no particular selection or avoidance, wj should approach one, which means that the habitat j is used in proportion to its availability.
We used Manly's selection ratios for habitat selection design III, which has been developed for studies where each animal is identified and both utilized and available RUs are measured for each individual. We considered all the RUs within an individual's MCP as available RUs, because the study area permits a mallard almost total freedom to roam wherever it chooses. Habitat selection was tested at the individual level using Pearson's Chi-square statistic by comparing the observed selection ratios to the expected selection ratios [45]. Then, for each habitat j and animal i, we computed the wij and associated confidence intervals to identify which habitats that were selected for. All analyses were conducted using the ADEhabitat package [49] for the R software 2.14.
Results
General description of daily routines
The 16 mallards remained in the study area for on average 31 days (range: 15-38) during their recorded stopover, all of them in the southern part of the island and within 40 km from where they were trapped. Most of them frequented the Ottenby area and the eastern side of the island (Figure 1); only one individual spent most of its time on the western side. Most of the 16 mallards showed a common daily routine that can be described as follows: 1) departure from day-roost 45 minutes after sunset; 2) 30-minute visit in a crop field (often maize); 3) arrival at a small permanent or seasonal wetland on the alvar steppe or close to the coast 45 minutes before complete darkness; 4) departure from “night-area” 45 minutes after first light; 5) 30-minute visit in a crop field (often the same field as the one used the previous evening); 6) arrival at day-roost 45 minutes before sunrise (see Figure 2B and 2C). However, there were many exceptions from this schedule, for example frequent flights between wetlands and crop fields during the night.
Figure 2. Example of typical mallard movements between frequently used sites on southeast Öland, Sweden, October – December 2010.

A (no. 1463): Orange ovals = coastal meadows; yellow ovals = maize fields; red ovals = flooded areas; blue ovals = coastal day-roosts (the upper is a sheltered bay, the lower is open water); green ovals = coastal reefs used as day-roosts; grey circle = duck trap location. B (no. 1426): Yellow oval (1) = maize field visited during dawn and dusk; red ovals (2) = various small (flooded) wetlands on alvar steppe (the upper one reaching into a maize field), visited at night; green oval (3) = coastal reef used as day-roost. C (no. 1440): Yellow oval (1) = two maize fields frequently visited, mostly during dawn and dusk; red oval (2) = flooded area (stream) visited most nights; light purple oval (3) = flooded pasture visited during two consecutive nights (probably for feeding); blue oval (4) = most frequented day-roost.
Habitat use
Fourteen of the 16 ducks spent about half of their total time along the coast (mean 47%, SD = 9.6%), which was the typical habitat for resting during the day. Two individuals used inland ponds as day-roosts instead, in which they spent 31% and 61% of their total time, respectively. For all individuals, when days were split in periods, 96% (SD = 8.0%) of the day (sunrise to sunset) was spent at either coast or pond (Table 2). Exclusion of one individual which temporarily used a wetland as its day-roost, increased this percentage to 98% (SD = 2.3). All 16 mallards spent on average 49% (SD = 8.4) at a presumed day-roost locality (Figure 3, Table 3).
Table 2. Habitat use time-budgets (in %) from sunrise to sunset for 16 mallards studied on southern Öland, Sweden, October – December 2010.
| Individual | Coast | Pond | Alvar (flooded) | Wetland (permanent) | Flooded area (outside alvar) | Coastal meadow | Maize | Other habitat | Number of days followed |
| 1425 | 29 | 71 | 0 | 0 | 0 | 0 | 0 | 0 | 16 |
| 1426 | 98 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 22 |
| 1427 | 98 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 20 |
| 1429 | 97 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 29 |
| 1440 | 99 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 16 |
| 1441 | 97 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 15 |
| 1445 | 2 | 98 | 0 | 0 | 0 | 0 | 0 | 0 | 23 |
| 1451 | 96 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 21 |
| 1452 | 97 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 31 |
| 1456 | 97 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 32 |
| 1457 | 92 | 6 | 0 | 0 | 0 | 2 | 0 | 0 | 23 |
| 1459 | 67 | 0 | 0 | 32 | 0 | 1 | 0 | 0 | 18 |
| 1460 | 99 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 34 |
| 1461 | 72 | 28 | 0 | 0 | 0 | 0 | 0 | 0 | 25 |
| 1462 | 90 | 1 | 0 | 0 | 2 | 7 | 0 | 0 | 38 |
| 1463 | 99 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 28 |
| Mean all | 85 | 11 | 0 | 2 | 0.1 | 2 | 0 | 0.1 | 14 |
| StDev | 28 | 29 | 0 | 8 | 0.5 | 2 | 0 | 0.3 | 6.9 |
Figure 3. Percentage of time spent by mallards in different habitats (all individuals, with standard deviation bars) on southern Öland, Sweden, October – December 2010.
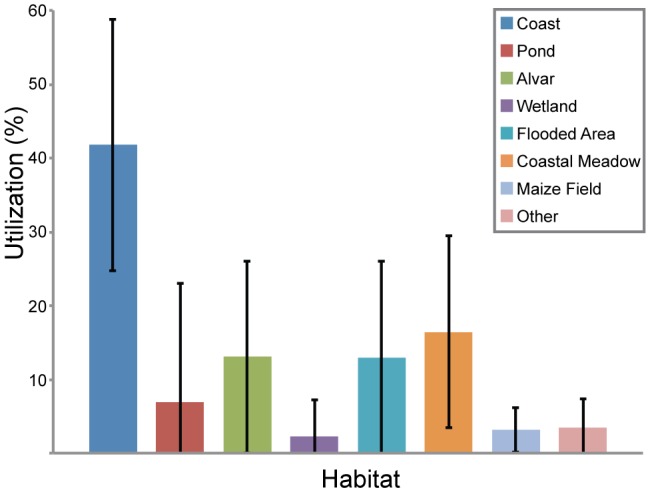
Table 3. Total habitat use time-budgets (in %), home-range size (100% MCP; in ha), and number of GPS fixes for 16 mallards studied on southern Öland, Sweden, October – December 2010.
| Individual (f) = female (m) = male | Coast | Pond | Alvar (flooded) | Wetland (permanent) | Flooded area (outside alvar) | Coastal meadow | Maize | Other habitat | Home-range size | Number of GPS fixes |
| 1425 (m) | 13 | 31 | 5 | 0 | 37 | 6 | 5 | 3 | 2176 | 1374 |
| 1426 (m) | 40 | 2 | 24 | 0 | 11 | 13 | 8 | 2 | 6519 | 2051 |
| 1427 (m) | 45 | 0 | 37 | 0 | 8 | 1 | 4 | 5 | 6822 | 1845 |
| 1429 (f) | 46 | 0 | 1 | 2 | 0 | 49 | 1 | 0 | 6031 | 2622 |
| 1440 (f) | 44 | 0 | 9 | 0 | 40 | 1 | 6 | 1 | 2393 | 1389 |
| 1441 (m) | 66 | 0 | 9 | 0 | 13 | 12 | 0 | 1 | 348 | 1330 |
| 1445 (f) | 1 | 61 | 0 | 0 | 6 | 28 | 0 | 3 | 1145 | 2102 |
| 1451 (m) | 63 | 0 | 2 | 1 | 1 | 19 | 0 | 15 | 1649 | 1850 |
| 1452 (m) | 54 | 0 | 31 | 0 | 4 | 4 | 3 | 4 | 11932 | 2797 |
| 1456 (m) | 42 | 0 | 17 | 0 | 17 | 20 | 3 | 1 | 6937 | 2913 |
| 1457 (m) | 38 | 3 | 10 | 0 | 33 | 10 | 5 | 1 | 11305 | 2051 |
| 1459 (m) | 52 | 0 | 1 | 14 | 0 | 33 | 0 | 0 | 4752 | 1571 |
| 1460 (m) | 51 | 0 | 4 | 0 | 17 | 22 | 4 | 2 | 7182 | 3083 |
| 1461 (m) | 30 | 12 | 32 | 17 | 2 | 0 | 1 | 5 | 4539 | 2284 |
| 1462 (m) | 39 | 2 | 25 | 0 | 6 | 20 | 4 | 4 | 23857 | 3390 |
| 1463 (m) | 46 | 0 | 1 | 0 | 13 | 25 | 6 | 9 | 12153 | 2513 |
| Mean all | 42 | 7 | 13 | 2 | 13 | 16 | 3 | 3 | 6859 | 2198 |
| StDev | 16 | 17 | 13 | 5 | 13 | 13 | 3 | 4 | 5872 | 640 |
The remaining 51% (SD = 8.3%) of their time use was mainly spent in freshwater habitats and, to a lesser extent, in crop fields (Figure 3, Table 3).
Most ducks spent a considerable proportion of the dark hours in small (most less than 50 m in diameter), permanent or seasonal, natural wetlands on the alvar steppe, 3–5 km from the coast. On average, the 16 ducks spent 23% (SD = 23.3%) of their night-time, and 13% (SD = 12.8%) of their total time, in such wetlands (Figure 3, Table 3, Table 4). The variation was large, however. For example, 5 mallards spent on average 54% (SD = 10.6%) of their nights at alvar wetlands. Also seasonally flooded areas (connected to small streams and located up to 3 km from the coast) were used intensively, mainly at night. Three mallards spent 58%, 65% and 72%, respectively, of their night-time in such flooded areas (Table 4). The third major freshwater habitat used by the mallards was flooded coastal meadows, where 6 of the birds spent >45% of their time away from day-roosts (mean = 63%, SD = 18.7%), and 8 ducks spent >25% of their night-time (mean = 46%, SD = 18.3%) (Table 4). Only 2 ducks spent a considerable proportion (29% and 30%, respectively) of their time away from day-roosts in permanent wetlands outside the alvar. Altogether, the four categories of shallow freshwater habitats were used on average 87% (SD = 9.6%) of the total time away from day-roosts (Figure 3, Table 3), and 76% (SD = 15.8%) of the total night-time (Table 4).
Table 4. Habitat use time-budgets (in %) from last light to first light for 16 mallards studied on southern Öland, Sweden, October – December 2010.
| Individual | Coast | Pond | Alvar (flooded) | Wetland (permanent) | Flooded area (outside alvar) | Coastal meadow | Maize | Other habitat | Number of days followed |
| 1425 | 1 | 6 | 8 | 0 | 65 | 10 | 5 | 5 | 16 |
| 1426 | 1 | 3 | 43 | 0 | 19 | 22 | 10 | 2 | 22 |
| 1427 | 10 | 0 | 66 | 0 | 14 | 1 | 5 | 5 | 20 |
| 1429 | 9 | 0 | 2 | 3 | 0 | 83 | 2 | 0 | 29 |
| 1440 | 6 | 0 | 15 | 0 | 72 | 0 | 6 | 0 | 16 |
| 1441 | 42 | 0 | 16 | 0 | 23 | 17 | 0 | 1 | 15 |
| 1445 | 0 | 37 | 0 | 0 | 10 | 51 | 0 | 2 | 23 |
| 1451 | 38 | 0 | 4 | 3 | 1 | 31 | 0 | 24 | 21 |
| 1452 | 22 | 0 | 59 | 0 | 7 | 5 | 3 | 5 | 31 |
| 1456 | 3 | 0 | 31 | 0 | 31 | 33 | 2 | 1 | 32 |
| 1457 | 1 | 1 | 18 | 0 | 58 | 15 | 5 | 0 | 23 |
| 1459 | 40 | 0 | 1 | 0 | 0 | 59 | 0 | 0 | 18 |
| 1460 | 18 | 0 | 7 | 0 | 30 | 37 | 5 | 2 | 34 |
| 1461 | 0 | 0 | 60 | 31 | 3 | 0 | 1 | 5 | 25 |
| 1462 | 6 | 3 | 43 | 0 | 9 | 29 | 5 | 5 | 38 |
| 1463 | 10 | 0 | 1 | 1 | 22 | 43 | 8 | 14 | 28 |
| Mean all | 13 | 3 | 23 | 2 | 23 | 27 | 4 | 5 | 14 |
| StDev | 15 | 9 | 23 | 8 | 23 | 24 | 3 | 6 | 6.9 |
Twelve mallards (75%) visited maize fields, spending on average 4% (SD = 2.0%) of their total time in that habitat (Figure 3, Table 3), and 12% (SD = 5.8%) of the twilight hours (first light to sunrise and sunset to last light). Pastures were visited by all individuals, but only 5 ducks spent more than a total of 3 hours in this habitat; on average 18 hours (SD = 13.9 h) or 3.1% (SD = 3.4%) of their total time-budgets. However, almost half (42%) of the data points ascribed to pastures referred to one mallard visiting a specific pasture that was most probably flooded. The presence of rain water pools probably explains why two mallards spent 6% and 5%, respectively, of their time in bean fields, whereas the rest spent no time at all in this habitat. Some mallards also visited harvested fields with residual rapeseed, wheat or barley, but such fields never made up >5% of an individual's total habitat use.
Home-range and distance analysis
The mallards in our study often flew short (<1 km) or fairly short (3–5 km) distances between foraging areas and day-roosts. However, some ducks regularly travelled up to 26 km between Ottenby and selected foraging sites. Hence, some used a limited area whereas others explored large areas. The 100% MCP home-ranges for the 16 mallards differed considerably in extent, ranging from 348 to 23,857 ha (mean = 6,859 ha; SD = 5,872 ha; Table 3). As expected, individual MCP size was positively related to the number of fixes recorded (linear model after log-transformation of the MCP size, slope = 0.001±3*10e-4, student t-test p<0.01). We therefore controlled for the number of fixes when analysing the effects of the different periods of the day (24 hours) on MCP size.
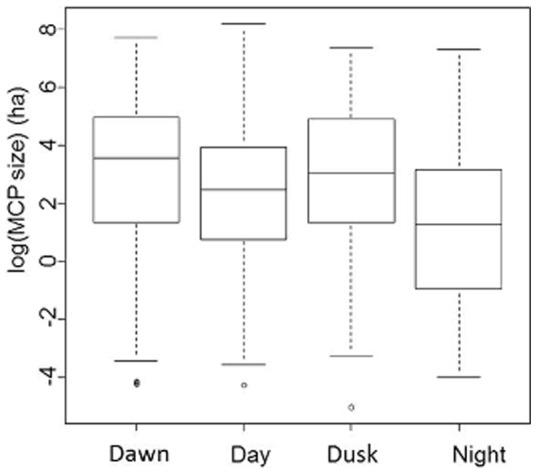
Although the time periods dawn and dusk each make up less than 10% of the total 24-hour-period, the areas explored during these time periods were significantly larger than during the time periods day and night (Figure 4, models MCP = nloc+per vs MCP = nloc, AIC = 7,066 vs 7,081).
Figure 4. Home-range size (MCP in ha on a log scale) of mallards on southern Öland, Sweden, October – December 2010, during different periods of the day; dawn = first light to sunrise (duration close to two hours), day = sunrise to sunset (7–9 hours), dusk = sunset to last light (duration close to two hours), and night = last light to first light (11–13 hours).
Distances between consecutive locations during different periods of the day revealed an even clearer period effect (Figure 5, models Dcl = per vs Dcl = constant, AIC = 107,652 vs 110,960). Overall, ducks moved over greater distances between fixes during dawn and dusk (average model predictions: 37.54 m, 95%CI [35.50, 39.72]; 56.54 m, 95%CI [53.31, 60.00], at dawn and dusk, respectively), whereas they were more stationary during day and night (average model predictions: 12.13 m, 95%CI [11.56, 12.72]; 7.53 m, 95%CI [7.20, 7.88], during day and night, respectively).
Figure 5. Distance between consecutive locations for 16 mallards on southern Öland, Sweden, October – December 2010, during different periods of the day; dawn = first light to sunrise (close to two hours duration), day = sunrise to sunset (7–9 hours), dusk = sunset to last light (close to two hours duration), and night = last light to first light (11–13 hours).

Habitat selection analysis
Habitat selection at the home-range level showed a strong preference for ponds and alvar wetlands in all ducks (Table 5; χ2 = 3,276; df = 96; p<<0.001). Despite the fact that ponds and alvar wetlands are relatively scarce habitat types on Öland, they were highly preferred by mallards in our dataset. This preference outweighed preference for more common habitat types to the point where it became difficult to determine whether these individuals also selected habitats other than ponds and alvar wetlands. In order to further explore selection, all wet habitats were pooled together. This resulted in a clear selection for maize fields (Table 6, test of overall departure from non-preference: χ2 = 18,843; df = 73; p<<0.001).
Table 5. Selection ratios (wi) at the home-range level, averaged over 16 mallards studied on southern Öland, Sweden, October – December 2010, with respective 95% confidence interval (CI) lower and upper limits.
| Habitat | wi | CI lower | CI upper |
| Ponds | 143.82 | −7.96 | 295.61 |
| Alvar wetlands | 29.99 | 12.57 | 47.41 |
| Other wetlands | 6.11 | −2.91 | 15.13 |
| Maize fields | 2.95 | 1.54 | 4.36 |
| Coast and sea | 1.49 | 0.80 | 2.19 |
| Coastal meadows | 1.20 | 0.44 | 1.95 |
| Alvar steppe | 0.77 | 0.04 | 1.50 |
| Pasture fields | 0.34 | 0.15 | 0.54 |
| Other crops | 0.17 | 0.02 | 0.32 |
| Other habitat | 0.08 | 0.03 | 0.13 |
The table shows the selection ratios when considering the habitat as a ten-level factor (wet habitats are divided in subcategories). The habitats are ordered from the most (top) to the least preferred habitat (bottom).
Table 6. Selection ratios (wi) at the home-range level, averaged over 16 mallards studied on southern Öland, Sweden, October – December 2010, with respective 95% confidence interval (CI) lower and upper limits.
| Habitat | wi | CI lower | CI upper |
| Maize fields | 2.95 | 1.63 | 4.28 |
| Coast/wetlands | 1.90 | 1.39 | 2.42 |
| Alvar steppe | 0.77 | 0.08 | 1.45 |
| Pasture fields | 0.34 | 0.16 | 0.53 |
| Other crops | 0.17 | 0.03 | 0.31 |
| Other habitat | 0.08 | 0.04 | 0.13 |
The table shows the selection ratios when considering the habitat as a six-level factor (all wet habitats are pooled together). The habitats are ordered from the most (top) to the least preferred habitat (bottom).
Investigating the pattern of habitat selection for each of the four periods of the day, we found a clear shift in habitat preference from the periods dawn and dusk compared to day and night (Figure 6). There was a clear preference for maize fields during dawn and dusk (tests of overall departure from non-preference, χ2 = 1,821; df = 55; p<<0.001 and χ2 = 1,922; df = 64; p<<0.001, respectively), while wetlands were preferentially selected during night and day (test of overall departure from non-preference: χ2 = 13,096; df = 65; p<<0.001 and χ2 = 2,695; df = 19; p<<0.001, respectively).
Figure 6. Manly et al.'s selection ratios (wij±95% CI) for each period of the day.

Habitat is considered as a six level factor, i.e. all wetland habitats have been pooled.
Discussion
Habitat use
Our study demonstrates that autumn-staging mallards have a clear diel pattern comprising major shifts in activity and habitat preference. The tracked mallards spent their daylight hours mostly at coastal localities, strikingly often at reefs in protected areas (often in nature reserves), and visited fields and inland wetlands during the night. Flight activity was highest at dawn and dusk (Figure 4, Figure 5), when ducks moved between day-roosts and foraging areas, which is in accordance with previous studies [32], [34]. Although some individuals were exceptions from this behaviour, and ducks often made additional flights, by and large mallard diel movements were similar among individuals and predictable in time. Some of the ducks in this study displayed fixed behavioural patterns, whereby they moved between the same habitat locations, at similar times, for many consecutive days. Moreover, we found that these predictable movements were often performed by pairs or groups of individuals evidently moving together.
Nocturnal foraging is generally attributed to more profitable foraging conditions (e.g. availability of nocturnally active prey) or lower predation risk [25]. Diurnal predators, such as raptors, are usually abundant in the study area in autumn, and Öland holds a vital population of red fox (Vulpes vulpes). Duck hunters are also present on Öland, but few in number. Since the hunting pressure is low (according to hunting reports and personal observations), it is unlikely to affect the foraging patterns of our study ducks to any large extent.
Day-roost and night-foraging patterns in winter have been described for the mallard, as well as for other dabbling ducks [28], [30], [34], [50], but the amount of night-foraging may differ within and between species [51]. In western France, wintering ducks spent 37–60% of their time foraging; on average 16% of daylight hours and 85% of the night [33]. The Eurasian teal has been shown to forage 88% of the total time on a daily basis in February – August [10], but only 46% in winter [26].
We found that the mallards in our study spent approximately equal time in “foraging” versus “resting” habitats (Figure 3, Table 2–4). This is an important insight, as we are not aware of any other study providing total time-budgets for habitat use of dabbling ducks during stopover. Admittedly, we cannot be certain that the mallards actually foraged in the shallow freshwater habitats that were visited mostly at night, as they may have filled their crops when visiting crop fields at dusk. However, it seems unlikely that they did not forage in the freshwater habitats, as it would have been equally possible, and probably much safer, to return to the coast to digest the crop contents. The obvious pattern of distinctly different habitat use in day and night also indicates that the mallards used freshwater habitats for foraging (and drinking).
There was considerable variation in how much time individuals spent at day-roosts versus foraging areas (Table 3). This could be explained by the possibility that some day-roosts offered foraging opportunities, thereby reducing the need for foraging at night, but may also reflect different needs related to individual body condition. Although some foraging probably occurred at the day-roost localities, it was likely limited as the reefs and bays do not offer good foraging habitats (brackish and fairly open water with little vegetation to feed on for a dabbling duck). Our data suggest that foraging times were approximately the same as for wintering ducks in France [26], [33].
Home-range and distance of movements
The home-ranges observed in our study are some of the largest ever documented for mallards, albeit with high standard deviations (Table 3, Table 7). This was mainly due to the relatively long travel distances performed by some individuals between day-roosts and night-time foraging areas. Observed distances between consecutive locations verify that longer movements were undertaken during dawn and dusk, while mainly shorter distances were covered during day and night (Figure 5).
Table 7. Previously published home-range sizes for mallards in comparison to the present study.
| Study/Author | Country (state) | Season | Home-range size (in ha) | Standard deviation | Number of ducks |
| This study | Sweden | autumn | 6859 | 5872 | 16 |
| Jorde et al. 1984 | USA (Nebraska) | winter | 3243.7** | 638.7 | 14 |
| Legagneux et al. 2009 | France | winter | 1257.3 | 1915.1 | 15 |
| Legagneux et al. 2009 | France | winter | 496.2 | 861.3 | 8 |
| Legagneux et al. 2009 | France | winter | 326.6 | 162.7 | 8 |
| Legagneux et al. 2009 | France | winter | 221.0 | 242.8 | 7 |
| Legagneux et al. 2009 | France | winter | 184.7 | 147.7 | 7 |
| Sauter et al. 2012 | Switzerland | winter | min 16* max 6367* | not available | 33 |
| Dwyer et al. 1979 | USA (North Dakota) | breeding (females) | 467.8 | 158.6 | 6 |
| Kirby et al. 1985 | USA (Minnesota) | Breeding (females) | 343.8 | 391.4 | 5 |
| Gilmer et al. 1975 | USA (Minnesota) | Breeding (males) | 240 | not available | 12 |
| Gilmer et al. 1975 | USA (Minnesota) | Breeding (females) | 210 | not available | 12 |
(Further examples exist, especially for breeding birds, but our aim here is to show a representative variation.).
*Mean values not published.
**This mean value does not include foraging areas.
The reefs where most ducks spent the days were fairly safe from predators, while the fields/wetlands where the ducks spent the nights were obviously attractive in terms of foraging opportunities. This suggests that the benefits obtained from these habitats outweighed the costs (such as energy expenditure and predation risk) incurred by moving between them. In areas where similar studies have been performed in winter, extensive wetlands make the ducks less prone to change foraging sites, and travelling distances are usually shorter. Wintering dabbling ducks usually move less than 25 km on a daily basis, based on studies of mallard [34], [52], [53] and pintail [54]. Home-ranges for wintering mallards seldom exceed 5,000 ha [34], [55], [56] and for breeding birds they are usually much smaller [57]–[60] (Table 7). It could be that birds learn to minimize home-range size more effectively in winter, compared to the shorter stopover periods in spring and autumn, respectively. Also, the need to minimize energy loss is usually higher in winter, as thermoregulation can be costly in cold temperatures.
The 3 females in our study had a much smaller average home-range size (3,190 ha; SD = 2,539 ha) than the 13 males (7,705 ha; SD = 6,105 ha). Our low sample size of females does not permit any conclusions to be drawn, although sex (and age) could possibly affect home-range size. Jorde et al. (1984) argued that young birds may need to explore larger areas due to inexperience [55]. The latter study also reported larger home-ranges for males than females, but with a rather small sample size (winter study; 14 mallards divided in 4 age/sex groups).
Habitat selection
The importance of alvar wetlands as a stopover habitat for mallards was previously unknown, although inland wetlands are generally considered very important to dabbling ducks [61], [62]. This finding has important implications beyond mallard autecology since dabbling ducks are considered to be main vectors of propagules (e.g. eggs, seeds, oogones) of aquatic organisms between isolated wetlands [63]. This can be considered an important ecosystem service [64], but possibly also a concern when it comes to spread of invasive species [65].
A large proportion of coastal southern Sweden is heavily farmed, but ducks have seldom been observed foraging in fields. However, our data show that they frequently do so. In the study area, maize fields are harvested at the end of the summer, leaving abundant residual maize for mallards to forage. Maize seems to be the most attractive crop to our staging mallards (Figure 6, Table 3, Table 4), a pattern also observed in other areas [53]. This is despite the fact that the birds in our study (all juveniles) most likely have never encountered maize before (it is not grown closer to the main breeding grounds of mallards farther northeast in Sweden, Finland, and Russia). This might imply a benefit for the mallard, being a generalist more than a specialist feeder. Furthermore, we found evidence in our data that some of the mallards moved in groups between fixed habitat locations. It is quite possible that the juveniles we tracked travelled alongside adult individuals who had previously stopped-over on Öland during previous migrations, and learnt to utilize grain fields from individuals with prior knowledge of the landscape.
The proportion of maize fields compared to other crops has increased in the study area in recent years. Until 2005, <1% of the farmland on southern Öland (the municipality of Mörbylånga) was used for maize, whereas in the last few years it has been grown to encompass >6% of the agricultural land. At the national level, maize fields cover only 0.6% of the total agricultural area used for crops in Sweden, but have increased by >400% from 2001 (3,161 ha) to 2012 (16,482 ha). This change of farming preferences could have an impact on the stopover ecology of mallards travelling through Sweden, as has been observed for other bird species. Several species of geese have recently become much more numerous in response to increasing availability of autumn-sown crops and sugar beet spill (due to machine harvesting) in Sweden and beyond [66]. The pink-footed goose (Anser brachyrhynchus) population on Svalbard has increased dramatically, mainly due to improved feeding conditions at stopover and wintering sites [67].
The frequent but usually very brief visits (most often 30 minutes or less) in maize fields observed in this study, could possibly be because most fields on Öland are located between the coast and the alvar steppe, i.e. between major foraging/roosting areas. But the geographically convenient location is probably not the main reason for the popularity of maize fields. For example, some ducks made several visits during a single night, by repeated flights between a wetland and a maize field.
We suggest that predation pressure, being a strong determinant of foraging behaviour [10], [68]–[70], is the most likely reason for the brief visits in maize fields. The risk of predation could also explain why the ducks preferred to visit maize fields in twilight, when diurnal raptors are usually not active and it is still possible to discern nocturnal predators (e.g. fox) by sight. According to optimal foraging theory, even a highly beneficial habitat or behaviour should only be used when the risk of predation is relatively low [71]–[73]. It could be that the ducks quickly fill up their crop and then leave for safer grounds.
Acknowledgments
We specifically thank the staff at Ottenby Bird Observatory for the work at the duck trap and all the hours spent tracking the mallards. Agricultural data were obtained from the Kalmar County Administrative Board (Länsstyrelsen i Kalmar län) and the Swedish Board of Agriculture (Jordbruksverket). Kalmar flygklubb was immensely valuable in helping out with the airborne tracking. Michelle Wille improved the figures and Jo Chapman improved the language. Two reviewers added highly relevant comments on a previous version of the manuscript. This is publication no. 281 from Ottenby Bird Observatory.
Funding Statement
Funding for this project was provided by the Swedish Research Councils VR (2010–3067, 2011–3568) and FORMAS (2009–1220). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Gunnarsson G, Elmberg J, Sjöberg K, Pöysä H, Nummi P (2006) Experimental evidence for density-dependent survival in mallard (Anas platyrhynchos) ducklings. Oecologia 149: 203–213. [DOI] [PubMed] [Google Scholar]
- 2. Nichols JD, Runge MC, Johnson FA, Williams BK (2007) Adaptive harvest management of North American waterfowl populations: a brief history and future prospects. Journal of Ornithology 148: 343–349. [Google Scholar]
- 3. Jourdain E, Gunnarsson G, Wahlgren J, Latorre-Margalef N, Bröjer C, et al. (2010) Influenza Virus in a Natural Host, the Mallard: Experimental Infection Data. Plos One 5: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Latorre-Margalef N, Gunnarsson G, Munster VJ, Fouchier RA, Osterhaus AD, et al. (2009) Effects of influenza A virus infection on migrating mallard ducks. Proceedings of the Royal Society B - Biological Sciences 276: 1029–1036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Wallensten A, Munster VJ, Latorre-Margalef N, Brytting M, Elmberg J, et al. (2007) Surveillance of influenza A virus in migratory waterfowl in northern Europe. Emerging Infectious Diseases 13: 404–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Arzel C, Elmberg J, Guillemain M (2006) Ecology of spring-migrating Anatidae: a review. Journal of Ornithology 147: 167–184. [Google Scholar]
- 7.Gunnarsson G, Waldenström J, Fransson T (2012) Direct and indirect effects of winter harshness on the survival of Mallards Anas platyrhynchos in northwest Europe. Ibis: 1–11.
- 8. Lindström A (1995) Stopover Ecology of Migrating Birds - Some Unsolved Questions. Israel Journal of Zoology 41: 407–416. [Google Scholar]
- 9. Arzel C, Elmberg J, Guillemain M, Lepley M, Bosca F, et al. (2009) A flyway perspective on food resource abundance in a long-distance migrant, the Eurasian teal (Anas crecca). Journal of Ornithology 150: 61–73. [Google Scholar]
- 10. Arzel C, Elmberg J, Guillemain M (2007) A flyway perspective of foraging activity in Eurasian Green-winged Teal, Anas crecca crecca . Canadian Journal of Zoology-Revue Canadienne De Zoologie 85: 81–91. [Google Scholar]
- 11. Callicutt JT, Hagy HM, Schummer ML (2011) The Food Preference Paradigm: A Review of Autumn–Winter Food Use by North American Dabbling Ducks (1900–2009). Journal of Fish and Wildlife Management 2: 29–40. [Google Scholar]
- 12.King JR (1974) Seasonal allocation of time and energy resources in birds. Pages 4–70 in Paynter RA (Ed) Avian Energetics. Nuttall Ornithological Club, no. 15, Cambridge, Massachusetts, USA.
- 13.Alerstam T, Lindström A (1990) Optimal bird migration: the relative importance of time, energy and safety. Pages 331–351 in Gwinner, E (Ed) Bird Migration: The Physiology and ecophysiology. Springer-Verlag, Berlin, Germany.
- 14. Weber TP, Houston AI (1997) Flight costs, flight range and the stopover ecology of migrating birds. Journal of Animal Ecology 66: 297–306. [Google Scholar]
- 15. Alerstam T, Hedenström A (1998) The development of bird migration theory. Journal of Avian Biology 29: 343–369. [Google Scholar]
- 16.Van Eerden MR (1984) Waterfowl movements in relation to food stocks. Pages 84–100 in Evans PR, Goss-Custard JD, Hale WG (Eds) Coastal waders and wildfowl in winter. Cambridge University Press, Cambridge, UK.
- 17.Martin AC, Uhler FM (1939) Food of game ducks in the United States and Canada. US Department of Agriculture, Technical Bulletin no 634.
- 18. Forsyth B (1965) December food habits of the mallard (Anas platyrhynchos Linn.) in the grand prairie of Arkansas. Arkansas Academy of Science Proceedings 19: 74–78. [Google Scholar]
- 19.Cramp S, Simmons KEL (1977) Birds of the Western Palearctic, vol 1 . Oxford University Press, Oxford, UK. [Google Scholar]
- 20.Glutz von Blotzheim UN (1990) Handbuch der Vögel Mitteleuropas. Band 2 Anseriformes, 2nd edition. Aula, Wiesbaden, Germany.
- 21.Krapu GL, Reinecke KJ (1992) Foraging ecology and nutrition. Pages 1–29 in Batt DJ, Afton AD, Anderson MG, Ankney CD, Johnson DH, Kadlec JA, Krapu GL, (Eds) Ecology and Management of Breeding Waterfowl. University Press of Minnesota, USA.
- 22. Tidwell PR, Webb EB, Vrtiska MP, Bishop AA (2013) Diets and Food Selection of Female Mallards and Blue-Winged Teal During Spring Migration. Journal of Fish and Wildlife Management 4: 63–74. [Google Scholar]
- 23. Foster MA, Gray MJ, Kaminski RM (2010) Agricultural Seed Biomass for Migrating and Wintering Waterfowl in the Southeastern United States. Journal of Wildlife Management 74: 489–495. [Google Scholar]
- 24. Brochet AL, Mouronval JB, Aubry P, Gauthier-Clerc M, Green AJ, et al. (2012) Diet and Feeding Habitats of Camargue Dabbling Ducks: What Has Changed Since the 1960s? Waterbirds 35: 555–576. [Google Scholar]
- 25. McNeil R, Drapeau P, Gosscustard JD (1992) The Occurrence and Adaptive Significance of Nocturnal Habits in Waterfowl. Biological Reviews of the Cambridge Philosophical Society 67: 381–419. [Google Scholar]
- 26.Tamisier A, Dehorter O (1999) Camargue. Canards et foulques. Centre Ornithologique du Gard, Nime, France.
- 27. Guillemain M, Fritz H, Duncan P (2002) The importance of protected areas as nocturnal feeding grounds for dabbling ducks wintering in western France. Biological Conservation 103: 183–198. [Google Scholar]
- 28. Kozulin A (1995) Ecology of Mallards Anas platyrhynchos wintering in low temperatures conditions in Belarus. Acta Ornithologica 30: 125–134. [Google Scholar]
- 29. Schoener TW (1983) Simple-Models of Optimal Feeding-Territory Size - a Reconciliation. American Naturalist 121: 608–629. [Google Scholar]
- 30. Legagneux P, Blaize C, Latraube F, Gautier J, Bretagnolle V (2009) Variation in home-range size and movements of wintering dabbling ducks. Journal of Ornithology 150: 183–193. [Google Scholar]
- 31. Roshier DA, Doerr VAJ, Doerr ED (2008) Animal movement in dynamic landscapes: interaction between behavioural strategies and resource distributions. Oecologia 156: 465–477. [DOI] [PubMed] [Google Scholar]
- 32.Paulus SL (1988) Time-activity budgets of nonbreeding Anatidae: a review. Pages 135–152 in Wennerberg L (Ed) Waterfowl in winter. University Press of Minnesota, USA.
- 33. Guillemain M, Fritz H, Duncan P (2002) Foraging strategies of granivorous dabbling ducks wintering in protected areas of the French Atlantic coast. Biodiversity and Conservation 11: 1721–1732. [Google Scholar]
- 34. Sauter A, Korner P, Fiedler W, Jenni L (2012) Individual behavioural variability of an ecological generalist: activity patterns and local movements of Mallards Anas platyrhynchos in winter. Journal of Ornithology 153: 713–726. [Google Scholar]
- 35. Guillemain M, Sadoul N, Simon GR (2005) European flyway permeability and abmigration in Teal Anas crecca, an analysis based on ringing recoveries. Ibis 147: 688–696. [Google Scholar]
- 36. Helseth A, Lindström A, Stervander M (2005) Southward migration and fuel deposition of Red Knots Calidris canutus . Ardea 93: 213–224. [Google Scholar]
- 37. Bergström U, Svärdson G (1938) Ringmärkningar och fågeliakttagelser vid Ölands södra udde under höststräcket 1937. [Ringing and observations at the southern point of island Öland during autumn migration 1937; in Swedish.] Fauna & Flora 33: 1–16. [Google Scholar]
- 38.Edelstam C (1972) The Visible Migration of Birds at Ottenby, Sweden. Vår fågelvärld, supplement no 7.
- 39. Roshier DA, Asmus MW (2009) Use of satellite telemetry on small-bodied waterfowl in Australia. Marine and Freshwater Research 60: 299–305. [Google Scholar]
- 40. Mohr CO (1947) Table of equivalent populations of North American small mammals. American Midland Naturalist 37: 223–249. [Google Scholar]
- 41. Worton BJ (1989) Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology 70: 164–168. [Google Scholar]
- 42. Getz WM, Fortmann-Roe S, Cross PC, Lyons AJ, Ryan SJ, et al. (2007) LoCoH: Nonparameteric Kernel Methods for Constructing Home Ranges and Utilization Distributions. PLOS ONE 2: e207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Sakia RM (1992) The Box-Cox transformation technique: a review. The statistician 41: 169–178. [Google Scholar]
- 44.Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edition. Springer, New York, USA.
- 45.Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP (2003) Resource selection by animals. Statistical design and analysis for field studies, 2nd edition. Kluwer Academic Publisher, Dordrecht, The Netherlands.
- 46. Jones J (2001) Habitat selection studies in avian ecology: A critical review. Auk 118: 557–562. [Google Scholar]
- 47. Pöysä H (2001) Dynamics of habitat distribution in breeding mallards: assessing the applicability of current habitat selection models. Oikos 94: 365–373. [Google Scholar]
- 48. Fretwell SD, Lucas HLJ (1970) On territorial behavior and other factors influencing habitat distributions of birds. Acta Biotheoretica 19: 16–36. [Google Scholar]
- 49. Calenge C (2007) Exploring Habitat Selection by Wildlife with adehabitat. Journal of Statistical Software 22: 2–19. [Google Scholar]
- 50. Guillemain M, Devineau O, Brochet AL, Fuster J, Fritz H, et al. (2010) What is the spatial unit for a wintering teal Anas crecca? Weekly day roost fidelity inferred from nasal saddles in the Camargue, southern France. Wildlife Biology 16: 215–220. [Google Scholar]
- 51. Johnson WP, Rohwer FC (2000) Foraging behavior of green-winged teal and mallards on tidal mudflats in Louisiana. Wetlands 20: 184–188. [Google Scholar]
- 52. Thomas GJ (1982) Autumn and Winter Feeding Ecology of Waterfowl at the Ouse Washes, England. Journal of Zoology 197: 131–172. [Google Scholar]
- 53. Jorde DG, Krapu GL, Crawford RD (1983) Feeding Ecology of Mallards Wintering in Nebraska. Journal of Wildlife Management 47: 1044–1053. [Google Scholar]
- 54. Cox RR, Afton AD (1996) Evening flights of female Northern Pintails from a major roost site. Condor 98: 810–819. [Google Scholar]
- 55. Jorde DG, Krapu GL, Crawford RD, Hay MA (1984) Effects of Weather on Habitat Selection and Behavior of Mallards Wintering in Nebraska. Condor 86: 258–265. [Google Scholar]
- 56. Tamisier A (1978) The functional units of wintering ducks: a spatial integration of their comfort and feeding requirements. Verhandlungen der Ornithologischen Gesellschaft in Bayern 23: 229–238. [Google Scholar]
- 57. Kirby RE, Riechmann JH, Cowardin LM (1985) Home Range and Habitat Use of Forest-Dwelling Mallards in Minnesota. Wilson Bulletin 97: 215–219. [Google Scholar]
- 58. Mack GG, Clark RG (2006) Home-range characteristics, age, body size, and breeding performance of female Mallards (Anas platyrhynchos). Auk 123: 467–474. [Google Scholar]
- 59. Gilmer DS, Ball IJ, Cowardin LM, Riechmann JH, Tester JR (1975) Habitat Use and Home Range of Mallards Breeding in Minnesota. Journal of Wildlife Management 39: 781–789. [Google Scholar]
- 60. Dwyer TJ, Krapu GL, Janke DM (1979) Use of Prairie Pothole Habitat by Breeding Mallards. Journal of Wildlife Management 43: 526–531. [Google Scholar]
- 61. Paracuellos M, Telleria JL (2004) Factors affecting the distribution of a waterbird community: The role of habitat configuration and bird abundance. Waterbirds 27: 446–453. [Google Scholar]
- 62. Paracuellos M (2006) How can habitat selection affect the use of a wetland complex by waterbirds? Biodiversity and Conservation 15: 4569–4582. [Google Scholar]
- 63. Green AJ, Figuerola J, Sanchez MI (2002) Implications of waterbird ecology for the dispersal of aquatic organisms. Acta Oecologica-International Journal of Ecology 23: 177–189. [Google Scholar]
- 64. Green AJ, Elmberg J (2013) Ecosystem services provided by waterbirds. Biological Reviews 89: 105–122. [DOI] [PubMed] [Google Scholar]
- 65. Brochet AL, Guillemain M, Fritz H, Gauthier-Clerc M, Green AJ (2009) The role of migratory ducks in the long-distance dispersal of native plants and the spread of exotic plants in Europe. Ecography 32: 919–928. [Google Scholar]
- 66. Nilsson L (2013) Field choice of autumn staging and wintering geese in south-western Sweden 1977/78 - 2011/12. Ornis Svecica 23: 46–60. [Google Scholar]
- 67. Kery M, Madsen J, Lebreton JD (2006) Survival of Svalbard pink-footed geese Anser brachyrhynchus in relation to winter climate, density and land-use. Journal of Animal Ecology 75: 1172–1181. [DOI] [PubMed] [Google Scholar]
- 68. Hereu B (2005) Movement patterns of the sea urchin Paracentrotus lividus in a marine reserve and an unprotected area in the NW Mediterranean. Marine Ecology - an Evolutionary Perspective 26: 54–62. [Google Scholar]
- 69. Hirsch BT, Morrell LJ (2011) Measuring marginal predation in animal groups. Behavioral Ecology 22: 648–656. [Google Scholar]
- 70. Ripple WJ, Larsen EJ, Renkin RA, Smith DW (2001) Trophic cascades among wolves, elk and aspen on Yellowstone National Park's northern range. Biological Conservation 102: 227–234. [Google Scholar]
- 71. Pöysä H (1987) Feeding-vigilance trade-off in the teal (Anas crecca) - effects of feeding method and predation risk. Behaviour 103: 108–122. [Google Scholar]
- 72. Curio E (1983) Time-energy budgets and optimization. Experientia 39: 25–34. [Google Scholar]
- 73. Houston AI, McNamara JM, Hutchinson JMC (1993) General results concerning the trade-off between gaining energy and avoiding predation. Philosophical Transactions of the Royal Society of London Series B-Biological Sciences 341: 375–397. [Google Scholar]