

Abstract
Human pluripotent stem cells (hPSCs) represent a novel source of hepatocytes for drug metabolism studies and cell-based therapy for the treatment of liver diseases. These applications are, however, dependent on the ability to generate mature metabolically functional cells from the hPSCs. Reproducible and efficient generation of such cells has been challenging to date, owing to the fact that the regulatory pathways that control hepatocyte maturation are poorly understood. Here, we show that the combination of three-dimensional cell aggregation and cAMP signaling enhance the maturation of hPSC-derived hepatoblasts to a hepatocyte-like population that displays expression profiles and metabolic enzyme levels comparable to those of primary human hepatocytes. Importantly, we also demonstrate that generation of the hepatoblast population capable of responding to cAMP is dependent on appropriate activin/nodal signaling in the definitive endoderm at early stages of differentiation. Together, these findings provide new insights into the pathways that regulate maturation of hPSC-derived hepatocytes and in doing so provide a simple and reproducible approach for generating metabolically functional cell populations.
Keywords: Endoderm, Hepatocyte, Human pluripotent stem cell, Maturation, Three dimensional culture, cAMP signaling
INTRODUCTION
The ability to direct the differentiation of human pluripotent stem cells (hPSCs) (embryonic and induced pluripotent stem cells) to specific lineages in culture does provide access to unlimited numbers of human primary cells for a wide range of applications that include the development of new treatments for a spectrum of diseases, the establishment of platforms for drug discovery and predictive toxicology, and the creation of in vitro models of human disease. Among the different lineages that can be derived from hPSCs, hepatocytes are of particular importance as they are the cells responsible for drug metabolism and thus for controlling xenobiotic elimination from the body (Guillouzo, 1998; Gebhardt et al., 2003; Hewitt et al., 2007). Given this role and the fact that individuals can differ in drug-metabolizing capacity (Byers et al., 2007), access to functional hepatocytes from a representative population sample would greatly facilitate drug discovery and testing within the pharmaceutical industry. Additionally, hepatocyte transplantation and bio-artificial liver devices developed with hPSC-derived hepatocytes represent potential life-saving therapies for individuals with specific types of liver disease who have no available matched donor organ.
Given the potential therapeutic and commercial importance of functional human hepatocytes, significant effort has been directed towards optimizing protocols for the generation of these cells from hPSCs over the past five years (Cai et al., 2007; Duan et al., 2007; Hay et al., 2008; Basma et al., 2009; Duan et al., 2010; Si-Tayeb et al., 2010b; Sullivan et al., 2010; Touboul et al., 2010; Chen et al., 2012). Almost all approaches have attempted to recapitulate the key stages of liver development in the differentiation cultures, including the induction of definitive endoderm, the specification of the endoderm to a hepatic fate, the generation of hepatic progenitors known as hepatoblasts and the differentiation of hepatoblasts to mature hepatocytes (Si-Tayeb et al., 2010a). In most studies, differentiation is induced in a monolayer format with the sequential addition of pathway agonists and antagonists that are known to regulate the early stages of development, including endoderm induction and hepatic specification. In contrast to the early developmental steps, the signaling pathways that promote the maturation of the hPSC-derived hepatocytes to functional cells as defined by Phase I and Phase II drug-metabolizing enzyme activity have not been well defined. As a consequence, the populations produced with the different protocols vary considerably in their maturation status and in most cases represent immature hepatocytes. The inability to reproducibly generate mature cells represents a significant bottleneck in the field, as drug development applications require cells that display functional levels of key drug-metabolizing enzymes.
In this study, we have addressed the issue of maturation by manipulating specific signaling pathways at different stages of hepatic development in hPSC differentiation cultures. We show that sustained activin/nodal signaling is important for appropriate patterning of the definitive endoderm population for hepatic specification, and that three-dimensional (3D) cellular aggregation of hepatoblast-stage cells initiates maturation of this progenitor population. Finally, we demonstrate that cAMP signaling within the 3D hepatoblast aggregates promotes further maturation to functional stage cells, as demonstrated by the upregulation of selected drug-metabolizing enzymes, including several Phase I cytochrome P450 and the Phase II enzymes that are responsible for the metabolism of many clinically and toxicologically important drugs.
MATERIALS AND METHODS
Human PSC culture and differentiation
hPSCs were maintained on irradiated mouse embryonic feeder cells in hESC medium as described previously (Kennedy et al., 2007). Prior to the generation of embryoid bodies (EBs), hESCs were passaged onto Matrigel-coated plates for 1 day to deplete the population of feeder cells and then dissociated with 0.25% Trypsin-EDTA to generate small clusters as previously described (Kennedy et al., 2007; Nostro et al., 2011). The clusters were cultured in serum-free differentiation (SFD) medium in the presence of BMP4 (3 ng/ml) for 24 hours (day 0 to day 1) and then in differentiation medium consisting of StemPro-34 supplemented with glutamine (2 mM), ascorbic acid (50 μg/ml; Sigma), MTG (4.5×10-4 M; Sigma), basic fibroblast growth factor (bFGF; 2.5 ng/ml), activin A (100 ng/ml), Wnt3a (25 ng/ml) and BMP4 (0.25 ng/ml) for 3 days. On day 4, the medium was changed and the amount of bFGF was increased to 5 ng/ml for an additional 48 hours of culture. At this stage, the EBs were harvested and dissociated with 0.25% Trypsin-EDTA and the cells cultured for 2 days on Matrigel-coated 12-well plates at a concentration of 4×105 cells in above differentiation medium without Wnt3A and a reduced amount of activin (50 ng/ml). On day 8, the differentiation medium was replaced with hepatic specification medium that consisted of Iscove’s Modified Dulbecco’s Medium (IMDM) supplement with 1% vol/vol B27 supplement (Invitrogen: A11576SA), ascorbic acid, MTG, FGF10 (50 ng/ml) (from day 8 to day 10), bFGF (20 ng/ml) (from day 10 to day 14) and BMP4 (50 ng/ml) (from day 8 to day 14). The medium was changed every 2 days until day 14 at which stage it was changed to maturation medium that consisted of IMDM with 1% vol/vol B27 supplement, ascorbic acid, glutamine, MTG, hepatocyte growth factor (HGF) (20 ng/ml), dexamethasone (Dex) (40 ng/ml) and oncostatin M (20 ng/ml). On day 26, the cells were dissociated with enzymatic treatment (collagenase type 1: Sigma C0130) and manual dissociation, and then cultured in six-well ultra-low cluster dishes at a concentration of 6×105 cells per well in above maturation medium supplemented with Rho-kinase inhibitor and 0.1% BSA to generate 3D aggregates. Aggregates were maintained under these conditions for 6 days, with medium changes every 3 days. On day 32, aggregates were cultured in hepatocyte culture medium (HCM) (Lonza: CC-4182) without EGF to promote the final stages of maturation. At this time point, 1 mM 8 bromo-cAMP (Biolog: B007) was added and the medium was changed every 3 days. To generate hepatocyte-like cells from H9 hESCs, H1 hESCs and 38-2 IPSCs, the following changes (summarized in supplementary material Table S1) were made to the hepatic specification medium. The concentration of bFGF was increased to 40 ng/ml and the base medium was switched from IMDM to H16 DMEM for culture from days 8-14 and then to H16 DMEM plus 25% Ham’s F12 and 0.1% BSA from days 14-20. IMDM was replaced with H21 DMEM plus 25% Ham’s F12 and 0.1% BSA for the maturation medium used from days 20-32. All cytokines were human and purchased from R&D Systems, unless stated otherwise. EB and monolayer cultures were maintained in a 5% CO2/5% O2/90% N2 environment. Aggregation cultures were maintained in a 5% CO2/ambient air environment.
Flow cytometry
Flow cytometric analyses were performed as described (Nostro et al., 2011). For cell surface markers, staining was carried out in PBS with 10% FCS, whereas for intracellular proteins, staining was performed on cells fixed with 4% paraformaldehyde (Electron Microscopy Science, Hatfield, PA, USA) in PBS. The conditions for SOX17 and FOXA2 staining were as previously described (Nostro et al., 2011). Albumin and α-fetoprotein staining was carried out in PBS with 10% FCS and 0.5% saponin (Sigma). Stained cells were analyzed using an LSRII flow cytometer (BD). The sources and concentrations of primary, secondary and isotype control antibodies are listed in supplementary material Table S7.
Immunostaining
To detect albumin and α-fetoprotein-positive cells, the populations were stained for 1 hour at room temperature with either a goat anti-ALB (Bethyl) or a rabbit anti-AFP antibody (DAKO). Concentrations of isotype controls were matched to primary antibodies. To visualize the signal, the cells were subsequently incubated for 1 hour at room temperature with either a donkey-anti-goat Alexa 488 (Invitrogen) or a donkey anti-rabbit-Cy3 antibody (Jackson ImmunoResearch). For SOX17 staining, the cells were fixed, permeabilized and blocked as described above. The stained cells were visualized using a fluorescence microscope (Leica CTR6000) and images captured using the Leica Application Suite software. For staining of the aggregates, they were fixed with 4% PFA at 37°C overnight, washed with normal saline (0.85% NaCl) and then embedded in 2% agar. The agar block was fixed with 10% PFA for 24 hours and embedded in a paraffin block and sectioned. For immunohistochemistry, the paraffin-embedded sections were dewaxed with xylene, rehydrated, placed in Tris-EGTA-buffer (TES; 10 mM Tris, 0.5 mM EGTA, pH 9.0) and subjected to heat-induced (microwave) epitope retrieval. The tissues were blocked by incubation with 10% normal donkey (Jackson ImmunoResearch) (for ALB/ASGR1, ALB/AFP and ALB/HNF4α staining) or goat serum (Jackson ImmunoResearch) (for ALB/E-cadherin staining) for 30 minutes. They were subsequently incubated with goat anti-ASGR1 (Santa Cruz), mouse-anti-E cadherin (BD) and goat anti-HNF4α (Santa Cruz) overnight at 4°C and then with anti-ALB and anti-AFP antibodies for 1 hour at room temperature. For double staining of ALB/ASGR1, ALB/AFP and ALB/HNF4α, the signals were visualized using donkey anti-goat Alexa 488 and anti-rabbit Cy3 antibodies. For ALB/E cadherin, the signals were visualized using goat anti-rabbit Alexa 488 and goat anti-mouse Cy3. The stained cells were analyzed using a confocal fluorescence microscope (Olympus Fluo View 1000 B laser scanning confocal) and images captured using the Olympus Application software. Primary and secondary antibodies were diluted in PBS+0.2% BSA+0.05% Triton-X100. Prolong Gold Antifade with DAPI (Invitrogen) was used to counterstain the nuclei.
Generation and dissociation procedure of 3D aggregates
Aggregates were generated from the monolayer by a combination of enzymatic treatment (collagenase type 1: Sigma C0130) and manual dissociation. For dissociation of the day 44 aggregates, they were incubated by gentle shaking overnight at room temperature in Hank’s solution containing 1 mg/ml collagenase Type II (Worthington #LS004176). On the following day, the solution was replaced with fresh dissociation medium consisting of Hank’s solution supplemented with 10 mM taurine, 0.1 mM EGTA, 1 mg/ml BSA and 1 mg/ml collagenase type II. The cells were dissociated by gentle pipetting.
Quantitative real-time PCR
Total RNA was prepared using RNA aqueous Micro Kit (Ambion) and treated with RNase-free DNase (Ambion). RNA (500 ng to 1 μg) was reverse transcribed into cDNA using random hexamers and Oligo(dT) with Superscript III Reverse Transcriptase (Invitrogen). QPCR was performed on a MasterCycler EP RealPlex (Eppendorf) using a QuantiFast SYBR Green PCR Kit (Quiagen) as described previously (Nostro et al., 2011). Expression levels were normalized to the housekeeping gene TATA box-binding protein (TBP). For UGT1A1, expression was calculated using the delta-delta CT method relative to the level in non-treated (8-Br-cAMP) cells. Oligonucleotide sequences are available in supplementary material Table S8. For controls, two samples of total adult and fetal liver RNA were purchased from Clontech (AL1, FL1), Agilent Technologies (AL2) and BioChain (FL2). Two of the primary hepatocyte samples (HH1892 and HH1901) used for RNA analyses were generated by culture of freshly isolated hepatocytes, as previously described (Kostrubsky et al., 1999). A third sample was purchased from Zenbio (lot: 2199). All RNA information is available in supplementary material Table S8.
Indocyanine green uptake
The indocyanine green (ICG, Sigma) solution was dissolved in HCM (Lonza) at a concentration of 5 mg/ml and added to the cells at final concentration of 1 mg/ml ICG in HCM. The cells were incubated at 37°C for 1 hour, washed three times with PBS and then examined with an inverted Microscope (Leica). To monitor release of the ICG, the cells were cultured in fresh medium without ICG for an additional for 24 hours.
Periodic acid-Schiff staining for glycogen
Cultured cells were fixed in 4% PFA for 15 minutes and stained according to the manufacturer’s instructions using a Periodic acid-Schiff (PAS) staining kit (Sigma).
Albumin secretion assay
Medium was harvested following 24 hours of culture of the different cell populations and the amount of albumin secreted was measured according to the manufacturer’s protocol using the Human Albumin ELISA Quantitation kit (Bethy Laboratories).
Drug metabolism assay by HPLC
Three lots of cryopreserved human hepatocytes (Celsis In Vitro Technologies, Baltimore, USA, lot No ONQ, OSI and JGM) were used as control. Cells were thawed and cultured in type 1 collagen-coated microtiter wells (5×104 cells per well) in InVitroGro HI medium (Celsis In Vitro Technologies) (Roymans et al., 2004). Following hepatocyte attachment (2-4 hours) the nonadherent dead cells were removed and replaced with fresh medium. To measure CYP1A2 and CYP3A4 induction, the primary hepatocytes (lots OSI and JGM) or hPSC-derived aggregates were treated with either lansoprazole (10 μM) or rifampicin (10 μM) for 72 hours. Medium with fresh inducer was changed daily. Following induction, the cells were incubated in the medium containing either the CYP1A2 substrate phenacetin (200 μM) or the CYP3A4 substrate testosterone (250 μM) for 24 or 2 hours, respectively. After incubation, aliquots of the medium were collected and the levels of metabolites were quantified by high-performance liquid chromatography. Controls were cultured with DMSO alone (final concentration 0.1%). Following the metabolic assays, the aggregates were harvested, dissociated and the cells counted.
To measure the CYP2B6, NAT1/2 and Total UGT activities, the hPSC-derived aggregates and cryopreserved hepatocytes (lot ONQ) were incubated in medium containing either the CYP2B6 substrate bupropion (900 μM), the NAT2-selective substrate sulfamethazine (SMZ) (500 μM) or the total UGT substrate 4-methylumbeliferone (4-MU) (200 μM) for either 24 or 48 hours. Hydroxybupropion, N-acetyl-SMZ and 4-MU glucuronide levels were quantified by HPLC using the methods of Loboz et al. (Loboz et al., 2005), Grant et al. (Grant et al., 1991) and Gagné et al. (Gagné et al., 2002), respectively.
Microarray processing and data analysis
RNA samples were run on Affymetrix Human Gene ST v1.0 chips following standard Affymetrix guidelines at the University Health Network Genomics Centre. Briefly, 300 ng of total RNA starting material for each sample was used as input to the Ambion WT Expression Kit. Amplified cDNA (2.7 μg) was then fragmented, labeled and hydridized to Affymetrix Human Gene ST v1.0 chips for 18 hours (45°C at 60 rpm). Arrays were washed using a GeneChip Fluidics Station and scanned with an Affymetrix GeneChip Scanner 7G. Raw CEL files were imported into Genespring (Agilent, v11.5.1) and probe level data were summarized using the ExonRMA16 algorithm based on the HuGene-1_0-st0v1_na31_hg19_2010-09-03 build. Each gene was normalized to the median value across all samples under consideration. All statistics were performed on log 2 transformed data. In total, 28,869 transcripts are represented on this array.
As a first step, transcripts were filtered to remove those that were consistently in the lower 20th percentile of measured expression across all of the three sample groups. An unsupervised hierarchical clustering analysis with a Pearson centered distance metric under average linkage rules was used to address overall similarity and differences between the samples and groups. Directed statistical analysis between the three sample groups was performed by ANOVA with a Benjamini and Hochberg False Discovery Rate (FDR, q<0.05) (Klipper-Aurbach et al., 1995). To find sets of differentially expressed transcripts with biological meaning, a gene ontology (GO) analysis was performed using a corrected Benjamini and Yuketieli hypergeometric test at the q<0.1 significance level (Benjamini et al., 2001). Two a priori defined sets of specific transcripts were examined in more detail: transcripts related to specific liver-related activity of interest; and transcripts found to be expressed and liver specific based on publicly available information from the HOMER database (Zhang and Chen, 2011). Microarray data have been deposited in GEO with Accession Number GSE39157.
RESULTS
Endoderm induction in EBs
The protocol to generate functional hepatocyte-like cells was developed with the HES2 hESC line using the embryoid body (EB) format for the early induction steps (Fig. 1A). Using this approach, EBs generated in the presence of BMP4 (24 hours) are exposed to activin A (activin) and Wnt3a for 5 days to induce definitive endoderm, a population defined by expression of the surface markers CXCR4, CKIT and EPCAM and the transcription factors SOX17 and FOXA2. Under optimal conditions, CKIT+CXCR4+, SOX17+ and FOXA2+ cells were detected within the EBs by day 3 of differentiation, and their number increased dramatically over the next 72 hours (Fig. 1B). By day 6 of differentiation, over 95% of the induced EB population co-expressed CXCR4 and CKIT or CXCR4 and EPCAM (Fig. 1B; supplementary material Fig. S1A). Intracellular flow cytometric analyses revealed that more than 95% of the cells expressed SOX17 and greater than 85% were FOXA2+ at this stage. Base culture medium influenced the efficiency of endoderm induction, as culture in neural basal medium rather than StemPro34 resulted in the generation of EBs with lower percentages of endoderm (supplementary material Fig. S1B). The expansion of the endoderm population was preceded by the transient expression of the primitive streak gene T, and accompanied by the upregulation of expression of genes that mark endoderm development, including SOX17, goosecoid (GSC) and FOXA2 (supplementary material Fig. S1C). The generation of highly enriched endoderm is an important first step in the protocol, as induction levels of less than 90% CXCR4+CKIT+ and 80% SOX17+ cells results in suboptimal hepatic lineage development (data not shown).
Fig. 1.
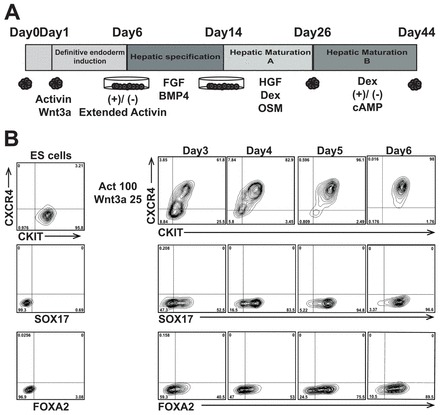
Endoderm induction in hESC-derived embryoid bodies. (A) The differentiation protocol. (B) Flow cytometric analyses showing the kinetics of development of the CXCR4+, CKIT+, SOX17+ and FOXA2+ populations in the activin/Wnt3a-induced EBs.
Duration of nodal/activin signaling impacts hepatic development
To specify the CXCR4+CKIT+ population to a hepatic fate, day 6 EBs were dissociated and the cells plated as a monolayer on Matrigel-coated plates in the presence of FGF10 and BMP4 for 48 hours, and then in bFGF and BMP4 for 6 days. As previously demonstrated in mouse and human ESC cultures (Gouon-Evans et al., 2006; Si-Tayeb et al., 2010b), the combination of BMP and FGF signaling was required for optimal hepatic induction under our conditions (supplementary material Fig. S2A). The FGF10/BMP4 step was included as it was found to increase albumin expression in the differentiation cultures (supplementary material Fig. S2B).
Although these induction conditions did lead to the development of albumin-positive cells by day 24 of differentiation (supplementary material Fig. S2C), they were not optimal as the proportion of SOX17+ and FOXA2+ cells within the culture had decreased from more than 90% to ∼50% by day 10 (Fig. 2A). As we have previously demonstrated that prolonged activin/nodal signaling promotes endoderm development in mouse ESC differentiation cultures (Gadue et al., 2006), we extended the activin induction step for an additional 2 days prior to the FGF/BMP4 specification step in an attempt to sustain the endoderm population in the human cultures. Extended activin signaling did lead to a significant increase in the proportion of SOX17+ and FOXA2+ cells detected at day 12 (Fig. 2A; supplementary material Fig. S3A). The prolonged activin treatment reduced the total cell number in the cultures (Fig. 2B), suggesting that these conditions may preferentially support the survival of endodermal cells.
Fig. 2.
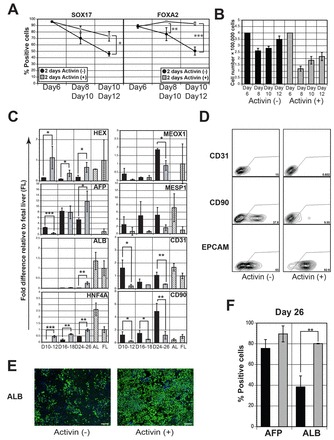
Duration of nodal/activin signaling impacts hepatic development. (A) Intracellular flow cytometric analysis showing the proportion of SOX17+ and FOXA2+ cells in day 6 activin/Wnt3A-induced EBs, as well as in monolayer populations derived from them. The monolayer populations were cultured either directly in the specification medium (-activin) or for 2 days in activin (50 ng/ml) and then in the specification medium (+activin). Populations were analyzed following 2 or 4 days of culture in the specification medium (total days: 8 and 10 for the -activin group; 10 and 12 for the +activin group). Bars represent s.d. of the mean of three independent experiments. (B) Total cell number in activin-treated and non-treated monolayer cultures. (C) RT-qPCR-based expression analyses of hepatic monolayer populations generated from activin-treated (black bars) and non-treated (gray bars) endoderm. Activin-treated populations (gray bars) were analyzed at days 12, 18 and 26 of total culture, whereas the non-treated population (black bar) was analyzed at days 10, 16 and 24 of culture. Values are determined relative to TBP and presented as fold change relative to expression in fetal liver, which is set at 1. AL (adult liver): n=2, AL1, AL2. FL (fetal liver): n=2, FL1, FL2. (D) Flow cytometric analysis showing the proportion of CD31+, CD90+ and EPCAM+ cells in monolayer populations derived from activin-treated (day 26) and non-treated (day 24) endoderm. The CD31+ and CD90+ populations were significantly larger in non-treated compared with the treated cultures (CD31, 13.6±2.3% versus 0.49±0.11%, P<0.001; CD90, 41.2±4.7% versus 8.5±1.19%, P<0.001, Student’s t-test, n=3). By contrast, a higher proportion of EPCAM+ cells was detected in the population derived from the activin-treated endoderm compared with the population generated from the non-treated cells (EPCAM, 90.7±2.7% versus 56.8±7.3%; P<0.01, n=3). (E) Immunostaining analyses showing the proportion of albumin-positive cells in cultures generated from activin-treated (day 26) and non-treated (day 24) endoderm. Albumin is visualized with Alexa 488 (green), nuclei are shown following staining with DAPI (blue). Scale bars: 200 μm. (F) Intracellular flow cytometric analyses indicating the proportion of albumin (ALB) and α-fetoprotein (AFP) cells in monolayer cultures generated from activin-treated (gray bars; day 26) and non-treated (black bars; day 24) endoderm. Error bars in all figures represent the s.d. of the mean of three independent experiments. *P<0.05, **P<0.01, ***P<0.001 (Student’s t-test; n=3).
The extended activin culture maintained the CXCR4+CKIT+ population until day 8 (supplementary material Fig. S3B) and resulted in higher levels of expression of genes indicative of hepatic progenitor (hepatoblast) development, including HEX (HHEX - Human Gene Nomenclature Database), AFP, ALB and HNF4A at day 26 (Fig. 2C). Cultures generated from non-treated CXCR4+CKIT+ endoderm contained contaminating mesoderm, as demonstrated by the expression of MEOX1, MESP1, CD31 (PECAM1 - Human Gene Nomenclature Database) and CD90 (THY1 - Human Gene Nomenclature Database), and by the presence of CD90+ cells and CD31+ endothelial cells at day 24 (Fig. 2C,D). Populations derived from the activin-treated endoderm showed reduced expression of the mesoderm genes, had a higher proportion of EPCAM+ cells, no detectable CD31+ cells and a much smaller CD90 population (Fig. 2D). Consistent with these differences, we observed a significantly higher proportion of albumin-positive cells in the treated compared with the non-treated population at day 26 of culture (Fig. 2E,F). The number of AFP-positive cells was not different between the two groups.
Aggregation promotes hepatic maturation
As previous studies have shown that cell aggregation can promote some degree of maturation of hESC-derived hepatic cells (Miki et al., 2011; Nagamoto et al., 2012; Sivertsson et al., 2013; Takayama et al., 2013), we next generated aggregates from the day 26 population (Fig. 3A) and cultured them for 6 days in the presence of HGF, dexamethasone (Dex) and oncostatin M (OSM) to determine whether these conditions would affect maturation of the hepatoblast-like cells in our cultures. Aggregation did increase the expression of a number of genes associated with liver function, including albumin, CPS1 (carbamonyl-phosphatase synthase 1), TAT (tyrosine aminotransferase), G6P (glucose 6 phosphatase) and TDO2 (tryptophan 2,3-dioxygenase) over that observed in monolayer culture. The expression of some (CYP7A1, CYP3A7 and CYP3A4) but not all (CYP1A2 and CYP2B6) P450 genes was upregulated by aggregation (Fig. 3C and data not shown). In addition to the different enzyme genes, aggregation also increased the proportion of cells expressing asialo-glycoprotein receptor 1 (ASGR1) a cell-surface marker found on mature hepatocytes (Basma et al., 2009) (Fig. 3D). Immunostaining showed that ASGR1 was detected on albumin+ cells (Fig. 3E). The albumin+ cells in the aggregates also expressed E-cadherin, indicating that they had acquired epithelial characteristics, a property of hepatocytes found in the intact liver (Fig. 3F). The ability to store glycogen, as measured by PAS staining, was not dependent on aggregation, as both monolayer cells and aggregates displayed this capacity (supplementary material Fig. S4). Collectively, these findings show that the simple process of aggregation into 3D structures promotes changes indicative of hepatic maturation.
Fig. 3.
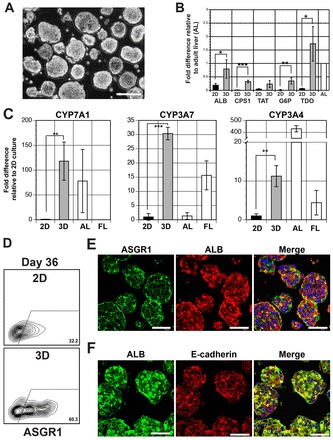
Aggregation promotes hepatoblast maturation. (A) Phase-contrast image of hepatic aggregates at day 28 of culture. Scale bar: 200 μm. (B) RT-qPCR based analyses of expression of indicated genes in monolayer (black bar) and 3D aggregate cultures (gray bar) at day 32 of differentiation. Values are determined relative to TBP and presented relative to expression in adult liver, which is set at 1. (C) RT-qPCR based analysis for CYP7A1, CYP3A7 and CYP3A4 expression at day 32 of differentiation in monolayer (black bar) and 3D aggregate cultures (gray bar). Values are determined relative to TBP and presented as fold change relative to expression in monolayer (2D) cells, which is set at one. AL (adult liver): n=2, AL1, AL2. FL (fetal liver): n=2, FL1, FL2. (D) Flow cytometric analysis showing the proportion of ASGR1 cells in the monolayer (2D) and aggregate (3D) cultures at day 36. The number of ASGR1+ cells was significantly higher in 3D aggregate cultures (2D, 28.8±3.1%; 3D, 64.7±4.26%, P<0.001, n=3). (E) Confocal microscopic images of immunostained day 32 aggregates showing co-expression of albumin and ASGR1. Albumin is visualized by Cy3 (red), ASGR1 by Alexa 488 (green) and the nuclei by DAPI (blue). Scale bars: 50 μm. (F) Confocal microscopic images of immunostained day 32 aggregates showing co-expression of albumin and E-cadherin. Albumin is visualized by Alexa 488 (green), E-cadherin by Cy3 (red) and the nuclei by DAPI (blue). Scale bars: 50 μm. Error bars in all graphs represent the s.d. of the mean of samples from three independent experiments, *P<0.05, **P<0.01, ***P<0.001, Student’s t-test.
cAMP signaling induces maturation of hESC-derived hepatocyte-like cells
To further mature the cells, we investigated the role of cAMP signalling, as studies using hepatic cell lines have shown that activation of this pathway induces hepatic gene expression, in part through the induction of the peroxisome proliferator-activated receptor γ co-activator 1α (PGC1A; PPARGC1A - Human Gene Nomenclature Database), a co-activator that functions together with HNF4A to regulate the expression of many genes involved in hepatocyte function (Bell and Michalopoulos, 2006; Arpiainen et al., 2008; Benet et al., 2010; Dankel et al., 2010). Treatment of the aggregates with 8-bromoadenosine-3′,5′-cyclic monophosphate (8-Br-cAMP), a cell-permeable analogue of cAMP, from days 32 to 44 of culture significantly enhanced the expression of PGC1A (15-fold), G6P (25-fold) and TAT (33-fold) but not that of HNF4A (Fig. 4A). By contrast, the expression levels of AFP and ALB were downregulated by 8-Br-cAMP. Flow cytometric analyses confirmed the AFP expression analyses and showed a reduction in the number of AFP-positive cells in the 8-Br-cAMP treated aggregates, compared with the non-treated controls. The proportion of ALB-positive cells was not reduced despite a decline in mRNA levels (Fig. 4B). These differences could reflect differences in RNA versus protein expression. The addition of 8-Br-cAMP did not impact the structure of the aggregates (supplementary material Fig. S5A) or the viability of the cells (typically >70%) during this culture period.
Fig. 4.
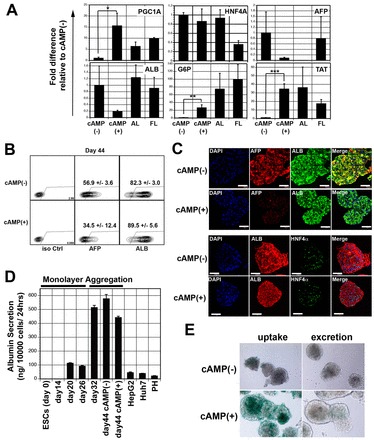
cAMP induces maturation of hESC-derived hepatocyte-like cells. (A) RT-qPCR-based expression analysis of indicated genes in hepatic aggregates cultured in the presence and absence of 8-Br-cAMP. Values are determined relative to TBP and presented as fold change relative to expression of the non-treated cells which is set at 1. AL (adult liver): n=2, AL1, AL2. FL (fetal liver): n=2, FL1, FL2. (B) Intracellular flow cytometric analysis showing the proportion of α-fetoprotein (AFP)+ and albumin (ALB)+ cells (day 44) in hepatic aggregates cultured in the presence and absence of 8-Br-cAMP. The number of AFP+ cells was significantly lower in the population induced with cAMP compared with the non-induced population (34.5±12.4% versus 56.9±3.6%, P<0.05, mean±s.d., n=3), whereas the proportion of ALB+ cells was higher in the treated population [89.5±5.6% versus 82.3±3.0%, P<0.05 (mean±s.d., n=3)]. (C) Confocal microscopic images showing co-expression of ALB and AFP or ALB and HNF4α in day 44 aggregates cultured in the presence and absence of 8-Br-cAMP. In the upper panel, albumin is visualized by Alexa 488 (green), AFP by Cy3 (red) and the nuclei by DAPI (blue). In the lower panel, albumin is visualized by Cy3 (red), HNF4α by Alexa 488 (green) and the nuclei by DAPI (blue). Scale bars: 50 μm. (D) The levels of albumin (ALB) secreted by hESC-derived monolayer and aggregate populations, as well as by HepG2 cells, Huh7 cells and cryopreserved hepatocytes (PH, lot OSI). Secretion was detected using an ELISA assay. (E) ICG uptake and release by cAMP-treated and non-treated day 44 aggregates. Error bars in all graphs represent the s.d. of the mean of the values from three independent experiments. *P<0.05, **P<0.01, ***P<0.001, Student’s t-test,
Immunostaining analyses were consistent with the flow cytometry data and showed that cAMP-treated aggregates expressed similar levels of ALB but lower levels of AFP compared with the non-treated ones (Fig. 4C). The levels of HNF4α protein in both aggregate populations were comparable, confirming the PCR analyses. Albumin secretion by the hESC-derived cells was not impacted by 8-Br-cAMP treatment but was dramatically enhanced by the aggregation step. (Fig. 4D). By contrast, the capacity to take up indocyanine green (ICG), a characteristic of adult hepatocytes (Stieger et al., 2012) was enhanced by cAMP signaling (Fig. 4E).
Other tissues, such as the pancreas, also express PGC1A. However, in contrast to the observed induction in hepatic cells, expression of PGC1A was not induced by cAMP signaling in hESC-derived insulin-positive pancreatic cells (supplementary material Fig. S5B), indicating that this response may be tissue specific.
cAMP signaling increases metabolic enzyme activity in hESC-derived hepatocytes
cAMP signaling also induced changes in the expression pattern of key Phase I cytochrome P450 genes, notably a reduction in the levels of expression of the fetal gene CYP3A7, and a significant increase in expression of the adult genes CYP3A4 (2.5-fold), CYP1A2 (18-fold) and CYP2B6 (4.7-fold) (Fig. 5A). UGT1A1, an important Phase II enzyme, was also significantly induced (11-fold) by 8-Br-cAMP (Fig. 5A). The inductive effects of cAMP signaling on the P450 genes were observed only in cells in the 3D aggregates, as little increase in expression of CYP1A2 and CYP3A4 was detected when it was added to monolayer cultures (Fig. 5B). Expression of PGC1A and TAT was induced in the monolayer format, likely due to the fact that the promoter regions of these genes contain cAMP-response element binding protein (CREB) sites.
Fig. 5.
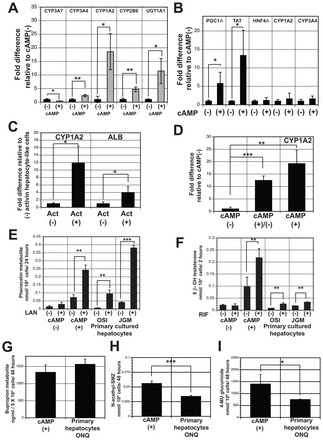
cAMP increases metabolic enzyme activity in hESC-derived hepatocytes. (A) RT-qPCR analysis showing expression of indicated genes in hepatic aggregates (day 44) cultured in the presence and absence of 8-Br-cAMP. Values are determined relative to TBP and presented as fold change relative to expression in non-treated cells, which is set at 1. (B) RT-qPCR analyses showing expression of indicated genes in untreated (-) and cAMP-treated (+) monolayer populations (day 44). Values are determined relative to TBP and presented as fold change relative to expression in non-treated cells, which is set at 1. (C) RT-qPCR analyses of CYP1A2 and ALB expression in cAMP-treated aggregates (day 44) generated from non-treated (-Act) or extended activin treated (+Act) endoderm. (D) RT-qPCR analyses of CYP1A2 expression in aggregates cultured for 6 (cAMP+/-) or 12 days in 8-Br-cAMP (cAMP+). (E) hESC-derived hepatic cells display lansoprazole (LAN)-inducible CYP1A2 activity in vitro. Generation of the O-de-ethylated metabolite acetaminophen from phenacetin was monitored by HPLC. Activity is presented per 10,000 cells. Non-induced cells (-) were cultured in 0.1% DMSO-containing medium (n=3). (F) hESC-derived hepatic cells display rifampicin (RIF)-inducible CYP3A4 activity in vitro. Generation of the 6β-hydroxytestosterone from testosterone was monitored by HPLC. Activity is presented per 10,000 cells. Non-induced cells (-) were cultured in 0.1% DMSO-containing medium (n=3). (G) hESC-derived hepatic cells display CYP2B6 activity in vitro. Formation of the metabolite O-hydroxy-bupropion from bupropion was measured by HPLC. Activity is presented per 50,000 cells (n=3). (H) Metabolism of sulfamethazine (SMZ) to N-acetylated SMZ indicates the presence of the Phase II enzyme(s) NAT2 and/or NAT1. Activity is presented per 10,000 cells (n=3). (I) HPLC analysis showing generation of 4-MU glucuronide (4-MUG) from 4-methylumbelliferone (4-MU) by the cAMP-treated aggregates indicative of total UGT activity. Activity is presented per 10,000 cells (n=3). Error bars in all graphs represent the s.d. of the mean of samples from three independent experiments. *P<0.05, **P<0.01, ***P<0.001, Student’s t-test. OSI, JGM and ONQ are three different lots of cultured primary hepatocytes.
cAMP signaling appeared to be most effective on highly enriched, appropriately patterned cells, as demonstrated by the fact that the levels of induction of CYP1A2 and ALB expression were significantly higher in the aggregates from the extended-activin treated endoderm (+Act) compared with the aggregates from the non-treated endoderm (-Act) (Fig. 5C). To determine whether the changes in gene expression are dependent on continuous signaling, cells induced with 8-Br-cAMP for 6 days and then maintained in the absence of 8-Br-cAMP for the remaining 6 days were compared with those cultured for the entire 12 days in 8-Br-cAMP (Fig. 5D). Expression of CYP1A2 was maintained following the shorter induction time, demonstrating that the higher levels of expression are not dependent on continuous signaling but rather reflect changes indicative of hepatocyte maturation.
To investigate the functional activity of the P450 enzymes, we determined the ability to metabolize isozyme-selective marker drugs. Additionally, the inducibility of the metabolic activity of two of the key enzymes, CYP1A2 and CYP3A4, was also evaluated. As shown in Fig. 5E, the 8-Br-cAMP-treated cells were able to metabolize the CYP1A2-selective substrate phenacetin. Induction of the cells with lansoprazole for 72 hours resulted in a 3.4-fold increase in this activity. The non-treated (8-Br-cAMP) cells had low levels of activity that were not inducible. Two independent primary hepatocyte samples showed lower or comparable levels of basal metabolic activity, but did display higher levels of induction (18- and 9-fold). CYP3A4 activity was measured by the ability of the cells to metabolize testosterone to 6β-hydroxyl testosterone. As shown in Fig. 5F the 8-Br-cAMP-treated cells displayed this activity. Addition of the CYP3A4 inducer rifampicin increased the activity 2.2-fold, indicating that this enzyme was also inducible in the hESC-derived cells. As observed with CYP1A2, little CYP3A4 activity was detected in the non-induced cells. The primary hepatocytes showed low but significant levels of CYP3A4 induction.
CYP2B6 activity, as measured by the hydroxylation of bupropion was also detected in the 8-Br-cAMP-treated cells, at levels comparable with those found in primary hepatocytes (Fig. 5G). Analyses of phase II metabolic enzymes, including the arylamine N-acetyltransferases NAT2 and/or NAT1 (Fig. 5H) and UDP-glucuronosyltransferase (UGT) (Fig. 5I) revealed activity higher than that of cryopreserved primary cultured hepatocytes, indicating that cAMP signaling induced the upregulation of expression of a broad range of enzymes, consistent with maturation of the population. Together, these observations indicate that cAMP signaling promotes maturation of the hESC-derived hepatocyte-like cells in the 3D aggregates to metabolically functional cells.
Hepatic specification and maturation from other hPSC lines
When induced with the above EB-based protocol, the hESC lines H9 and H1 and the induced pluripotent cell (iPSC) line 38-2 generated populations that contained high frequencies of CKIT+CXCR4+ and CKIT+EPCAM+ cells (Fig. 6A). Differences were, however, observed in the proportion of cells within the EBs that expressed SOX17+ and FOXA2+ (Fig. 6A), indicating that surface marker analysis alone is not sufficient to monitor endoderm development. Extended activin/nodal signaling also improved hepatic development of the CKIT+CXCR4+ population from these hPSC lines; however, the time of treatment necessary to generate significant levels of ALB-positive cells varied between them. Whereas populations consisting of 90% ALB+ cells were obtained following 2 days of activin treatment with H9-derived cells, both H1 and 38-2 cells required 4 days of additional activin signaling to generate populations that contained at least 70% ALB+ cells (Fig. 6B). H9-derived cells at day 26 of differentiation showed a cobblestone morphology very similar to that of cultured hepatocytes (supplementary material Fig. S6B). Both the H9 and 38-2-derived hepatocytes stained with PAS, demonstrating their ability store glycogen (supplementary material Fig. S6C).
Fig. 6.
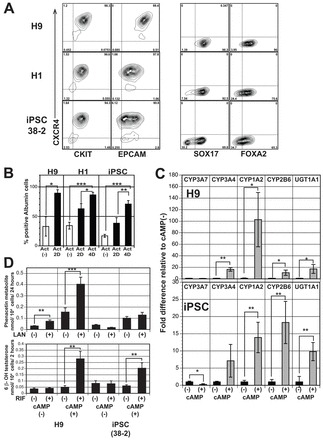
Hepatic specification and maturation from other hPSCs lines. (A) Flow cytometric analyses showing the proportion of CXCR4+, CKIT+, EPCAM+, SOX17+ and FOXA2+ cells in activin/Wnt3a-induced day 6 EBs generated from H9 hESCs, H1 hESCs and 38-2 iPSCs. (B) Intracellular flow cytometric analyses showing the number of ALB-positive cells generated from the different hPSC lines [no activin (-), day 24; 2-day activin, day 26; 4-day activin, day 28 of differentiation]. (C) RT-qPCR analyses showing expression of indicated genes in H9- and iPSC (38-2)-derived hepatic aggregates (day 44) cultured in the presence and absence of 8-Br-cAMP. Values are determined relative to TBP and presented as fold change relative to expression in non-treated cells, which is set at 1. (D) CYP1A2 and CYP3A4 activity in hepatic cells derived from H9 hESCs and 38-2 iPS. Activity is presented per 10,000 cells. Non-induced cells (-) were cultured in 0.1% DMSO-containing medium (n=3). Primary hepatocyte controls are the same as in Fig. 5E,F. Error bars in all graphs represent the s.d. of the mean of the values from three independent experiments, *P<0.05, **P<0.01, ***P<0.001, Student’s t-test.
Addition of 8-Br-cAMP did induce significant levels of expression of CYP3A4 (16-fold), CYP1A2 (100-fold) and CYP2B6 (10-fold), and the Phase II enzyme UGT1A1 (16-fold) in the H9-derived aggregates (Fig. 6D). 8-Br-cAMP also induced the expression of these enzymes in hiPSC-derived derived aggregates (Fig. 6D); however, in the case of CYP3A4 the differences were not significant. As observed with the HES2 line, the H9-derived cells possessed that lansoprazole-inducible CYP1A2 activity. H9 and iPSC-derived cells also showed CYP3A4 activity that was inducible with rifampicin. Inducible CYP1A2 activity was not detectable in the iPSC-derived cells, possibly reflecting suboptimal differentiation of this population.
Microarray analyses of cAMP stimulated hepatic populations
To further assess the developmental status of the H9-derived hepatic populations, we carried out a microarray analysis comparing the global expression profiles of the cAMP-induced and non-induced cells with that of cultured (48 hours) primary hepatocytes. A total of 23,038 filtered transcripts were used in the final analysis. A two-way unsupervised hierarchical cluster analysis revealed that the three groups appear as distinct populations (supplementary material Fig. S7). The three cAMP-induced populations were the most similar to one another, whereas the three primary hepatocyte populations showed the most divergent expression patterns. A FDR corrected ANOVA (q<0.05) identified 784 transcripts that showed the most statistically significant variability across all three sample groups. A hierarchically clustered visualization of these data identified clusters of highly expressed transcripts in each of the biological groups (Fig. 7A). These clusters consisted of 181 transcripts in the primary hepatocytes, (purple bar) 106 transcripts in the 8-Br-cAMP-induced cells (yellow bar) and 80 transcripts (blue bar) in the non-treated cells (supplementary material Table S2). Genes enriched in 8-Br-cAMP-induced cells included most of the key P450 enzymes, those involved in different aspects of liver function (including gluconeogenesis, glucose homeostasis and lipid metabolism) and those involved in mitochondria function, such as carnitine palmitoyltransferase 1A (CPT1A) and PTEN induced putative kinase 1 (PINK1) [supplementary material Table S2 (q<0.1), Table S11]. The cluster expressed at highest levels in the primary hepatocytes consisted of immune system, inflammatory-related and MHC genes (supplementary material Tables S2, S9, S10). The cluster detected in the non-induced hESC-derived cells did not contain any enriched gene ontology categories.
Fig. 7.
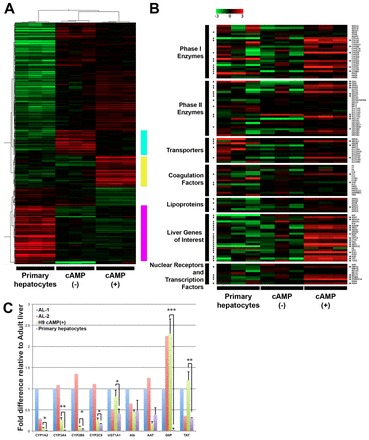
Microarray analyses comparing primary hepatocytes with non-treated and cAMP-treated hESC-derived hepatic populations. (A) Heat map summarizing expression of 784 transcripts that showed the most statistically significant variability across the sample groups. The bars on the right indicate clusters of transcripts highly expressed in each of the biological groups. (B) Heat map showing expression of selected transcripts for each of the indicated categories. Asterisks on the right indicate those genes that are expressed at significantly higher levels in the cAMP-treated hESC-derived cells compared with primary hepatocytes, whereas markings on the left indicate those that are significantly higher in the cAMP-treated hESC-derived cells compared with non-treated hESC population. (C) RT-qPCR analyses comparing expression levels of CYP1A2, CYP3A4, CYP2B6, CYP2C9, UGT1A1, ALB, a1-anti trypsin (AAT), G6P and TAT expression in day 44 H9-derived cAMP-treated aggregates (day 44) with those in adult liver and cultured primary hepatocytes. Values are determined relative to TBP and presented as fold change relative to expression in the adult liver sample 1 (AL1), which is set at 1. AL1: total human adult liver RNA. AL2: total human adult liver RNA. Error bars in all graphs represent the s.d. of the mean of the values from three independent experiments. *P<0.05, **P<0.01, ***P<0.001, Student’s t-test.
For a more detailed comparison of the populations, we next analyzed selected sets of transcripts that included a subset of Phase I and II drug metabolizing enzymes, transporters, coagulation factors, lipoproteins, nuclear receptors and transcription factors and general liver enzymes, and other functional molecules (Fig. 7B). The complete gene lists with fold differences in expression are provided in supplementary material Tables S2-S6. Analyses of these data revealed that many of the genes were expressed at comparable levels in the 8-Br-cAMP-treated hESC-derived cells and the primary hepatocytes. Select genes in each category were expressed at significantly higher levels in the 8-Br-cAMP-treated cells compared with the untreated cells or the primary hepatocytes. These include the Phase I enzymes CYP1A2 and CYP3A4; the Phase II enzyme SULT2A1; ASGR1, ALB and the transporter SLCO1B1; and the general liver enzymes TAT, G6P and TDO2.
RT-qPCR analyses showed that the levels of CYP1A2, CYP3A4, CYP2B6 and UGT1A1 were significantly higher in 8-Br-cAMP treated cells than in the primary cultured hepatocytes (Fig. 7C), confirming the findings from the microarray analyses. Comparison of the hESC-derived cells to the adult liver revealed levels of CYP1A2, CYP3A4, CYP2B6, CYP2C9 and UGT1A1 at 6.8±1.4%, 26.5±5.01%, 11±1.6%, 26.3±5.8% and 82.3±14.5%, respectively, of the levels detected in the intact tissue. Other genes, such as G6P and TAT, were expressed at similar levels to those found in the adult liver. The levels of expression in the cultured hepatocytes were substantially lower than the levels in the adult liver samples, reflecting the loss of metabolic activity of cultured hepatocytes. Taken together, these findings demonstrate that induction of hepatoblast-stage aggregates with cAMP results in global expression changes indicative of hepatocyte maturation.
DISCUSSION
Previous studies have shown that it is possible to generate immature hepatic lineage cells from both hESCs and hiPSCs using staged protocols designed to recapitulate crucial developmental steps in the embryo (Cai et al., 2007; Hay et al., 2008; Si-Tayeb et al., 2010b; Touboul et al., 2010; Funakoshi et al., 2011; Kajiwara et al., 2012). The success of these studies reflects the fact that the pathways controlling the early stages of differentiation are reasonably well defined. In this report, we extended the differentiation protocol to provide insights into pathways that regulate maturation of hESC-derived hepatocyte-like cells and demonstrate that the combination of 3D aggregation and cAMP signaling play a pivotal role at this stage of development. We also show that the duration of activin/nodal signaling following endoderm induction is crucial for the generation of an enriched progenitor population that can respond to cAMP. With these manipulations, it is possible to routinely generate hESC-derived populations that display measurable levels of Phase I and II metabolic enzymes and gene expression profiles indicative of hepatocyte maturation.
Our expression analyses showed that the cAMP-induced cells expressed higher levels of metabolic genes and other genes involved in hepatocyte function than found in cultured primary hepatocytes. Comparison with adult liver revealed that the hESC-derived cells had levels of the Phase I enzyme genes in the range of 7-27% and of the Phase II gene UGT1A1 at 82% of those found in the adult tissue. Several previous studies have reported the development of hPSC-derived hepatocytes that express some P450 enzyme activity (Duan et al., 2010; Hay et al., 2011; Takayama et al., 2011; Nagamoto et al., 2012; Takayama et al., 2012; Takayama et al., 2013). Duan et al. (Duan et al., 2010) were the first to successfully generate cells that had CYP1A2, CYP3A4, CYP2C9 and CYP2D6 enzyme activities comparable with those found in primary hepatocytes. These findings were encouraging as they demonstrated that it is possible to derive metabolically active cells from hESCs. This study did not, however, provide any insights into the pathways that promote maturation or show that the approach was applicable to different cell lines. More recent studies have shown that culture on specific polymers (Hay et al., 2011), culture as 3D aggregates (Sivertsson et al., 2013) or the combination of enforced expression of key transcription factors together with 3D aggregation (Takayama et al., 2013) promote the development hPSC-derived hepatocytes that express CYP genes. Of these, only two showed inducible CYP activity. Hay et al. reported general CYP3A activity, whereas Takayama and colleagues showed rifampicin-inducible CYP3A4 activity in iPSC-derived cells. Although the cells in the later study display Phase I and II gene expression profiles at levels similar to those in primary hepatocytes, this approach does have the drawback in that the development of the cells is dependent on viral transduction of different transcription factors.
The observation that sustained activin/nodal signaling within the CXCR4+CKIT+ population is crucial for the generation of mature hepatocytes highlights the importance of appropriate manipulation of early-stage cells for the efficient generation of mature cells. The effect of extended activin/nodal signaling between days 6 and 8 of differentiation (for HES2 cells) is striking, as it dramatically impacted gene expression patterns and the proportion of albumin-positive cells detected at day 26 of culture. Most importantly, this step promoted the development of a population of hepatic cells that, in response to cAMP, mature to give rise to metabolically functioning hepatocytes. This additional signaling step is not compensation for poor endoderm induction, as the day 6 EB target population consisted of greater than 95% CXCR4+CKIT+EPCAM+SOX17+ cells. Rather, it appears to reduce contaminating mesoderm-derivatives (CD90+ and CD31+ cells), possibly due to the inability of activin to promote their survival in the absence of BMP or FGF. The extended activin step may also play a role in endoderm patterning, as previous studies have shown that the duration of activin/nodal signaling does influence lineage specification from hESC-derived endoderm (Green et al., 2011; Nostro et al., 2011; Spence et al., 2011).
The maturation stage of our protocol involves two distinct, but interdependent, steps. The first is the generation of 3D aggregates. As shown in previous studies (Miki et al., 2011; Sivertsson et al., 2013) and in the work reported here, culture of hPSC-derived hepatic cells as 3D aggregates leads to the upregulation of expression of a wide range of genes involved in different aspects of liver function. Aggregation alone, however, does not appear to promote maturation of the population to the stage at which the cells have functional levels of enzyme activity. Development of such cells is dependent on additional maturation signals, one of which we have shown to be cAMP. Importantly, the 3D aggregation step does induce maturation to the stage at which the cells can respond to cAMP. The mechanism by which aggregation promotes this differentiation step is currently not known, but could be related to enhanced cellular interactions and the generation of polarized epithelial cells that mimic the morphology of the hepatocytes within the liver.
The second step of our maturation strategy is the activation of the cAMP pathway within the 3D aggregates that results in broad changes in gene expression indicative of maturation of the hepatic lineage. Notable among these changes was the upregulation of expression of two key CYP genes, CYP1A2 and CYP3A4, that are not expressed in the liver until after birth and function to metabolize many of the clinically relevant drugs (Hines and McCarver, 2002). These changes were indicative of function as the cAMP-treated cells displayed inducible CYP1A2 and CYP3A enzyme activity. cAMP and PGC1A have been shown to regulate gene expression patterns in the liver in vivo. For example, under conditions of fasting, cAMP levels are upregulated, resulting in the rapid induction of PGC1A, a co-factor for HNF4A (Iordanidou et al., 2005; Bell and Michalopoulos, 2006) that plays a crucial role in liver metabolism through the control of Phase I and Phase II drug-metabolizing enzyme activities, glucose metabolism and lipid production (Parviz et al., 2003; Rhee et al., 2003; Odom et al., 2004). Expression of PGC1A is also dramatically upregulated in mouse liver immediately after birth (Lin et al., 2003) possibly to promote maturation of the neonatal hepatocytes. Through the upregulation of PGC1A expression, the effects of cAMP signaling on the hPSC-derived hepatoblasts may be recapitulating the change observed in the liver during fasting and/or in hepatocyte lineage at birth, resulting the generation of cells that display many features of mature cells.
In summary, our findings have, for the first time, defined crucial steps that promote the maturation of hepatic lineage cells from hPSCs resulting the generation of cells that display functional properties of hepatocytes. The development of metabolically functional cells is an important end point that will enable the routine production of hPSC-derived hepatocyte-like cells for drug metabolism analyses in the pharmaceutical industry. The cAMP-induced cells also provide an ideal candidate population for the development of bio-artificial liver devices and ultimately for transplantation for cell replacement therapy for the treatment of liver disease. Both the drug metabolism and therapeutic applications will require scalable production that enables the routine generation of large numbers of these cells. Current efforts are aimed at optimizing expansion strategies at different stages of the protocol.
Supplementary Material
Acknowledgments
We thank members of the G.K. laboratory for discussion and critical reading of the manuscript; M. C. Nostro and F. Sarangi for generating the islet-like cells from hESCs; G. Daley (Harvard Medical School, Boston) for providing the human iPSC line (38-2); S. C. Strom for providing the RNA samples of isolated primary human hepatocytes; and R. Snodgrass for discussion and suggestions.
Footnotes
Funding
This work was supported by funding from VistaGen Therapeutics (San Francisco) and by a grant [SCN 12091] from the Canadian Stem Cell Network (Canada) to G.K., R.F.T., R.R.L. and D.M.G.
Competing interests statement
The authors declare no competing financial interests.
Author contributions
S.O. and G.K. designed the study and wrote the paper. S.O., J.S., M.O. and M.N. designed and carried out the experiments and analyzed the data. C.V. performed microarray experiment and analyzed the data. K.S.S., S.W., L.T., C.G., E.H., Z.B., R.R.L., R.F.T. and D.M.G. designed and performed the drug metabolism assay, and analyzed the data.
Supplementary material
Supplementary material available online at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.090266/-/DC1
References
- Arpiainen S., Järvenpää S. M., Manninen A., Viitala P., Lang M. A., Pelkonen O., Hakkola J. (2008). Coactivator PGC-1alpha regulates the fasting inducible xenobiotic-metabolizing enzyme CYP2A5 in mouse primary hepatocytes. Toxicol. Appl. Pharmacol. 232, 135–141 [DOI] [PubMed] [Google Scholar]
- Basma H., Soto-Gutiérrez A., Yannam G. R., Liu L., Ito R., Yamamoto T., Ellis E., Carson S. D., Sato S., Chen Y., et al. (2009). Differentiation and transplantation of human embryonic stem cell-derived hepatocytes. Gastroenterology 136, 990–999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell A. W., Michalopoulos G. K. (2006). Phenobarbital regulates nuclear expression of HNF-4alpha in mouse and rat hepatocytes independent of CAR and PXR. Hepatology 44, 186–194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benet M., Lahoz A., Guzmán C., Castell J. V., Jover R. (2010). CCAAT/enhancer-binding protein alpha (C/EBPalpha) and hepatocyte nuclear factor 4alpha (HNF4alpha) synergistically cooperate with constitutive androstane receptor to transactivate the human cytochrome P450 2B6 (CYP2B6) gene: application to the development of a metabolically competent human hepatic cell model. J. Biol. Chem. 285, 28457–28471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benjamini Y., Drai D., Elmer G., Kafkafi N., Golani I. (2001). Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 125, 279–284 [DOI] [PubMed] [Google Scholar]
- Byers J., Bachmann K., Eng H., Katta A., White D., Ghosh R., Hewitt N. J., Silber P., Chen G. (2007). An estimate of the number of hepatocyte donors required to provide reasonable estimates of human hepatic clearance from in vitro experiments. Drug Metab. Lett. 1, 91–95 [DOI] [PubMed] [Google Scholar]
- Cai J., Zhao Y., Liu Y., Ye F., Song Z., Qin H., Meng S., Chen Y., Zhou R., Song X., et al. (2007). Directed differentiation of human embryonic stem cells into functional hepatic cells. Hepatology 45, 1229–1239 [DOI] [PubMed] [Google Scholar]
- Chen Y. F., Tseng C. Y., Wang H. W., Kuo H. C., Yang V. W., Lee O. K. (2012). Rapid generation of mature hepatocyte-like cells from human induced pluripotent stem cells by an efficient three-step protocol. Hepatology 55, 1193–1203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dankel S. N., Hoang T., Flågeng M. H., Sagen J. V., Mellgren G. (2010). cAMP-mediated regulation of HNF-4alpha depends on the level of coactivator PGC-1alpha. Biochim. Biophys. Acta 1803, 1013–1019 [DOI] [PubMed] [Google Scholar]
- Duan Y., Catana A., Meng Y., Yamamoto N., He S., Gupta S., Gambhir S. S., Zern M. A. (2007). Differentiation and enrichment of hepatocyte-like cells from human embryonic stem cells in vitro and in vivo. Stem Cells 25, 3058–3068 [DOI] [PubMed] [Google Scholar]
- Duan Y., Ma X., Zou W., Wang C., Bahbahan I. S., Ahuja T. P., Tolstikov V., Zern M. A. (2010). Differentiation and characterization of metabolically functioning hepatocytes from human embryonic stem cells. Stem Cells 28, 674–686 [DOI] [PubMed] [Google Scholar]
- Funakoshi N., Duret C., Pascussi J. M., Blanc P., Maurel P., Daujat-Chavanieu M., Gerbal-Chaloin S. (2011). Comparison of hepatic-like cell production from human embryonic stem cells and adult liver progenitor cells: CAR transduction activates a battery of detoxification genes. Stem Cell Rev. 7, 518–531 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gadue P., Huber T. L., Paddison P. J., Keller G. M. (2006). Wnt and TGF-beta signaling are required for the induction of an in vitro model of primitive streak formation using embryonic stem cells. Proc. Natl. Acad. Sci. USA 103, 16806–16811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gagné J. F., Montminy V., Belanger P., Journault K., Gaucher G., Guillemette C. (2002). Common human UGT1A polymorphisms and the altered metabolism of irinotecan active metabolite 7-ethyl-10-hydroxycamptothecin (SN-38). Mol. Pharmacol. 62, 608–617 [DOI] [PubMed] [Google Scholar]
- Gebhardt R., Hengstler J. G., Müller D., Glöckner R., Buenning P., Laube B., Schmelzer E., Ullrich M., Utesch D., Hewitt N., et al. (2003). New hepatocyte in vitro systems for drug metabolism: metabolic capacity and recommendations for application in basic research and drug development, standard operation procedures. Drug Metab. Rev. 35, 145–213 [DOI] [PubMed] [Google Scholar]
- Gouon-Evans V., Boussemart L., Gadue P., Nierhoff D., Koehler C. I., Kubo A., Shafritz D. A., Keller G. (2006). BMP-4 is required for hepatic specification of mouse embryonic stem cell-derived definitive endoderm. Nat. Biotechnol. 24, 1402–1411 [DOI] [PubMed] [Google Scholar]
- Grant D. M., Blum M., Beer M., Meyer U. A. (1991). Monomorphic and polymorphic human arylamine N-acetyltransferases: a comparison of liver isozymes and expressed products of two cloned genes. Mol. Pharmacol. 39, 184–191 [PubMed] [Google Scholar]
- Green M. D., Chen A., Nostro M. C., d’Souza S. L., Schaniel C., Lemischka I. R., Gouon-Evans V., Keller G., Snoeck H. W. (2011). Generation of anterior foregut endoderm from human embryonic and induced pluripotent stem cells. Nat. Biotechnol. 29, 267–272 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillouzo A. (1998). Liver cell models in in vitro toxicology. Environ. Health Perspect. 106 Suppl. 2, 511–532 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay D. C., Fletcher J., Payne C., Terrace J. D., Gallagher R. C., Snoeys J., Black J. R., Wojtacha D., Samuel K., Hannoun Z., et al. (2008). Highly efficient differentiation of hESCs to functional hepatic endoderm requires ActivinA and Wnt3a signaling. Proc. Natl. Acad. Sci. USA 105, 12301–12306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hay D. C., Pernagallo S., Diaz-Mochon J. J., Medine C. N., Greenhough S., Hannoun Z., Schrader J., Black J. R., Fletcher J., Dalgetty D., et al. (2011). Unbiased screening of polymer libraries to define novel substrates for functional hepatocytes with inducible drug metabolism. Stem Cell Res. 6, 92–102 [DOI] [PubMed] [Google Scholar]
- Hewitt N. J., Lechón M. J., Houston J. B., Hallifax D., Brown H. S., Maurel P., Kenna J. G., Gustavsson L., Lohmann C., Skonberg C., et al. (2007). Primary hepatocytes: current understanding of the regulation of metabolic enzymes and transporter proteins, and pharmaceutical practice for the use of hepatocytes in metabolism, enzyme induction, transporter, clearance, and hepatotoxicity studies. Drug Metab. Rev. 39, 159–234 [DOI] [PubMed] [Google Scholar]
- Hines R. N., McCarver D. G. (2002). The ontogeny of human drug-metabolizing enzymes: phase I oxidative enzymes. J. Pharmacol. Exp. Ther. 300, 355–360 [DOI] [PubMed] [Google Scholar]
- Iordanidou P., Aggelidou E., Demetriades C., Hadzopoulou-Cladaras M. (2005). Distinct amino acid residues may be involved in coactivator and ligand interactions in hepatocyte nuclear factor-4alpha. J. Biol. Chem. 280, 21810–21819 [DOI] [PubMed] [Google Scholar]
- Kajiwara M., Aoi T., Okita K., Takahashi R., Inoue H., Takayama N., Endo H., Eto K., Toguchida J., Uemoto S., et al. (2012). Donor-dependent variations in hepatic differentiation from human-induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 109, 12538–12543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy M., D’Souza S. L., Lynch-Kattman M., Schwantz S., Keller G. (2007). Development of the hemangioblast defines the onset of hematopoiesis in human ES cell differentiation cultures. Blood 109, 2679–2687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klipper-Aurbach Y., Wasserman M., Braunspiegel-Weintrob N., Borstein D., Peleg S., Assa S., Karp M., Benjamini Y., Hochberg Y., Laron Z. (1995). Mathematical formulae for the prediction of the residual beta cell function during the first two years of disease in children and adolescents with insulin-dependent diabetes mellitus. Med. Hypotheses 45, 486–490 [DOI] [PubMed] [Google Scholar]
- Kostrubsky V. E., Ramachandran V., Venkataramanan R., Dorko K., Esplen J. E., Zhang S., Sinclair J. F., Wrighton S. A., Strom S. C. (1999). The use of human hepatocyte cultures to study the induction of cytochrome P-450. Drug Metab. Dispos. 27, 887–894 [PubMed] [Google Scholar]
- Lin J., Tarr P. T., Yang R., Rhee J., Puigserver P., Newgard C. B., Spiegelman B. M. (2003). PGC-1beta in the regulation of hepatic glucose and energy metabolism. J. Biol. Chem. 278, 30843–30848 [DOI] [PubMed] [Google Scholar]
- Loboz K. K., Gross A. S., Ray J., McLachlan A. J. (2005). HPLC assay for bupropion and its major metabolites in human plasma. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 823, 115–121 [DOI] [PubMed] [Google Scholar]
- Miki T., Ring A., Gerlach J. (2011). Hepatic differentiation of human embryonic stem cells is promoted by three-dimensional dynamic perfusion culture conditions. Tissue Eng. Part C Methods 17, 557–568 [DOI] [PubMed] [Google Scholar]
- Nagamoto Y., Tashiro K., Takayama K., Ohashi K., Kawabata K., Sakurai F., Tachibana M., Hayakawa T., Furue M. K., Mizuguchi H. (2012). The promotion of hepatic maturation of human pluripotent stem cells in 3D co-culture using type I collagen and Swiss 3T3 cell sheets. Biomaterials 33, 4526–4534 [DOI] [PubMed] [Google Scholar]
- Nostro M. C., Sarangi F., Ogawa S., Holtzinger A., Corneo B., Li X., Micallef S. J., Park I. H., Basford C., Wheeler M. B., et al. (2011). Stage-specific signaling through TGFβ family members and WNT regulates patterning and pancreatic specification of human pluripotent stem cells. Development 138, 861–871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Odom D. T., Zizlsperger N., Gordon D. B., Bell G. W., Rinaldi N. J., Murray H. L., Volkert T. L., Schreiber J., Rolfe P. A., Gifford D. K., et al. (2004). Control of pancreas and liver gene expression by HNF transcription factors. Science 303, 1378–1381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parviz F., Matullo C., Garrison W. D., Savatski L., Adamson J. W., Ning G., Kaestner K. H., Rossi J. M., Zaret K. S., Duncan S. A. (2003). Hepatocyte nuclear factor 4alpha controls the development of a hepatic epithelium and liver morphogenesis. Nat. Genet. 34, 292–296 [DOI] [PubMed] [Google Scholar]
- Rhee J., Inoue Y., Yoon J. C., Puigserver P., Fan M., Gonzalez F. J., Spiegelman B. M. (2003). Regulation of hepatic fasting response by PPARgamma coactivator-1alpha (PGC-1): requirement for hepatocyte nuclear factor 4alpha in gluconeogenesis. Proc. Natl. Acad. Sci. USA 100, 4012–4017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roymans D., Van Looveren C., Leone A., Parker J. B., McMillian M., Johnson M. D., Koganti A., Gilissen R., Silber P., Mannens G., et al. (2004). Determination of cytochrome P450 1A2 and cytochrome P450 3A4 induction in cryopreserved human hepatocytes. Biochem. Pharmacol. 67, 427–437 [DOI] [PubMed] [Google Scholar]
- Si-Tayeb K., Lemaigre F. P., Duncan S. A. (2010a). Organogenesis and development of the liver. Dev. Cell 18, 175–189 [DOI] [PubMed] [Google Scholar]
- Si-Tayeb K., Noto F. K., Nagaoka M., Li J., Battle M. A., Duris C., North P. E., Dalton S., Duncan S. A. (2010b). Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 51, 297–305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sivertsson L., Synnergren J., Jensen J., Björquist P., Ingelman-Sundberg M. (2013). Hepatic differentiation and maturation of human embryonic stem cells cultured in a perfused three-dimensional bioreactor. Stem Cells Dev. 22, 581–594 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spence J. R., Mayhew C. N., Rankin S. A., Kuhar M. F., Vallance J. E., Tolle K., Hoskins E. E., Kalinichenko V. V., Wells S. I., Zorn A. M., et al. (2011). Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro. Nature 470, 105–109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stieger B., Heger M., de Graaf W., Paumgartner G., van Gulik T. (2012). The emerging role of transport systems in liver function tests. Eur. J. Pharmacol. 675, 1–5 [DOI] [PubMed] [Google Scholar]
- Sullivan G. J., Hay D. C., Park I. H., Fletcher J., Hannoun Z., Payne C. M., Dalgetty D., Black J. R., Ross J. A., Samuel K., et al. (2010). Generation of functional human hepatic endoderm from human induced pluripotent stem cells. Hepatology 51, 329–335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takayama K., Inamura M., Kawabata K., Tashiro K., Katayama K., Sakurai F., Hayakawa T., Furue M. K., Mizuguchi H. (2011). Efficient and directive generation of two distinct endoderm lineages from human ESCs and iPSCs by differentiation stage-specific SOX17 transduction. PLoS ONE 6, e21780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takayama K., Inamura M., Kawabata K., Katayama K., Higuchi M., Tashiro K., Nonaka A., Sakurai F., Hayakawa T., Furue M. K., et al. (2012). Efficient generation of functional hepatocytes from human embryonic stem cells and induced pluripotent stem cells by HNF4alpha transduction. Mol. Ther. 20, 127–137 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takayama K., Kawabata K., Nagamoto Y., Kishimoto K., Tashiro K., Sakurai F., Tachibana M., Kanda K., Hayakawa T., Furue M. K., et al. (2013). 3D spheroid culture of hESC/hiPSC-derived hepatocyte-like cells for drug toxicity testing. Biomaterials 34, 1781–1789 [DOI] [PubMed] [Google Scholar]
- Touboul T., Hannan N. R., Corbineau S., Martinez A., Martinet C., Branchereau S., Mainot S., Strick-Marchand H., Pedersen R., Di Santo J., et al. (2010). Generation of functional hepatocytes from human embryonic stem cells under chemically defined conditions that recapitulate liver development. Hepatology 51, 1754–1765 [DOI] [PubMed] [Google Scholar]
- Zhang F., Chen J. Y. (2011). HOMER: a human organ-specific molecular electronic repository. BMC Bioinformatics 12 Suppl. 10, S4 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.