

Abstract
Wnt signalling is a key regulator of vertebrate heart development, yet it is unclear which specific Wnt signalling components are required to regulate which aspect of cardiogenesis. Previously, we identified Wnt6 as an endogenous Wnt ligand required for controlling heart muscle differentiation via canonical Wnt/β-catenin signalling. Here we show for the first time a requirement for an endogenous Wnt signalling inhibitor for normal heart muscle differentiation. Expression of sfrp1 is strongly induced in differentiating heart muscle. We show that sfrp1 is not only able to promote heart muscle differentiation but is also required for the formation of normal size heart muscle in the embryo. sfrp1 is functionally able to inhibit Wnt6 signalling and its requirement during heart development relates to relieving the cardiogenesis-restricting function of endogenous wnt6. In turn, we discover that sfrp1 expression in the heart is regulated by Wnt6 signalling, which for the first time indicates that sfrp genes can function as part of a Wnt negative-feedback regulatory loop. Our experiments indicate that sfrp1 controls the size of the differentiating heart muscle primarily by regulating cell fate within the cardiac mesoderm between muscular and non-muscular cell lineages. The cardiac mesoderm is therefore not passively patterned by signals from the surrounding tissue, but regulates its differentiation into muscular and non-muscular tissue using positional information from the surrounding tissue. This regulatory network might ensure that Wnt activation enables expansion and migration of cardiac progenitors, followed by Wnt inhibition permitting cardiomyocyte differentiation.
Keywords: Xenopus, Wnt signalling, Heart development
INTRODUCTION
During vertebrate embryonic development, heart progenitors acquire the potential to subsequently differentiate as cardiomyocytes in the myocardium and form the functional heart muscle (Mohun et al., 2000; Mohun et al., 2003; Nakajima et al., 2009). Identifying how these cardiac progenitors are controlled and the source of signals regulating differentiation is of fundamental importance for our understanding of embryonic heart development. Moreover, injury to the adult heart, for instance after myocardial infarction, can cause serious damage to mature cardiomyocytes. Even though cardiomyocytes have some potential to proliferate and there is encouraging evidence for resident cardiac progenitors in the adult heart, this is normally insufficient to repair the damage. Understanding how to promote cardiomyocyte differentiation from endogenous progenitor cells in order to replace damaged cardiomyocytes and regenerate damaged heart muscle would be an exciting prospect and impact greatly on clinical treatments of heart patients (reviewed by Mummery et al., 2010).
The Wnt signalling pathway is a key regulator of heart development and particularly of cardiomyocyte differentiation (reviewed by Gessert and Kühl, 2010). Experimental manipulation of Wnt ligands and extracellular Wnt inhibitors was shown to influence heart development (e.g. Marvin, 2001; Schneider and Mercola, 2001). Furthermore, the endogenous expression of some of these factors in, or close to, embryonic heart tissue further supported their possible role in this process (e.g. Monaghan et al., 1999). However, evidence of the requirement for specific endogenous Wnt signalling component genes in early embryonic heart development is slow to emerge. We set out to identify the important endogenous Wnt signalling components required for vertebrate heart development and to uncover the Wnt-regulated molecular gene regulatory mechanisms promoting myocardium development and cardiomyocyte differentiation. This will prove paramount for understanding heart development in the embryo but also for developing future regenerative therapies for heart patients. However, Wnt signalling in heart development appears complicated. So-called canonical β-catenin-dependent Wnt signalling has apparently opposing effects at different developmental stages on subsequent cardiomyocyte differentiation (reviewed by Tzahor, 2007), which may be linked to a positive effect on progenitor expansion and a brake on differentiation (reviewed by Bergmann, 2010). Additionally, non-canonical (i.e. β-catenin-independent) Wnt signalling mechanisms also regulate heart development (e.g. Eisenberg and Eisenberg, 1999; Pandur et al., 2002; Afouda et al., 2008; Cohen et al., 2012; Onizuka et al., 2012).
The Xenopus model system has been instrumental in uncovering fundamental molecular mechanisms and conserved functions in vertebrate heart development (reviewed by Warkman and Krieg, 2007), particularly the role of Wnt signalling during the early specification of heart development (e.g. Marvin, 2001; Schneider and Mercola, 2001). We identified in Wnt6 the first endogenous Wnt ligand required for regulating vertebrate heart development via the canonical Wnt pathway, but surprisingly discovered a function at later stages of development, during organogenesis preceding cardiomyocyte differentiation (Lavery et al., 2008b). The identification of a specific Wnt ligand that is expressed close to, but outside of, the cardiac mesoderm (Lavery et al., 2008a) allowed us to consider a spatial model of how heart muscle differentiation is regulated in the embryo. The cardiogenic mesoderm is further patterned into muscular (i.e. myocardium) and non-muscular (e.g. pericardium) tissues (e.g. Raffin et al., 2000). We wondered whether a simple Wnt gradient would be sufficient to pattern the cardiac mesoderm tissue or whether additional mechanisms were at play.
sfrp1 is a member of the secreted frizzled-related protein (sFRP) family of potential Wnt inhibitors [originally referred to as FrzA (Duplàa et al., 1999)], which was found to be expressed in the myocardium of the developing embryo (Xu et al., 1998). sFRP proteins share structural domains with members of the frizzled family of Wnt receptors, which is consistent with their initially proposed role as extracellular Wnt inhibitors (reviewed by Jones and Jomary, 2002).
Here we test the hypothesis that sfrp1, as an endogenous Wnt inhibitor, interacts with wnt6 in the embryonic Xenopus heart to promote myocardial development. We confirm sfrp1 expression in the myocardium, that endogenous sfrp1 is required for normal heart muscle differentiation and that experimental overexpression of sfrp1 is sufficient to expand heart muscle development. We also demonstrate that sfrp1 is capable of inhibiting Wnt6 activity and that sfrp1 function in the heart is primarily required for regulating Wnt6 function. The regulation of sfrp1 expression suggests that it functions as part of an extracellular feedback regulatory loop. Since sfrp1 expression is conserved in vertebrate heart development, our findings are of relevance for mammalian heart development and regeneration.
MATERIALS AND METHODS
Embryo manipulations
Xenopus laevis (Daudin) embryos were collected according to Hoppler (Hoppler, 2008) and staged according to Nieuwkoop and Faber (Nieuwkoop and Faber, 1967).
RNA and morpholino injections
Capped RNA and morpholino antisense oligonucleotides (MOs) were injected as previously described (Lavery and Hoppler, 2008b; Lavery et al., 2008b) (see supplementary material Table S1 for plasmid templates for RNA synthesis). MOs were designed to target Xenopus laevis transcripts and synthesised by Gene Tools: sfrp1 MOs, see Fig. 2; gata4 and gata6 MOs (Peterkin et al., 2007); wnt6MO3 (Lavery et al., 2008b). Either 80 ng (CoMO and sfrp1MO), 40 ng (wnt6MO3), 20 ng (gata4MO) or 10 ng (gata6MO) was injected per embryo at stage 3 of development into the marginal zone of both dorsal blastomeres. All results are from at least five independent experiments (i.e. with independent batches of embryos), except where indicated (see Figs 6, 9).
Fig. 2.
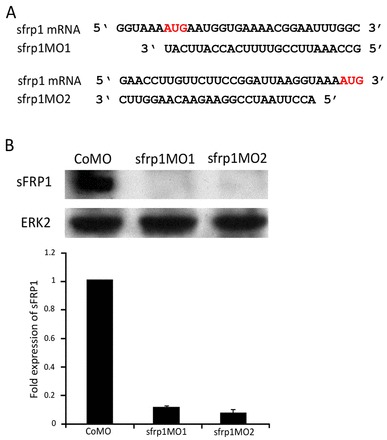
sfrp1 protein expression is knocked down by non-overlapping antisense morpholino oligonucleotides. (A) Non-overlapping sfrp1 morpholino oligonucleotide (MO) sequences with the AUG translation initiation codon highlighted. (B) Western blot and analysis of sfrp1 protein expression levels in embryos injected with control MO (CoMO), sfrp1MO1 and sfrp1MO2 at the 4-cell stage into all four blastomeres. Protein extraction was carried out at stage 32 of development. Representative blots and mean ± s.e.m. of data are shown.
Fig. 6.
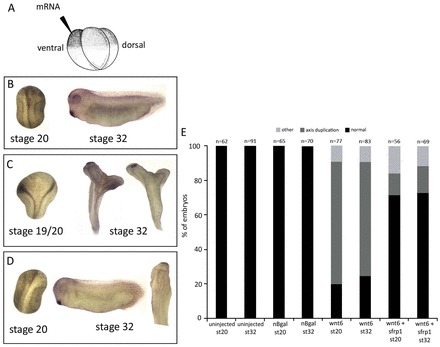
sfrp1 inhibits wnt6 in vivo. (A) In the sfrp1 and wnt6 functional interaction assay, mRNA was injected into one ventral blastomere of 4-cell embryos. Embryos were left to develop until stages 20 or 32 for analysis. (B) Control embryo injected with nβ-gal-encoding mRNA showing normal axis development (100%). (C) Injection of nβ-gal-encoding mRNA and wnt6 mRNA into the same ventral blastomere causes secondary axis formation in over 70% of injected embryos. (D) Embryos injected with sfrp1 mRNA followed 30 minutes later by wnt6 mRNA into the same ventral blastomere show rescue of wnt6-induced secondary axis formation, being reduced to less than 20%. (E) The number of embryos injected and their phenotype. The results presented here are from at least three independent experiments (i.e. with independent batches of embryos).
Fig. 9.
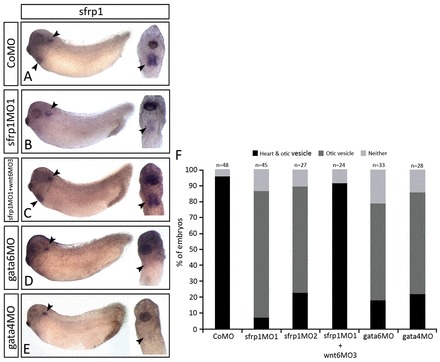
sfrp1 expression is regulated by cardiogenic transcription factors. (A-E) WISH analysis on stage 32 embryos highlighting the sfrp1 gene expression pattern (arrowheads) after injection with either CoMO (A), sfrp1MO1 (B), sfrp1MO1 and wnt6MO3 combined (C), gata6MO (D) or gata4MO (E) into both dorsal blastomeres at the 4-cell stage of development. Note the loss of sfrp1 expression in the myocardium in B,D,E and the recovery of lost myocardial sfrp1 expression in C. (F) The number of embryos injected and their corresponding sfrp1 expression pattern. The results presented here are from at least three independent experiments (i.e. with independent batches of embryos).
Heat shock-inducible gene expression in transgenics
Transgenic embryos were created and transgene expression induced by heat treatment at stage 22 with three 15-minute treatments at 37°C following 30-minute intervals at 16°C (Amaya and Kroll, 1999; Wheeler et al., 2000; Lavery et al., 2008b; Wheeler et al., 2008).
Whole-mount RNA in situ hybridisation
Digoxigenin (DIG)-labelled antisense RNA probes to detect Xenopus laevis mRNA were synthesised from linearised template plasmids (supplementary material Table S2) using the High Yield Megascript Kit (Ambion).
Whole-mount RNA in situ hybridisation was performed with a standard protocol (e.g. Lavery and Hoppler, 2008a; Thompson et al., 2009). Embryos were imaged using a Leica MZ6 stereomicroscope at 1.6× magnification for whole embryos and 3.2× magnification for the heart region with an RS Photometrics CoolSNAP digital camera with Improvision Openlab and Adobe Photoshop software on a Macintosh computer.
Cryostat sections of whole-mount RNA in situ embryos were obtained at 14 μm, mounted in 90% glycerol and imaged using a Nikon SMZ1500 stereomicroscope with an attached Nikon DS-Fi1c camera at 10× magnification.
Cryosectioning and immunofluorescence staining
Sections were cut at 14 μm using a Bright Clinical Cryostat. Cardiac Troponin T (cTnT/tnnt2) immunohistochemistry analysis was as described (Lavery et al., 2008b). Sections were imaged the following day using a Zeiss Axioplan 2 fluorescence microscope.
Quantitative analysis of immunofluorescence sections was performed using ImageJ (NIH). For each experimental sample the fluorescence intensity of cTnT (red) was measured, giving a percentage area of cTnT expression. An average was taken between all sections from different embryos and graphed as shown. P-values were obtained using a paired Student’s t-test.
Protein analysis
Protein was extracted from five embryos per experimental condition and western blotting performed using standard procedures with minor modifications. Primary antibodies were polyclonal rabbit anti-sFRP1 (Abcam, ab4193) diluted 1:230, and polyclonal rabbit anti-ERK2 (C-14) (Santa Cruz Biotechnology, sc-154) diluted 1:1000. Primary antibodies were left overnight at 4°C rolling, followed by polyclonal swine anti-rabbit immunoglobulins/HRP (DAKO, P0217) secondary antibody diluted 1:1000.
Immunoreactive bands were quantified by densitometry using Multi-Analyst software 1.1 (Bio-Rad). The housekeeping protein Ornithine decarboxylase (ODC) was used to normalise expression levels.
Quantitative RT-PCR analysis of cardiac gene expression
Total RNA was extracted using the RNeasy Mini Kit (Qiagen) from 15 embryos per experimental condition. cDNA synthesis was carried out using the QuantiTect Reverse Transcription Kit (Qiagen), using 2 μg RNA of each experimental condition per 40 μl cDNA synthesis reaction. Quantitative (q) PCR was performed in triplicate using the LightCycler 480 SYBR Green I Master Mix (Roche) (see supplementary material Table S3 for primers). Relative quantification was calculated using the standard curve method after normalising to expression levels of ornithine decarboxylase (odc1).
Modelling
Gene regulatory network (GRN) models were designed in BioTapestry Editor (version 4.1.0, http://www.biotapestry.org/), the mathematical model was exported as systems biology mark-up language code (http://sbml.org) and imported into the Dizzy simulator (version 6.1, http://magnet.systemsbiology.net/software/Dizzy) for simulation with the Gillespie stochastic algorithm. A serial arrangement of cardiogenic transcription factor (cTF) and inhibitor (Inh) genes in different responding cells was designed in a flat model because the current version of BioTapestry only supports SBML export for top-level flat models. The default parameter settings were used for modelling the cTF genes (Bolouri and Davidson, 2003). Other parameters were adjusted as described in the text, figure legend and supplementary material. The exported SBML models were simulated within Dizzy.
RESULTS
Overexpression of Wnt inhibitors such as Dickkopf and Crescent revealed an important role for Wnt signalling in regulating early vertebrate heart development (Marvin et al., 2001; Schneider and Mercola, 2001; Yang et al., 2008). While in pursuit of endogenous Wnt signals that mediate this regulation of early vertebrate heart development, we identified wnt6 (Lavery et al., 2008a), but discovered unexpectedly a function in restricting heart muscle development at the later stages of organogenesis (Lavery et al., 2008b). We adopted a simple modelling approach (see below), which supported our intuition that a simple Wnt gradient was likely to be insufficient to bring about clear patterning of the cardiogenic mesoderm into heart muscle tissue (i.e. myocardium) and non-muscular tissue (e.g. pericardium), without additional reinforcing cooperativity or negative-feedback mechanisms (supplementary material Models 1, 7, 8). We therefore wondered whether Wnt6 signalling during organogenesis stages was regulated by potential Wnt inhibitors and encountered sfrp1 (also known as FrzA) expressed in the differentiating heart (Xu et al., 1998). We initially studied the expression of sfrp1 (Fig. 1), and confirmed expression in the myocardium during organogenesis stages (Fig. 1F-L) but also earlier in the common cardiac precursors immediately posterior to the cement gland [compare nkx2-5 and gata6b in this tissue (Fig. 1M,N) with sfrp1 expression at the same stage (Fig. 1E)]. This expression pattern supported the hypothesis that endogenous sfrp1 might function to promote myocardium development by inhibiting Wnt6 signalling.
Fig. 1.
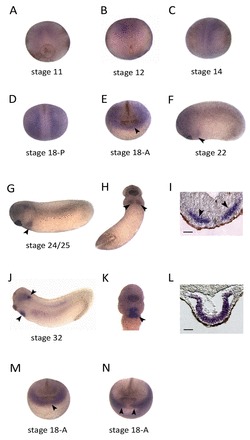
sfrp1 expression in cardiac precursors and differentiating myocardium. (A-H,J,K) Whole-mount RNA in situ hybridisation (WISH) analysis of sfrp1 on Xenopus embryos at the indicated stages. (I,L) WISH analysis on sections of stage 24 (I) and stage 32 (L) embryos, showing sfrp1 expression in cardiac progenitors (arrowheads in I) and in the differentiated myocardium (L). (M,N) WISH analysis on stage 18 embryos showing nkx2-5 (M) and gata6b (N) expression in the common cardiac progenitor population (arrowheads), confirming sfrp1 expression in this region (arrowhead in E). Scale bars: 100 μm in I,L.
Endogenous sfrp1 function is required for normal myocardium development
In order to study the requirement of endogenous sfrp1 in embryonic development we developed morpholino antisense oligonucleotides (MOs) to specifically interfere with translation initiation of sfrp1 protein synthesis. We designed two independent MOs targeting non-overlapping sequences close to the translation initiation site of the sfrp1 mRNA (Fig. 2A). We tested the efficacy of these MOs initially in in vitro transcription and translation reactions (data not shown), and confirmed the efficiency by western blot analysis, which showed a loss of endogenous sfrp1 protein expression in both sfrp1MO1- and sfrp1MO2-injected embryos (Fig. 2B). sfrp1MO1 and sfrp1MO2 result in indistinguishable phenotypes (Fig. 3J), suggesting negligible, if any, off-target effects. All loss-of-function analyses were subsequently carried out in parallel with sfrp1MO1 and sfrp1MO2, but, as indicated in the figure legends, we mostly illustrate our results with examples using sfrp1MO1.
Fig. 3.
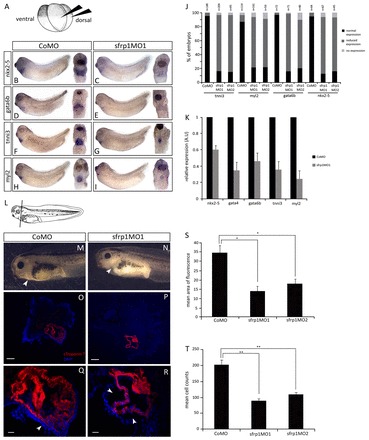
Loss-of-function experiments show that endogenous sfrp1 is required for formation of a normal size heart. (A) For sfrp1 loss-of-function experiments, MOs were injected into the marginal zone of both dorsal blastomeres at the 4-cell stage. (B-I) WISH analysis on stage 32 embryos indicating the expression of cardiac-specific marker genes. Control embryos injected with CoMO showed normal expression of cardiogenic transcription factor (B,D) and myocardial differentiation marker (F,H) genes. By contrast, embryos injected with sfrp1MO1 (C,E,G,I) revealed a loss of cardiogenic transcription factor gene expression and a reduced expression domain of myocardial differentiation marker genes. (J) Quantification of WISH of heart marker gene expression showing reduced expression in sfrp1MO1- and sfrp1MO2-injected compared with CoMO-injected embryos. (K) qPCR analysis on MO-injected embryos at stage 32 showing reduced cardiogenic gene expression in sfrp1MO1-injected embryos relative to controls. (L) Schematic of a stage 41 embryo indicating the orientation of sections in O-R. (M,N) Phenotype of stage 41 MO-injected embryos after terminal differentiation. CoMO-injected embryos (M) show a normal size heart within the pericardial cavity (arrowhead). sfrp1MO1-injected embryos (N) show reduced heart size (arrowhead) within a relatively empty pericardial cavity. (O-R) Terminal differentiation of the heart was analysed in stage 41 embryos by immunofluorescence for cardiac Troponin T (cTnT, red) with DAPI counterstaining (blue). Whereas control embryos show normal cTnT expression (O,Q), sfrp1MO1-injected embryos show reduced cTnT expression indicating loss of myocardial tissue. Arrowheads highlight the largely empty pericardial cavity due to loss of heart tissue (compare Q with R). (S) Analysis of cTnT immunofluorescence illustrating significant loss of myocardial tissue in sfrp1MO1-injected compared with CoMO-injected embryos. Measurements were taken from the middle section through each heart at stage 41, with three to four different embryos analysed per experimental condition (i.e. CoMO- and sfrp1MO1-injected embryos). (n=4, *P<0.05). (T) Mean cell counts in CoMO-injected embryo sections compared with sfrp1MO1 and sfrp1MO2 morphants. Although the size of individual cardiomyocytes appears unchanged, a significant reduction in cell number was observed in the differentiated myocardium of embryos injected with sfrp1MO (n=3, **P<0.01). See supplementary material Fig. S3 for heart marker tissue specificity. Data are mean ± s.e.m. Scale bars: 50 μm in O,P (10×); 200 μm in Q,R (40×).
In order to study the requirement of endogenous sfrp1 for normal vertebrate heart development we injected early Xenopus embryos with sfrp1 MOs to target subsequent heart-forming tissues (Fig. 3A). We did not observe any phenotype during early stages (supplementary material Fig. S2), but there was a visible reduction of heart tissue as the tadpole embryos developed, with an almost empty pericardial cavity and almost no beating heart tissue (compare Fig. 3N with 3M; supplementary material Movie 1). We examined whether this phenotype was a result of changes to embryonic heart development at organogenesis stages by monitoring the expression of different heart development-associated molecular markers (Fig. 3; supplementary material Figs S4, S5). Heart development-promoting cardiogenic transcription factor genes (gata6b and nkx2-5) and early myocardium-specific differentiation markers [cardiac Troponin I (tnni3) and Myosin light chain 2 (myl2)] were analysed during late organogenesis stages (typically stage 32) by RNA in situ hybridisation and qRT-PCR, whereas the myocardium differentiation marker cardiac Troponin T (tnnt2) was analysed after heart muscle differentiation (stage 41) by immunohistochemistry.
Interestingly, the expression of myocardium differentiation markers is confined to a smaller population of cells when endogenous sfrp1 is knocked down; however, in those remaining cells we detect what appears to be strong expression (Fig. 3G,I; supplementary material Fig. S4). This suggests that sfrp1 is not regulating expression levels as such, but rather the size of the tissue expressing heart muscle differentiation genes, consistent with the reduced size of the subsequently differentiating heart muscle in sfrp1 knockdown embryos (Fig. 3N,P,R). In most of our experimental samples, we detect a much reduced heart muscle in a mostly empty pericardial cavity (Fig. 3N). Analysis of images also reveals that this phenotype is not due to a reduced size of individual cardiomyocytes but rather to reduced numbers of normally proportioned cardiomyocytes in a less populous myocardium (Fig. 3T). Again, within this much reduced heart muscle, the expression of the differentiation marker was strong (Fig. 3P,R). Expression domains and levels of cardiogenic transcription factors were, however, reduced by sfrp1 knockdown (Fig. 3B-E).
sfrp1 can promote myocardium development
In order to study the functional potential of sfrp1 in vertebrate heart development we performed overexpression experiments. Injection of sfrp1-encoding mRNA into Xenopus embryos causes early developmental defects to axial and neural patterning, which would make it problematic for analysing any specific functions of sfrp1 in vertebrate heart development (not shown). Since Wnt signalling has prominent roles during axial and neural patterning in early Xenopus embryos (reviewed by Hoppler, 2008), overexpression of a known modulator of Wnt signalling, such as sfrp1, is expected to interfere with those processes (e.g. Mii and Taira, 2009).
Since sfrp1 is expressed during mid-organogenesis stages in heart-forming tissues, we adapted a transgenic approach designed to deliver stage-specific inducible expression (e.g. Wheeler et al., 2008) (Fig. 4A-C). When experimental sfrp1 overexpression was induced during organogenesis (stage 22), tadpole embryos developed with enlarged, but apparently normally functioning, heart muscle (Fig. 4P) that almost filled the pericardial cavity and, remarkably, caused the ventral body wall to extend and retract with every heartbeat (supplementary material Movie 2). We analysed whether this phenotype was a consequence of changes to heart development during organogenesis stages by monitoring heart development-associated molecular marker gene expression (see above). The expression of myocardium differentiation markers was not only strong but the expression domain was also extended (compare Fig. 4I,K with 4H,J), consistent with the subsequent formation of an enlarged heart muscle (Fig. 4P,R,T,U). Analysis of images revealed that this enlarged heart muscle phenotype was not due to unusually large individual cardiomyocytes (cardiac hypertrophy) but rather to increased numbers of cardiomyocytes (Fig. 4V) (i.e. cardiac hyperplasia, but see below regarding cell proliferation). sfrp1 overexpression during organogenesis stages also increased cardiogenic transcription factor gene expression (Fig. 4M) and particularly expanded their expression domains (compare Fig. 4E,G with 4D,F).
Fig. 4.
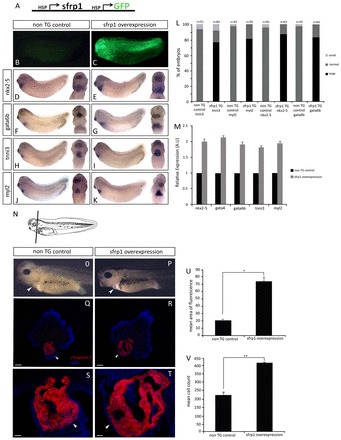
Gain-of-function experiments show that sfrp1 promotes myocardium differentiation. (A) Transgene construct containing sfrp1 and GFP under the control of heat shock promoters allowing for stage-specific induction of overexpression. (B,C) Identification of non-transgenic control (B) and GFP-positive transgenic (C) embryos. (D-K) WISH analysis at stage 32 on non-transgenic control and on sfrp1 transgene-expressing embryos (induced to express transgenes at stage 22). Note the increase in gene expression of cardiogenic transcription factors (E,G) and myocardial differentiation markers (I,K) compared with controls (D,F,H,J), indicating an increase in myocardial tissue. (L) Quantification of WISH analysis showing an increase in cardiac gene expression in sfrp1-overexpressing transgenic (TG) embryos compared with non-transgenic controls. (M) qPCR analysis at stage 32 shows an increase in expression of both cardiogenic transcription factor and myocardial differentiation marker genes in sfrp1-overexpressing transgenic embryos relative to their normal expression in non-transgenic controls. (N) Schematic of a stage 41 embryo indicating the orientation of sections in Q-T. (O,P) Phenotype of non-transgenic control embryo (O) and embryo with stage-specific sfrp1 overexpression (P). Note the increase in heart size and loss of pericardial cavity space (arrowheads) in sfrp1 transgenic embryos compared with control. (Q-T) Differentiation of the heart muscle was analysed at stage 41 by cTnT immunofluorescence (red), with DAPI counterstaining (blue). Non-transgenic control embryos show normal cTnT expression confirming normal myocardial development (Q,S). sfrp1-expressing transgenic embryos show increased expression of cTnT indicating more myocardial tissue (R,T). Note loss of the pericardial cavity due to the increase in myocardial tissue in sfrp1 transgenic embryos (arrowheads). (U) Quantification showing the significant increase in the amount of myocardial tissue as represented by cTnT expression in sfrp1 transgenic embryos (n=4, *P<0.05). (V) Mean cell counts showing a significant increase in cell number in embryos overexpressing sfrp1 compared with controls, whereas cell size appeared unaffected (n=3, **P<0.01). Data are mean ± s.e.m. Scale bars: 50 μm in Q,R; 200 μm in S,T.
sfrp1 regulates cell fate for myocardium development
Our loss- and gain-of-function experiments show that sfrp1 controls the size of the developing heart muscle by regulating cell numbers in the myocardium. We wanted to test possible mechanisms: whether sfrp1 promoted cell proliferation or moderated apoptosis in the myocardium, or, alternatively, whether sfrp1 regulated cell fate decisions in the cardiac mesoderm, inciting cells to follow a differentiation pathway in the myocardium leading to cardiomyocytes.
We were unable to find evidence for altered cell proliferation or apoptosis in heart development in response to sfrp1 (supplementary material Fig. S1; data not shown). Lineage-tracing experiments further showed that differentiating cells do not disappear but appear to undergo a cell fate change when sfrp1 is knocked down (supplementary material Fig. S6). Indeed, marker genes associated with non-muscular heart tissue [fzd7 (Wheeler and Hoppler, 1999), gata5 (Gessert and Kühl, 2009)] were affected in our sfrp1 experiments in the opposite way to myocardium markers (Fig. 5), consistent with such a cell fate change. There was no apparent change to the expression of marker genes associated with the second heart field [bmp4 and isl1 (Gessert and Kühl, 2009); supplementary material Fig. S5].
Fig. 5.
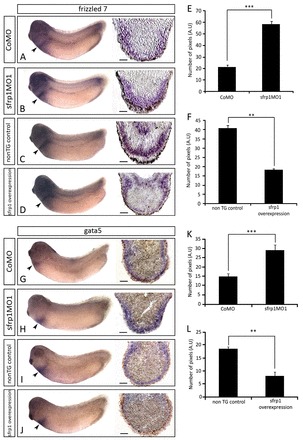
Experimental sfrp1 manipulation causes complementary effects on non-myocardial marker gene expression. (A-D) WISH analysis showing frizzled 7 (fzd7) expression (arrowheads) in CoMO-injected (A), sfrp1MO1-injected (B), non-transgenic control (C) and sfrp1-overexpressing transgenic (D) embryos, together with corresponding sections through the posterior pericardium. (G-J) WISH analysis showing gata5 expression (arrowheads) in CoMO (G), sfrp1MO1 (H), non-transgenic control (I) and sfrp1-overexpressing (J) embryos, with corresponding posterior pericardial sections. (E,F,K,L) Analysis of fzd7 (E,F) and gata5 (K,L) sections representing the number of pixels measured from the apex of the section to the edge of gene expression staining. CoMO compared with sfrp1MO1: n=4, ***P<0.001 (E,K). Non-transgenic control compared with sfrp1 overexpression: n=3, **P<0.01 (F,L). Data are mean ± s.e.m.
sfrp1 regulates heart development as an inhibitor of wnt6 function
Although sfrp1 was initially described as an extracellular Wnt inhibitor (Xu et al., 1998), evidence for a more elaborate Wnt signalling modulating activity has emerged (Mii and Taira, 2009). We had previously identified wnt6 as an endogenous regulator of cardiogenic transcription factors that subsequently regulate the expression of downstream structural heart muscle genes (Afouda et al., 2008; Lavery et al., 2008b; Martin et al., 2010). We therefore tested whether sfrp1 was promoting heart muscle development during organogenesis stages by interfering with Wnt6 signalling.
We first tested in a functional assay whether sfrp1 protein was capable of interfering with the signalling activity of Wnt6. wnt6 is prominently expressed during organogenesis stages (Lavery et al., 2008a), and there is little evidence to suggest an earlier function in axial patterning (Lavery et al., 2008b). However, endogenous Wnt signalling is clearly pivotal for establishing the embryonic axis (e.g. reviewed by Hoppler, 2008), and artificial Wnt6 overexpression in the early embryo is able to interfere with axial patterning, for instance to induce axis duplication (Fig. 6C,E) (see also Lavery et al., 2008b). We found that this artificial axis-inducing activity of Wnt6 signalling was reduced by artificial co-expression of sfrp1 (Fig. 6D,E), which shows that sfrp1 is capable of inhibiting Wnt6 signalling function in vivo.
If sfrp1 function during heart organogenesis is dependent on its Wnt6-inhibiting activity then the phenotype that we describe above for the sfrp1 knockdown would be a consequence of unchecked excessive Wnt6 signalling. To test this hypothesis, we carried out a double-knockdown experiment with co-injection of both sfrp1- and wnt6-inhibiting MOs (Fig. 7G-N). Indeed, heart muscle development was dramatically restored when Wnt6 was knocked down in addition to sfrp1. The amount of heart muscle in the double-knockdown embryos exceeded that in normal embryos and resembled much more closely that of the Wn6 single knockdown [compare Fig. 7E,F,I with figure 2 in Lavery et al. (Lavery et al., 2008b)].
Fig. 7.
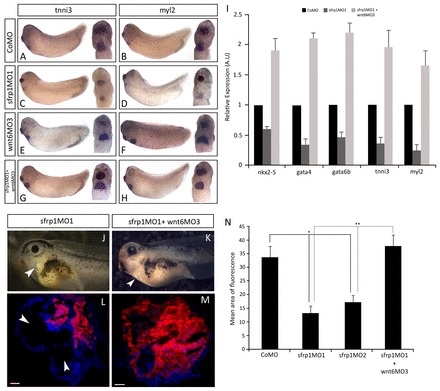
sfrp1 promotes heart development by restraining wnt6-mediated repression. (A-H) WISH analysis on stage 32 embryos highlighting the gene expression pattern of structural myocardial differentiation markers. Embryos injected with CoMO showed normal expression (A,B), whereas embryos injected with sfrp1MO1 showed reduced myocardial gene expression (C,D). Co-injection of wnt6MO3 and sfrp1MO1 caused an enlarged area of myocardial marker gene expression (G,H) reminiscent of the phenotype caused by injection of wnt6MO3 alone [E,F; compare with figure 2 in Lavery et al. (Lavery et al., 2008b)]. (I) qRT-PCR analysis at stage 32 of MO-injected embryos shows decreased gene expression of both cardiogenic transcription factors and myocardial differentiation markers in sfrp1MO1-injected embryos and an increase in both when sfrp1MO1 was co-injected with wnt6MO3, relative to the normal expression in CoMO-injected embryos. (J,K) Phenotype of sfrp1MO1-injected embryo and for embryo co-injected with sfrp1MO1 and wnt6MO3 at stage 41. Note the change in heart size (arrowheads). (L,M) Differentiation of the heart was analysed at stage 41 by cTnT immunofluorescence (red) with DAPI counterstaining (blue). sfrp1MO1 injection causes reduced cTnT expression (L), but embryos co-injected with sfrp1MO1 and wnt6MO3 show enlarged cTnT expression indicating an increase in the amount of differentiated myocardial tissue [M; compare with figure 2 in Lavery et al. (Lavery et al., 2008b)]. Arrowheads indicate loss of myocardial tissue (J,L) and increase in heart size (K). (N) Quantification showing a significant increase in the amount of myocardial tissue as represented by cTnT expression in co-injected embryos compared with embryos injected with sfrp1MO1 alone, which exhibited a significant loss of myocardial tissue relative to CoMO. n=4, **P<0.01 and *P<0.05. Data are mean ± s.e.m. Scale bars: 200 μm in L,M.
Our results allow us to conclude that endogenous sfrp1 regulates the size of the developing heart muscle by inhibiting endogenous Wnt6 signalling, which functions to restrict the size of the differentiating myocardium.
Mathematical modelling predicts and in vivo experiments confirm sfrp1 function in a negative-feedback regulatory loop
It is not intuitively clear why regulation of heart muscle development by Wnt signalling requires components that activate and others that inhibit Wnt signalling. In order to explore the evolved architecture of this gene regulatory network (GRN) in vertebrate heart muscle development, we developed a one-dimensional mathematical model. Our model simulates seven cells along the distal to medial axis (Fig. 8), containing the ectodermal cell as the source of Wnt6 signalling and cardiac mesoderm cells patterned into myocardium (heart muscle) and non-muscular heart tissue (in this model simply referred to as pericardium). The model ignores the endocardium or any other tissues and assumes that they do not contribute to patterning of the cardiac mesoderm into muscular and non-muscular tissues. The model focuses on exploring the extracellular signalling network architecture and therefore reduces the intracellular cardiogenic GRN in responding cardiac mesoderm cells to a bare minimum (Fig. 8A); the generally mutually supporting function of different cardiogenic transcription factor genes (e.g. Searcy et al., 1998; Davidson and Erwin, 2006; Peterkin et al., 2007) is represented by just one autoregulated gene. Our model also assumes that wnt6 expression in the ectoderm is not influenced by events in the cardiac mesoderm and assumes relatively short-range diffusion and therefore a steep gradient, following current thinking in the field (e.g. Mii and Taira, 2009).
Fig. 8.
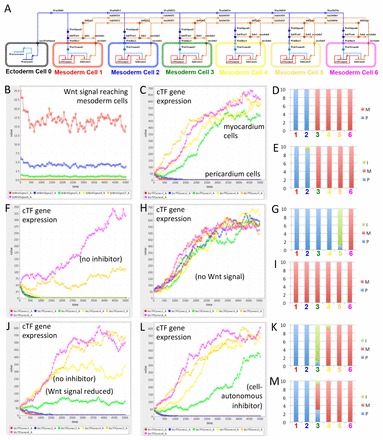
One-dimensional multicellular model of a Wnt negative-feedback gene regulatory network controlling myocardium differentiation. (A) One-dimensional model of a gene regulatory network (GRN) designed in BioTapestry Editor. Note the ectodermal cell (source of Wnt signal) at the far left, with distal to medial cardiac mesoderm cells from left to right, in which myocardium differentiation potential is controlled by cardiogenic transcription factor (cTF) gene expression. Colours of cells correspond to simulation output from the Dizzy simulator (see below). (B) Simulated value of Wnt signal (in arbitrary units, but related to amount) reaching cardiac mesoderm cells. (C) Simulated expression of cTF gene expression (in arbitrary units related to amount) in different cardiac mesoderm cells. Note that simulation of the full model (as illustrated in A) predicts clear patterning into two populations of cells according to cTF gene expression: into prospective non-muscular cells that have lost cTF expression (here simply called pericardium) and prospective muscular cells gaining strong cTF expression (myocardium). (D) Repeated simulation shows reproducibility of output suggesting reliability of patterning. Ten random simulations are illustrated in a stacked column chart. Cells were attributed to the pericardium (P) if the cTF expression was lower than 30 and to the myocardium (M) if higher than 300. Coloured numbers correspond to cells illustrated in A and simulation output in B and C. (E) Simulation of a GRN lacking negative regulation of inhibitor gene expression (i.e. inhibitor gene expression only regulated by cTF genes) suggests still relatively reliable patterning with only occasional cells with predicted intermediate (I) (i.e. between 30 and 300) values of cTF expression. (F) Simulation of an inhibitor loss-of-function experiment predicting many fewer cells with sufficient cTF gene expression for myocardium differentiation. (G) Repeated simulation of experiment as in F. (H) Simulation of Wnt signal loss-of-function experiment (or Wnt and inhibitor double loss-of-function experiment) confirming that the GRN is designed with the default for cardiac mesoderm cells being myocardium differentiation. (I) Repeated simulation of experiment as in H. (J) Simulation of an alternative GRN with only Wnt regulation (i.e. no inhibitor) but with values for Wnt signal adjusted so that the simulation usually predicts at least two pericardial and two myocardial cells suggests less clear patterning giving rise to cells with intermediate identity. (K) Repeated simulation of the alternative GRN as in J. (L) Simulation of an alternative GRN with a cell-autonomous inhibitor predicts relatively clear patterning into pericardial and myocardial cells. (M) Repeated simulation of the alternative GRN as in L suggests less reliable patterning into two pericardium and four myocardium cells than the GRN illustrated in A (compare with D). See supplementary material Models 1-9.
A model without feedback regulation suggested that a Wnt signal was unable to pattern cardiac mesoderm cells into two distinct populations (i.e. muscular versus non-muscular; Fig. 8J). An alternative model with altered cooperativity (equivalent to introducing a Hill coefficient) in the autoregulation of cardiogenic transcription factor expression clearly improved the simulated outcome, predicting clear patterning into two distinct populations of cells (supplementary material Model 8), but a model involving inhibition of sfrp1 expression by Wnt signalling (Fig. 8A; supplementary material Model 1) predicted reliable patterning of the cardiac mesoderm not only into two distinct populations of cells (Fig. 8C) but also in proportions corresponding approximately to the situation observed in the embryo [i.e. relatively few non-muscular cells and a substantial myocardium (Raffin et al., 2000)].
Thus, the modelling predicts that the inhibitor of Wnt signalling (here sfrp1) needs to be negatively regulated by Wnt signalling itself, either by cardiogenic transcription factors, the expression of which is regulated by Wnt signalling activity, or by additional mechanisms (supplementary material compare Models 1 or 8 with 6). This was a modelling prediction that we could verify in the biological system. Indeed, expression of endogenous sfrp1 was found to be absent in the heart-forming region of sfrp1 knockdown embryos (Fig. 9B), which is consistent with sfrp1 expression being inhibited by endogenous Wnt6 signalling. The absence of sfrp1 mRNA after MO-mediated inhibition could possibly be an artefact facilitated by processes such as nonsense-mediated mRNA decay. However, strong sfrp1 mRNA expression was restored to the myocardium in Wnt6 and sfrp1 double-knockdown embryos (Fig. 9C). These findings confirm negative regulation of sfrp1 expression by Wnt signalling in embryos, as predicted by mathematical modelling. We additionally tested in embryos whether sfrp1 expression was dependent on cardiogenic transcription factor gene function as assumed in our model. Indeed, knockdown of cardiogenic gata4 or gata6 resulted in loss of detectable sfrp1 expression in the heart (Fig. 9D,E).
Mathematical modelling allows the simulation of experiments that would be impossible to carry out in real embryos. One such simulation provided an unexpected result. sfrp1 encodes a secreted Wnt inhibitor; thus, when we modified the model to simulate a cell-autonomous Wnt inhibitor instead, we expected a dramatic effect on the patterning of cardiac mesoderm cells, but to our surprise clear patterning into muscular and non-muscular tissue still occurred (Fig. 8L). Although both the in silico simulations and the in vivo experiments emphasised the importance of negative-feedback regulation, the relevance of involving a secreted Wnt inhibitor such as sfrp1 remained initially unclear.
By altering the chosen parameters for any specific simulation, in particular the strength of wnt6 expression in the ectoderm cell, the precise location of the resulting split between the myocardium and non-muscular heart tissue can be changed. Repeated simulations assuming a secreted Wnt inhibitor (mimicking the endogenous sfrp1) led to more reliable patterning to occur close to the Wnt source than repeated simulations with an assumed cell-autonomous Wnt inhibitor (when other parameters were optimised; compare Fig. 8M with 8D). This does of course reflect the normal situation in the embryo, with a large portion of the cardiac mesoderm differentiating into muscular tissue (e.g. Fig. 1L) and fewer cells destined for non-cardiac heart tissues, such as the pericardium (e.g. Fig. 5A,G) (Raffin et al., 2000).
DISCUSSION
Schneider and Mercola (Schneider and Mercola, 2001) assayed the heart development-promoting activity of Dickkopf (dkk1) and other Wnt signalling inhibitors, including sfrp1, in early Xenopus embryos. Whereas dkk1 proved at those early stages to be a potent inducer of many later aspects of cardiac development, sfrp1 in the same assay turned out to be much weaker and only capable of causing a slight increase in nkx2-5 expression. We have now discovered that sfrp1 function is required at later stages - during organogenesis to promote heart muscle differentiation: endogenous sfrp1 is expressed in the differentiating myocardium during organogenesis stages; overexpression at the same stages causes the subsequent differentiation of an enlarged heart muscle; and interference with endogenous sfrp1 expression reduces heart muscle tissue. Although our experiments cannot rule out additional activities [such as those described previously (Lopez-Rios et al., 2008; Kobayashi et al., 2009; Mii and Taira, 2009)], both our gain- and loss-of-function experiments support the notion that sfrp1 functions in this tissue as an inhibitor of wnt6.
Wnt signalling is a prominent mitogen in some developing tissues; however, we were unable to find evidence for sfrp1 regulating the size of the myocardium by altering the cell cycle (supplementary material Fig. S1), which is in accordance with previous findings (Movassagh and Philpott, 2008) that early cardiac differentiation is largely independent of cell cycle control [but see Goetz et al. (Goetz et al., 2006) for later stage cardiomyocyte differentiation]. Nor could we find evidence for sfrp1 function affecting apoptosis in the heart (data not shown), in accordance with relatively little apoptosis being found during normal early heart development (e.g. Hensey and Gautier, 1998). Lineage-tracing experiments further indicate that these cells do not multiply or disappear (supplementary material Fig. S6). Our findings suggest instead that sfrp1 function regulates cell identity by allowing maintenance of cardiogenic gene expression of not only markers for myocardium differentiation but also cardiogenic transcription factors. This is consistent with our previous findings that Wnt/β-catenin signalling restricts myocardium differentiation by regulating the expression of cardiogenic transcription factors (Afouda et al., 2008; Lavery et al., 2008b; Martin et al., 2010). Indeed, the phenotype of our stage-specific sfrp1 overexpression is remarkably similar to that previously described for the overexpression of cardiogenic transcription factors [such as nkx2-5 (Cleaver et al., 1996)]. However, if sfrp1 activity is not primarily affecting cell proliferation or apoptosis but cell identity for myocardium differentiation, the extra cells recruited to the myocardium in the sfrp1 overexpression experiments must be missing elsewhere and the cells lost from the myocardium in the sfrp1 loss-of-function experiments must end up somewhere else. Cardiac mesoderm progenitors give rise to muscular myocardium and non-muscular tissues (e.g. Raffin et al., 2000; Kruithof et al., 2006; Zhou et al., 2008). There are currently few specific markers that we can rely on for differentiated cell types deriving from cardiac mesoderm other than myocardium; however, our experiments suggest effects on marker gene expression previously linked to the pericardium [fzd7 (Wheeler and Hoppler, 1999), gata5 (Gessert and Kühl, 2009)].
A requirement for endogenous sfrp1 in regulating embryonic heart muscle differentiation is likely to represent an ancient function that is conserved in mammalian heart development. Mammalian Sfrp1 is also strongly expressed in embryonic myocardium, differentiated heart muscle and cardiomyocytes, and, as in Xenopus, expression is specifically excluded from the pericardium (Jaspard et al., 2000). Remarkably, Sfrp1 is transiently upregulated in a mouse model following experimentally induced myocardial infarction, predominantly at the infarct border zone, and, moreover, transgenic overexpression of Sfrp1 in the mouse model resulted in reduced infarct size and improved recovery (Barandon et al., 2003), which has since been found to be part of a coordinated response to myocardial infarction that also involves increased neovascularisation and reduced inflammatory response and scar formation (Barandon et al., 2011). During early mammalian heart development, Sfrp1 appears to function at least partially redundantly with other factors (Trevant et al., 2008), specifically Sfrp2, which, however, also mediates prominent Wnt-independent functions in the heart (Kobayashi et al., 2009). Modulation of Wnt signalling, particularly by sFRPs, appears promising for therapeutic intervention, particularly for left ventricle remodelling in adult heart patients (reviewed by Bergmann, 2010).
Mathematical modelling provided us with a useful tool for exploring the network architecture of the gene regulatory relationships discovered by functional experiments but also for suggesting additional regulatory links, which were then confirmed, such as the inhibition of sfrp1 expression by Wnt signalling and the dependence on cardiogenic gata transcription factor expression. It might be true, to some extent, that by tinkering with parameter values many modelled networks can be made to fit, at least qualitatively, with the expected outcomes from previous experimental analysis. Also, ultimately, much more quantitative and detailed measurements of gene expression in the biological system will be required for building more accurate and more predictive mathematical models. However, the tinkering with parameters to fit models to qualitative biological results was substantially more difficult with some network structures and much easier with others, suggesting that the particular network architecture might be much more important in mathematical models for reliable simulation of patterning than precise parameter values, and the same might of course also apply to the real biological networks that they are designed to represent. A limitation of our model is possibly that the multicellular arrangement of the simulated cells is static, whereas in the embryo there are morphogenetic movements of cardiac mesoderm cells not only early in heart development, as associated with the migration of the heart fields, but also physical separation with the formation of a pericardial cavity.
Mathematical modelling of our GRN emphasised the importance of negative-feedback regulation of Wnt signalling but it did not immediately offer obvious explanations for why this negative feedback needed to be non-cell-autonomous. ‘Why’ questions can be problematic when considering evolved biological processes. Repeated simulation suggests, however, that non-cell-autonomy of the feedback Wnt inhibitor might facilitate more reliable patterning close to the source of the Wnt signal, which in this biological context results in fewer non-muscular cells and a substantially larger myocardium. Such reliability is of particular importance because the size of the future heart muscle is determined primarily by regulating cell fate within a given population of cells (here the cardiac mesoderm), with little apparent regulation of cell proliferation or apoptosis (see above). If the Wnt gradient were not as steep as is currently assumed (e.g. Mii and Taira, 2009), differences in the simulated outcome between models with a cell-autonomous versus a secreted inhibitor would be expected to be more pronounced. Alternatively, our one-dimensional model might not capture aspects of the biological system within the myocardium for which there may be a requirement for a non-cell-autonomous inhibitor [such as a community effect (Saka et al., 2011)].
Although feedback regulation in Wnt signalling is well established [e.g. axin2 (Lustig et al., 2002)], to our knowledge this is the first evidence for sfrp genes acting in a regulatory negative-feedback loop in canonical Wnt/β-catenin signalling, where sfrp protein not only regulates Wnt signalling but sfrp1 expression is in turn negatively controlled by Wnt/β-catenin signalling. sfrp genes might mediate non-cell-autonomous feedback regulation of Wnt/β-catenin signalling in other tissues. Interestingly, sfrp1 appears to be involved in the endogenous response to myocardial infarction, to rescue cardiomyocyte function (see above). It is tempting to speculate that a similar regulatory network to the one we describe in Xenopus embryonic development operates in the adult infarcted heart.
Supplementary Material
Acknowledgments
We thank Yvonne Turnbull for technical assistance, Grant Wheeler (now University of East Anglia) for help with initial sfrp1 overexpression experiments, and Gioia Petrighi Polidori and Yasushi Saka for discussions. S.H. thanks the Woods Hole Marine Laboratory course on Gene Regulatory Networks for Development for excellent and inspirational training in modelling.
Footnotes
Funding
N.G. was supported by a non-clinical PhD research scholarship from the British Heart Foundation. D.L.L. was supported by the James Mearns Charitable Trust.
Competing interests statement
The authors declare no competing financial interests.
Supplementary material
Supplementary material available online at http://dev.biologists.org/lookup/suppl/doi:10.1242/dev.088047/-/DC1
References
- Afouda B. A., Martin J., Liu F., Ciau-Uitz A., Patient R., Hoppler S. (2008). GATA transcription factors integrate Wnt signalling during heart development. Development 135, 3185–3190 [DOI] [PubMed] [Google Scholar]
- Amaya E., Kroll K. L. (1999). A method for generating transgenic frog embryos. Methods Mol. Biol. 97, 393–414 [DOI] [PubMed] [Google Scholar]
- Barandon L., Couffinhal T., Ezan J., Dufourcq P., Costet P., Alzieu P., Leroux L., Moreau C., Dare D., Duplàa C. (2003). Reduction of infarct size and prevention of cardiac rupture in transgenic mice overexpressing FrzA. Circulation 108, 2282–2289 [DOI] [PubMed] [Google Scholar]
- Barandon L., Casassus F., Leroux L., Moreau C., Allières C., Lamazière J. M. D., Dufourcq P., Couffinhal T., Duplàa C. (2011). Secreted frizzled-related protein-1 improves postinfarction scar formation through a modulation of inflammatory response. Arterioscler. Thromb. Vasc. Biol. 31, e80–e87 [DOI] [PubMed] [Google Scholar]
- Bergmann M. W. (2010). WNT signaling in adult cardiac hypertrophy and remodeling: lessons learned from cardiac development. Circ. Res. 107, 1198–1208 [DOI] [PubMed] [Google Scholar]
- Bolouri H., Davidson E. H. (2003). Transcriptional regulatory cascades in development: initial rates, not steady state, determine network kinetics. Proc. Natl. Acad. Sci. USA 100, 9371–9376 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cleaver O. B., Patterson K. D., Krieg P. A. (1996). Overexpression of the tinman-related genes XNkx-2.5 and XNkx-2.3 in Xenopus embryos results in myocardial hyperplasia. Development 122, 3549–3556 [DOI] [PubMed] [Google Scholar]
- Cohen E. D., Miller M. F., Wang Z., Moon R. T., Morrisey E. E. (2012). Wnt5a and Wnt11 are essential for second heart field progenitor development. Development 139, 1931–1940 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson E. H., Erwin D. H. (2006). Gene regulatory networks and the evolution of animal body plans. Science 311, 796–800 [DOI] [PubMed] [Google Scholar]
- Drysdale T. A., Tonissen K. F., Patterson K. D., Crawford M. J., Krieg P. A. (1994). Cardiac troponin I is a heart-specific marker in the Xenopus embryo: expression during abnormal heart morphogenesis. Dev. Biol. 165, 432–441 [DOI] [PubMed] [Google Scholar]
- Duplàa C., Jaspard B., Moreau C., D’Amore P. A. (1999). Identification and cloning of a secreted protein related to the cysteine-rich domain of frizzled. Evidence for a role in endothelial cell growth control. Circ. Res. 84, 1433–1445 [DOI] [PubMed] [Google Scholar]
- Eisenberg C. A., Eisenberg L. M. (1999). WNT11 promotes cardiac tissue formation of early mesoderm. Dev. Dyn. 216, 45–58 [DOI] [PubMed] [Google Scholar]
- Evans S. M., Yan W., Murillo M. P., Ponce J., Papalopulu N. (1995). tinman, a Drosophila homeobox gene required for heart and visceral mesoderm specification, may be represented by a family of genes in vertebrates: XNkx-2.3, a second vertebrate homologue of tinman. Development 121, 3889–3899 [DOI] [PubMed] [Google Scholar]
- Gessert S., Kühl M. (2009). Comparative gene expression analysis and fate mapping studies suggest an early segregation of cardiogenic lineages in Xenopus laevis. Dev. Biol. 334, 395–408 [DOI] [PubMed] [Google Scholar]
- Gessert S., Kühl M. (2010). The multiple phases and faces of wnt signaling during cardiac differentiation and development. Circ. Res. 107, 186–199 [DOI] [PubMed] [Google Scholar]
- Goetz S. C., Brown D. D., Conlon F. L. (2006). TBX5 is required for embryonic cardiac cell cycle progression. Development 133, 2575–2584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gove C., Walmsley M., Nijjar S., Bertwistle D., Guille M., Partington G., Bomford A., Patient R. (1997). Over-expression of GATA-6 in Xenopus embryos blocks differentiation of heart precursors. EMBO J. 16, 355–368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hensey C., Gautier J. (1998). Programmed cell death during Xenopus development: a spatio-temporal analysis. Dev. Biol. 203, 36–48 [DOI] [PubMed] [Google Scholar]
- Hoppler S. (2008). Studying Wnt signaling in Xenopus. Methods Mol. Biol. 469, 319–332 [DOI] [PubMed] [Google Scholar]
- Jaspard B., Couffinhal T., Dufourcq P., Moreau C., Duplàa C. (2000). Expression pattern of mouse sFRP-1 and mWnt-8 gene during heart morphogenesis. Mech. Dev. 90, 263–267 [DOI] [PubMed] [Google Scholar]
- Jiang Y., Evans T. (1996). The Xenopus GATA-4/5/6 genes are associated with cardiac specification and can regulate cardiac-specific transcription during embryogenesis. Dev. Biol. 174, 258–270 [DOI] [PubMed] [Google Scholar]
- Jones S. E., Jomary C. (2002). Secreted Frizzled-related proteins: searching for relationships and patterns. BioEssays 24, 811–820 [DOI] [PubMed] [Google Scholar]
- Kobayashi K., Luo M., Zhang Y., Wilkes D. C., Ge G., Grieskamp T., Yamada C., Liu T. C., Huang G., Basson C. T., et al. (2009). Secreted Frizzled-related protein 2 is a procollagen C proteinase enhancer with a role in fibrosis associated with myocardial infarction. Nat. Cell Biol. 11, 46–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruithof B. P., van Wijk B., Somi S., Kruithof-de Julio M., Pérez Pomares J. M., Weesie F., Wessels A., Moorman A. F., van den Hoff M. J. (2006). BMP and FGF regulate the differentiation of multipotential pericardial mesoderm into the myocardial or epicardial lineage. Dev. Biol. 295, 507–522 [DOI] [PubMed] [Google Scholar]
- Lavery D. L., Hoppler S. (2008a). Analysis of gene expression in Xenopus embryos. Methods Mol. Biol. 469, 335–361 [DOI] [PubMed] [Google Scholar]
- Lavery D. L., Hoppler S. (2008b). Gain-of-function and loss-of-function strategies in Xenopus. Methods Mol. Biol. 469, 401–415 [DOI] [PubMed] [Google Scholar]
- Lavery D. L., Davenport I. R., Turnbull Y. D., Wheeler G. N., Hoppler S. (2008a). Wnt6 expression in epidermis and epithelial tissues during Xenopus organogenesis., Dev. Dyn. 237, 768–779 [DOI] [PubMed] [Google Scholar]
- Lavery D. L., Martin J., Turnbull Y. D., Hoppler S. (2008b). Wnt6 signaling regulates heart muscle development during organogenesis. Dev. Biol. 323, 177–188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopez-Rios J., Esteve P., Ruiz J. M., Bovolenta P. (2008). The Netrin-related domain of Sfrp1 interacts with Wnt ligands and antagonizes their activity in the anterior neural plate. Neural Dev. 3, 19 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lustig B., Jerchow B., Sachs M., Weiler S., Pietsch T., Karsten U., van de Wetering M., Clevers H., Schlag P. M., Birchmeier W., et al. (2002). Negative feedback loop of Wnt signaling through upregulation of conductin/axin2 in colorectal and liver tumors. Mol. Cell. Biol. 22, 1184–1193 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin J., Afouda B. A., Hoppler S. (2010). Wnt/beta-catenin signalling regulates cardiomyogenesis via GATA transcription factors. J. Anat. 216, 92–107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marvin M. J., Di Rocco G., Gardiner A., Bush S. M., Lassar A. B. (2001). Inhibition of Wnt activity induces heart formation from posterior mesoderm. Genes Dev. 15, 316–327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mii Y., Taira M. (2009). Secreted Frizzled-related proteins enhance the diffusion of Wnt ligands and expand their signalling range. Development 136, 4083–4088 [DOI] [PubMed] [Google Scholar]
- Mohun T. J., Leong L. M., Weninger W. J., Sparrow D. B. (2000). The morphology of heart development in Xenopus laevis. Dev. Biol. 218, 74–88 [DOI] [PubMed] [Google Scholar]
- Mohun T., Orford R., Shang C. (2003). The origins of cardiac tissue in the amphibian, Xenopus laevis. Trends Cardiovasc. Med. 13, 244–248 [DOI] [PubMed] [Google Scholar]
- Monaghan A. P., Kioschis P., Wu W., Zuniga A., Bock D., Poustka A., Delius H., Niehrs C. (1999). Dickkopf genes are co-ordinately expressed in mesodermal lineages. Mech. Dev. 87, 45–56 [DOI] [PubMed] [Google Scholar]
- Movassagh M., Philpott A. (2008). Cardiac differentiation in Xenopus requires the cyclin-dependent kinase inhibitor, p27Xic1. Cardiovasc. Res. 79, 436–447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mummery C. L., Davis R. P., Krieger J. E. (2010). Challenges in using stem cells for cardiac repair. Sci. Transl. Med. 2, 27ps17 [DOI] [PubMed] [Google Scholar]
- Nakajima Y., Sakabe M., Matsui H., Sakata H., Yanagawa N., Yamagishi T. (2009). Heart development before beating. Anat. Sci. Int. 84, 67–76 [DOI] [PubMed] [Google Scholar]
- Nieuwkoop P. D., Faber J. (1967). Normal Table of Xenopus laevis (Daudin): a Systematical and Chronological Survey of the Development from the Fertilized Egg to the End of Metamorphosis. Amsterdam: North Holland Publishing; [Google Scholar]
- Onizuka T., Yuasa S., Kusumoto D., Shimoji K., Egashira T., Ohno Y., Kageyama T., Tanaka T., Hattori F., Fujita J., et al. (2012). Wnt2 accelerates cardiac myocyte differentiation from ES-cell derived mesodermal cells via non-canonical pathway. J. Mol. Cell. Cardiol. 52, 650–659 [DOI] [PubMed] [Google Scholar]
- Pandur P., Läsche M., Eisenberg L. M., Kühl M. (2002). Wnt-11 activation of a non-canonical Wnt signalling pathway is required for cardiogenesis. Nature 418, 636–641 [DOI] [PubMed] [Google Scholar]
- Peterkin T., Gibson A., Patient R. (2007). Redundancy and evolution of GATA factor requirements in development of the myocardium. Dev. Biol. 311, 623–635 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raffin M., Leong L. M., Rones M. S., Sparrow D., Mohun T., Mercola M. (2000). Subdivision of the cardiac Nkx2.5 expression domain into myogenic and nonmyogenic compartments. Dev. Biol. 218, 326–340 [DOI] [PubMed] [Google Scholar]
- Saka Y., Lhoussaine C., Kuttler C., Ullner E., Thiel M. (2011). Theoretical basis of the community effect in development. BMC Syst. Biol. 5, 54 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneider V. A., Mercola M. (2001). Wnt antagonism initiates cardiogenesis in Xenopus laevis. Genes Dev. 15, 304–315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Searcy R. D., Vincent E. B., Liberatore C. M., Yutzey K. E. (1998). A GATA-dependent nkx-2.5 regulatory element activates early cardiac gene expression in transgenic mice. Development 125, 4461–4470 [DOI] [PubMed] [Google Scholar]
- Smith W. C., Harland R. M. (1991). Injected Xwnt-8 RNA acts early in Xenopus embryos to promote formation of a vegetal dorsalizing center. Cell 67, 753–765 [DOI] [PubMed] [Google Scholar]
- Thompson H., Andrews W., Parnavelas J. G., Erskine L. (2009). Robo2 is required for Slit-mediated intraretinal axon guidance. Dev. Biol. 335, 418–426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tonissen K. F., Drysdale T. A., Lints T. J., Harvey R. P., Krieg P. A. (1994). XNkx-2.5, a Xenopus gene related to Nkx-2.5 and tinman: evidence for a conserved role in cardiac development. Dev. Biol. 162, 325–328 [DOI] [PubMed] [Google Scholar]
- Trevant B., Gaur T., Hussain S., Symons J., Komm B. S., Bodine P. V., Stein G. S., Lian J. B. (2008). Expression of secreted frizzled related protein 1, a Wnt antagonist, in brain, kidney, and skeleton is dispensable for normal embryonic development. J. Cell. Physiol. 217, 113–126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tzahor E. (2007). Wnt/beta-catenin signaling and cardiogenesis: timing does matter. Dev. Cell 13, 10–13 [DOI] [PubMed] [Google Scholar]
- Warkman A. S., Krieg P. A. (2007). Xenopus as a model system for vertebrate heart development. Semin. Cell Dev. Biol. 18, 46–53 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wheeler G. N., Hoppler S. (1999). Two novel Xenopus frizzled genes expressed in developing heart and brain. Mech. Dev. 86, 203–207 [DOI] [PubMed] [Google Scholar]
- Wheeler G. N., Hamilton F. S., Hoppler S. (2000). Inducible gene expression in transgenic Xenopus embryos. Curr. Biol. 10, 849–852 [DOI] [PubMed] [Google Scholar]
- Wheeler G. N., Lavery D. L., Hoppler S. (2008). Inducible gene expression in transient transgenic Xenopus embryos. Methods Mol. Biol. 469, 431–449 [DOI] [PubMed] [Google Scholar]
- Xu Q., D’Amore P. A., Sokol S. Y. (1998). Functional and biochemical interactions of Wnts with FrzA, a secreted Wnt antagonist. Development 125, 4767–4776 [DOI] [PubMed] [Google Scholar]
- Yang L., Soonpaa M. H., Adler E. D., Roepke T. K., Kattman S. J., Kennedy M., Henckaerts E., Bonham K., Abbott G. W., Linden R. M., et al. (2008). Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 453, 524–528 [DOI] [PubMed] [Google Scholar]
- Zhou B., Ma Q., Rajagopal S., Wu S. M., Domian I., Rivera-Feliciano J., Jiang D., von Gise A., Ikeda S., Chien K. R., et al. (2008). Epicardial progenitors contribute to the cardiomyocyte lineage in the developing heart. Nature 454, 109–113 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.