

Abstract
Background. Maggot debridement therapy (MDT), using Lucilia sericata larvae, represents efficient, simple, and low-cost therapy for the treatment of chronic wounds. Aim. The aim was to investigate the antibiofilm activity of maggot excretions/secretions (ES) against biofilm of wound isolates Staphylococcus aureus (S. aureus), Enterobacter cloacae (E. cloacae), and Proteus mirabilis (P. mirabilis). Methods. Quantification of biofilm formation, was carried out using a microtiter plate assay. Proteolytic activity of maggot ES was performed using skim milk agar plates. A solid phase extraction and reverse phase HPLC C18 chromatography were employed to the isolate of maggot ES antibiofilm compounds. Results. Maggot ES at 100 mg/mL concentration significantly reduced biofilm formation thus disrupting established biofilm of E. cloacae. Heat-treated ES did not show any antibiofilm activity towards E. cloacae. Similar results were obtained in the case of S. aureus; however, the heat-treatment of maggot ES did not affect its antibiofilm activity. Moreover, a compound with molecular weight of 25 kDa exhibiting antibiofilm activity was identified in maggot ES. On the other hand, maggot ES protected and even stimulated P. mirabilis biofilm formation. Conclusions. Our results suggest that maggot ES may act selectively against different bacterial strain.
1. Introduction
It has been known for decades that some chronic bacterial infections are caused by the ability of bacteria to form biofilm [1–7]. The classic example of biofilm involvement in chronic infections is nonhealing dermal wounds. Biofilm growth and its persistence within wounds have recently been suggested as being a contributing factor towards impaired healing [3, 8]. Using cultural and molecular techniques, it has been shown that pathogenic biofilm in chronic wound is generally polymicrobial, with certain species such as Pseudomonas aeruginosa (P. aeruginosa) and Staphylococcus aureus (S. aureus) which are often predominant. Recently, several studies have demonstrated that a wide variety of bacteria with different physiological and phenotypic preferences are common as part of the pathogenic biofilm communities in chronic wounds [9–12]. There is an estimated average of 6.3 bacterial species in chronic ulcers [13]. Chronic wounds associated with biofilm are a worldwide problem joined with high economical costs, social and psychological deprivation, and pain. The treatment of bacterial biofilm in wound is complicated due to the underlying mechanisms of biofilm growth. Furthermore, mixed species of biofilms have complementary metabolic strategies for obtaining nutrients and degrading host immune molecules [14]. Bacteria that reside within mature biofilms are highly resistant to many traditional therapies.
Presently, maggot debridement therapy (MDT) has attracted much attention due to its successful application and efficacy in the elimination of multidrug resistant wound pathogens. In the last few years, several comparative clinical trials investigating the efficacy of MDT have been performed [15, 16]. In terms of debridement, MDT is more effective than conventional therapies [15].
Although, the beneficial debridement effect of larval therapy is well documented, the underlying mechanisms of action, particularly antibacterial and antibiofilm effects, have not been fully elucidated. Nevertheless, maggot Lucilia sericata excretions/secretions (ES) are effective for the disruption of P. aeruginosa and S. aureus biofilms [17, 18]; little is known about antibiofilm activity against another predominant wound pathogen such as Enterobacter cloacae (E. cloacae) and Proteus mirabilis (P. mirabilis).
Therefore, the aim of the study was to investigate the antibiofilm activity of sterile maggot ES against E. cloacae, P. mirabilis, and S. aureus biofilms.
2. Materials and Methods
2.1. Bacterial Strains
Bacterial isolates S. aureus 1141, E. cloacae 2383/10, and P. mirabilis 719/10 from nonhealing wounds were collected from the Department of Clinical Microbiology in Liptovsky Mikulas Hospital (Liptovsky Mikulas, Slovakia) and Prievidza Hospital (Bojnice, Slovakia). The isolates were transported to the Department of Microbiology, Faculty of Medicine, Slovak Medical University (Bratislava, Slovakia).
2.2. Preparation of Sterile and Nonsterile L. sericata Larvae
L. sericata flies were maintained at the Institute of Zoology SAS under the constant conditions according to [19]. The oviposited eggs were divided into two groups. The first group consisted of sterilized eggs in order to obtain sterile larvae (producing sterile ES products) while the second group consisted of nonsterile eggs to hatch nonsterile larvae producing nonsterile ES products. In the sterile group, the eggs were incubated on the blood agar plate until the larvae reached the third instar stage, where sterile ES products were collected. In the second group, the eggs were incubated on a beef liver; nonsterile larvae were also incubated until they reached the third instar, where nonsterile ES products were collected.
2.3. Maggot ES Collection and Preparation
Maggot ES products were collected according to van der Plas et al. [20] with minor modifications. Briefly, the third instar nonsterile and sterile larvae were washed and incubated in Milli-Q ultrapure water for 60 min (50 larvae/200 μL water) at 4°C in a dark place. The incubation was performed at 4°C due to prevention of proteolysis. After incubation, the generated ES products were transferred into a new tube, centrifuged (20 000 g, 4°C, 30 min) to remove solid parts, and finally freeze-dried. Lyophilized ES products were dissolved in Tryptic Soy Broth (TSB) medium (Oxoid, UK) to a final concentration of 100 mg/mL, filtered, and stored at −20°C. The subsequent concentrations of ES products were used in the study: 5 mg/mL, 7.5 mg/mL, 10 mg/mL, 20 mg/mL, 50 mg/mL, and 100 mg/mL. Heat-treated ES (HES) was prepared by incubating native ES products at 100°C for 10 min, centrifuged, and transferred into a new tube.
2.4. Determination of ES Influence on Bacterial Growth
An overnight culture of wound isolates was suspended in phosphate buffered saline (PBS) buffer (pH 7.2); the suspension turbidity was adjusted to a 108 colony forming unit (CFU)/mL and was diluted with a medium to the final concentration of 106 CFU/mL. Ten μL aliquots of suspension were inoculated in each well of sterile 96-well polystyrene plates and supplemented with 90 μL of sterile medium; the medium was diluted with ES and HES products. Plates were incubated at 37°C for 22 h; during this period, the bacterial growth was determined by monitoring the absorbance at 570 nm. Adhesive foils (Sigma Aldrich, Germany) were used to avoid plate contamination during the examination of absorbance in a plate reader (Bio-Rad Laboratories, Hercules, CA, USA).
2.5. Microtiter Plate Biofilm Formation Assay
Biofilm formation assays were performed according to Stepanovi et al. [21] using plastic 96-well tissue culture microtiter plates (Sarstedt, Germany). Briefly, a loop full of cells from a blood agar plate was transferred to a sterile 15 mL polystyrene tube containing 4 mL of PBS. The cells were dispersed with a vortex for 1 min and suspension was passed through a 5 μm pore-size syringe filter to remove large clumps of cells. The suspension turbidity was adjusted to 108 CFU/mL and diluted with TSB broth to a final concentration of 105 CFU/mL. Aliquots of bacterial suspension (10 μL) were transferred to each well of the microtiter plate. After incubating bacterial cultures for 24 h at 37°C, the content of each well was aspirated and wells were washed three times with sterile PBS. The remaining attached bacteria were fixed with 100 μL methanol for 15 min. The plates were then stained with 100 μL of 2% (w/v) crystal violet. Afterwards, the dye bound to the adherent cells was resolubilized with 100 μL of 33% (v/v) glacial acetic acid. The optical density (OD) of each well was measured at 570 nm using an automated microtiter plate reader.
2.6. Biofilm Inhibition Assay
Biofilms were cultured for 24 h as described above, but 10 μL aliquots of bacterial suspension were inoculated in each well of sterile 96-well polystyrene plates and were supplemented with 90 μL of ES and HES diluted in TSB broth in a range of 5–100 mg/mL. After the incubation time, the remaining attached bacteria were quantified as described in the previous section.
2.7. Biofilm Disruption Assay
Biofilms were cultured in a 96-well microtiter plate as described above, with modification according to Stepanovi et al. [21]. Briefly, after incubating bacterial biofilms for 7 h at 37°C, the content of each well was aspirated; the wells were washed three times with sterile PBS, treated with 100 μL ES and HES products diluted in TSB broth (concentration range of 10–100 mg/mL), and cultivated for an additional 16 h at 37°C. Control biofilms were cultured in TSB broth alone. After the incubation period, biofilms were quantified by CFU enumeration and crystal violet staining as described above. In the case of CFU enumeration, biofilms were rinsed three times; adherent bacteria were detached using a swabbing technique. A cotton-tipped swab was then transferred to 5 mL of PBS buffer and mixed by vortex agitating for 60 s. The bacterial suspensions of E. cloacae and S. aureus were serially diluted and spread on blood agar plates. In the case of P. mirabilis MacConkey agar plates (Oxoid, UK) were used. The plates were incubated at 37°C for 24 h.
2.8. Biofilm and Proteinase K Treatment
We examined the effect of proteinase K on biofilm development, on the disruption of established biofilm, and on the cell viability within the biofilm as a positive control and compared potential maggot ES proteolytic activity.
2.9. Determination of Protease Activity
Proteolytic activity assay was performed according to [22], using skim milk agar plates. Briefly, 1 g of bacto-agar (Oxoid, UK) and 10 g nonfat dried milk (Sigma Aldrich) were dissolved in 100 mL distilled water and sterilized. Five μL of ES and HES products in range of 5–100 mg/mL was added in a hole in skim milk agar plates and incubated at 37°C for 24 h. Proteinase K at concentration range from 0.1 to 10 μg/mL was used as positive control.
2.10. Purification of Antibiofilm Compounds from Maggot ES Products
Potential maggot antibiofilm compounds were purified using a solid phase extraction (SPE) methodology according to [23] with minor modifications. SPE was performed using the column Maxi-Clean SPE C18 600 mg (Grace, USA). One hundred mg of ES products was diluted in 50 mL Milli-Q ultrapure water and centrifuged (20 000 g, 4°C, 15 min) to remove solid parts. The SPE column was conditioned with 10 mL acetonitrile (Sigma Aldrich, Germany) and washed with 10 mL distilled water before loading the sample. It was then washed with 10 mL distilled water and eluted with 5 mL acetonitrile. Final eluate was lyophilized and dissolved in TSB medium to a concentration of 35 mg/mL and tested for protease and antibiofilm activity against S. aureus and E. cloacae as described above.
2.11. Chromatography Fractionation
In order to identify the antibiofilm components of ES products, SPE eluate was fractionated using a reverse phase (RP)-HPLC chromatography (Beckman System Gold, USA) coupled with a C18 column (250 mm × 4.6 mm; 5 μm) (Grace, USA) at a flow rate 0.3 mL/min, by using a gradient from 0 to 90% (v/v) acetonitrile (containing 0.1% (v/v) trifluoroacetic acid), during 70 min. The fractions were lyophilized and dissolved in 100 μL of TSB and tested for antibiofilm activity against S. aureus. The active fraction was then fractionated using a C4 column (250 mm × 4.6 mm; 5 μm) (Grace, USA) and tested under the same conditions. The purity of active fractions was checked by electrophoresis on 16.5% Tricine-SDS-PAGE gel using a Mini-Protean II electrophoresis cell (Bio-Rad, CA, USA). Protein bands were detected after staining with Coomassie Brilliant Blue R-250.
2.12. Statistical Analysis
Results are presented as the mean with standard error (SEM). All data were statistically analysed from three independent experiments using a Student's t-test to determine whether there were significant differences between each ES/HES treatment and untreated control. P ≤ 0.05 was considered to be significant. Analyses were performed using GraphPad Prism (GraphPad Software Inc., USA).
3. Results
3.1. The Effect of Maggot ES on Biofilm Formation of Wound Isolates
The ability of maggot ES products to inhibit biofilm formation of E. cloacae, S. aureus, and P. mirabilis was assessed in a 96-well microtiter plate. Maggot ES products with concentration gradient 5–100 mg/mL and HES at concentration of 100 mg/mL did not significantly affect the growth of planktonic bacterial cells of tested isolates. Figure 1 shows significant inhibition of biofilm formation of E. cloacae and of S. aureus in the presence of maggot ES compared to control. Biofilm formation was averagely inhibited by 47% in S. aureus and by approximately 45% in E. cloacae isolate. The most effective maggot ES concentration which inhibited biofilm formation in S. aureus was 50 mg/mL. The maggot ES concentration of 100 mg/mL was observed as the most effective in E. cloacae where 63% decrease in biofilm formation was observed.
Figure 1.
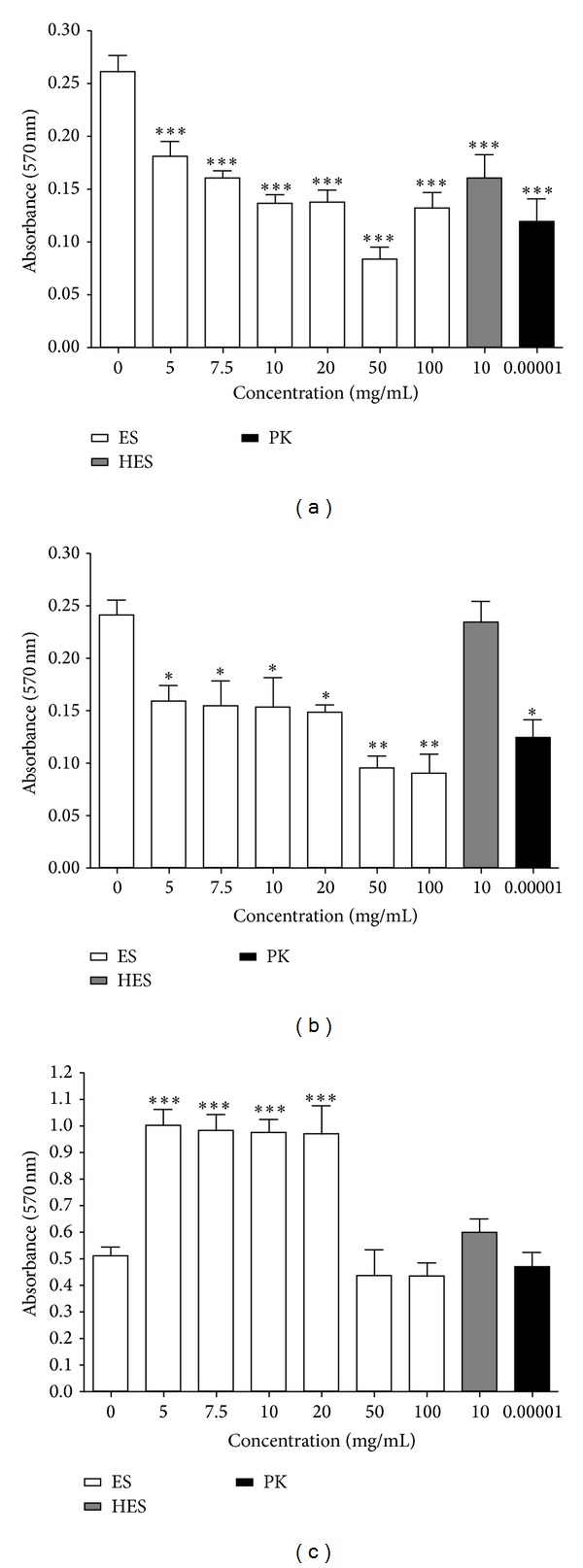
Effects of maggot ES, HES at concentration of 10 mg/mL, and proteinase K (PK) at concentration of 0.01 μg/mL on (a) Staphylococcus aureus, (b) Enterobacter cloacae, and (c) Proteus mirabilis biofilm formation in Tryptone Soya Broth medium after 24 h in 96-well plate. The values of absorbance are mean SEM of three independent assays. The data were statistically analyzed by the Student's t-test (*P < 0.05, **P < 0.01, ***P < 0.001).
In case of P. mirabilis, the side effect of maggot ES was observed. A statistically significant stimulation of P. mirabilis biofilm formation was documented in the presence of maggot ES at concentration range 5–20 mg/mL. In order to partially identify the components responsible for the antibiofilm effect of maggot ES, HES was used. HES did not affect biofilm formation of E. cloacae and P. mirabilis, while in the case of S. aureus, HES caused a 51% reduction of biofilm formation. To determine the role of heat unstable ES compounds such as larval protease(s) in inhibition process of biofilm formation, we compared the effect of proteinase K with maggot ES treatment. E. cloacae and S. aureus exhibited reduction of biofilm formation comparable with ES treatment when treated with proteinase K. The biofilm formation of P. mirabilis was unaffected by proteinase K.
3.2. The Disruption of Established Biofilm of Wound Isolates by Maggot ES
The positive clinical outcomes of MDT suggest that maggot ES products are able to disperse the matured wound biofilm. We determined the ability of maggot ES and HES to detach already established S. aureus, E. cloacae, and P. mirabilis biofilms. We found that established biofilm of E. cloacae was nearly fully disrupted (95% reduction) in the presence of maggot ES at concentration 100 mg/mL and concentration 50 mg/mL partially disrupted (56% reduction) in the case of S. aureus compared to the control biofilm (Figure 2). On the other hand, no decrease in P. mirabilis biofilm mass in the presence of maggot ES was observed. Furthermore, maggot ES at lower concentrations (10 and 20 mg/mL) significantly increased biofilm mass of P. mirabilis. HES had no antibiofilm activity against matured biofilm of S. aureus and E. cloacae, while it increased the biofilm mass of P. mirabilis. The potential biofilm-disrupting trend of termolabile ES products was confirmed by proteinase K. Proteinase K significantly decreased S. aureus and E. cloacae biofilm mass comparable to observed maggot ES effects.
Figure 2.
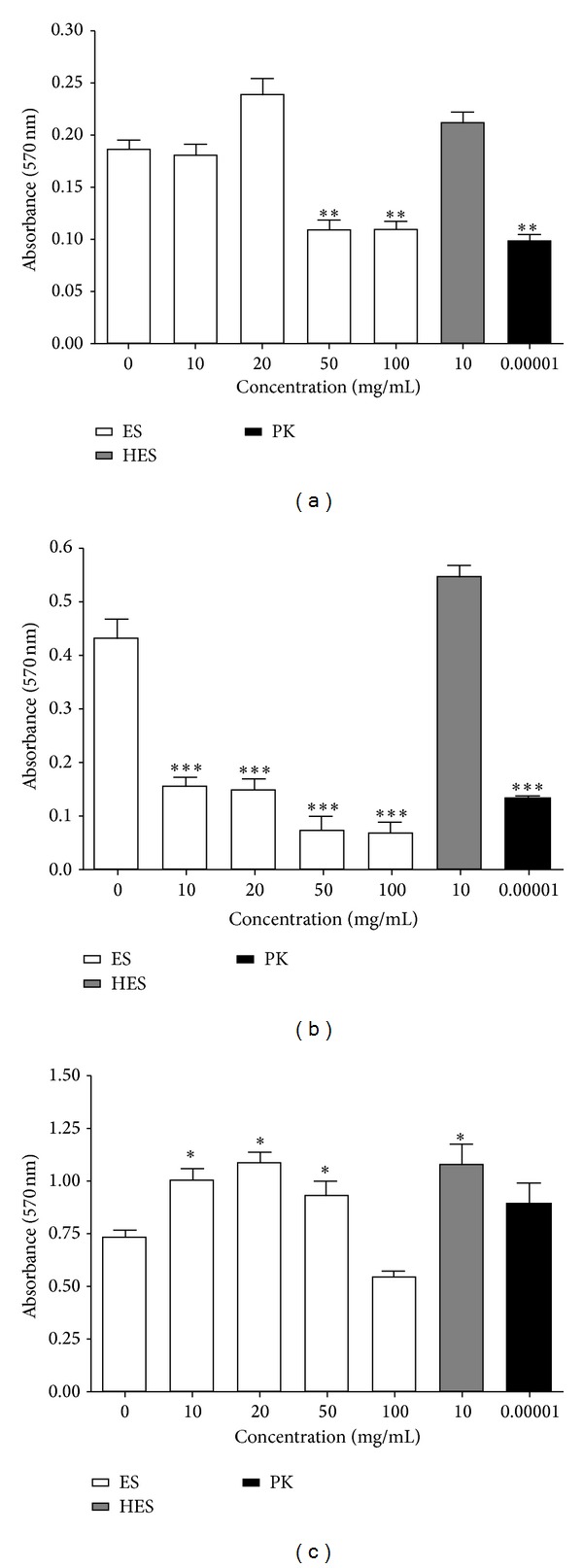
Effects of maggot ES, HES at concentration of 10 mg/mL, and proteinase K (PK) at concentration of 10 mg/mL on established biofilms of (a) Staphylococcus aureus, (b) Enterobacter cloacae, and (c) Proteus mirabilis. Biofilms of wound pathogens were grown in Tryptone Soya Broth medium for 7 h in 96-well plate and treated with maggot ES, HES, and proteinase K for additional 24 h. The values of absorbance are mean SEM of three independent assays. The data were statistically analyzed by the Student's t-test (*P < 0.05).
3.3. Maggot ES Products Kill the Cells within the Bacterial Biofilm
We investigated the maggot ES ability to kill bacteria within the established biofilm of E. cloacae, S. aureus, and P. mirabilis. Already established biofilm was treated with either maggot ES at concentrations of 10, 20, 50, and 100 mg/mL or HES for an additional 17 h. Afterwards we determined the number of viable bacteria (CFU) within the biofilm (Figure 3). Quantification of biofilm formation by CFU enumeration revealed that S. aureus, E. cloacae, and P. mirabilis isolates produced about 107-108 biofilm cells per well. Treatment of S. aureus and P. mirabilis biofilms with maggot ES did not result in a significant decrease in CFU/well values in log units compared with nontreated biofilms. The killing activity of maggot ES was significant only in the case of E. cloacae biofilm; maggot ES decreased the number of viable biofilm cells within a concentration of 100 mg/mL by 10.1% in CFU/well values in log units. Samples treated with heated ES did not show a significant difference in CFU/well values in log units compared with control.
Figure 3.
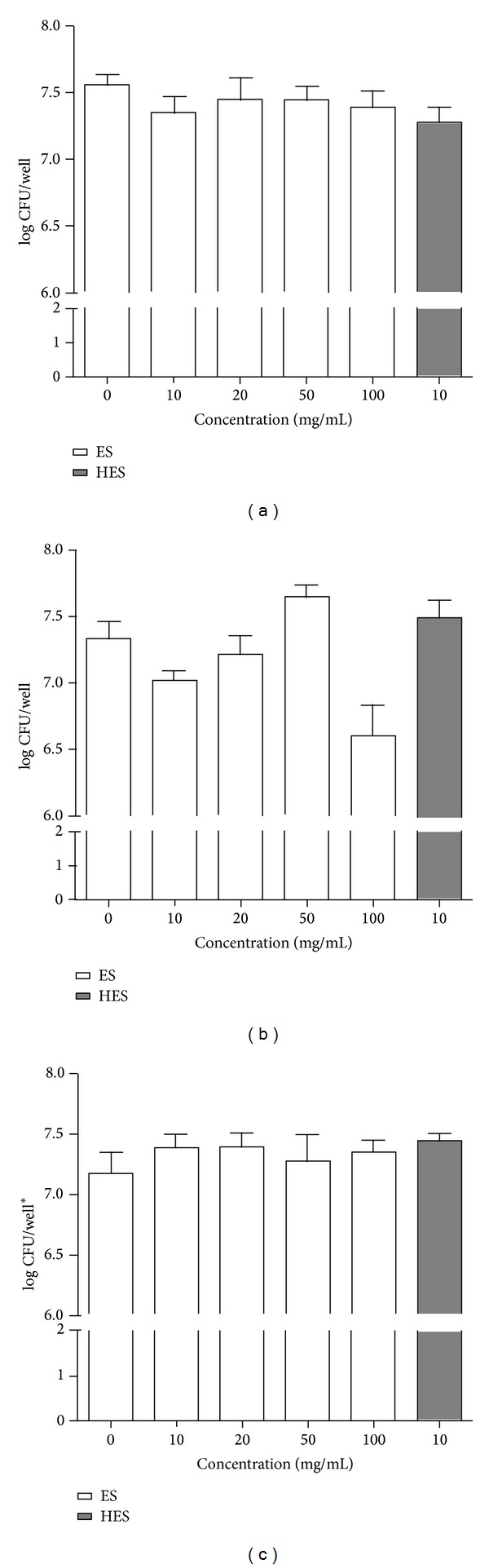
The viable bacteria number in maggot ES and HES treated (a) Staphylococcus aureus, (b) Enterobacter cloacae, and (c) Proteus mirabilis biofilms by log of colony forming unit (CFU) enumeration. The values are number of log CFU per well SEM of three independent assays.
3.4. Determination of Protease Activity in Maggot ES
We investigated the maggot ES proteolytic activity using protease activity assay on milk agar plates. As expected, maggot ES exhibited high proteolytic activity, which could be observed as clear zone by degrading milk proteins. The zone dimension increased with an increasing concentration of maggot ES (Figure 4). There was no visible zone when HES was used. As a positive control we used proteinase K.
Figure 4.
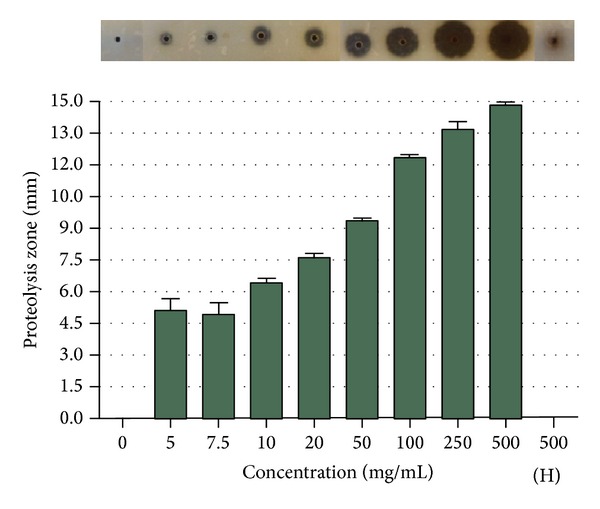
The determination of proteolytic activity in maggot ES products. Maggot ES at concentration range 5–500 mg/mL showed high proteolytic activity which could be observed as a circle zone in the milk agar plate. Heat-treated maggot ES at concentration of 500 mg/mL (500(H)) showed no proteolytic zone.
3.5. Partial Purification of Antibiofilm Compound(s) from Maggot ES Products
The first purification step of maggot antibiofilm compounds was based on the use of SPE-C18 cartridge. The obtained eluate was tested against established biofilm of S. aureus and E. cloacae. The results showed strong antibiofilm activity against matured biofilm. It decreased the S. aureus and E. cloacae biofilm by 74% and 83% in eluate fraction. Afterwards, SPE-C18 eluate was fractionated in HPLC system using C18 column (Figure 5(a)). The obtained fractions were tested for activity against S. aureus established biofilm. The fraction with retention time between 56.6 and 60 min (fraction nos. 15 and 16) degraded matured biofilm by 73%. This active fraction was then fractionated on C4 column employing the same conditions (Figure 5(b)). The fractions with retention time 54–61 min (fractions nos. 15 and 16) significantly degraded S. aureus established biofilm by 76 to 71%. The antibiofilm fractions contained a protein with MW of around 25 kDa (Figure 6).
Figure 5.
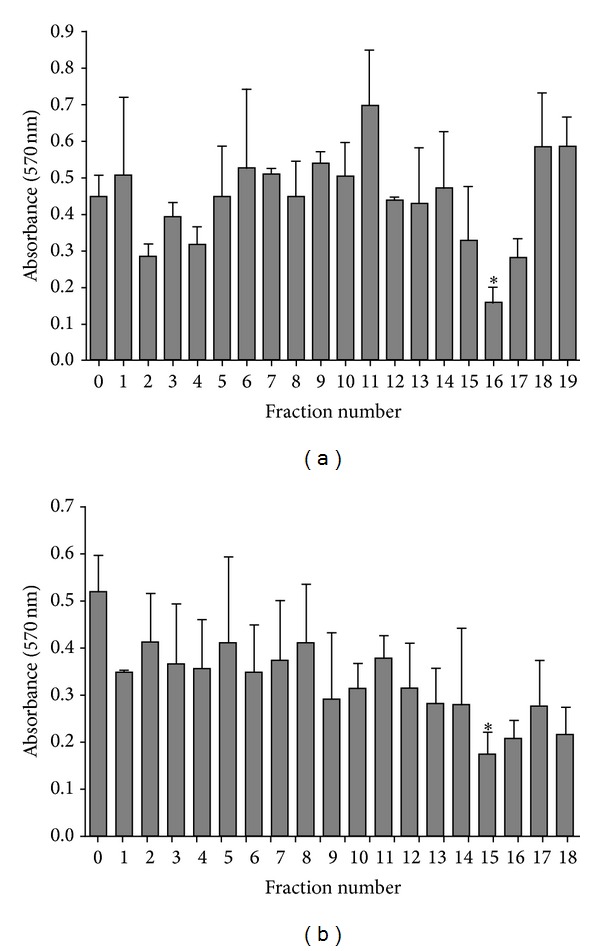
Chromatography fractionation of maggot ES SPE eluate. Maggot ES SPE eluate was fractionated by HPLC system using C18 column (250 mm × 4.6 mm; 5 μm) at a flow rate 0.3 mL/min, by using a gradient from 0 to 90% (v/v) acetonitrile (containing 0.1% (v/v) trifluoroacetic acid), during 70 min (a). Fractions with antibiofilm activity against S. aureus established biofilm were then fractionated by C4 column with the same conditions (b) and checked for its activity against Staphylococcus aureus established biofilm. PC is positive control, established biofilm without ES treatment.
Figure 6.
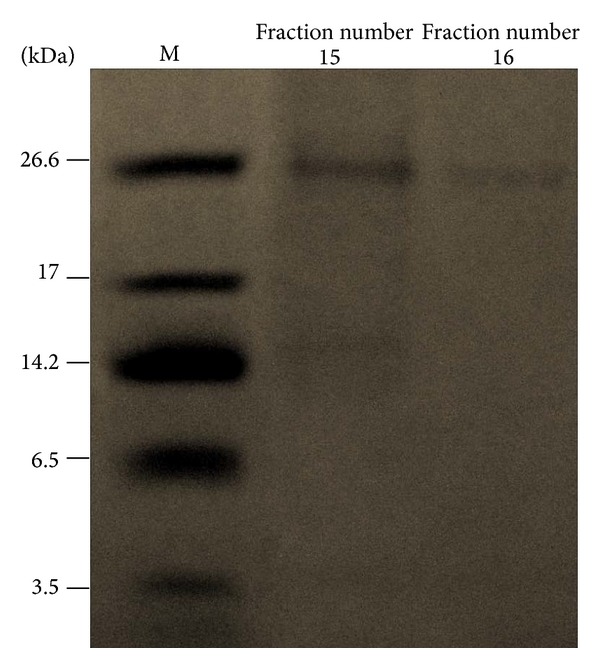
The size (kDa) of partially purified antibiofilm fraction against Staphylococcus aureus isolate. Fraction numbers 15 and 16 were done on Tricine SDS-PAGE. The gels were stained by Coomassie Blue-R. The active band was about 25 kDa in size, present in fraction number 15 and fraction number 16, but in the case of fraction number 16, the band is clearer.
4. Discussion
Bacterial biofilms formed by pathogenic bacteria cause serious troubles in the human health and delay wound healing process [23]. We investigated the antibiofilm activity of maggot ES against wound isolates S. aureus, E. cloacae, and P. mirabilis, some of the most clinically relevant species. Our results suggested that maggot ES prevented biofilm formation of S. aureus and E. cloacae but stimulated biofilm formation of P. mirabilis. Maggot ES treatment of preformed bacterial biofilms revealed differences in the ability to disrupt biofilm and kill the bacteria within S. aureus and E. cloacae biofilms. In case of P. mirabilis, we observed that maggot ES increased biofilm mass and exhibited no effect on cell viability in biofilm. There are a limited number of studies that attempted to elucidate the effect of maggot ES on the ability of bacteria to form biofilm communities and the potential use of ES as an agent to disrupt existing bacterial biofilms [17, 24–27].
Maggot ES are continuously secreted/excreted to the wounds by L. sericata larvae during the MDT. Antibiofilm action of maggot ES depends on the wound characteristics and the wound bacterial environment [28]. Based on our results, maggot ES may either reduce or stimulate biofilm formation in wound depending on the presented bacterial strain. Furthermore, maggot ES-induced reduction and disruption of bacterial biofilm depend on the concentration of active substances.
According to the previous studies of maggot ES antibiofilm activity [17, 26] we confirmed that maggot ES contain substances able to prevent biofilm formation and disrupt established biofilm of S. aureus. In our study, we report strong antibiofilm activity of maggot ES against wound pathogen E. cloacae. Interestingly, maggot ES were more effective in preventing the biofilm formation and dispersing the matured biofilm of E. cloacae than the biofilm of S. aureus. Also, we observed that maggot ES were able to significantly affect the cell viability within the biofilm of E. cloacae while it failed in the case of S. aureus. The present data confirm the fact that maggot ES are differentially effective against different bacterial species. Our results indicate that maggot ES eradicate the bacterial biofilm of different bacterial strains through different mechanisms.
In the present study, maggot ES induced the elimination of S. aureus biofilm mass but the number of viable cells was not significantly decreased. In case of E. cloacae we observed complete disruption of the preformed biofilm and significant decrease of viable cells within the biofilm. It is observed that the antibacterial maggot ES compounds were able to diffuse and kill the bacterial cells through the established biofilm matrix. Heat-treated maggot ES lost antibiofilm activity against E. cloacae but remained in the inhibition of forming S. aureus biofilm. This shifted the attention to heat-labile compounds of maggot ES such as larval proteases.
We found that maggot ES exhibited high proteolytic activity. Proteases play an important role in biofilm regulation of gram-positive and gram-negative bacteria [29]. Maggot ES contain a mix of different extracellular proteases. These proteases play a crucial role in cleaning the wound bed of chronic wounds during MDT [30, 31]. Three groups of proteolytic enzymes (metalloproteinases, serine and aspartyl proteases) were identified in the maggot ES products [30]. In addition, we have very recently identified five novel putative proteases of L. sericata maggots and demonstrated that they could be secreted into the wound during the MDT [32].
The importance of antibiofilm properties of heat sensitive compounds within maggot ES which may be responsible for breaking down S. aureus biofilm was discussed in previous studies [20]. Brown and coworkers [18] found that nucleases present in maggot ES can digest DNA associated with P. aeruginosa biofilm formation. In a very recent study it has been presented that maggots chymotrypsin disrupts protein-adhesin mediated biofilm formation of S. aureus [33]. Van der Plas and coworkers [27] indicated that potential molecules in maggot ES responsible for breaking down S. aureus biofilm are proteases belonging to the group of serine proteases. Harris and coworkers identified a molecule with antibiofilm activity against S. epidermidis biofilm; the particular responsible molecule(s) was >10 kDa in size and appeared to have protease or glucosaminidase activity [25]. In the present paper we also partially purified a compound of maggot ES with antibiofilm properties against S. aureus isolate with a size of around 25 kDa. However, the exact structure of the molecules responsible for the antibiofilm properties of maggot ES is yet unknown.
Interestingly, maggot ES in particular concentrations stimulated biofilm formation as well as significantly increased the biomass of P. mirabilis preformed biofilm. The number of viable cells within P. mirabilis biofilm was not affected after maggot ES treatment. P. mirabilis is a commensal organism living in digestive system of dipteran larval stage [34]. This symbiotic bacterium adheres to peritrophic membrane with the help of lectins and it is important for maggots surviving in their natural pathogenic environment [35]. The contaminated food is easily ingested and microorganisms are rapidly eliminated due to interaction with substances produced by P. mirabilis [36]. One of the beneficial substances produced by P. mirabilis is mirabilicide [37] and other diverse proteolytic enzymes [25, 38, 39]. These compounds contribute to protect the larvae from other harmful bacteria in food intake [36]. This relationship may explain why maggot ES did not show any antibiofilm activity against P. mirabilis.
In conclusion, maggot ES products were shown to be effective in the reduction of biofilm formation and the eradication of established biofilms of E. cloacae as well as S. aureus wound pathogens. The antibiofilm effects of maggot ES are mainly mediated by proteases. On the other hand, maggot ES protected or stimulated P. mirabilis biofilm formation. Therefore, MDT could be used as a potential therapy for the treatment of wounds containing E. cloacae and S. aureus but not for P. mirabilis.
Acknowledgments
This work was supported by the Operational Program Research and Development and was cofinanced by the European Fund for Regional Development (EFRD), Grant: ITMS 26240220020—“Establishment of biotherapeutic facility and technology proposal for production and development of bio-drugs.”
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.James GA, Swogger E, Wolcott R, et al. Biofilms in chronic wounds. Wound Repair and Regeneration. 2008;16(1):37–44. doi: 10.1111/j.1524-475X.2007.00321.x. [DOI] [PubMed] [Google Scholar]
- 2.Kirketerp-Møller K, Jensen PØ, Fazli M, et al. Distribution, organization, and ecology of bacteria in chronic wounds. Journal of Clinical Microbiology. 2008;46(8):2717–2722. doi: 10.1128/JCM.00501-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bjarnsholt T, Kirketerp-Møller K, Jensen PØ, et al. Why chronic wounds will not heal: a novel hypothesis. Wound Repair and Regeneration. 2008;16(1):2–10. doi: 10.1111/j.1524-475X.2007.00283.x. [DOI] [PubMed] [Google Scholar]
- 4.Bjarnsholt T, Jensen PØ, Fiandaca MJ, et al. Pseudomonas aeruginosa biofilms in the respiratory tract of cystic fibrosis patients. Pediatric Pulmonology. 2009;44(6):547–558. doi: 10.1002/ppul.21011. [DOI] [PubMed] [Google Scholar]
- 5.Bjarnsholt T, Tolker-Nielsen T, Givskov M, Janssen M, Christensen LH. Detection of bacteria by fluorescence in situ hybridization in culture-negative soft tissue filler lesions. Dermatologic Surgery. 2009;35(2):1620–1623. doi: 10.1111/j.1524-4725.2009.01313.x. [DOI] [PubMed] [Google Scholar]
- 6.Homøe P, Bjarnsholt T, Wessman M, Sørensen HCF, Johansen HK. Morphological evidence of biofilm formation in Greenlanders with chronic suppurative otitis media. European Archives of Oto-Rhino-Laryngology. 2009;266(10):1533–1538. doi: 10.1007/s00405-009-0940-9. [DOI] [PubMed] [Google Scholar]
- 7.Burmølle M, Thomsen TR, Fazli M, et al. Biofilms in chronic infections—a matter of opportunity—monospecies biofilms in multispecies infections. FEMS Immunology & Medical Microbiology. 2010;59(3):324–336. doi: 10.1111/j.1574-695X.2010.00714.x. [DOI] [PubMed] [Google Scholar]
- 8.Davis SC, Ricotti C, Cazzaniga A, Welsh E, Eaglstein WH, Mertz PM. Microscopic and physiologic evidence for biofilm-associated wound colonization in vivo. Wound Repair and Regeneration. 2008;16(1):23–29. doi: 10.1111/j.1524-475X.2007.00303.x. [DOI] [PubMed] [Google Scholar]
- 9.Dowd SE, Sun Y, Secor PR, et al. Survey of bacterial diversity in chronic wounds using Pyrosequencing, DGGE, and full ribosome shotgun sequencing. BMC Microbiology. 2008;8, article 43 doi: 10.1186/1471-2180-8-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Frank DN, Wysocki A, Specht-Glick DD, et al. Microbial diversity in chronic open wounds. Wound Repair and Regeneration. 2009;17(2):163–172. doi: 10.1111/j.1524-475X.2009.00472.x. [DOI] [PubMed] [Google Scholar]
- 11.Price LB, Liu CM, Melendez JH, et al. Community analysis of chronic wound bacteria using 16S rRNA gene-based pyrosequencing: impact of diabetes and antibiotics on chronic wound microbiota. PLoS ONE. 2009;4(7) doi: 10.1371/journal.pone.0006462.e6462 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Martin JM, Zenilman JM, Lazarus GS. Molecular microbiology: new dimensions for cutaneous biology and wound healing. Journal of Investigative Dermatology. 2010;130(1):38–48. doi: 10.1038/jid.2009.221. [DOI] [PubMed] [Google Scholar]
- 13.Gjødsbøl K, Christensen JJ, Karlsmark T, Jørgensen B, Klein BM, Krogfelt KA. Multiple bacterial species reside in chronic wounds: a longitudinal study. International Wound Journal. 2006;3(3):225–231. doi: 10.1111/j.1742-481X.2006.00159.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rickard AH, Gilbert P, High NJ, Kolenbrander PE, Handley PS. Bacterial coaggregation: an integral process in the development of multi-species biofilms. Trends in Microbiology. 2003;11(2):94–100. doi: 10.1016/s0966-842x(02)00034-3. [DOI] [PubMed] [Google Scholar]
- 15.Sherman RA. Maggot versus conservative debridement therapy for the treatment of pressure ulcers. Wound Repair and Regeneration. 2002;10(4):208–214. doi: 10.1046/j.1524-475x.2002.10403.x. [DOI] [PubMed] [Google Scholar]
- 16.Dumville JC, Worthy G, Bland JM, et al. Larval therapy for leg ulcers (VenUS II): randomised controlled trial. British Medical Journal. 2009;338, article b773 doi: 10.1136/bmj.b773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.van der Plas MJA, Jukema GN, Wai SW, et al. Maggot excretions/secretions are differentially effective against biofilms of Staphylococcus aureus and Pseudomonas aeruginosa . Journal of Antimicrobial Chemotherapy. 2008;61(1):117–122. doi: 10.1093/jac/dkm407. [DOI] [PubMed] [Google Scholar]
- 18.Brown A, Horobin A, Blount DG, et al. Blow fly Lucilia sericata nuclease digests DNA associated with wound slough/eschar and with Pseudomonas aeruginosa biofilm. Medicinal and Veterinary Entomology. 2012;26(4):432–439. doi: 10.1111/j.1365-2915.2012.01029.x. [DOI] [PubMed] [Google Scholar]
- 19.Jaklic D, Lapanje A, Zupancic K, Smrke D, Gunde-Cimerman N. Selective antimicrobial activity of maggots against pathogenic bacteria. Journal of Medicinal Microbiogy. 2008;57, part 5:617–625. doi: 10.1099/jmm.0.47515-0. [DOI] [PubMed] [Google Scholar]
- 20.van der Plas MJ, van der Does AM, Baldry M, et al. Maggot excretions/secretions inhibit multiple neutrophil pro-inflammatory responses. Microbes and Infection. 2007;9(4):507–514. doi: 10.1016/j.micinf.2007.01.008. [DOI] [PubMed] [Google Scholar]
- 21.Stepanović S, Vuković D, Hola V, et al. Quantification of biofilm in microtiter plates: overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Acta Pathologica, Microbiologica, et Immunologica Scandinavica. 2007;115(8):891–899. doi: 10.1111/j.1600-0463.2007.apm_630.x. [DOI] [PubMed] [Google Scholar]
- 22.Quiblier C, Zinkernagel AS, Schuepbach RA, Berger-Bächi B, Senn MM. Contribution of SecDF to Staphylococcus aureus resistance and expression of virulence factors. BMC Microbiology. 2011;11, article 72 doi: 10.1186/1471-2180-11-72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Saito ST, Trentin DS, Macedo AJ, et al. Bioguided fractionation shows Cassia alata extract to inhibit Staphylococcus epidermidis and Pseudomonas aeruginosa growth and biofilm formation. Evidence-Based Complementary and Alternative Medicine. 2012;2012:13 pages. doi: 10.1155/2012/867103.867103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cazander G, van Veen KE, Bouwman LH, Bernards AT, Jukema GN. The influence of maggot excretions on PAO1 biofilm formation on different biomaterials. Clinical Orthopaedics and Related Research. 2009;467(2):536–545. doi: 10.1007/s11999-008-0555-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Harris LG, Bexfield A, Nigam Y, Rohde H, Ratcliffe NA, Mack D. Disruption of Staphylococcus epidermidis biofilms by medicinal maggot Lucilia sericata excretions/secretions. International Journal of Artificial Organs. 2009;32(9):555–564. doi: 10.1177/039139880903200904. [DOI] [PubMed] [Google Scholar]
- 26.Cazander G, van de Veerdonk MC, Vandenbroucke-Grauls CMJE, Schreurs MWJ, Jukema GN. Maggot excretions inhibit biofilm formation on biomaterials. Clinical Orthopaedics and Related Research. 2010;468(10):2789–2796. doi: 10.1007/s11999-010-1309-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.van der Plas MJA, Dambrot C, Dogterom-Ballering HCM, Kruithof S, van Dissel JT, Nibbering PH. Combinations of maggot excretions/secretions and antibiotics are effective against Staphylococcus aureus biofilms and the bacteria derived therefrom. Journal of Antimicrobial Chemotherapy. 2010;65(5):917–923. doi: 10.1093/jac/dkq042. [DOI] [PubMed] [Google Scholar]
- 28.Jiang KC, Sun XJ, Wang W, et al. Excretions/secretions from bacteria-pretreated maggot are more effective against Pseudomonas aeruginosa biofilms. PLoS ONE. 2012;7(11) doi: 10.1371/journal.pone.0049815.e49815 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Park JH, Lee JH, Cho MH, Herzberg M, Lee J. Acceleration of protease effect on Staphylococcus aureus biofilm dispersal. FEMS Microbiology Letters. 2012;335(1):31–38. doi: 10.1111/j.1574-6968.2012.02635.x. [DOI] [PubMed] [Google Scholar]
- 30.Chambers L, Woodrow S, Brown AP, et al. Degradation of extracellular matrix components by defined proteinases from the greenbottle larva Lucilia sericata used for the clinical debridement of non-healing wounds. The British Journal of Dermatology. 2003;148(1):14–23. doi: 10.1046/j.1365-2133.2003.04935.x. [DOI] [PubMed] [Google Scholar]
- 31.Telford G, Brown AP, Seabra RAM, et al. Degradation of eschar from venous leg ulcers using a recombinant chymotrypsin from Lucilia sericata . The British Journal of Dermatology. 2010;163(3):523–531. doi: 10.1111/j.1365-2133.2010.09854.x. [DOI] [PubMed] [Google Scholar]
- 32.Valachova I, Majtan T, Takac P, Majtan J. Identification and characterisation of different proteases in Lucilia sericata medicinal maggots involved in maggot debridement therapy. Journal of Applied Biomedicine. 2014 [Google Scholar]
- 33.Harris LG, Nigam Y, Sawyer J, Mack D, Pritchard DI. Lucilia sericata chymotrypsin disrupts protein adhesin-mediated staphylococcal biofilm formation. Applied and Environmental Microbiology. 2013;79(4):1393–1395. doi: 10.1128/AEM.03689-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Greenberg B, Kowalski JA, Klowden MJ. Factors affecting the transmission of salmonella by flies: natural resistance to colonization and bacterial interference. Infection and Immunity. 1970;2(6):800–809. doi: 10.1128/iai.2.6.800-809.1970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Peters W, Kolb H, Kolb-Bachofen V. Evidence for a sugar receptor (lectin) in the peritrophic membrane of the blowfly larva, Calliphora erythrocephala Mg. (Diptera) Journal of Insect Physiology. 1983;29(3):275–280. [Google Scholar]
- 36.Erdmann GR, Khalil SKW. Isolation and identification of two antibacterial agents produced by a strain of Proteus mirabilis isolated from larvae of the screwworm (Cochliomyia hominivorax) (Diptera: Calliphoridae) Journal of Medical Entomology. 1986;23(2):208–211. doi: 10.1093/jmedent/23.2.208. [DOI] [PubMed] [Google Scholar]
- 37.Greenberg B. Model for destruction of bacteria in the midgut of blow fly maggots. Journal of Medical Entomology. 1968;5(1):31–38. doi: 10.1093/jmedent/5.1.31. [DOI] [PubMed] [Google Scholar]
- 38.Hobson RP. On an enzyme from blow-fly larvre Lucilia sericata which digests collagen in alkaline solution. The Biochemical Journal. 1931;25(5):1458–1463. doi: 10.1042/bj0251458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ziffren SE, Heist HE, May SC, Womack NA. The secretion of collagenase by maggots and its implication. Annals of surgery. 1953;138(6):932–934. doi: 10.1097/00000658-195312000-00023. [DOI] [PMC free article] [PubMed] [Google Scholar]