

Abstract
Differentiation of erythroblasts to mature red blood cells involves dynamic changes of the membrane and cytoskeleton networks that are not fully characterized. Using a mouse fetal liver erythroblast culture system and a targeted shRNA functional screening strategy, we identified a critical role of pleckstrin-2 in actin dynamics and protection of early stage terminal erythroblasts from oxidative damage. Knockdown of pleckstrin-2 in the early stage of terminal erythropoiesis disrupted the actin cytoskeleton and led to differentiation inhibition and apoptosis. This pro-survival and differentiation function of pleckstrin-2 was mediated through its interaction with cofilin, by preventing cofilin’s mitochondrial entry when the intracellular level of reactive oxygen species was higher in the early stage of terminal erythropoiesis. Treatment of the cells with a scavenger of reactive oxygen species rescued cofilin’s mitochondrial entry and differentiation inhibition induced by pleckstrin-2 knockdown. In contrast, pleckstrin-2 knockdown in late stage terminal erythroblasts had no effect on survival or differentiation but blocked enucleation due to disorganized actin cytoskeleton. Thus, our study identified a dual function of pleckstrin-2 in the early and late stages of terminal erythropoiesis through its regulations of actin dynamics and cofilin’s mitochondrial localization, which reflects intracellular level of reactive oxygen species in different developmental stages.
Introduction
Erythropoiesis is the process of differentiation of hematopoietic stem cells to mature erythrocytes. This stepwise process includes the formation of committed erythroid burst-forming units (BFU-Es) followed by rapidly dividing erythroid colony-forming units (CFU-Es). Differentiation from CFU-Es to mature red blood cells, generally termed terminal erythropoiesis, is driven by multiple erythropoietin (Epo)-induced signaling transduction pathways.1 Epo and its receptor EpoR are essential for terminal erythropoiesis. Binding of Epo to EpoR triggers the activation of Jak2, which is followed by the induction of several signal transduction cascades, including Stat5, PI3K/Akt, and Ras-MAP kinase pathways.2–4 Mice with genetic deletion of Epo or EpoR die at embryonic Day 13 (E13) with severe defects in erythropoiesis even though normal numbers of CFU-Es are present.5
During the late stage of terminal erythropoiesis, erythroblasts undergo terminal cell cycle exit, chromatin condensation, and extrusion of nuclei. Recent studies revealed that multiple signaling pathways are involved in the generation of enucleated erythroid cells.6,7 These include histone deacetylation,8,9 actin cytoskeleton,10–13 cytokinesis,14,15 cell-matrix interactions,16 specific microRNAs17 and vesicle trafficking.18
Many of these studies used a mouse fetal liver erythroblast culture system, which makes it possible to analyze erythroid cell differentiation and enucleation both quantitatively and step by step.19,20 In this system, erythroid progenitor cells from mouse fetal liver can easily be isolated by a single-step purification of TER119 (marker for the mature erythroid cells) negative cells. These progenitor cells can be further studied in an in vitro culture system that supports their normal proliferation and differentiation. During the 2-day culture, cell differentiation can be monitored by a flow cytometric analysis based on the surface expression of the transferrin receptor CD71 and the TER119 antigen. In addition, enucleation of the terminally differentiated erythroblasts can be detected by a DNA marker Hoechst 33342 on Day 2. Thus, the extruded nuclei and incipient reticulocytes can be further analyzed in vitro, given that this is impossible in vivo since nuclei are rapidly engulfed by macrophages.21
With the same fetal liver erythroblast culture system, a recent report using next generation sequencing identified that the expression of nearly 500 genes increased greater than 2-fold during terminal erythropoiesis.22 Many of these genes encode proteins that are important for erythroid cells such as α and β globins, heme biosynthetic enzymes, erythroid membrane and cytoskeleton proteins, as well as erythroid transcription factors. However, the roles that many of the up-regulated genes play in erythroid cells are still unknown. To identify and characterize genes that have novel functions in different stages of terminal erythropoiesis, we used the mouse fetal liver erythroblast in vitro culture technology together with a targeted array-based high throughput screening system and discovered more than 30 genes that have novel functions in the early and late stages of terminal erythropoiesis. Among these genes, we identified pleckstrin-2 (plek2), which was previously reported to be involved in T-cell cytoskeleton reorganization,23 play critical roles in terminal erythropoiesis. We demonstrated that plek2 is important in the regulation of actin cytoskeleton, cell differentiation and apoptosis. We also found that plek2 blocks apoptosis in the early stage of terminal erythropoiesis through preventing cofilin’s mitochondrial entry. Furthermore, we dissected these functions of plek2 in the early and late stages of terminal erythropoiesis reflecting different intracellular levels of reactive oxygen species (ROS).
Methods
Materials
Rabbit polyclonal antibody against plek2 was purchased from Proteintech. Rabbit polyclonal antibodies against cofilin and phospho-cofilin were purchased from Cell Signaling Technology. Monoclonal antibody against Hsc70 was purchased from Santa Cruz Biotechnology. All the antibodies for flow cytometric analysis were purchased from eBioscience. Alexa Fluor 488 phalloidin and Alexa Fluor 544 for immunofluorescence were purchased from Invitrogen Molecular Probes. The Hairpin-pLKO.1 lentiviruses were from the Broad Institute (Cambridge, MA, USA).
Purification and culture of fetal liver cells for targeted screening
Purification of mouse fetal liver erythroblast progenitors (CFU-E; TER119-negative cells) was based on previously described procedures.10 Details of procedures including viral transduction and the screening protocol are available in the Online Supplementary Appendix.
Retroviral shRNA construction
Retroviral shRNA oligonucleotides against plek2 were designed using an automated shRNA selection web server (http://jura.wi.mit.edu/bioc/siRNAext/home.php).24 Cloning of shRNA into MSCV-U3-H1 was performed as previously described.25 The shRNA nucleotide sequences used in the experiments are as follows. Non-targeting shRNA: 5′-GGACTAGTCTCCACGGTGA-3′. Plek2 shRNA: 5′ – CAACTCAGTATCTCTGGGA – 3′.
Flow cytometric analysis and immunoprecipitation
Flow cytometric analysis of differentiation and enucleation of cultured mouse fetal erythroblasts was as previously described.10 Details of the protocol for immunoprecipitation are available in the Online Supplementary Appendix.
Immunofluorescence stain and microscopic analysis
Details of the protocol for immunofluorescence stain and microscopic analysis are available in the Online Supplementary Appendix.
Quantitative analysis of intracellular reactive oxygen species
The intracellular ROS levels were determined using CM-H2DCFDA (Molecular Probes). Briefly, erythroid cells were incubated with 10 μM CM-H2DCFDA or DMSO control. After incubation for 30 min at 37°C, the cells were harvested, washed, and resuspended in PBS. The levels of fluorescence were immediately detected using flow cytometric analysis.
Results
Targeted shRNA screening identified genes that play novel functions in the early and late stages of terminal erythropoiesis
To identify genes that play novel functions in different stages of terminal erythropoiesis, we chose 100 genes (Online Supplementary Table S1) that were most abundantly expressed during differentiation in a recent genetic study,22 but that have not been studied in erythroid cells, for an shRNA-based targeted functional screening assay. TER119-negative mouse fetal liver erythroblasts from E13.5 were used for the screening culture.
Since there are significant differences in terms of differentiation, cell cycle status and morphology between the early and late stages of terminal erythropoiesis, we employed a unique screening strategy to identify genes that have specific roles in different stages of terminal erythropoiesis (Figure 1A). This consists of two independent conditions to test candidate gene function in the early and late stages of terminal erythropoiesis. For the early stage of terminal erythropoiesis (Figure 1A, dashed lines), TER119-negative fetal liver erythroblasts were purified. These cells showed minimal apoptosis and most were CD117-positive (Online Supplementary Figure S1A). After lentiviral shRNA transduction, the cells were cultured for 12 h in Epo-free medium but containing stem cell factor (SCF), interleukin-6 and FLT3 ligand to maintain their progenitor status, maintain cell survival, and allow expression of the infected shRNA.26 After 12 h, these cells underwent mild downregulation of CD117 with minimal apoptosis and remained TER119-negative (Online Supplementary Figure S1B). We call this SCF medium. The cells were then transferred to Epo-containing media with puromycin to select the transduced cells, which were continually cultured for two days. During the 2-day culture in Epo medium, the TER119-negative cells gradually became negative for CD117 and positive for CD71 and TER119 with the presence of Hoechst 33342 low reticulocytes (Online Supplementary Figure S1B). To test genes in the late stage of terminal erythropoiesis (Figure 1A, solid lines), the TER119-negative cells were plated in Epo-containing media immediately after infection, added with puromycin after 12 h, and further cultured for two days. The differentiation and enucleation profiles of these cells after 60 h were similar to cells cultured in SCF medium before Epo medium (Online Supplementary Figure S1C). In both culture conditions, the TER119-negative erythroblasts underwent some degree of differentiation after 12 h, as judged by CD71 expression. As expected, this differentiation was more prominent in those cells cultured immediately in Epo conditions. Both screenings were carried out with E13.5 TER119-negative mouse fetal liver erythroblasts in 96-well plates. Five different shRNAs were designed for each gene to mitigate spurious gene identification due to off-target effects.27
Figure 1.
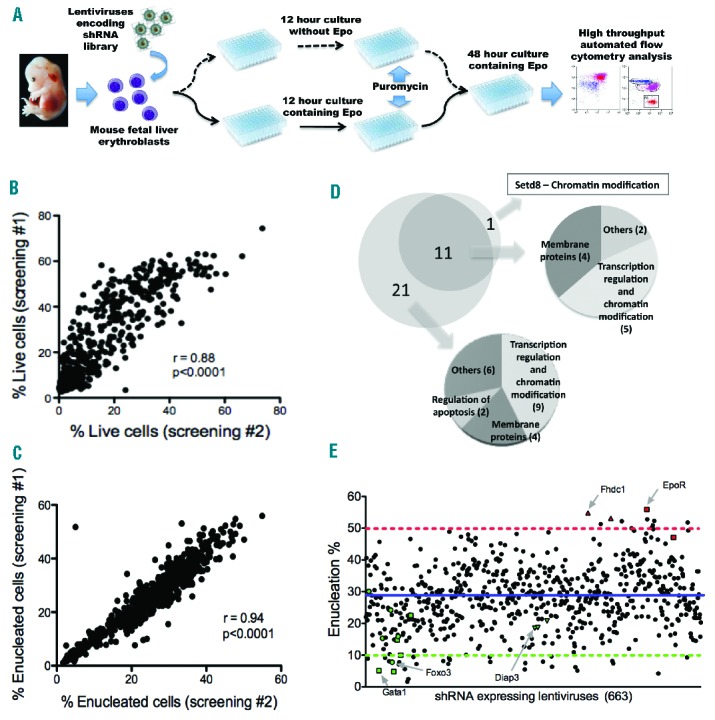
Targeted shRNA screening identified genes that play novel functions in terminal erythropoiesis. (A) A schematic overview of the screening strategies. Dashed and solid lines represent the screening strategies for the early and late stages of terminal erythropoiesis, respectively. (B–C) Correlation coefficient of duplicate sets of screens for the dashed (B) and solid (C) line methods. (D) Pie graph of hit genes and their functions in other types of cells based on DAVID bioinformatics resources.28,29 (E) Summary of flow cytometric analysis of enucleation screening. Each dot indicates the enucleation rate of cells transduced with single distinct shRNA encoding lentiviruses. Control genes (green) and genes that gave higher enucleation rate after knockdown (red) are highlighted in arrow. The blue line indicates mean enucleation rate. The dotted red and green lines indicate the upper and lower limits for the selection of hit genes, respectively.
ShRNA against luciferase was used as a general positive control in which approximately 40% of the cells were transduced that survived after puromycin treatment (Online Supplementary Figure S2A, left panel). The transduced cells showed TER119-positivity at the end of the culture since no endogenous genes were down-regulated (Online Supplementary Figure S2A, right panel). Cells without lentiviral infection were used as a general negative control in which all the cells underwent apoptosis after 2.5 days due to the lack of transduction (Online Supplementary Figure S2B). To validate the screening system, a group of genes with well-known functions in different stages of terminal erythropoiesis were used as functional controls. For the screening of the early stage of terminal erythropoiesis, we used GATA1, which is a transcription factor in erythropoiesis, as a control. Cells transduced with lentiviral shRNA against GATA1 mimicked those without lentiviral transduction in which most of the cells died after 2.5 days of culture (Online Supplementary Figure S2C, left panel). Less than 5% of the cells survived, which made the analysis of cell differentiation unmeaningful (Online Supplementary Figure S2C, right panel). Therefore, we used apoptosis as the readout for this screening method. For the screening of genes that function in the late stage of terminal erythropoiesis, cells transduced with lentiviruses encoding shRNA against luciferase showed approximately 40% enucleation rate in the 96-well plate screening condition (Online Supplementary Figure S2D, left panel). The enucleation assay was validated with the evidence that enucleated cells were smaller than the non-enucleated late-stage erythroblasts (Online Supplementary Figure S2D, right panel). Similarly, we tested the genes that play well-established roles in terminal erythropoiesis and enucleation, which showed dramatically decreased enucleation rates when they were down-regulated (Online Supplementary Figure S2E). Taken together, these data validated the screening system.
Using these conditions, we performed duplicate sets of screenings for each method with high correlation coefficients (Figure 1B and C). Our criteria to define a hit gene are: 1) for the screening of the early stage of terminal erythropoiesis, at least three unique shRNAs should lead to less than 5% of live cells; 2) for late-stage screening, at least three unique shRNAs should lead to less than 20% of enucleation (Hoechst 33342 low, TER119 high population), with one shRNA less than 10%; 3) for both screenings, the same shRNAs should give a similar phenotype in each of the duplicate experiment. With these criteria, 15 false positive hits (defined by either less than three different shRNAs with the same phenotype or inconsistent results in duplicates) were excluded (data not shown). We putatively identified 32 genes that have novel functions in the early stage of terminal erythropoiesis. These genes are classified into different groups based on their known functions in other cell types (Figure 1D and Table 1). Interestingly, 11 of these genes play dual functions in both early and late stages of terminal erythropoiesis (Figure 1D and Online Supplementary Table S2). In addition, Setd8 plays a unique function only in the late stage of terminal erythropoiesis. Knockdown of the control shRNAs that target luciferase led to an average of 35% enucleation rate (Figure 1E). Surprisingly, knockdown of Fdhc1, one of the formin proteins, and erythropoietin receptor (EpoR) almost doubled the enucleation rate (Figure 1E and Online Supplementary Figure S2F). While it is known that Epo modulates erythroid cell adhesion to bone marrow niche in the late stage of terminal erythropoiesis,30 the EpoR knockdown result suggests an anti-enucleation role for Epo. In fact, while we were optimizing the system, we found that Epo diminished enucleation rate in a dose-dependent manner (Online Supplementary Figure S2G).
Table 1.
List of hit genes with novel functions in terminal erythropoiesis.
Pleckstrin-2 is critical for terminal erythropoiesis
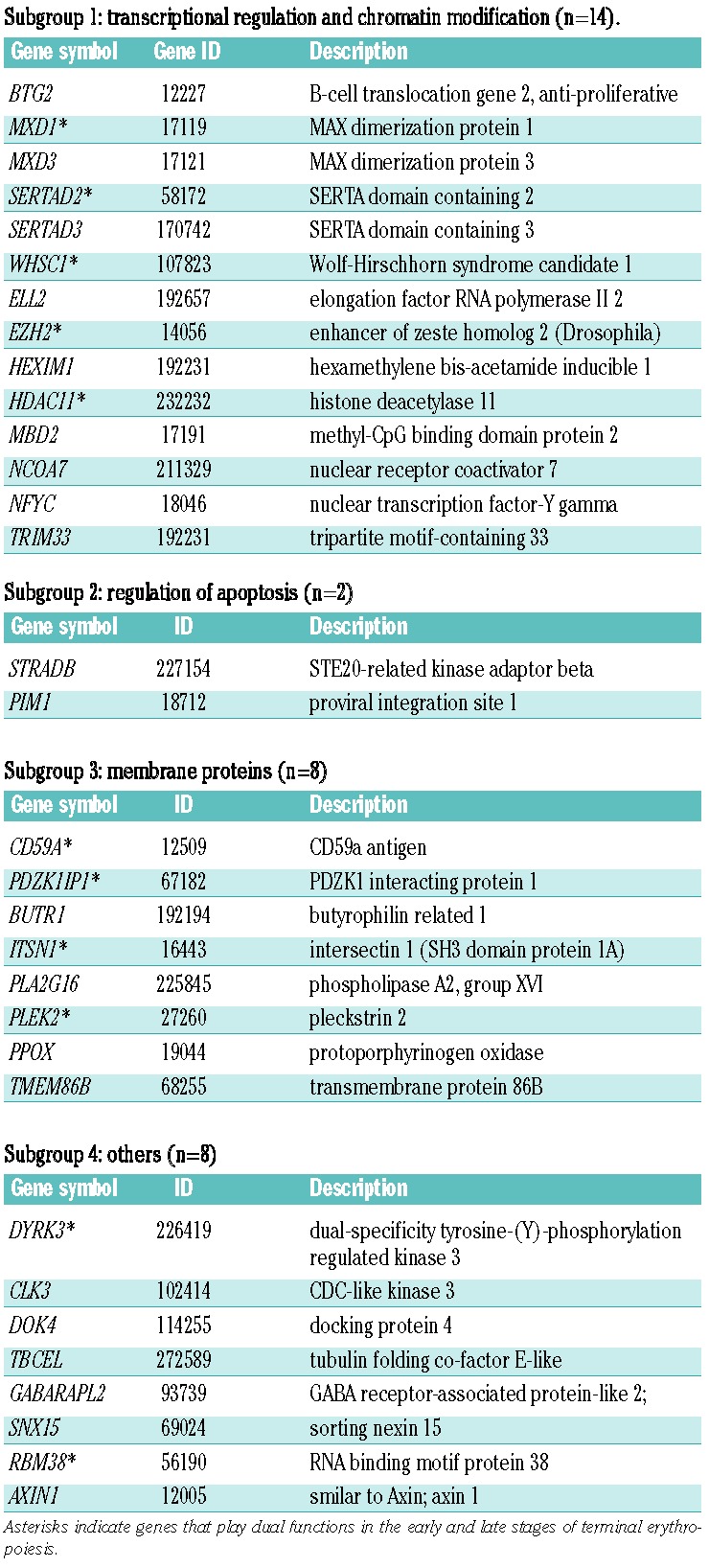
Among the hit genes, pleckstrin-2 (plek2) is particularly interesting since it showed a broad functional impact both in the early and late stages of terminal erythropoiesis. In T cells, plek2 functions in cytoskeleton reorganization,23,31 whereas its roles in other hematopoietic lineages, especially erythroid cell, are completely unknown. During culture of mouse fetal liver erythroblasts, plek2 reached the highest protein levels on Day 1 in culture, and decreased on Day 2 (Figure 2A). A slightly up-shifted band was observed in freshly purified TER119 negative erythroblasts (Day 0), indicating a possible post-translational modification (Figure 2A). Interestingly, plek2 protein reached an undetectable level when cultured in SCF medium for 12 h after purification, but regained strong expression after switching to Epo medium (Figure 2B). This indicates that Epo signaling pathways could control plek2 expression. In order to further study the mechanism of plek2 in erythropoiesis, we generated an independent shRNA encoded by a murine stem cell retroviral vector (MSCV), which has a stable and high level infection efficiency in the mouse hematopoietic cells.32 It efficiently down-regulated plek2, in contrast to the vector control or a non-targeting shRNA (Figure 2C and D). With this shRNA, downregulation of plek2 in the early stage of terminal erythropoiesis (Figure 1A, dashed line) led to a dramatic reduction in total cell number (Figure 2E). Flow cytometric analysis demonstrated a significant inhibition of differentiation (Figure 2F) and subsequent enucleation (Figure 2G), which was accompanied by a significant cell death (Figure 2H). Cell cycle analysis further confirmed apoptosis with an increased sub-G1 population after plek2 knockdown. However, other phases of cell cycle were not significantly affected (Online Supplementary Figure S3A). To determine whether these phenotypes are indeed due to plek2 deficiency rather than off-target effects of the shRNA, we rescued the differentiation inhibition and apoptosis by co-infecting a non-targeting plek2 over-expressing retrovirus (Online Supplementary Figure S3B–D).
Figure 2.
Plek2 is required for the early stage of terminal erythropoiesis. (A) Western blot analysis of plek2 on different days of cultured mouse fetal liver erythroblasts in Epo medium together with TER119 positive fetal liver cells. Hsc70 was used as a loading control. (B) Western blot analysis of plek2 on freshly purified TER119 negative fetal liver erythroblasts, cells cultured in SCF medium for 12 h, and cells cultured in SCF medium for 12 h followed by 24-h Epo medium culture. Hsc70 was used as a loading control. (C) TER119 negative fetal liver erythroblasts were transduced with retroviruses encoding the indicated shRNAs and cultured in SCF medium for 12 h. The cells were changed to Epo medium and continued to culture for 12 h followed by Western blot analysis of plek2. Nt-shRNA represents a non-targeting shRNA. Hsc70 was used as a loading control. (D) Quantification of (C). Data were obtained from 3 independent experiments. (E) Cell growth and quantity analysis after knockdown by indicated shRNAs. Same as C except that the cells were cultured for two more days after change to Epo medium. The cells were counted using a hemocytometer on each day. (F) Flow cytometric analysis of TER119 level in the indicated cells from (E). Right bar graph is the statistical analysis using Student’s t-test. *** P<0.001. (G–H) Analysis of mouse fetal erythroblast enucleation and apoptosis during in vitro culture. Same as (E), the cells after culture were analyzed by flow cytometric analysis for enucleation (G) and apoptosis (H) using Hoechst 33342 and TER119, and propidium iodide (PI) and Annexin V, respectively. Statistical analysis using Student’s t-test is on the right. ***P<0.001.
Plek2 is required for the integrity of actin cytoskeleton in the early stage of terminal erythropoiesis
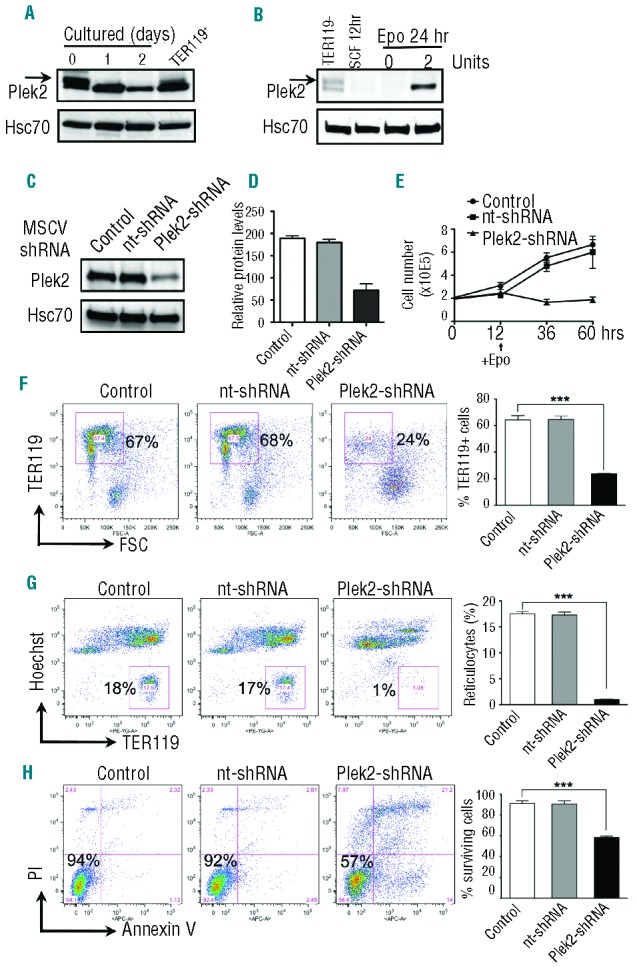
Since plek2 was reported to be membrane bound23 with a putative function in actin rearrangement in other cell types,31 we next tested the sub-cellular localization of plek2 in the cultured mouse fetal liver erythroblasts with confocal microscopy. Consistent with previous reports, plek2 localized primarily on the plasma membrane in the early stage of terminal erythropoiesis (Figure 3A). Plek2 partially co-localized with actin filaments on Day 1 and Day 2 of the cells in Epo medium, suggesting a role for plek2 in actin cytoskeleton regulation (Figure 3A). The actin cytoskeleton network plays critical roles in cell differentiation. Disruption of actin cytoskeleton leads to apoptosis in many cell types,33 including erythroid cells.10,34,35 We hypothesized that the plek2 knockdown phenotypes in early stage terminal erythroblasts could be due to the disruption of actin cytoskeleton. To test this, we analyzed actin localization when plek2 was down-regulated in the early stage approximately 12 h after Epo exposure and before apoptosis was observed. As expected, knockdown of plek2 dramatically abolished actin localization on plasma membrane (Figure 3B).
Figure 3.
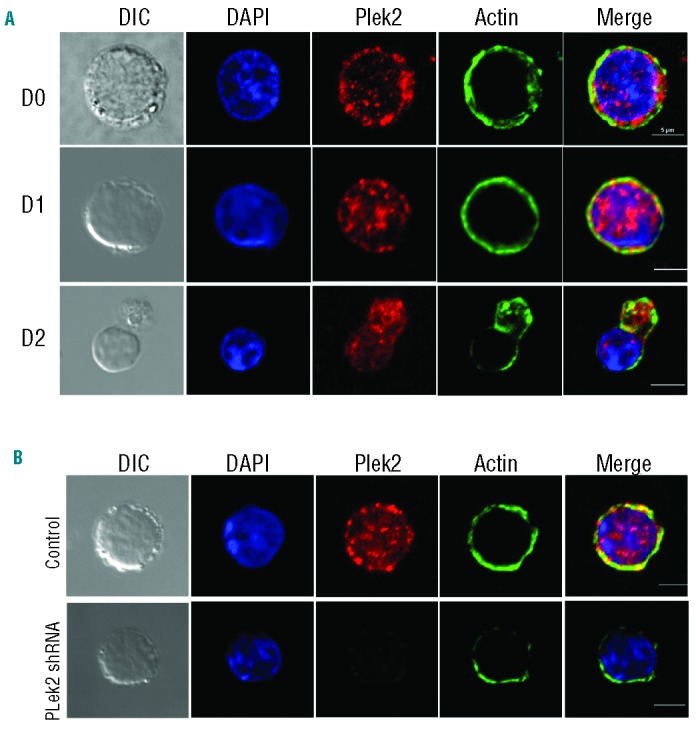
Plek2 is required for actin cytoskeleton integrity of mouse fetal liver erythroblasts. (A) Confocal immunofluorescence microscopy of actin (green) and plek2 (red) localizations on different days of Epo medium cultured mouse fetal liver erythroblasts. Blue is for DAPI DNA stain. Scale bar: 5 μm. (B) Confocal microscopy of actin and plek2 localizations in control (upper panels) and plek2 knockdown (lower panels) mouse fetal liver erythroblasts. The cells were transduced with indicated MSCV retroviruses and cultured in SCF medium for 12 h, which was followed by Epo medium culture for 12 h before being fixed for staining. Scale bar: 5 μm. The confocal images were representative optical sections of the cell from at least 50 cells in each condition in 3 different experiments.
Plek2 interacts with cofilin to prevent its mitochondrial entry in the early stage of terminal erythropoiesis
To gain insights into the detailed mechanism of plek2 in terminal erythropoiesis, we performed a proteomic study by immunoprecipitating plek2 followed by mass spectrometric analysis, which revealed many novel plek2-binding proteins as well as actin (Figure 4A). Among these proteins, cofilin-1 (hereafter cofilin) is particularly revealing for its critical roles in the regulation of actin dynamics and apoptosis,36–38 which is in line with the phenotypes we observed. In addition, translocation of cofilin to mitochondria is an early step in the induction of apoptosis.39 We confirmed the interaction of plek2 and cofilin by a co-immunoprecipitation assay. Both ectopically overexpressed HA tagged and endogenous plek2 bind to cofilin in the cultured mouse fetal liver erythroblasts (Figure 4B). Cofilin was abundant and constantly expressed in the Epo medium cultured mouse fetal liver erythroblasts (Figure 4C). Knockdown of plek2 in early stage terminal erythroblasts did not influence the total protein level of cofilin or the phosphorylated form (Figure 4D). The level of plek1 also remained unchanged after plek2 knockdown, which ruled out a compensatory effort by plek1 (Figure 4D). The colocalization of plek2 with cofilin was further confirmed by a confocal immunofluorescence microscopic analysis (Figure 4E).
Figure 4.

Plek2 interacts with cofilin. (A) Proteomic study of plek2 interacting proteins. TER119 negative mouse fetal liver erythroblasts were purified and cultured for 30 h in Epo medium. The cells were harvested and lysed. Immunoprecipitation of the cell lysates using rabbit normal IgG and rabbit anti-plek2 were performed separately followed by a silver stain. The distinct protein bands were extracted and submitted for a mass spectrometry analysis. The identified proteins are listed on the right. (B) Immunoprecipitation (IP) of plek2 was performed on HA-plek2 retroviral transduced and non-transduced mouse fetal liver erythroblasts (Endo-plek2) followed by a Western blot analysis of cofilin to detect co-IP. IgG was used as an IP control. (C) Western blot analysis of cofilin and phospho-cofilin levels in freshly purified mouse fetal liver TER119 negative erythroblasts (Day 0) and Epo medium cultured erythroblasts on different days. TER119+ represents freshly purified TER119 positive fetal liver erythroid cells. Hsc70 was used as a loading control. (D) Western blot analysis of plek2, cofilin and plek1 in control and plek2 shRNA knockdown mouse fetal liver erythroblasts in the early stage of terminal erythropoiesis. Hsc70 was used as a loading control. (E) Confocal microscopy of plek2 (red) and cofilin (green) in control cells and cells with plek2 knockdown. The cells were transduced with MSCV encoding control or plek2 shRNA and cultured in SCF medium for 12 h, which was followed by Epo medium culture for 12 h before being fixed for staining. Scale bar: 5 μm. The confocal images were representative optical sections of the cell from at least 50 cells in each condition in 3 different experiments.
The activity of cofilin is tightly regulated by its post-translational modifications.40–42 Oxidation inactivates cofilin and leads to its mitochondrial localization to induce apoptosis.36,39,42 Therefore, it is likely that plek2 interacts with cofilin to inhibit apoptosis by preventing cofilin’s mitochondrial entry in the early stage of terminal erythropoiesis, when the level of ROS is higher (Figure 5A). To directly test this hypothesis, we performed confocal microscopy of cofilin and mitochondria. While cofilin showed a universal nuclear and cytoplasmic localization in the control cells, knockdown of plek2 in the early stage of terminal erythropoiesis induced a relocalization of cofilin to mitochondria (Figure 5B). To further confirm this, we treated the fetal liver erythroblasts with ROS scavenger N-acetyl-cysteine (NAC) to determine whether NAC could rescue the phenotypes induced by plek2 knockdown. NAC by itself at low concentration did not affect cell differentiation but dramatically decreased the cellular ROS level (Figure 5 C and D). With the same dose (5 mM), NAC significantly rescued the differentiation inhibition (Figure 5D and E) in plek2 knockdown cells. Confocal microscopy further demonstrated that NAC rescued cofilin-mitochondria colocalization induced by plek2 knockdown (Figure 5F and G).
Figure 5.

Plek2 is required to prevent cofilin’s mitochondrial entry in the early stage of terminal erythropoiesis. (A) Flow cytometric analysis of the levels of reactive oxidative species (ROS), based on the intensity of CM-H2DCFDA, on different days of Epo medium cultured mouse fetal liver erythroblasts. (B) Confocal microscopy of cofilin and mitochondria localization in control (upper panels) and plek2 shRNA (lower panels) retroviral transduced mouse fetal liver erythroblasts. The TER119 negative mouse fetal liver erythroblasts were purified and transduced with indicated retroviruses followed by culture in SCF medium for 12 h. The cells were then changed to Epo medium for 12 h before harvesting. Scale bar: 5 μm. The confocal images were representative optical sections of the cell from at least 50 cells in each condition in 3 different experiments. (C) Flow cytometric analysis of the ROS levels in control and NAC treated erythroblasts. TER119 negative erythroblasts were treated with different concentrations of NAC on Day 0 in Epo medium. The cells were harvested for flow cytometric assay on Day 1. (D) Flow cytometric analysis of the differentiation profiles in control and plek2 shRNA retroviral transduced mouse fetal liver erythroblasts with or without NAC treatment. The cells were infected, cultured and harvested as in (B). (E) Statistical summary of (D). **P<0.01. (F) Confocal microscopy of cofilin and mitochondria localizations for cells in panel (D). The confocal images were representative optical sections of the cell from at least 50 cells in each condition in 3 different experiments. (G) Statistical analysis of panels (B) and (F) for the percentage of colocalization of cofilin and mitochondria.
Plek2 is required for enucleation but not differentiation or apoptosis in the late stage of terminal erythropoiesis
We further determined the role of plek2 in the late stage of terminal erythropoiesis. To this end, we used the same strategy as in the screening experiments in which the infected mouse fetal liver erythroblasts were cultured immediately in Epo medium. Distinct from the early stage, knockdown of plek2 in late stage terminal erythroblasts neither inhibited differentiation (Figure 6A and B) nor induced apoptosis (Figure 6C). This is consistent with decreased ROS in the late stage of terminal erythroblasts (Figure 5A), and increased phospho-cofilin (Figure 4C), which is mutually exclusive with the oxidized form inducing apoptosis.42,43 In addition, mitochondria are cleared through autophagy in the late stage of terminal erythropoiesis.44 We also detected that the hemoglobin concentration of cells with plek2 knockdown was the same as that in the control cells, which further confirmed intact differentiation (Figure 6D). However, enucleation was affected with plek2 knockdown as demonstrated by the flow cytometric analysis where plek2 shRNA transduced cells showed significantly lower reticulocyte count (TER119highHoechstlow) (Figure 6E and F). Benzidine-Giemsa stains of the cells with plek2 knockdown showed a decreased enucleation rate compared to the controls. However, the cytoplasm was hemoglobin positive with orange stains, again indicating intact differentiation (Figure 6G). Since actin cytoskeleton is critical for enucleation, we next determined actin localization when plek2 was down-regulated in the late stage of terminal erythropoiesis. As an alternative method, we utilized a super-resolution structured illumination microscopy (SIM) technique to visualize detailed structural changes when plek2 was knocked-down. Consistent with the enucleation defect, SIM microscopy illustrated a dramatic disruption of actin cytoskeleton after plek2 knockdown in the late stage of terminal erythropoiesis (Figure 6H).
Figure 6.

Plek2 is required for enucleation but does not influence differentiation or apoptosis in the late stage of terminal erythropoiesis. (A) Flow cytometric analysis of differentiation in the late stage of terminal erythropoiesis. The cells were infected with indicated retroviruses and cultured immediately in Epo medium for two days. (B) Statistical analysis of panel (A). (C) Statistical analysis of flow cytometry of apoptosis using PI and Annexin V with the cells in panel (A). (D) Hemoglobin concentration of the indicated cells in panel (A). (E) Flow cytometric analysis of enucleation in the late stage of terminal erythropoiesis of indicated cells. The cells were infected, cultured, and harvested as in A. (F) Statistical analysis of panel (E) using Students t-test. ***P<0.001. (G) Benzidine-Giemsa staining of cells from A. Arrows indicate incipient reticulocytes. Scale bar: 5 μm. (H) SIM analysis of actin and plek2 localizations in control (upper panels) and plek2 shRNA lentiviral transduced (lower panels) mouse fetal liver erythroblasts. Cells were infected, cultured and harvested as in (A–C). The boxed regions are highlighted on the right. Scale bars: 5 μm. The images are representatives of approximately 20 cells from 3 independent experiments analyzed in each condition. (I) Treatment with H2O2 induced apoptosis in cells with plek2 knockdown. The mouse fetal liver TER119 negative erythroblasts were purified and infected with indicated retroviruses followed by culture in Epo medium. H2O2 was added at 30 h in culture. Apoptosis was analyzed at 48 h in culture. The percentage of surviving cells is presented. **P<0.01.
If lack of apoptosis after plek2 knockdown in the late stage of terminal erythropoiesis were due to decreased ROS level, exposure to exogenous ROS would induce apoptosis in late stage terminal erythroblasts with plek2 downregulation. To confirm this, we treated the cells with hydrogen peroxide (H2O2), a physiological ROS precursor, and tested apoptosis in cells transduced with control and plek2 shRNA in the late stage of terminal erythropoiesis. As we expected, while the control cells survived in the presence of H2O2, the plek2 knockdown cells showed dramatically decreased viable cells under the same condition (Figure 6I).
Discussion
In this study, we used a targeted screening strategy and identified many genes that play novel functions in terminal erythropoiesis. Temporal controlled shRNA expression method allowed us to dissect the roles of the hit genes in different stages of terminal erythropoiesis. We also applied stringent selection criteria to minimize possible false-positive candidates. This is especially important for the discovery of genes in the early stage of terminal erythropoiesis since we used apoptosis as the readout, which could be due to poor transduction efficiency. In this case, a hit gene must fit the criteria that at least three independent shRNAs in duplicated screening experiments should give the same phenotype. Although using apoptosis as the readout may exclude genes that affect or slow down differentiation, it increases the likelihood of the hit genes to be true positive with the severe phenotype. The hit genes were further validated in that one-third of the hit genes important in the early stage of terminal erythropoiesis also functioned in the late stage, where inhibition of enucleation, but not apoptosis, was the phenotype observed. Given the fact that the same lentiviruses produced different phenotypes in the early and late stages of terminal erythropoiesis, these results also demonstrated adequate transduction efficiencies of the shRNA lentiviruses used in the system. It should also be noted that for the screening of genes involved in the late stage of terminal erythropoiesis, downregulation of the hit genes could lead to a slowdown rather than inhibition of enucleation. Although we cannot completely rule this out, the fact that the enucleation rate did not increase after 48 h (data not shown) suggests that these genes are involved in enucleation per se.
Our screening results also revealed that the same pathways are commonly used in the early and late stages of terminal erythropoiesis. Among the hit genes, many are functionally important in transcriptional regulation, chromatin modification, membrane dynamics and apoptosis in other cell types. Correlating with previous reports of their high expression levels,22 genes involved in these pathways represent potential key regulators in terminal erythropoiesis. This is directly confirmed by the finding that Ezh2, one of our hit genes and an enzymatic component of the polycomb repressive complex 2 (PRC2), is important for fetal erythropoiesis.45 This work was reported after our screening and served as a perfect example of the significance of the genes we had discovered in erythropoiesis.
It is unexpected that knockdown of Epo receptor (EpoR) promoted enucleation. An anti-enucleation effect of Epo and EpoR was further suggested by an inhibitory effect of enucleation with high concentration of Epo in culture. Epo signaling pathway was reported to be able to activate Rac GTPases.46 Perhaps an increased Epo signaling may hyperactivate Rac GTPases, which is detrimental to enucleation.10 It is also possible that Epo signaling regulates the expression or function of plek2 in terminal erythropoiesis, as indicated in Figure 2B. Further studies would reveal how Epo signaling is involved in the late stage of terminal erythropoiesis.
We focused on the mechanistic study of plek2 due to its dual functions in the early and late stages of terminal erythropoiesis, as well as high expression level in erythroid cells. Plek2 was reported to regulate actin cytoskeleton rearrangement31,47 and T-cell cytoskeleton reorganization.23 It contains two pleckstrin homology (PH) domains and one disheveled-Egl-10-pleckstrin (DEP) domain, which are present in a broad spectrum of proteins involved in cellular signaling and cytoskeleton organization.48 These domains are important for their membrane association as evidenced in T cells, where plek2 is mostly localized on the plasma membrane.23 However, the roles of plek2 in other lineages of hematopoietic cells, especially erythroid cells, are completely unknown. In this paper, we discovered a novel and critical role of plek2 in erythroid cells by regulating erythroid cell actin cytoskeleton dynamics and apoptosis through its interaction with cofilin. In the early stage of terminal erythropoiesis, plek2 regulates actin cytoskeleton integrity that is critical for cell proliferation and differentiation. Plek2 is also required to interact with cofilin to prevent cofilin’s mitochondrial entry, when the ROS level is high. Downregulation of plek2 leads to the disruption of actin cytoskeleton, which by itself can induce apoptosis. In addition, apoptosis is further enhanced by the mitochondrial entry of cofilin (Figure 7A). On the other hand, in the late stage of terminal erythropoiesis, since ROS is decreased, which is consistent with the clearance of mitochondria by autophagy,44 cells escape apoptosis when plek2 is knocked-down. However, plek2 is still required to regulate normal actin cytoskeleton that is critical for enucleation (Figure 7B).
Figure 7.
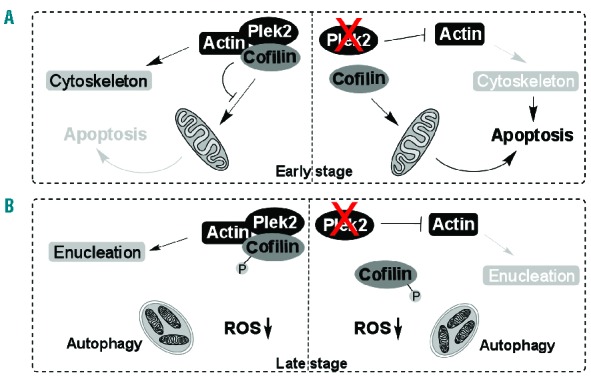
Schematic illustration of the mechanisms of plek2 in the early and late stage of terminal erythropoiesis. (A) In the early stage of terminal erythropoiesis, plek2 is required to regulate actin cytoskeleton integrity and prevent apoptosis by inhibiting cofilin’s mitochondrial entry. (B) In the late stage, ROS is decreased due to mitochondria autophagy. Phospho-cofilin is increased. Plek2 mainly functions in enucleation through the regulation of actin cytoskeleton.
Broadly, plek2-cofilin interaction could have a significant functional impact in other cell types in which plek2 recruits cofilin and regulates apoptosis. Cofilin is pivotal in cell biology as a functional node to regulate a variety of cellular processes.36 Much of the work on the regulation of cofilin has focused on its post-translational modifications and sub-cellular localization. However, how cofilin is regulated through other interacting proteins is unclear. Our study revealed a dual function of plek2 in regulating actin cytoskeleton reorganization and apoptosis through cofilin interaction. In this way, plek2 serves as a docking site and controls the localization of cofilin during erythropoiesis. In addition, plek2 could also interact with cofilin to indirectly function on actin cytoskeleton. These functions of plek2 could be equally important for its regulation of other interacting proteins that we discovered, such as desmoplakin, which is not within the scope of this study. It would also be interesting to explore the role of plek2 in other lineages of the hematopoietic cells.
Acknowledgments
The authors would like to thank Drs. John D. Crispino and David A. Williams for critical reading of the manuscript; Drs. Jing Zhang, William A. Muller, Charles S. Abrams and Deyu Fang for helpful comments. This study was supported by NIH R00HL102154 and an American Society of Hematology scholar award to PJ. The work is also partially supported by the National Center for Advancing Translational Sciences Grant (8UL1TR000150) and the NCI Cancer Center Support Grant (NCI CA060553 to GF)
Footnotes
The online version of this article has a Supplementary Appendix.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Broxmeyer HE. Erythropoietin: multiple targets, actions, and modifying influences for biological and clinical consideration. J. Exp. Med. 2013;210(2):205–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fried W. Erythropoietin and erythropoiesis. Exp Hematol. 2009;37(9):1007–15 [DOI] [PubMed] [Google Scholar]
- 3.Richmond TD, Chohan M, Barber DL. Turning cells red: signal transduction mediated by erythropoietin. Trends Cell Biol. 2005;15(3):146–55 [DOI] [PubMed] [Google Scholar]
- 4.Hattangadi SM, Wong P, Zhang L, Flygare J, Lodish HF. From stem cell to red cell: regulation of erythropoiesis at multiple levels by multiple proteins, RNAs, and chromatin modifications. Blood. 2011;118(24):6258–68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wu H, Liu X, Jaenisch R, Lodish HF. Generation of committed erythroid BFU-E and CFU-E progenitors does not require erythropoietin or the erythropoietin receptor. Cell. 1995;83(1):59–67 [DOI] [PubMed] [Google Scholar]
- 6.Ji P, Murata-Hori M, Lodish HF. Formation of mammalian erythrocytes: chromatin condensation and enucleation. Trends Cell Biol. 2011;21(7):409–15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Keerthivasan G, Wickrema A, Crispino JD. Erythroblast enucleation. Stem Cells Int. 2011;2011:139851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Popova EY, Krauss SW, Short SA, Lee G, Villalobos J, Etzell J, et al. Chromatin condensation in terminally differentiating mouse erythroblasts does not involve special architectural proteins but depends on histone deacetylation. Chromosome Res. 2009;17(1):47–64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ji P, Yeh V, Ramirez T, Murata-Hori M, Lodish HF. Histone deacetylase 2 is required for chromatin condensation and subsequent enucleation of cultured mouse fetal erythroblasts. Haematologica. 2010;95(12):2013–21 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ji P, Jayapal SR, Lodish HF. Enucleation of cultured mouse fetal erythroblasts requires Rac GTPases and mDia2. Nat Cell Biol. 2008;10(3):314–21 [DOI] [PubMed] [Google Scholar]
- 11.Konstantinidis DG, Pushkaran S, Johnson JF, Cancelas JA, Manganaris S, Harris CE, et al. Signaling and cytoskeletal requirements in erythroblast enucleation. Blood. 2012;119(25):6118–27 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sui Z, Nowak RB, Bacconi A, Kim NE, Liu H, Li J, et al. Tropomodulin3-null mice are embryonic lethal with anemia due to impaired erythroid terminal differentiation in the fetal liver. Blood. 2013;123(5):758–67 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ubukawa K, Guo Y-M, Takahashi M, Hirokawa M, Michishita Y, Nara M, et al. Enucleation of human erythroblasts involves non-muscle myosin IIB. Blood. 2012;119(4):1036–44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Watanabe S, De Zan T, Ishizaki T, Yasuda S, Kamijo H, Yamada D, et al. Loss of a Rho-Regulated Actin Nucleator, mDia2, Impairs Cytokinesis during Mouse Fetal Erythropoiesis. CellReports. 2013;5(4):926–32 [DOI] [PubMed] [Google Scholar]
- 15.Keerthivasan G, Liu H, Gump JM, Dowdy SF, Wickrema A, Crispino JD. A novel role for survivin in erythroblast enucleation. Haematologica. 2012;97(10):1471–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chasis JA, Mohandas N. Erythroblastic islands: niches for erythropoiesis. Blood. 2008;112(3):470–8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Zhang L, Flygare J, Wong P, Lim B, Lodish HF. miR-191 regulates mouse erythroblast enucleation by down-regulating Riok3 and Mxi1. Genes Dev. 2011;25(2):119–24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Keerthivasan G, Small S, Liu H, Wickrema A, Crispino JD. Vesicle trafficking plays a novel role in erythroblast enucleation. Blood. 2010;116(17):3331–40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhang J, Socolovsky M, Gross AW, Lodish HF. Role of Ras signaling in erythroid differentiation of mouse fetal liver cells: functional analysis by a flow cytometry-based novel culture system. Blood. 2003;102(12):3938–46 [DOI] [PubMed] [Google Scholar]
- 20.Socolovsky M, Fallon AE, Wang S, Brugnara C, Lodish HF. Fetal anemia and apoptosis of red cell progenitors in Stat5a−/−5b−/− mice: a direct role for Stat5 in Bcl-X(L) induction. Cell. 1999;98(2):181–91 [DOI] [PubMed] [Google Scholar]
- 21.Yoshida H, Kawane K, Koike M, Mori Y, Uchiyama Y, Nagata S. Phosphatidylserine-dependent engulfment by macrophages of nuclei from erythroid precursor cells. Nature. 2005;437(7059):754–8 [DOI] [PubMed] [Google Scholar]
- 22.Wong P, Hattangadi SM, Cheng AW, Frampton GM, Young RA, Lodish HF. Gene induction and repression during terminal erythropoiesis are mediated by distinct epigenetic changes. Blood. 2011;118(16):e1–38 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bach TL, Kerr WT, Wang Y, Bauman EM, Kine P, Whiteman EL, et al. PI3K regulates pleckstrin-2 in T-cell cytoskeletal reorganization. Blood. 2007;109(3):1147–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Yuan B, Latek R, Hossbach M, Tuschl T, Lewitter F. siRNA Selection Server: an automated siRNA oligonucleotide prediction server. Nucleic Acids Res. 2004;32(Web Server issue):W130–4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Luo B, Heard AD, Lodish HF. Small interfering RNA production by enzymatic engineering of DNA (SPEED). Proc Natl Acad Sci USA. 2004;101(15):5494–9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tong W, Zhang J, Lodish HF. Lnk inhibits erythropoiesis and Epo-dependent JAK2 activation and downstream signaling pathways. Blood. 2005;105(12):4604–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Moffat J, Grueneberg DA, Yang X, Kim SY, Kloepfer AM, Hinkle G, et al. A lentiviral RNAi library for human and mouse genes applied to an arrayed viral high-content screen. Cell. 2006;124(6):1283–8 [DOI] [PubMed] [Google Scholar]
- 28.Huang DW, Sherman BT, Lempicki RA. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009;37(1–13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Huang DW, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4(1):44–57 [DOI] [PubMed] [Google Scholar]
- 30.Sathyanarayana P, Menon MP, Bogacheva O, Bogachev O, Niss K, Kapelle WS, et al. Erythropoietin modulation of podocalyxin and a proposed erythroblast niche. Blood. 2007;110(2):509–18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hu MH, Bauman EM, Roll RL, Yeilding N, Abrams CS. Pleckstrin 2, a widely expressed paralog of pleckstrin involved in actin rearrangement. J. Biol. Chem. 1999;274(31):21515–8 [DOI] [PubMed] [Google Scholar]
- 32.Hawley RG, Lieu FH, Fong AZ, Hawley TS. Versatile retroviral vectors for potential use in gene therapy. Gene Ther. 1994;1(2):136–8 [PubMed] [Google Scholar]
- 33.Dominguez R, Holmes KC. Actin structure and function. Annu Rev Biophys. 2011;40:169–86 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kalfa TA, Pushkaran S, Mohandas N, Hartwig JH, Fowler VM, Johnson JF, et al. Rac GTPases regulate the morphology and deformability of the erythrocyte cytoskeleton. Blood. 2006;108(12):3637–45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ji P, Lodish HF. Rac GTPases play multiple roles in erythropoiesis. Haematologica. 2010;95(1):2–4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Bernstein BW, Bamburg JR. ADF/cofilin: a functional node in cell biology. Trends Cell Biol. 2010;20(4):187–95 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Oser M, Condeelis J. The cofilin activity cycle in lamellipodia and invadopodia. J Cell Biochem. 2009;108(6):1252–62 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Blume von J, Duran JM, Forlanelli E, Alleaume A-M, Egorov M, Polishchuk R, et al. Actin remodeling by ADF/cofilin is required for cargo sorting at the trans-Golgi network. J Cell Biol. 2009;187(7):1055–69 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chua BT, Volbracht C, Tan KO, Li R, Yu VC, Li P. Mitochondrial translocation of cofilin is an early step in apoptosis induction. Nat Cell Biol. 2003;5(12):1083–9 [DOI] [PubMed] [Google Scholar]
- 40.Yoo Y, Ho HJ, Wang C, Guan J-L. Tyrosine phosphorylation of cofilin at Y68 by v-Src leads to its degradation through ubiquitin-proteasome pathway. Oncogene. 2010;29(2):263–72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kim J-S, Huang TY, Bokoch GM. Reactive oxygen species regulate a slingshot-cofilin activation pathway. Mol. Biol. Cell. 2009;20(11):2650–60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Klemke M, Wabnitz GH, Funke F, Funk B, Kirchgessner H, Samstag Y. Oxidation of cofilin mediates T cell hyporesponsiveness under oxidative stress conditions. Immunity. 2008;29(3):404–13 [DOI] [PubMed] [Google Scholar]
- 43.Klamt F, Zdanov S, Levine RL, Pariser A, Zhang Y, Zhang B, et al. Oxidant-induced apoptosis is mediated by oxidation of the actin-regulatory protein cofilin. Nat Cell Biol. 2009;11(10):1241–6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sandoval H, Thiagarajan P, Dasgupta SK, Schumacher A, Prchal JT, Chen M, et al. Essential role for Nix in autophagic maturation of erythroid cells. Nature. 2008;454(7201):232–5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mochizuki-Kashio M, Mishima Y, Miyagi S, Negishi M, Saraya A, Konuma T, et al. Dependency on the polycomb gene Ezh2 distinguishes fetal from adult hematopoietic stem cells. Blood. 2011;118(25):6553–61 [DOI] [PubMed] [Google Scholar]
- 46.Arai A, Kanda E, Miura O. Rac is activated by erythropoietin or interleukin-3 and is involved in activation of the Erk signaling pathway. Oncogene. 2002;21(17):2641–51 [DOI] [PubMed] [Google Scholar]
- 47.Ma AD, Abrams CS. Pleckstrin induces cytoskeletal reorganization via a Rac-dependent pathway. J Biol Chem. 1999;274(40):28730–5 [DOI] [PubMed] [Google Scholar]
- 48.Lemmon MA, Ferguson KM, Abrams CS. Pleckstrin homology domains and the cytoskeleton. FEBS Lett. 2002;513(1):71–6 [DOI] [PubMed] [Google Scholar]