

Abstract
The adaptive calibration model (ACM) is an evolutionary–developmental theory of individual differences in stress responsivity. In this article, we tested some key predictions of the ACM in a middle childhood sample (N = 256). Measures of autonomic nervous system activity across the sympathetic and parasympathetic branches validated the 4-pattern taxonomy of the ACM via finite mixture modeling. Moreover, the 4 patterns of responsivity showed the predicted associations with family stress levels but no association with measures of ecological stress. Our hypotheses concerning sex differences in responsivity were only partly confirmed. This preliminary study provides initial support for the key predictions of the ACM and highlights some of the methodological challenges that will need to be considered in future research on this topic.
Keywords: adaptive calibration model, developmental plasticity, respiratory sinus arrhythmia, skin conductance level, stress response
Individual differences in physiological responsivity to stress correlate with an impressive array of psychologically relevant variables, including aggression and antisocial behavior, risk taking, attachment, personality, memory and learning, depression, and anxiety (e.g., Alink et al., 2008; Oskis, Loveday, Hucklebridge, Thorn, & Clow, 2011; Quirin, Pruessner, & Kuhl, 2008; Shirtcliff et al., 2009; Stark et al., 2006; Staton, El-Sheikh, & Buckhalt, 2009; van den Bos, Harteveld, & Stoop, 2009; van Goozen, Fairchild, Snoek, & Harold, 2007). Understanding the causes of such individual differences and their developmental course is a major goal of developmental science. Empirical research in this field, however, has lacked an integrative theoretical framework, making it difficult to synthesize diverse empirical findings and gain a coherent “big picture” of the subject matter.
The Adaptive Calibration Model
Recently, a comprehensive evolutionary–developmental theory of individual differences in stress responsivity has been advanced by Del Giudice, Ellis, and Shirtcliff (2011). The adaptive calibration model (ACM) extends and refines the theory of biological sensitivity to context (BSC; Boyce & Ellis, 2005; Ellis, Essex, & Boyce, 2005). The main elements of the ACM are (a) an evolutionary analysis of the functions of the stress response system, defined as an integrated, hierarchically organized system comprising the autonomic nervous system and the hypothalamic–pituitary–adrenal (HPA) axis; (b) a theory of the adaptive match between environmental conditions and stress responsivity; and (c) a taxonomy of four prototypical responsivity patterns (labeled sensitive [I], buffered [II], vigilant [III], and unemotional [IV]). The four patterns are characterized by combinations of physiological parameters indexing the functioning of the parasympathetic nervous system (PNS), the sympathetic nervous system (SNS; i.e., the two branches of the autonomous nervous system, ANS), and the HPA axis (see Figure 1 and Table 1). For each pattern, the ACM makes predictions about neurobiological correlates (e.g., serotonergic function), behavioral correlates (e.g., aggression, self-regulation), and developmental trajectories (including general patterns of gene–environment interaction).
Figure 1.
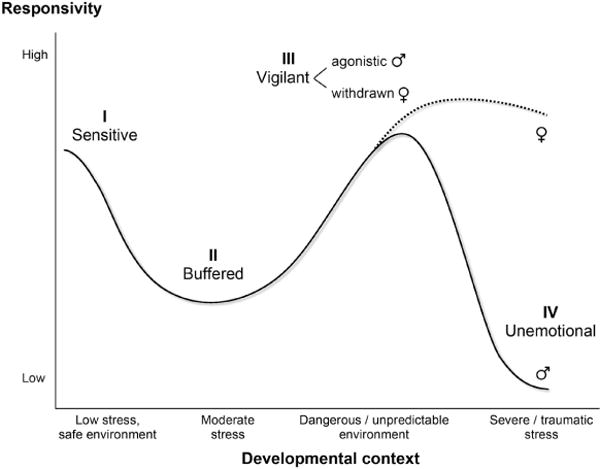
The adaptive calibration model (ACM) of individual differences in development of stress responsivity. At a very general level, a nonlinear relation exists between exposures to environmental stress and support during development and optimal levels of stress responsivity. Although this nonlinear relation is specified for the stress response system (see Table 1), it may apply to other neurobiological systems as well. The figure does not imply that all components of the system will show identical responsivity profiles or that they will activate at the same time or over the same time course. Male/female symbols indicate sex-typical patterns of responsivity, but substantial within-sex differences in responsivity are expected as well. Roman numerals indicate the four prototypical responsivity patterns in the ACM. From “The Adaptive Calibration Model of Stress Responsivity,” by M. Del Giudice, B. J. Ellis, and E. A. Shirtcliff, 2010, Neuroscience & Biobehavioral Reviews, 35, p. 1577. Copyright 2010 by Elsevier.
Table 1.
Predicted Physiological Profiles of the Four Responsivity Patterns
| Physiological profile | Responsivity patterns
|
|||
|---|---|---|---|---|
| I/Sensitive | II/Buffered | III/Vigilant | IV/Unemotional | |
| Parasympathetic nervous system | ||||
| Responsivity | High | Moderate | Low/moderate | Low* |
| Basal | High | Moderate | Low | Low |
| Sympathetic nervous system | ||||
| Responsivity | High/moderate | Low/moderate | High | Low* |
| Basal | Moderate | Low/moderate | High | Low |
| Hypothalamic–pituitary–adrenal (HPA) axis | ||||
| Responsivity | High | Moderate | High | Low |
| Basal | Moderate | Moderate | High/moderate | Low |
Note. Unemotional individuals may display autonomic activation when faced with immediate physical threats and during agonistic confrontations, in contrast with their general pattern of unresponsivity to nonagonistic stressors. From “The Adaptive Calibration Model of Stress Responsivity,” by M. Del Giudice, B. J. Ellis, and E. A. Shirtcliff, 2010, Neuroscience & Biobehavioral Reviews, 35, p. 1578. Copyright 2010 by Elsevier.
The main theoretical foundations of the ACM are life history theory, a biological framework for describing the developmental decisions of organisms and their allocation of resources to different life tasks (life history strategies; see Ellis, Figueredo, Brumbach, & Schlomer, 2009; Hill, 1993; Kaplan & Gangestad, 2005), and the theory of adaptive developmental plasticity (West-Eberhard, 2003). The model integrates many concepts from previous evolutionary accounts of stress responsivity, including those by Flinn (2006; see also Flinn, Nepomnaschy, Muehlenbein, & Ponzi, 2011); Korte, Koolhaas, Wingfield, and McEwen (2005); Porges (1995, 2007); and Taylor et al. (2000). In the ACM, individual differences in the functioning of the stress response system are thought to result largely (though not exclusively) from the operation of evolved mechanisms that match the individual’s physiology and behavior to its local environmental conditions. Thus, patterns of stress responsivity are seen as adaptive in the biological sense, as they function in a way that ultimately tends to maximize the individual’s survival and reproduction. Note that biological adaptiveness is orthogonal to subjective well-being and socially desirable behavior; the two meanings of the word adaptive should never be confused (see Frankenhuis & Del Giudice, 2012). In fact, successful biological adaptation can significantly reduce subjective well-being, the quality of social relationships, and even health—especially when organisms must adapt to harsh or risky environments.
The ACM can be summarized in seven points (for a detailed explanation, see Del Giudice et al., 2011):
The stress response system has three main biological functions: to coordinate the organism’s allostatic response to physical and psychosocial challenges; to encode and filter information from the environment, thus mediating the organism’s openness to environmental inputs; and to regulate a broad range of life history–relevant traits and behaviors.
The stress response system works as a mechanism of conditional adaptation, regulating the development of alternative life history strategies (i.e., suites of coordinated traits including physical growth, sexual maturation, and reproduction-related behavior). Different patterns of activation and responsivity in early development modulate differential susceptibility to environmental influence and shift susceptible children on alternative pathways, leading to individual differences in life history strategies and in the adaptive calibration of stress responsivity.
Activation of the stress response system during the first years of life provides crucial information about life history–relevant dimensions of the child’s environment, namely, danger and unpredictability (see Ellis et al., 2009). This information is used to adaptively regulate the development of the child’s life history strategy.
At a very general level, a nonlinear relationship exists between environmental stress during ontogenetic development and the optimal level of stress responsivity (Figure 1). Note that the environment–responsivity relationship need not be the same for all the components of the stress response system (see Table 1 for detailed predictions for each of the main components). Furthermore, stress responsivity is expected to show domain-specific effects; for example, a generally unresponsive component of the stress response system may respond strongly to some particular type of challenge.
Because of sex differences in optimal life history strategies, sex differences are expected in the distribution of responsivity patterns and in their specific behavioral correlates. Sex differences should become more pronounced at increasing levels of environmental stress; in particular, contexts characterized by severe or traumatic stress should favor the emergence of a male-biased pattern of low responsivity (the unemotional pattern).
Prenatal and early postnatal development, the transition from early to middle childhood, and puberty are likely “switch points” for the calibration of stress responsivity. Individual and sex differences in the functioning of the stress response system are predicted to emerge according to the evolutionary function of each developmental stage.
Responsivity profiles develop under the joint effects of environmental and genetic factors. Genotypic variation may have directional effects on stress responsivity and associated life history strategies, thus predisposing some individuals to follow a certain developmental trajectory. Genotypic variation, in part through effects on the stress response system, may also affect their sensitivity to environmental inputs, resulting in gene–environment interactions whereby some individuals display a broader range of possible developmental outcomes (i.e., broader “reaction norms”) than others.
Patterns of Responsivity
Here we provide a short summary of the main features of the four responsivity patterns, while directing the reader to Del Giudice et al. (2011) for an extensive treatment of this subject. At the end of the section, we briefly discuss the most widely adopted taxonomy of stress responsivity patterns, the one derived from the doctrine of autonomic space (Berntson, Cacioppo, & Quigley, 1991), and compare it with the ACM.
Sensitive (Type I)
Sensitive patterns are hypothesized to develop in safe, predictable conditions and warm family environments. High stress responsivity in sensitive individuals increases their openness to the social and physical environment (Boyce & Ellis, 2005; see also Ellis, Boyce, Belsky, Bakermans-Kranenburg, & van IJzendoorn, 2011). The physiological profile of individuals with this pattern favors sustained but flexible attention and sensitivity to social feedback. Sensitive individuals are reflective, self-and other-conscious, and engaged with the environment; they are also high in inhibitory control, executive function, and delay of gratification. Collectively, these traits promote sustained learning and cooperation.
Buffered (Type II)
Buffered patterns are predicted to develop preferentially in conditions of moderate environmental stress, where they strike a balance between the costs and benefits of responsivity. Buffered responsivity is predicted to arise primarily through moderate, repeated activation of the stress response system during the first years of life. Compared with individuals with Type III and IV patterns, buffered individuals are predicted to be lower in anxiety and aggression, less risk prone, and more sensitive to social feedback. Buffered responsivity can thus look like a “protective factor,” as postulated by the stress inoculation hypothesis (Garmezy, 1991; Rutter, 1993).
Vigilant (Type III)
Vigilant patterns develop in stressful contexts, where they enable people to cope effectively with dangers and threats in the physical and social environment. Their SNS-dominated physiological profile mediates heightened attention to threats and high trait anxiety. In the ACM, vigilance is not associated with a single behavioral pattern but rather with a distribution of patterns involving different mixtures of aggressive/externalizing and withdrawn/internalizing behaviors. In males, vigilant responsivity should be associated more often with increased risk taking, impulsivity, agonistic social competition, and reactive aggression (the vigilant–agonistic subtype; III–A). In females, the typical pattern should involve social anxiety, lower risk taking and impulsivity, and fearful or withdrawn behavior (the vigilant–withdrawn subtype; III–W). Vigilant children who display high levels of both agonistic and withdrawn behaviors (Rogosch & Cicchetti, 1994) may be best described as belonging to a third subtype, the vigilant–agonistic/withdrawn pattern (III–A/W).
Unemotional (Type IV)
Unemotional patterns are marked by a profile of low stress responsivity. Generalized unresponsivity inhibits social learning and sensitivity to social feedback; it can also increase risk taking by blocking information about dangers and threats in the environments. The predicted correlates of this pattern are low empathy and cooperation, impulsivity, competitive risk taking, and antisocial behavior, including high levels of proactive/instrumental aggression (van Goozen et al., 2007), especially in males. Based on life history theory, the distribution of Pattern IV is expected to be male biased, and its behavioral correlates are expected to differ between the sexes. For example, one of the key features of unemotional responsivity in females may be a generalized pattern of aloof social relationships with parents, siblings, and peers. In the ACM, two main developmental pathways are hypothesized to lead to unemotional responsivity patterns. In the first pathway, an initially responsive phenotype shifts toward unresponsivity following chronic severe stress; the shift to Pattern IV should often take place during juvenility or adolescence. In the second pathway, unresponsivity may develop even in low-stress environments because of strong genetic predispositions and may be already apparent in early childhood.
Comparison With the Doctrine of Autonomic Space
The doctrine of autonomic space concerns the simultaneous action of the SNS and PNS on target organs that are innervated by both branches of the autonomic nervous system (e.g., heart; Berntson & Cacioppo, 2004; Berntson et al., 1991). In this model, a two-dimensional space of autonomic responses to a stressor is described, and four main “modes” of response can be identified: reciprocal sympathetic and reciprocal parasympathetic activation (i.e., increased activation in one of the autonomic braches coupled with decreased activation in the other), coactivation (i.e., increased activation of both the SNS and PNS), and coinhibition (i.e., decreased activation of both the SNS and PNS). Note that these are all coupled modes of autonomic response; uncoupled responses are also possible, in which SNS and PNS responses are uncorrelated with one another (Berntson et al., 1991, Berntson, Cacioppo, & Quigley, 1993). The patterns specified by the autonomic space model pertain to modes of activation across the SNS and PNS that may be adaptive or maladaptive in particular contexts. For example, reciprocal sympathetic activation may be well suited for adjustments to challenges, whereas reciprocal parasympathetic activation may be appropriate in situations in which a calm physiological state is beneficial.
The four responsivity patterns of the ACM (see Table 1) differ in important ways from the four patterns described by the doctrine of autonomic space (Berntson et al., 1991, 1993; Cacioppo, Uchino, & Berntson, 1994). Descriptively, the autonomic profiles of sensitive and vigilant types may be thought of as versions of reciprocal sympathetic activation, though the physiological effects of reciprocal sympathetic activation in the vigilant type are likely more pronounced due to lower basal levels of PNS activity and higher basal levels of SNS activity. Autonomic activity of the buffered type might fit into the doctrine of autonomic space as uncoupled parasympathetic withdrawal (moderate withdrawal of PNS influence and minimal-to-moderate sympathetic activation), while the autonomic profile of the unemotional type might best be described as one of coinhibition. These descriptive labels, however, fail to capture the essence of the ACM responsivity patterns. ACM patterns attempt to model the functional organization of the stress response system in different ecologies rather than describe a physiological response to a specific event; for example, one of the predictions of the ACM is that (generally unresponsive) unemotional individuals should display comparatively strong sympathetic activation (or even SNS–PNS coactivation) in response to agonistic stressors or immediate physical threats (Table 1; see Del Giudice et al., 2011, p. 1583). In addition, ACM patterns include HPA functioning as a crucial component of the stress response and explicitly model individual differences in basal activity levels as well as differences in responsivity. Indices of basal autonomic activity have been included in some (but not all) applications of the autonomic space model (e.g., El-Sheikh et al., 2009).
Extensions and empirical testing of the autonomic space model across electrodermal and cardiovascular systems have been conducted (e.g., El-Sheikh et al., 2009). Findings from these studies illustrate a consistent pattern of effects in which environmental risk (e.g., marital conflict and child maltreatment) interacts with both skin conductance and respiratory sinus arrhythmia to predict children’s adjustment problems (El-Sheikh et al., 2009; Gordis, Feres, Olezeski, Rabkin, & Trickett, 2010). Specifically, opposing actions of the PNS and SNS (coinhibition and coactivation) predicted higher levels of aggression and externalizing behavior in the context of stressful, high-risk environments (for further review, see El-Sheikh & Erath, in press). Additionally, evidence has been found for within-physiological system interactions in the context of negative family environments (El-Sheikh, Hinnant, & Erath, 2011), which further suggests that a model encompassing multiple physiological systems at rest and in response to stress would make a significant contribution to understanding the developing stress response systems.
The Present Study
Aim of the Study
The ACM is an exciting theoretical development that promises to integrate the developmental study of stress responsivity in a modern evolutionary framework. An important feature of the ACM is the unified treatment of developmental outcomes in both supportive and risky environments. As yet, however, there have been no direct empirical tests of the model and its novel predictions. Moreover, while the proposed taxonomy of individual differences is theoretically plausible and broadly consistent with the available evidence, it has never been validated empirically.
Previous work in this area has only tested for the hypothesized U-shaped relation between childhood conditions (i.e., levels of early support–adversity) and development of sensitive, buffered, and vigilant phenotypes, as specified by BSC theory. Exploratory analyses in two studies (Ellis et al., 2005) offered confirmatory evidence for this three-pattern classification: the highest prevalences of low reactivity phenotypes or buffered children (as indicated by low-to-average levels of autonomic and adrenocortical reactivity to laboratory stressors) were found under conditions of moderate stress. By contrast, both tails of the support-adversity distribution were associated with higher proportions of high reactivity children (i.e., sensitive and vigilant individuals, respectively). However, Ellis et al. (2005) did not employ high-risk samples and thus was not adequately positioned to identify an unemotional pattern.
In the current study, we extended Ellis et al.’s (2005) study by examining responsivity patterns in a diverse sample of high-risk children (8–10 year-olds). Specifically, our main goals were to (a) validate the four-pattern classification and (b) test the predicted associations between responsivity patterns and environmental conditions. In addition, we formulated a number of hypotheses concerning sex differences in responsivity and the variability of environmental conditions in different responsivity patterns.
We examined autonomic measures across the SNS and PNS branches. SNS activity was indexed by skin conductance level (SCL) and PNS activity was indexed by respiratory sinus arrhythmia (RSA). Both RSA and SCL were assessed during basal conditions and in response to a laboratory task. To operationalize environmental stress and support, we aggregated a wide range of measures of ecological adversity (such as low socioeconomic status [SES], economic strain, and alcohol use by parents) and family relationships, both negative (e.g., harsh parenting, conflict between parents) and positive (e.g., warm parenting, predictable home environment).
Study Hypotheses
Responsivity patterns
We hypothesized the existence of four main patterns of responsivity, each characterized by a specific combination of physiological parameters, as specified in Table 1. Although the four patterns described in Del Giudice et al. (2011) are not intended to be an exhaustive taxonomy of stress responsivity profiles, they should emerge reliably as a broadband descriptive classification.
Environmental stress
The four responsivity patterns should be associated with different levels of environmental stress. Specifically, sensitive (Type I) and buffered (Type II) patterns should be associated with low-to-moderate-stress environmental conditions, whereas vigilant (Type III) and unemotional (Type IV) patterns should be associated with high-stress contexts.
Sex differences
In the ACM, sex differences in responsivity are predicted to increase in high-stress environments. We hypothesized males to be overrepresented in unemotional patterns and underrepresented in vigilant patterns. On the basis of general predictions from life history theory, Del Giudice et al. (2011) also hypothesized that a similar (but much weaker) unbalance might be observed in sensitive versus buffered patterns, with males slightly underrepresented in the former and overrepresented in the latter (see Del Giudice et al., 2011, p. 1581).
Variability in environmental conditions
Our final predictions concerned the relative degree of variability in the environmental conditions associated with each pattern. In the ACM, unemotional patterns are thought to originate from two distinct developmental pathways, one associated with intense chronic stress and the other determined by strong genetic predispositions, even in absence of high levels of stress. For this reason, we expected unemotional children to display wide variation in their exposure to environmental stress. A related (though more speculative) prediction was that buffered patterns should arise in a wider range of conditions compared with sensitive patterns because of the effects of buffering genetic variants on children who would otherwise develop sensitive or vigilant profiles (for details, see Del Giudice et al., 2011, p. 1582).
Method
Participants
Children in third and fourth grades were recruited from three local public school districts in the southeastern United States. Inclusion/exclusion criteria were established for a larger study and were implemented to reduce potential confounds; these criteria included age ranging between 8 and 10 years and absence of mental retardation, learning disability, or diagnosed sleep disorder. The majority of recruited families were reached through letters distributed to schools that were sent home with children. A total of 2,700 letters were sent out to families; of the 314 families who contacted our lab in response to the letters and fit our inclusion criteria, about 90% participated in the study.
The current study included 256 children (135 boys, 121 girls; M age = 9.44 years, SD = 8.31 months). Representative of the community, European American children comprised 64% of the sample; 36% were African American. Both European American and African American families were oversampled across a wide socioeconomic range, which resulted in 31% being at Level 1 or 2 (semiskilled workers), 24% at Level 3 (skilled workers), 37% at level 4 (minor professionals), and 8% at Level 5 (professionals; Hollingshead, 1975). Percentages of families earning the following annual incomes were 17% < $10,000; 11% = $10,000–$20,000; 18% = $20,000–$35,000; 19% = $35,000–$50,000; 16% = $50,000–$75,000; and 13% > $75,000; 6% of participants did not provide income information. Based on mother-reported pubertal status information (1 = prepubertal, 2 = early pubertal, 3 = midpubertal, 4 = late pubertal, 5 = postpubertal; Petersen, Crockett, Richards, & Boxer, 1988), the mean puberty score was 1.45 (SD = 0.29) for boys and 1.71 (SD = 0.43) for girls, indicating that most children were prepubertal.
The majority of children lived with their biological mother (N = 230); of these, 58% (n = 133) also lived with their biological father, 21% (n = 48) lived with their mothers’ partner, and the remaining 21% lived with a single mother (n = 49). The 26 children who did not live with their biological mother most commonly lived with their biological father (57%), and all had a significant mother figure in their lives (e.g., stepmother, grandmother). In the Measures section, we include these individuals as giving mother reports. Twenty-six additional children participated in the larger study but were not included in this article because they did not consent to participate in the physiological data collection session or because of large amounts of pertinent missing data due to equipment malfunction, session termination, and so on.
Procedure
This report is based on a larger study, and only pertinent procedures are discussed. Mothers and children were scheduled to visit our laboratory to complete physiological data collection and questionnaire measures. Upon arrival, informed consent and assent were attained (in preparation for physiological data collection). Children were then seated in the physiological data collection room where measures were collected in two domains of psycho-physiological functioning. Parasympathetic nervous system activity was indexed by RSA, which is a well-validated and commonly used measure of PNS activity (Grossman & Taylor, 2007). Sympathetic nervous system activity was indexed by SCL, a well-validated and commonly used measure of SNS activity (Boucsein, 1992). The two measures were collected under conditions of rest and in response to a laboratory stressor.
After electrodes and a bellows belt were attached to the children, they had a 3-min adaptation period during which they were asked to sit quietly and relax. This was followed immediately by a 3-min baseline measurement during which children were sitting quietly. Pertinent to the current study, this initial baseline was followed by a 3-min star-tracing task in which children were asked to trace an outline of a star on a piece of paper while looking into a mirror to guide their movements (Mirror Tracer; Lafayette Instrument Co., Lafayette, IN). This frustrating task consistently elicits significant stress responses in multiple physiological systems including RSA and SCL reactivity (e.g., El-Sheikh, Hinnant, & Erath, 2011). Children traced the star with their dominant hand on a board placed on their lap; electrodes to measure SCL were attached to the nondominant hand so as to minimize movement artifacts.
Mothers completed questionnaire measures of economic stress, family stress, family warmth, and child adjustment while children were engaged in physiological data collection. Children also completed similar measures via an interview with a well-trained research assistant. Research assistants were also available to help parents or children if they did not understand a question. Families were compensated monetarily for participating in the laboratory session.
Measures
Physiological measures
RSA and SCL during the aforementioned baseline and star-tracing task were obtained for analyses. Data were collected via the MW1000A acquisition system (Mind-ware Technologies, Gahanna, OH) and analyzed with the Mind-ware analysis system. A Mindware BioNex 8-slot chassis was used to collect ECG and SCL data. Cardiovascular responses were recorded with the ECG activity amplifier module and disposable snap pediatric ECG electrodes using a standard or modified lead-II configuration. Respiration was calculated through spectral analysis of thoracic impedance (Zo; Ernst, Litvack, Lozano, Cacioppo, & Berntson, 1999). Physiological data were scored in 1-min intervals with Mindware software (Heart Rate Variability Version 3.0.1). We inspected the cardiovascular data for artifacts and missing R peaks on the basis of improbable interbeat intervals. Missing or misplaced R peaks were inserted manually. RSA was calculated as the natural log of the high frequency power (.15–.40 Hz), a validated method for isolating parasympathetic, vagal influence on the heart (Berntson et al., 1997).
We examined SCL (expressed in microSiemens) using two disposable 1-cm silver metal–silver chloride (Ag–AgCl) skin conductance electrodes and filled with 0% Ag–AgCl wet gel (Mind-ware Technologies). Skin conductance electrodes were placed on the palm of the nondominant hand. Care was taken to make sure that the electrodes did not touch and were taped down to help maintain contact with the skin. Collected data were scored in 1-min intervals using a gain of 10 mV and a low-pass filter of 10-Hz. Mindware’s Electrodermal Activity Version 3.0.1 computer program captured SCL continuously throughout the assessment.
For both RSA and SCL, the three 1-min collections during the baseline period (RSA–B and SCL–B, respectively) and during star-tracing period were averaged to form composite baseline and task scores. RSA and SCL reactivity were calculated as difference scores (task level minus baseline level). Positive reactivity scores reflect RSA augmentation and SCL activation, respectively. Thus, an individual with positive reactivity scores for both RSA and SCL exhibits increased RSA (indexing PNS activation) and increased SCL (indexing SNS activation) to the star-tracing task, a pattern of response analogous to coactivation (Berntson et al., 1991, 1993; El-Sheikh et al., 2009). Conversely, negative reactivity scores reflect decreased RSA and decreased SCL, analogous to coinhibition. Negative RSA and positive SCL reactivity scores would be analogous to reciprocal sympathetic activation, and positive RSA scores and negative SCL scores would be analogous to reciprocal parasympathetic activation. RSA and SCL reactivity scores are referred to as RSA–R and SCL–R, respectively, in the remainder of the article. Consistent with the BSC/ACM theoretical frameworks, more negative RSA scores (reflecting greater parasympathetic withdrawal) and more positive SCL scores (reflecting greater sympathetic activation) were conceptualized as indicators of greater openness or susceptibility to environmental influence.
Although 260 children participated in the physiological data collection, data from four children were lost due to insufficiently gelled skin conductance electrodes. Other sources of missing data appeared to be random; 10 children had individual pieces of physiological data that were missing or excluded due to their outlier status (±4 SDs), but this missing information was not related to demographic characteristics. Children were included in our substantive analyses if they had at least two of four physiological indicators (N = 256).
Measures of environmental stress and support
Summary measures of environmental stress and support were obtained in a two-stage procedure. In the first stage, we computed seven unit-weighted composite variables, based on multiple informants and indicators, with each composite representing a different domain of environmental stress or support. To create these unit-weighted composites, we combined constituent items into scales, and then all of the scale scores within a given domain were standardized and averaged. Some of these composites also included single-item measures (e.g., income-to-needs). The rationale for building unit-weighted composites is that exposure to more stressors contributes cumulatively to increased environmental stress; from a causal standpoint, environmental stress is a consequence (not a cause) of the number and severity of stressors in one’s environment. Thus, stress-related variables should be treated as formative (or causal) indicators of environmental stress rather than as indicators of a latent “stress” dimension (see Edwards & Bagozzi, 2000; Kline, 2006; MacCallum & Browne, 1993); linear composite variables provide a simple way to preserve the formative meaning of the variables (Kline, 2006). Note that in formative measurement (and contrary to reflective measurement), indicators are not expected to correlate with one another; thus, reliability scores based on interitem correlations are irrelevant to formative constructs. In the second stage, we reduced the dimensionality of the composite variables via principal component analysis (PCA). Thus, our approach was partly theory-driven (creation of linear composites based on conceptual affinity) and partly data-driven (PCA reduction).
The seven composite variables were economic strain, maternal depression, alcohol use by parents, interparental conflict, harsh parenting, warm parenting, and home chaos. Mother and child reports of the same constructs (interparental conflict [r = .15], harsh parenting, [r = .19], and warm parenting [r = .27]) were averaged in the same composite variable. As is typically the case, mother and child reports were only weakly correlated; we chose to average them so as to obtain a summary measure of the evaluations made by the two actors. With this method, moderately high scores are obtained when either the mother or the child report high levels of the behavior of interest, whereas the highest scores correspond to families in which both the mother and the child report high levels of the behavior. In absence of specific information on the reasons for discrepancy between mother- and child-reports, we consider this a sensible approach. Moreover, relying exclusively on children’s evaluation of family conflict has been shown to increase the confounding effect of genetic factors (Schermerhorn et al., 2011). Before computing composite measures, we imputed missing cases with stochastic regression in SPSS; random error was modeled with regression residuals (see Allison, 2002). The percentage of missing cases in the original variables ranged from 0% to 27.7% (median = 12.1%).
Economic strain
Consistent with recommendations regarding the importance of examining multiple facets of SES and economic adversity (Braveman et al., 2005), we used several measures of mother-reported economic stress. An overall SES score (Hollingshead, 1975) was derived from parents’ income and education (M = 36.32, SD = 11.88). Familial income-to-needs ratio was also calculated (M = 1.71, SD = 1.04); this measure is based on income as a proportion of the federal poverty line per family size (Institute of Research on Poverty, 2010) and may give a more nuanced view of economic stress (McLoyd, 1998). Mothers also reported on their experience of economic stress with making ends meet (a three-item measure of reported difficulty in being able to pay all bills and expenses; M = 0.01, SD = 2.63, α = .71) and financial cutbacks (a 22-item measure of financial adjustments the family has had to make in the last year in order to get by; M = 17.86, SD = 5.29, α = .86; see Conger, Ge, Elder, Lorenz, & Simon, 1994). Making ends meet may be thought of as reflecting chronic economic stress, while financial cutbacks may be more reflective of acute economic strain that has forced recent changes in budgeting. All economic measures were coded so that higher scores reflect greater economic strain.
Maternal depression
Mothers self-reported on their depressive symptoms with two well-validated measures: the Center for Epidemiologic Studies Depression Scale (CES-D; Radloff, 1977; M = .56, SD = .46, α = .89) and the Symptoms Checklist 90-Revised (Derogatis & Cleary, 1977; M depression = .62, SD = .59, α = .88; M hostility = .35, SD = .42, α = .78). Higher scores reflect higher levels of symptoms.
Alcohol use
Mothers’ and fathers’ alcohol use was assessed by mother report with the Parental Alcohol Experiences scale (Windle, 1997; M mother = 1.08, SD = 0.19; M father = 1.15, SD = 0.35; α = .88). This scale measures frequency of alcohol use in combination with symptoms of alcohol dependence (e.g., “Drank to get rid of a hangover”) and consequences of excessive alcohol use (e.g., “Missed work because of drinking”). Higher scores indicate higher frequency of alcohol use and problems with alcohol use.
Interparental conflict
Both mothers and children reported on conflict between parents using the well-established Conflict Tactics Scale (CTS-2; Straus, Hamby, Boney-McCoy, & Sugarman, 1996). The scale measures both psychological (e.g., “Called spouse fat or ugly”) and physical (e.g., “Punched or hit spouse with an object”) aggression between parents/parent figures. Mothers reported on their spouses’ psychological (α = .79) and physical (α = .87) aggression. Children reported on both parents’ psychological and physical aggression toward each other. Reliabilities (a) for child reports ranged from .68 to .80. Higher scores indicate higher levels of interparental conflict.
Harsh parenting
Mothers and children also reported on harsh parenting with two established and validated measures: the Parent-Child Conflict Tactics Scale (Straus, 1999) and the Parental Behavior Inventory (Schwarz, Barton-Henry, & Pruzinsky, 1985). Subscales used in this study assessed mothers’ self-reported verbal aggression (five items; e.g., “Shouted, yelled, or screamed at child”) and physical aggression (nine items; e.g., “Spanked on the bottom”) toward the child and child-reported verbal and physical aggression from both parents. Reliabilities for mother-reported verbal and physical aggression were .66 and .71, respectively. Child-reported reliabilities for parental verbal and physical aggression ranged from .63 to .70. The second measure of harsh parenting was mother- and child-reported Hostile Control from the Parental Behavior Inventory. The eight items from this subscale index parental control through hostile or threatening interactions (e.g., “Losing temper with child for noncompliance”). Reliability for mothers was .55, and reliabilities for child reports of maternal and paternal hostile control were .54 and .53, respectively. For each measure, higher scores reflect greater levels of harsh parenting. It should be noted that relatively low reliabilities were countered to some extent by taking harsh parenting in the context of multiple reporters and overall family stress in the PCA analysis.
Warm parenting
Warm parenting was assessed through three mother- and child-reported eight-item subscales of the Parental Behavior Inventory: Acceptance (e.g., “Understands problems and worries of child”), Child Centeredness (e.g., “Gives child a lot of care and attention”), and Positive Involvement (e.g., “Enjoys reading to or talking to child about reading”). Mother-reported reliabilities for these scales ranged from .64 to .72 while child-reported reliabilities ranged from .56 to .76 for mothers’ behavior and from .72 to .79 for fathers’ behavior. Higher scores on these scales indicate greater warmth.
Home chaos
Chaotic home environment was assessed through mother reports on the 15-item Confusion, Hubbub, and Order Scale (Matheny, Wachs, Ludwig, & Phillips, 1995). This scale uses true-false items to assess home activity (e.g., “It’s a real zoo in our home”), organization (e.g., “We can find things in our home when we need them”), and schedule (e.g., “We have a regular routine at home”). Reliability for this scale was .77.
Principal component analysis (PCA)
In order to obtain a small number of summary variables representing environmental stress and support, the seven composite variables were further reduced via PCA. Parallel analysis (Horn, 1965; see Hayton, Allen, & Scarpello, 2004) indicated three components, which we then extracted and varimax rotated. Allowing for correlated components by oblique rotation only resulted in weak correlations (rs from −.13 to .22), so an orthogonal structure was preferred for ease of interpretation. The three dimensions of environmental stress thus identified accounted for 63% of the variance in the seven composite variables previously described and were labeled ecological stress, negative family relationships, and family warmth/predictability. Ecological stress had loadings ≥ .40 on economic strain (.84), maternal depression (.69), and alcohol use (.47). The negative family relationships component had loadings ≥ .40 on harsh parenting (.83) and parental conflict (.77). Family warmth/predictability had loadings ≥ .40 on warm parenting (.82) and home chaos (−.71). Parallel analysis and PCA were performed in R Version 2.11, psych package 1.0.
Plan of Analysis
Using the ACM, we sought to identify different patterns of psychophysiological function and to test their association with environmental stress. Whereas any single measure of psychophysiological function during baseline or stress responsivity may be continuously and normally distributed, consideration of multiple indices of psychophysiological function across multiple systems simultaneously may yield distinct patterns of function (see Table 1). In order to capture the complex multivariate distribution characterizing patterns of psychophysiological function, we employed finite mixture modeling.
Finite mixture modeling is a broad term encompassing a new generation of person-centered analyses that is being used increasingly when researchers believe that a sample distribution may be composed of distinct groups of individuals (or classes), each with its own distribution (McLachlan & Peel, 2000); another common name for mixture models dealing with continuous indicators that researchers may be familiar with is latent profile analysis. Mixture modeling is becoming increasingly important in clinical psychology where scientists seek to find specific clusters of related symptoms that differentiate individuals’ disorders (e.g., Vaughn, Edens, Howard, & Smith, 2009); in this respect, finite mixture modeling shares some commonalities with taxometric analysis (Meehl, 2001). In the present case, our mixture model should be regarded as a useful approximation, as both the BSC and the ACM postulate a continuum of individual variation. In particular, the four patterns of responsivity predicted by the ACM are descriptive prototypes rather than qualitatively different categories. However, nonlinear associations between environmental stress and physiological function across multiple systems are much easier to capture in a typological model based on prototypical profiles. We reflected these assumptions by relaxing the constraint of no within-class variability that is typically applied to this kind of model (Sterba & Bauer, 2010).
A significant aspect of building support for multiple classes of individuals involves the use of theoretically derived predictors to differentiate classes from the average as well as classes from one another (Bauer & Curran, 2003; Muthén, 2003). We expected to find four classes of psychophysiological function based on the four indicators of class (RSA–B, RSA–R, SCL–B, and SCL–R) and five predictors of class membership (the three environmental stress variables, race, and sex). In finite mixture modeling, each class indicator is compared with another class (i.e., dummy coded) or the grand mean (i.e., effect coded) and is interpreted in mean differences (e.g., RSA-B for children in one class is higher/lower than that of children in another class or higher/lower than the average). Each class predictor estimate is a logistic regression coefficient indicating the relation between the predictor and membership in one class versus a comparison class. These logistic regression coefficients can be translated into class membership probabilities (or odds ratios) to aid in interpretation (see Peng, Lee, & Ingersoll, 2002, for an overview). Conventional p values were used to assess significant relations (p < .05 indicating significance; p < .10 indicating marginal significance). Post-hoc dummy coded analyses also allowed us to test whether the mean values of predictors of a specific class were significantly different from those of another class.
All mixture analyses were conducted with Latent Gold (a program designed for mixture modeling); AMOS and SPSS were used for post hoc analyses and descriptive statistics. Post hoc analyses consisted of class mean and variance comparisons on indicators and predictors through tests of change in chi square and degrees of freedom (Bentler & Bonett, 1980). Constraints of equality on class means or variances and subsequent change in chi square and degrees of freedom were used to indicate whether means or variances for a given variable were significantly different between any two classes. Missing data in the mixture models and post hoc analyses were handled with full information maximum likelihood estimation. This procedure does not impute missing data but uses all available information to estimate model parameters (Acock, 2005).
Results
Descriptive Statistics
Means, standard deviations, and correlations between the variables used in the finite mixture modeling analysis are shown in Table 2. Physiological measures were generally uncorrelated with one another and with environmental stress, except for a negative correlation of between RSA–B and RSA–R, p < .001; children with higher RSA levels during baselines tended to show greater RSA withdrawal in response to the stressor task. The majority of children (65%) exhibited some level of RSA withdrawal to the stressor. The average RSA response was significantly different from 0, t(254) = −6.70, p < .001. The majority of children (85%) also exhibited some level of SCL activation to the stressor. The average SCL response was also significantly different from 0, t(218) = 10.38, p < .001.
Table 2.
Correlations and Descriptive Statistics for Continuous Variables
| Variable | 1 | 2 | 3 | 4 | 5 | 6 | M | SD |
|---|---|---|---|---|---|---|---|---|
| 1. SCL–baseline | 1.00 | 6.65 | 5.78 | |||||
| 2. SCL–reactivity | .04 | 1.00 | 2.03 | 2.89 | ||||
| 3. RSA–baseline | .00 | −.03 | 1.00 | 6.96 | 1.02 | |||
| 4. RSA–reactivity | .02 | .07 | −.29* | 1.00 | 0.28 | 0.67 | ||
| 5. Ecological stress | −.04 | .06 | .01 | .08 | 1.00 | 0.00 | 1.02 | |
| 6. Negative family relationships | .09 | .02 | −.06 | .04 | .00 | 1.00 | 0.01 | 1.00 |
| 7. Family warmth/predictability | −.11 | −.09 | .03 | .02 | −.03 | .01 | 0.02 | 1.00 |
Note. SCL = skin conductance level; RSA = respiratory sinus arrhythmia.
p < .001.
Number of Classes
To find the number of classes that provided the best fit to the data, we tested a series of models with each successive model estimating an additional class, up to six classes. To avoid local maxima, we used 500 random starts for each model and estimated all analyses twice to ensure that results were replicated (which would be highly unlikely if the first solution was due to arriving at a local maxima; McLachlan & Peel, 2000). We evaluated the models using recommended indices of fit, the Bayesian information criterion (BIC) and the bootstrap likelihood ratio test (BLRT; McLachlan & Peel, 2000; Nylund, Asparouhov, & Muthén, 2007). The BLRT is a formal test of whether a bootstrapped (500 in our models) log likelihood difference distribution between a model with k classes is a significant improvement over a model with k – 1 classes. The BRLT also provides significant p values, indicating that the addition of a model with k classes fits better than a model with k – 1 classes. Conversely, a nonsignificant p value indicates that a model with k classes does not fit better than a model with k – 1 classes (i.e., it would indicate that the simpler model with fewer classes should be accepted). Because of its superior performance in simulation studies (Nylund et al., 2007), we used the BLRT as our first choice in determining the number of classes.
When testing our models, we assumed that at least for some classes, baseline levels of autonomic activity would be correlated with reactivity levels within systems (i.e., RSA–B and RSA–R, SCL–B, and SCL–R), baseline levels would be correlated across systems (RSA–B and SCL–B), and reactivity levels would be correlated across systems (RSA–R and SCL–R; see El-Sheikh et al., 2009). Thus, these indicators were allowed to covary within classes in our models. The empirical model is shown in Figure 2. As indicated by the BLRT model comparisons (shown in Table 3), we found that the best fitting model contained four classes of individuals. This provides support to the four-pattern structure hypothesized on the basis of the ACM.
Figure 2.
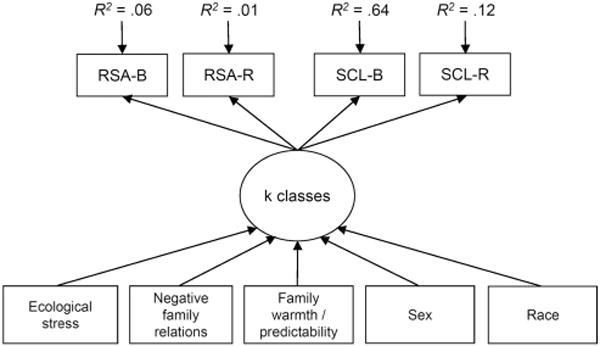
Depiction of the finite mixture model. RSA–B = respiratory sinus arrhythmia at baseline; RSA–R = respiratory sinus arrhythmia reactivity; SCL–B = skin conductance level at baseline; SCL–R = skin conductance level reactivity.
Table 3.
Nested Model Comparisons for Latent Classes of Physiological Activity
| Class | Log likelihood | BIC | BLRT | p |
|---|---|---|---|---|
| 1 | − 1855.03 | 3776.61 | ||
| 2 | − 1755.62 | 3677.60 | 198.83 | <.001 |
| 3 | − 1723.24 | 3712.64 | 64.77 | <.001 |
| 4 | − 1691.47 | 3748.91 | 63.54 | <.001 |
| 5 | − 1669.08 | 3803.96 | 44.77 | .14 |
| 6 | − 1646.25 | 3858.10 | 45.67 | .13 |
Note. BIC = Bayesian information criterion; BLRT = bootstrap likelihood ratio test.
Physiological Profiles
After having identified four classes of individuals, we set out to describe the physiological profile associated with each class and evaluate differences in autonomic functioning between classes. The latent class variable accounted for a significant amount of variance in the indicators SCL–B (64%) and SCL–R (12%). Although the latent class variable did not account for a significant amount of variance in RSA–B (6%), several classes differed from average on RSA–B. This was not the case, however, for RSA–R; the latent class variable accounted for only 1% of variance in this indicator. In other words, physiological differences between the classes were dominated by SNS activity and (to a smaller extent) PNS basal activity, with little role for PNS reactivity. In Table 4, we present class sizes and descriptive statistics for each class. Figure 3 depicts, in Z-score form, prediction of psychophysiological functioning from the unobserved latent class variable; this gives a sense of the relative importance of the physiological indicators that define the classes. Following the descriptive taxonomy of the ACM, the four classes were labeled sensitive, buffered, vigilant, and unemotional.
Table 4.
Class Descriptive Statistics for Indicators and Predictors
| Variable | Sensitive (n = 70; 27%) |
Buffered (n = 115; 45%) |
Vigilant (n = 26; 10%) |
Unemotional (n = 45; (18%) |
|---|---|---|---|---|
| Physiological measures | ||||
| RSA–baseline | 6.99 | 7.17 | 6.97 | 6.50 |
| RSA–reactivity | −0.32 | −0.29 | −0.33 | −0.17 |
| SCL–baseline | 8.96 | 2.49 | 16.48 | 5.03 |
| SCL–reactivity | 2.43 | 1.83 | 3.98 | 0.54 |
| Environmental stress/support | ||||
| Ecological stress | 0.21 | −0.06 | −0.28 | −0.01 |
| Negative family relationships | 0.02 | −0.37 | 0.28 | .62 |
| Family warmth/predictability | 0.22 | 0.16 | −0.27 | −0.37 |
| Demographics | ||||
| Sex (male = 1) | 0.71 | 0.51 | 0.34 | 0.41 |
| Race (African American = 1) | 0.33 | 0.27 | 0.52 | 0.47 |
Note. Estimated class size, class means for physiological parameters, and class means for environmental and demographic predictors. Values for sex and race are proportions. RSA = respiratory sinus arrhythmia; SCL = skin conductance level.
Figure 3.
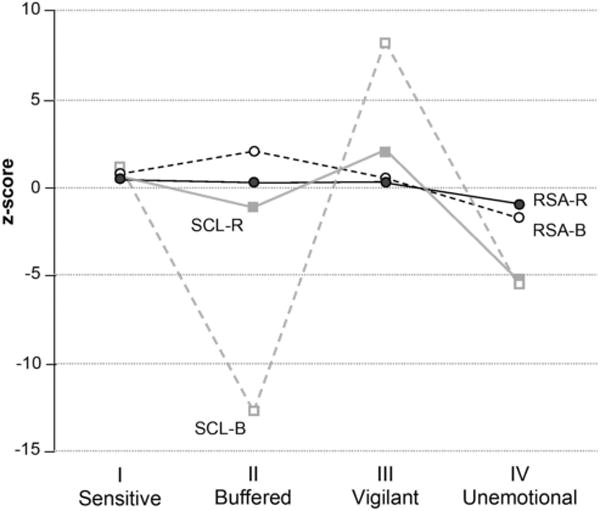
Prediction of physiological indicators of the four latent classes (z scores). Roman numerals indicate the four prototypical responsivity patterns in the adaptive calibration model. SCL–R = skin conductance level reactivity; SCL–B = skin conductance level at baseline; RSA–B = respiratory sinus arrhythmia at baseline; RSA–R = respiratory sinus arrhythmia reactivity. Roman numerals indicate the four prototypical responsivity patterns in the adaptive calibration model.
The largest class (45%) was labeled the buffered class for its similarity with Pattern II of the ACM. Children in this class exhibited high PNS basal activity, low SNS basal activity, and comparatively low SNS reactivity. Specifically, for these children, RSA–B was significantly higher than average and SCL-B was significantly lower than average. Post hoc class comparisons indicated that children in the buffered class had RSA–B levels that were significantly higher than those of children in the unemotional class but not different from children in the other two classes. Children in the buffered class had SCL–B levels that were significantly lower than those of children in any other class. Children in this class also had SCL–R levels that were significantly higher than those in the unemotional class yet lower than levels for children in the vigilant class.
The second largest class (27%) was labeled sensitive for its similarity with Pattern I of the ACM. Although children in this class were not significantly different from average on any psychophysiological characteristic, they had PNS and SNS baselines that were the second highest of any class and PNS and SNS reactivity scores that were the second greatest of any class. RSA–B levels of children in this class were significantly higher than those of children in the unemotional class. SCL–B of children in this class was significantly higher than that of children in both the buffered and the unemotional classes, but lower than those in the vigilant class. Sensitive children’s SCL–R was higher than that of those in the unemotional class, but lower than that of children in the vigilant class.
The third class (18%) was named unemotional for its similarity with Pattern IV of the ACM. Children in this class displayed a consistent pattern of psychophysiological underarousal, with low basal activity in both the PNS and SNS indices and low SNS reactivity. Specifically, the RSA–B of these children was marginally lower than average, SCL–B was lower than average, and SCL–R was lower than average. Compared with children in the other classes, RSA–B and SCL–R were significantly lower. SCL–B for children in this class was significantly lower than that of those in the sensitive and vigilant classes, but higher than that of children in the buffered class.
The smallest class of children (10%) was named vigilant for its similarity with Pattern III of the ACM. Children in this class exhibited high SNS activity at rest and in response to stress, as indicated by higher SCL–B levels than average and higher SCL–R than average. RSA–B levels of vigilant children were significantly higher than those of children in the unemotional class, while SCL–B and SCL–R were significantly higher than in any other class.
Class Predictors: Environmental Stress, Sex, and Race
The next step in the analysis was to test the hypothesized association between the four physiological patterns and measures of environmental stress. In addition, we tested whether class membership was significantly affected by sex and race. We chose the buffered class as the comparison class because it was the largest one in this sample, and because buffered responsivity may be the most common pattern in the general population. Results for prediction of class membership are presented in Table 5. Overall Wald statistics (Table 5) showed that negative family relationships, family warmth/predictability, and sex significantly predicted children’s class membership. In Figure 4, we depict mean levels of environmental stress and support for the four classes.
Table 5.
Estimates for Predictors of Class Membership
| Variable | Sensitive
|
Vigilant
|
Unemotional
|
Wald χ2 | |||
|---|---|---|---|---|---|---|---|
| B | SE | B | SE | B | SE | ||
| Intercept | −0.74† | 0.43 | −1.10* | 0.46 | −0.78 | 0.80 | 7.48† |
| Environmental stress/support | |||||||
| Ecological stress | 0.33 | 0.20 | −0.26 | 0.31 | −0.01 | 0.33 | 4.41 |
| Negative family relationships | 0.62* | 0.30 | 1.08** | 0.37 | 1.33** | 0.46 | 9.94* |
| Family warmth/predictability | 0.06 | 0.23 | − 0.59* | 0.29 | −0.67* | 0.31 | 7.64* |
| Demographics | |||||||
| Sex (male =1) | 0.74 | 0.46 | −1.15~ | 0.64 | −0.88 | 0.62 | 8.34* |
| Race (African American =1) | 0.26 | 0.46 | 1.24* | 0.58 | 1.05 | 0.70 | 4.76 |
Note. Buffered class was used as the comparison class. Estimates are logistic regression coefficients. SE = standard error.
p < .10.
p < .05.
p < .01.
Figure 4.
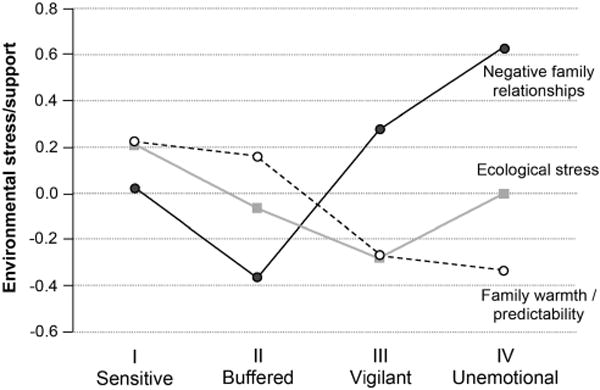
Mean levels of environmental stress/support in the four latent classes. Roman numerals indicate the four prototypical responsivity patterns in the adaptive calibration model.
Consistent with expectations, as the level of children’s negative family relationships increased, odds of membership in the vigilant and unemotional classes also increased. In comparison to children in the buffered class, children at very high levels of negative family relationships (+ 2 SDs) were 2.92 and 6.55 times more likely to be in the vigilant and unemotional classes, respectively. Contrary to expectations, the negative family relationships component was positively related to odds of membership in the sensitive class (compared with the buffered class), but this relation was not as strong as for the vigilant and unemotional classes. As children’s levels of family warmth/predictability increased, odds of membership in the vigilant and unemotional classes decreased. Odds of membership in the vigilant and unemotional classes at very low levels of family warmth/predictability (−2 SDs) were 1.08 and 1.75, respectively, whereas at high levels of family warmth/predictability, children were much less likely to be in the vigilant and unemotional classes. These findings are summarized in Figure 5. Finally, girls were marginally more likely to be in the vigilant class than in the buffered class.
Figure 5.
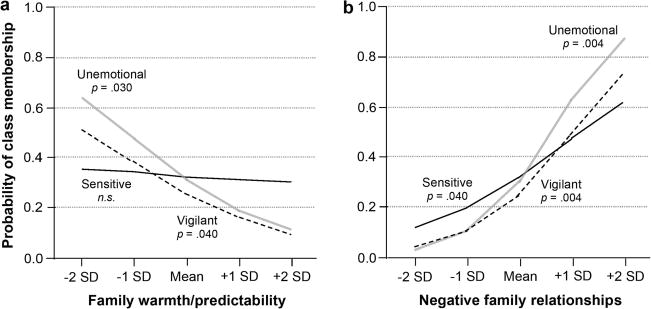
Probability of class membership as predicted by (a) family warmth/predictability and (b) negative family relationships. Probabilities are relative to the Buffered class (a probability of 0.5 indicates that membership in the two classes has the same likelihood).
Post hoc class comparisons were performed on the mean levels of the two stress variables that significantly predicted class membership, (i.e., negative family relationships and family warmth/predictability). In regard to negative family relations, children in the buffered class had scores that were significantly lower than those of the other classes. Children in the sensitive class were lower in negative family relationships than those in the unemotional class, and not significantly different from those in the vigilant class. Consistent with our hypotheses, children in the unemotional class had the highest scores of negative family relationships, which were significantly elevated in comparison to any other class. Also consistent with our hypotheses, children in the sensitive and buffered classes experienced significantly more family warmth/predictability than those in either the unemotional or vigilant classes. The sensitive and buffered classes, however, did not differ significantly from one another on family warmth/predictability, and neither did the unemotional and vigilant classes.
To summarize, our predictions concerning the levels of environmental stress associated with the four classes were broadly confirmed. Two components of environmental stress emerged as significant predictors of class membership: negative family relationships and family warmth/predictability. In contrast, we found no significant effect of ecological stress. Our analysis clearly suggested the existence of two low-stress classes (the high-reactivity sensitive class and the low-reactivity buffered class) and two high-stress classes (the high-reactivity vigilant class and the low-reactivity unemotional class).
The analysis of sex differences in class membership provided a less clear-cut picture. Consistent with our predictions, girls were overrepresented in the vigilant class. However, we found no support for the hypothesized male bias in the unemotional class. Race was not a significant predictor overall, although African American children were marginally less likely to be in the buffered class and marginally more likely to be in the vigilant class.
Within-Class Variability
All of the classes exhibited significant within-class variability, both on the four physiological indicators and on the three environmental stress predictors. Constraining variances for each predictor to be equal across all classes resulted in a significant decrement in model fit. This suggests that it would have been inappropriate to assume that within-class variability is nonexistent or consists only of error variance. It also suggests that, consistent with the ACM, the finite mixture model we fitted to our data is a simplified version of a true multivariate distribution that is more continuous in nature.
Children in the sensitive and unemotional classes experienced significantly more variability in ecological stress than did children in either the buffered or vigilant classes (variance ratio [VR] = 2.0 and 2.2, respectively). Variability in both negative family relationships and family warmth/predictability was highest in the unemotional class and lowest in the buffered class. The unemotional class had significantly more variability in negative family relationships than the buffered and sensitive classes (VR = 4.9 and 1.9, respectively) and marginally more variability than the vigilant class (VR = 1.9). In contrast, the buffered class had significantly less variability in negative family relationships than any other class (VRs from 0.2 to 0.4) and less variability in family warmth/predictability than the unemotional class (VR = 0.6).
In summary, the unemotional class displayed the highest variability in environmental conditions, a finding consistent with our hypotheses. In contrast with our hypotheses, however, children in the buffered class appeared to live in relatively homogeneous contexts compared with children in the sensitive class.
Discussion
In this article, we used a data set with a diverse sample of children (8–10 years old) to test several predictions from the recently developed ACM of stress responsivity (Del Giudice et al., 2011). The results lend initial support to the model, opening the way to further investigations and more fine-grained empirical studies. Despite the limitations of the present study (see Limitations section), most of our key predictions were supported. These results suggest that the ACM may provide a useful, heuristically powerful framework to organize research on stress responsivity.
First, we were able to identify four classes of autonomic nervous system activity indexed by SCL and RSA during resting conditions and in response to a stressful task; the four classes could be reasonably mapped on the four responsivity patterns described in the ACM. Even if variation is treated in the ACM as continuously distributed, discrete latent classes provide a useful approximation of the complex multivariate structure predicted by the model. Furthermore, the four patterns were associated with different levels of family stress; as predicted, high- and low-responsivity patterns were found in both low-stress and high-stress conditions. These results extend the initial three-pattern classification proposed by BSC theory (Boyce & Ellis, 2005) and supported in previous research on relatively low-risk, normative samples (Ellis et al., 2005). Most critically, the current research demonstrates the need for diverse samples, including high-risk children, to capture the full four-pattern classification.
These relationships between family stress and physiological responsivity can be captured only by a highly nonlinear model, such as the ACM and its approximation by finite mixture modeling. As is apparent from Table 2, there are no straightforward, linear associations between any of the environmental measures and the physiological variables. In the absence of an explicit model, standard statistical procedures might have suggested that stress responsivity is virtually unrelated to the family environment. Although high-order interactions can be used effectively to represent nonlinear relations between variables (e.g., El-Sheikh et al., 2011; Keller & El-Sheikh, 2009), they may be difficult to interpret and require very large samples to reach acceptable levels of statistical power.
Whereas the quality of family relationships significantly predicted class membership in the hypothesized direction, we found no significant effect of the ecological stress component (defined by low SES, low income, maternal depression, and alcohol use). Though in need of replication and further investigation, this surprising finding underlines the critical role of the quality of family relationships in the modulation of the stress response, above and beyond that of general socioeconomic variables (Flinn, 2006; Flinn & England, 1997; Flinn et al., 2011). Other studies have found long-term effects of childhood SES on the development of life history strategies (e.g., Ellis et al., 2003; Griskevicius, Delton, Robertson, & Tybur, in press; Griskevicius, Tybur, Delton, & Robertson, 2011). In our sample, however, the severity of ecological stress showed only weak-to-null correlations with the quality of family relationships (see the Method section); this may not generalize to other samples and may not reflect a general pattern in the population. In particular, our sample was atypical with respect to others in the literature, as it included higher levels of poverty, children from single-parent homes, and a large proportion of African American children. Further research is clearly warranted on this issue.
Support for our predictions concerning sex differences was considerably more mixed. As hypothesized, girls were overrepresented in the vigilant class; however, and contrary to predictions, boys were overrepresented in the sensitive class. We also failed to detect the predicted male-biased distribution in unemotional patterns. It should be noted, however, that sample sizes for many of the classes were rather small (e.g., N = 26 for the vigilant class, and N = 45 for the unemotional class). Thus, our ability to test for sex differences in responsivity patterns was rather limited. Larger samples would be required for an adequate statistical test of our predictions. Another factor that may confound the effects of sex is the use of a cognitive stressor to elicit autonomic activation, as males and females differ in their response to achievement-related stressors (Stroud, Salovey, & Epel, 2002). Another factor to consider is that our sample was largely prepubertal, and sex differences in stress responsivity are expected to emerge in middle childhood but intensify in adolescence (Del Giudice et al., 2011).
Finally, the unemotional pattern was associated with a much wider range of environmental conditions compared with the other patterns. This finding is consistent with the hypothesis of two distinct developmental pathways leading to unemotional profiles—one associated with intense, chronic stress, and another driven by genetic predispositions, relatively independent of local stress levels. We had also predicted that buffered patterns would be found in a wider range of environments compared with sensitive patterns. This (admittedly more speculative) prediction was not supported—indeed, the buffered class displayed the lowest amount of environmental variability.
Limitations
This study had a number of methodological limitations, especially in view of the complexity of the model we set out to evaluate. These limitations should be kept in mind when interpreting the results, as they highlight the preliminary nature of the present findings.
To begin with, measures of physiological activity were limited to two indices of the SNS (SCL and SCL reactivity) and two indices of the PNS (RSA and RSA reactivity). Incorporation of other autonomic indices (e.g., pre-ejection period) may yield a different pattern of effects. Since the doctrine of autonomic space was originally conceived to model dual autonomic innervation of the same organ or system, including cardiac indices of both PNS and SNS functioning may result in patterns that match more closely the typologies proposed by Berntson and colleagues (1991; Berntson & Cacioppo, 2004). Additionally, prior studies of these typologies have been based solely on cross-tabulations of autonomic stress reactivity scores (Alkon et al., 2003; Salomon, Matthews, & Allen, 2000) as opposed to the person-centered clustering approach in the current study. It is notable that despite differences in theoretical and analytical approaches, results from the current study support some aspects of the autonomic space typologies.
While we examined physiological profiles in the autonomic nervous system, the responsivity profiles in Table 1 include HPA axis functioning as a key element. In addition, the four classes we identified were dominated by variation in SCL and SCL reactivity, as RSA measures made a much smaller contribution to the responsivity profiles (Figures 2 and 3). Thus, the information available for the identification of responsivity patterns was partial and limited, which may have affected the assignment of children to the four classes. It is likely that many children in the sample would have been assigned to a different class had more physiological parameters been available. Specifically, children in the sensitive and buffered classes had very similar environmental correlates, with sensitive children falling midway between buffered and vigilant ones on the dimension of negative family relationships. It is quite possible that some proportion of the children in the sensitive and vigilant classes was misclassified, partly obscuring the relation between environmental conditions and stress physiology. Furthermore, because sex differences in stress functioning may be especially apparent across HPA responses (see Kudielka & Kirschbaum, 2005; Stroud et al., 2002), limiting our physiological measures to autonomic functioning may have reduced the ability to detect sex differences in responsivity patterns.
A related limitation concerns the task used to elicit RSA and SCL reactivity. Although the star-tracing task is a valid and effective procedure, multiple assessments of responsivity to different types of stressor are probably needed to firmly classify children into one of the four responsivity patterns of the ACM. Indeed, the stress response system undergoes strong activation in response to social challenges; for example, the HPA axis is powerfully activated by social–evaluative threats and social conflict (e.g., Dickerson & Kemeny, 2004; Flinn, 2006). Cognitive tasks alone may not be able capture the full range of the system’s responses (for evidence of different autonomic responses to different types of laboratory stressors, see also Obradović, Bush, & Boyce, 2011). Other studies (e.g., Ellis et al., 2005) made use of a range of standardized stressors from multiple domains (see Boyce et al., 2001); we regard such procedures as the ideal way to elicit robust, ecologically valid responsivity data. Furthermore, domain-specific tasks may be required to test some of the more detailed predictions of the ACM; for example, we anticipate that unemotional children may show strong sympathetic responses to agonistic challenges (Del Giudice et al., 2011). Another possible issue is that autonomic reactivity during the star-tracing task was referenced to resting conditions, rather than autonomic activity during a control task. However, a potential problem in using baselines that correspond closely with the task used to evoke reactivity is that they frequently require enhanced attention, which will evoke RSA and SCL reactivity. In our view, this is a worse confound than using a resting baseline.
While negative family relationships and family warmth/predictability were significant predictors of class membership, there was no clear-cut differentiation between membership in the vigilant and unemotional classes based on these variables; high levels of negative family relationships and low family warmth/predictability were linked to increased odds of membership in both classes. Differentiating children who develop vigilant phenotypes from those who develop unemotional phenotypes may require a fuller assessment of the environmental context; for example, other dimensions of stress such as unpredictability (Ellis et al., 2009) and chronicity should be explicitly measured in addition to stressor intensity (the focus of the current study). Additionally, there is much work to be done regarding which children are likely to transition from response patterns adapted to safe contexts (i.e., sensitive, buffered) to higher risk patterns (i.e., vigilant, unemotional) when the environment undergoes rapid changes for the worse (see El-Sheikh & Hinnant, in press, for additional evidence pertaining to the effects of changes in the family environment on PNS function).
The existence of different responsivity patterns at different frequencies in the population raises additional challenges concerning sample size. If a given pattern is comparatively rare (e.g., the vigilant pattern in the present study), large and diverse samples are required to permit accurate estimates of its characteristics. This was already noted in relation to sex differences but applies equally well to the ecological correlates of responsivity patterns. The issue of sample size would become even more pressing in studies focusing on a finer scale of detail, for example, on the differences between the two hypothesized subtypes of Pattern III (vigilant–agonistic and vigilant–withdrawn).
Finally, our study is not genetically informative, and as such, it cannot evaluate the role of genetic factors in producing the observed association between stress responsivity and the quality of family relationships. In the ACM, genetic variation and environmental inputs interact to determine the development of responsivity patterns; also, different pathways may involve a different causal balance of genetic and environmental factors (see Del Giudice et al., 2011). The present results are certainly consistent with the hypothesis that children’s patterns of stress responsivity reflect conditional adaptations to their developmental context; however, genetically controlled studies will be needed to detail the gene–environment interplay involved in the calibration of the stress response system.
Concluding Remarks
Theoretical models are useful insofar as they make novel, testable predictions. The ACM is a complex model, and it can be used to derive dozens of predictions at different levels of analysis, including hypotheses about the relationship between stress responsivity and behavior, individual differences in neuromodulation, Gene × Environment interactions, and responses to domain-specific stressors (such as agonistic confrontations). Clearly, no single study can address all of these predictions, and multiple studies (both cross-sectional and longitudinal) will be required even to evaluate the more basic ones. The present investigation offers a preliminary test of the model and highlights some of the methodological challenges that will need to be considered in future research on this topic.
Evolutionary psychologists are sometimes criticized for supposedly advancing untestable or unfalsifiable theories (see discussion in Conway & Schaller, 2002; Ketelaar & Ellis, 2000). On the contrary, we showed how an evolutionary-informed perspective on human behavior can generate eminently testable theories and predictions about the stress response system (see also Ellis, Jackson, & Boyce, 2006). For example, we advanced two novel predictions about within-class variability in environmental conditions and tested them. Crucially, one of the hypotheses was supported, while the other one was not supported by the present data. Whereas the limitations of the study render these results provisional, the predictions derived from the ACM are clearly testable and will be sorted out as new evidence accumulates.
Likewise, the original ACM will certainly need revisions as new evidence emerges; however, its firm rooting in modern biological theory make it less likely that the model will need to be discarded. Just as the ACM was developed as an extension of BSC theory (Boyce & Ellis, 2005), a future extension of the ACM may become an integral part of a general theory of human development. Such cumulative theory building is one of the most attractive features of the evolutionary paradigm; we believe that developmental psychology has much to gain from adopting this approach and its outstanding heuristic potential.
Acknowledgments
This research was supported by National Institute of Health Grant R01-HL093246 awarded to Mona El-Sheikh. Marco Del Giudice was supported by a grant (Bando Scienze Umane e Sociali 2008, L.R. n. 4/2006) from Regione Piemonte (Piedmont Regional Administration), Italy. We wish to thank the Auburn University lab staff for data collection and preparation, and the school personnel, children, and parents who participated.
Footnotes
Editor’s Note. David F. Bjorklund served as the action editor for this article—JSE.
Contributor Information
Marco Del Giudice, Department of Psychology, University of Turin, Turin, Italy.
J. Benjamin Hinnant, Department of Psychology, The Catholic University of America.
Bruce J. Ellis, Norton School of Family and Consumer Sciences, University of Arizona
Mona El-Sheikh, Human Development and Family Studies, Auburn University.
References
- Acock AC. Working with missing values. Journal of Marriage and Family. 2005;67:1012–1028. doi: 10.1111/j.1741-3737.2005.00191.x. [DOI] [Google Scholar]
- Alink LRA, van IJzendoorn MH, Bakermans-Kranenburg MJ, Mesman J, Juffer F, Koot HM. Cortisol and externalizing behavior in children and adolescents: Mixed meta-analytic evidence for the inverse relation of basal cortisol and cortisol reactivity with externalizing behavior. Developmental Psychobiology. 2008;50:427–450. doi: 10.1002/dev.20300. [DOI] [PubMed] [Google Scholar]
- Alkon A, Goldstein LH, Smider N, Essex MJ, Kupfer DJ, Boyce WT. Developmental and contextual influences on autonomic reactivity in young children. Developmental Psychobiology. 2003;42:64–78. doi: 10.1002/dev.10082. [DOI] [PubMed] [Google Scholar]
- Allison PD. Missing data. Thousand Oaks, CA: Sage; 2002. [Google Scholar]
- Bauer DJ, Curran PJ. Distributional assumptions of growth mixture models: Implications for overextraction of latent trajectory classes. Psychological Methods. 2003;8:338–363. doi: 10.1037/1082-989X.8.3.338. [DOI] [PubMed] [Google Scholar]
- Bentler PM, Bonett DG. Significance tests and goodness of fit in the analysis of covariance structures. Psychological Bulletin. 1980;88:588–606. [Google Scholar]
- Berntson GG, Bigger JT, Jr, Eckberg DL, Grossman P, Kaufmann PG, Malik M, Van Der Molen MW. Heart rate variability: Origins, methods, and interpretive caveats. Psychophysiology. 1997;34:623–648. doi: 10.1111/j.1469-8986.1997.tb02140.x. [DOI] [PubMed] [Google Scholar]
- Berntson GG, Cacioppo JT. Heart rate variability: Stress and psychiatric conditions. In: Malik M, Camm AJ, editors. Dynamic electrocardiography. New York, NY: Blackwell/Futura; 2004. pp. 57–64. [DOI] [Google Scholar]
- Berntson GG, Cacioppo JT, Quigley KS. Autonomic determinism: The modes of autonomic control, the doctrine of autonomic space, and the laws of autonomic constraint. Psychological Review. 1991;98:459–487. doi: 10.1037/0033-295X.98.4.459. [DOI] [PubMed] [Google Scholar]
- Berntson GG, Cacioppo JT, Quigley KS. Cardiac psychophysiology and autonomic space in humans: Empirical perspectives and conceptual implications. Psychological Bulletin. 1993;114:296–322. doi: 10.1037/0033-2909.114.2.296. [DOI] [PubMed] [Google Scholar]
- Boucsein W. Electrodermal activity. New York, NY: Plenum; 1992. [Google Scholar]
- Boyce WT, Ellis BJ. Biological sensitivity to context: I. An evolutionary-developmental theory of the origins and functions of stress reactivity. Development and Psychopathology. 2005;17:271–301. doi: 10.1017/S0954579405050145. [DOI] [PubMed] [Google Scholar]
- Boyce WT, Quas J, Alkon A, Smider N, Essex M, Kupfer DJ. Autonomic reactivity and psychopathology in middle childhood. British Journal of Psychiatry. 2001;179:144–150. doi: 10.1192/bjp.179.2.144. [DOI] [PubMed] [Google Scholar]
- Braveman PA, Cubbin C, Egerter S, Chideya S, Marchi KS, Metzler M, Posner S. Socioeconomic status in health research: One size does not fit all. Journal of the American Medical Association. 2005;294:2879–2888. doi: 10.1001/jama.294.22.2879. [DOI] [PubMed] [Google Scholar]
- Cacioppo JT, Uchino BN, Berntson GG. Individual differences in the autonomic origins of heart rate reactivity: The psychometrics of respiratory sinus arrhythmia and preejection period. Psychophysiology. 1994;31:412–419. doi: 10.1111/j.1469-8986.1994.tb02449.x. [DOI] [PubMed] [Google Scholar]
- Conger RD, Ge X, Elder GH, Lorenz FO, Simon RL. Economic stress, coercive family process, and developmental problems of adolescents. Child Development. 1994;65:541–561. doi: 10.2307/1131401. [DOI] [PubMed] [Google Scholar]
- Conway LG, III, Schaller M. On the verifiability of evolutionary psychological theories: An analysis of the psychology of scientific persuasion. Personality and Social Psychology Review. 2002;6:152–166. doi: 10.1207/S15327957PSPR0602_04. [DOI] [Google Scholar]
- Del Giudice M, Ellis BJ, Shirtcliff EA. The adaptive calibration model of stress responsivity. Neuroscience & Biobehavioral Reviews. 2011;35:1562–1592. doi: 10.1016/j.neubiorev.2010.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derogatis LR, Cleary PA. Confirmation of the dimensional structure of the SCL-90: A study in construct validation. Journal of Clinical Psychology. 1977;33:981–989. doi: 10.1002/1097-4679(197710)33:4<981::AID-JCLP2270330412>3.0.CO;2-0. [DOI] [Google Scholar]
- Dickerson SS, Kemeny ME. Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychological Bulletin. 2004;130:355–391. doi: 10.1037/0033-2909.130.3.355. [DOI] [PubMed] [Google Scholar]
- Edwards JR, Bagozzi R. On the nature and direction of relationships between constructs and measures. Psychological Methods. 2000;5:155–174. doi: 10.1037/1082-989X.5.2.155. [DOI] [PubMed] [Google Scholar]
- Ellis BJ, Bates JE, Dodge KA, Fergusson DM, Horwood JL, Pettit GS, Woodward L. Does father absence place daughters at special risk for early sexual activity and teenage pregnancy? Child Development. 2003;74:801–821. doi: 10.1111/1467-8624.00569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ellis BJ, Boyce WT, Belsky J, Bakermans-Kranenburg MJ, van IJzendoorn MH. Differential susceptibility to the environment: A neurodevelopmental theory. Development and Psychopathology. 2011;23:7–28. doi: 10.1017/S0954579410000611. [DOI] [PubMed] [Google Scholar]
- Ellis BJ, Essex MJ, Boyce WT. Biological sensitivity to context: II. Empirical explorations of an evolutionary-developmental theory. Development and Psychopathology. 2005;17:303–328. doi: 10.1017/S0954579405050157. [DOI] [PubMed] [Google Scholar]
- Ellis BJ, Figueredo AJ, Brumbach BH, Schlomer GL. The impact of harsh versus unpredictable environments on the evolution and development of life history strategies. Human Nature. 2009;20:204–268. doi: 10.1007/s12110-009-9063-7. [DOI] [PubMed] [Google Scholar]
- Ellis BJ, Jackson JJ, Boyce WT. The stress response systems: Universality and adaptive individual differences. Developmental Review. 2006;26:175–212. doi: 10.1016/j.dr.2006.02.004. [DOI] [Google Scholar]
- El-Sheikh M, Erath SA. Family conflict, autonomic nervous system functioning, and child adaptation: State of the science and future directions. Development and Psychopathology. 23:703–721. doi: 10.1017/S0954579411000034. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Sheikh M, Hinnant JB. Marital conflict, respiratory sinus arrhythmia, and allostatic load: Interrelations and associations with the development of children’s externalizing behavior. Development and Psychopathology. 23:815–829. doi: 10.1017/S0954579411000320. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Sheikh M, Hinnant JB, Erath SA. Developmental trajectories of delinquency symptoms in childhood: The role of marital conflict and autonomic nervous system activity. Journal of Abnormal Psychology. 2011;120:16–32. doi: 10.1037/a0020626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El-Sheikh M, Kouros CD, Erath S, Cummings EM, Keller P, Staton L. Marital conflict and children’s externalizing behavior: Interactions between parasympathetic and sympathetic nervous system activity. Monographs of the Society for Research in Child Development. 2009;74(1, Serial No. 292):1–101. doi: 10.1111/j.1540-5834.2009.00501.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ernst JM, Litvack DA, Lozano DL, Cacioppo JT, Berntson GG. Impedance pneumography: Noise as signal in impedance cardiography. Psychophysiology. 1999;36:333–338. doi: 10.1017/S0048577299981003. [DOI] [PubMed] [Google Scholar]
- Flinn MV. Evolution and ontogeny of the stress response to social challenges in the human child. Developmental Review. 2006;26:138–174. doi: 10.1016/j.dr.2006.02.003. [DOI] [Google Scholar]
- Flinn MV, England BG. Social economics of childhood glucocorticoid stress response and health. American Journal of Physical Anthropology. 1997;102:33–53. doi: 10.1002/(SICI)1096-8644(199701)102:1<33::AID-AJPA4>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Flinn MV, Nepomnaschy PA, Muehlenbein MP, Ponzi D. Evolutionary functions of early social modulation of hypothalamic-pituitary-adrenal axis development in humans. Neuroscience & Biobehavioral Reviews. 2011;35:1611–1629. doi: 10.1016/j.neubiorev.2011.01.005. [DOI] [PubMed] [Google Scholar]
- Frankenhuis WE, Del Giudice M. When do adaptive developmental mechanisms yield maladaptive outcomes? Developmental Psychology. 2012;48:628–642. doi: 10.1037/a0025629. [DOI] [PubMed] [Google Scholar]
- Garmezy N. Resilience in children’s adaptation to negative life events and stressed environments. Pediatric Annals. 1991;20:459–466. doi: 10.1177/104345429901600105. [DOI] [PubMed] [Google Scholar]
- Gordis EB, Feres N, Olezeski CL, Rabkin AN, Trickett PK. Skin conductance reactivity and respiratory sinus arrhythmia among maltreated and comparison youth: Relations with aggressive behavior. Journal of Pediatric Psychology. 2010;35:547–558. doi: 10.1093/jpepsy/jsp113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griskevicius V, Delton AW, Robertson TE, Tybur JM. The environmental contingency of life history strategies: Influences of mortality and socioeconomic status on reproductive timing. Journal of Personality and Social Psychology. doi: 10.1037/a0021082. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griskevicius V, Tybur JM, Delton AW, Robertson TE. The influence of mortality and socioeconomic status on risk and delayed rewards: A life history theory approach. Journal of Personality and Social Psychology. 2011;100:1015–1026. doi: 10.1037/a0022403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grossman P, Taylor EW. Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution, and biobehavioral functions. Biological Psychology. 2007;74:263–285. doi: 10.1016/j.biopsycho.2005.11.014. [DOI] [PubMed] [Google Scholar]
- Hayton JC, Allen DG, Scarpello V. Factor retention decisions in exploratory factor analysis: A tutorial on parallel analysis. Organizational Research Methods. 2004;7:191–205. doi: 10.1177/1094428104263675. [DOI] [Google Scholar]
- Hill K. Life history theory and evolutionary anthropology. Evolutionary Anthropology. 1993;2:78–88. doi: 10.1002/evan.1360020303. [DOI] [Google Scholar]
- Hollingshead AB. Four factor index of social status. 1975 Unpublished manuscript. [Google Scholar]
- Horn JL. A rationale and test for the number of factors in factor analysis. Psychometrika. 1965;30:179–185. doi: 10.1007/BF02289447. [DOI] [PubMed] [Google Scholar]
- Institute for Research on Poverty. What are poverty thresholds and poverty guidelines. 2010 Retrieved from http://www.irp.wisc.edu/faqs/faq1.htm.
- Kaplan HS, Gangestad SW. Life history theory and evolutionary psychology. In: Buss DM, editor. Handbook of evolutionary psychology. Hoboken, NJ: Wiley; 2005. pp. 68–95. [Google Scholar]
- Keller PS, El-Sheikh M. Salivary alpha-amylase as a longitudinal predictor of children’s externalizing symptoms: Respiratory sinus arrhythmia as a moderator of effects. Psychoneuroendocrinology. 2009;34:633–643. doi: 10.1016/j.psyneuen.2008.12.016. [DOI] [PubMed] [Google Scholar]
- Ketelaar T, Ellis BJ. Are evolutionary explanations unfalsifiable? Evolutionary psychology and the Lakatosian philosophy of science. Psychological Inquiry. 2000;11:1–21. doi: 10.1207/S15327965PLI1101_01. [DOI] [Google Scholar]
- Kline RB. Reverse arrow dynamics: Formative measurement and feedback loops. In: Hancock GR, Mueller RO, editors. Structural equation modeling: A second course. Greenwich, CT: Information Age; 2006. pp. 43–68. [Google Scholar]
- Korte SM, Koolhaas JM, Wingfield JC, McEwen BS. The Darwinian concept of stress: Benefits of allostasis and costs of allostatic load and the trade-offs in health and disease. Neuroscience & Biobehavioral Reviews. 2005;29:3–38. doi: 10.1016/j.neubiorev.2004.08.009. [DOI] [PubMed] [Google Scholar]
- Kudielka BM, Kirschbaum C. Sex differences in HPA axis responses to stress: A review. Biological Psychology. 2005;69:113–132. doi: 10.1016/j.biopsycho.2004.11.009. [DOI] [PubMed] [Google Scholar]
- MacCallum RC, Browne MW. The use of causal indicators in covariance structure models: Some practical issues. Psychological Bulletin. 1993;114:533–541. doi: 10.1037/0033-2909.114.3.533. [DOI] [PubMed] [Google Scholar]
- Matheny AP, Wachs TD, Ludwig JL, Phillips K. Bringing order out of chaos: Psychometric characteristics of the confusion, hubbub, and order scale. Journal of Applied Developmental Psychology. 1995;16(95):429–444. 90028–4. doi: 10.1016/0193-3973. [DOI] [Google Scholar]
- McLachlan GJ, Peel D. Finite mixture models. New York, NY: Wiley; 2000. [Google Scholar]
- McLoyd VC. Socioeconomic disadvantage and child development. American Psychologist. 1998;53:185–204. doi: 10.1037/0003-066X.53.2.185. [DOI] [PubMed] [Google Scholar]
- Meehl PE. Comorbidities and taxometrics. Clinical Psychology: Science and Practice. 2001;8:507–519. doi: 10.1093/clipsy.8.4.507. [DOI] [Google Scholar]
- Muthén B. Statistical and substantive checking in growth mixture modeling: Comment on Bauer and Curran 2003. Psychological Methods. 2003;8:369–377. doi: 10.1037/1082-989X.8.3.369. [DOI] [PubMed] [Google Scholar]
- Nylund KL, Asparouhov T, Muthén BO. Deciding on the number of classes in latent class analysis and growth mixture modeling: A Monte Carlo simulation study. Structural Equation Modeling. 2007;14:535–569. doi: 10.1080/10705510701575396. [DOI] [Google Scholar]
- Obradović J, Bush NR, Boyce WT. The interactive effect of marital conflict and stress reactivity on externalizing and internalizing symptoms: The role of laboratory stressors. Development and Psychopathology. 2011;23:101–114. doi: 10.1017/S0954579410000672. [DOI] [PubMed] [Google Scholar]
- Oskis A, Loveday C, Hucklebridge F, Thorn L, Clow A. Anxious attachment style and salivary cortisol dysregulation in healthy female children and adolescents. Journal of Child Psychology and Psychiatry. 2011;52:111–118. doi: 10.1111/j.1469-7610.2010.02296.x. [DOI] [PubMed] [Google Scholar]
- Peng CJ, Lee KL, Ingersoll GM. An introduction to logistic regression analysis and reporting. The Journal of Educational Research. 2002;96:3–14. doi: 10.1080/00220670209598786. [DOI] [Google Scholar]
- Petersen AC, Crockett L, Richards M, Boxer A. A self-report measure of pubertal status: Reliability, validity, and initial norms. Journal of Youth and Adolescence. 1988;17:117–133. doi: 10.1007/BF01537962. [DOI] [PubMed] [Google Scholar]
- Porges SW. Orienting in a defensive world: Mammalian modifications of our evolutionary heritage. A polyvagal theory. Psychophysiology. 1995;32:301–318. doi: 10.1111/j.1469-8986.1995.tb01213.x. [DOI] [PubMed] [Google Scholar]
- Porges SW. The polyvagal perspective. Biological Psychology. 2007;74:116–143. doi: 10.1016/j.biopsycho.2006.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quirin M, Pruessner JC, Kuhl J. HPA system regulation and adult attachment anxiety: Individual differences in reactive and awakening cortisol. Psychoneuroendocrinology. 2008;33:581–590. doi: 10.1016/j.psyneuen.2008.01.013. [DOI] [PubMed] [Google Scholar]
- Radloff LS. The CES–D scale: A self-report depression scale for research in the general population. Applied Psychological Measurement. 1977;1:385–401. doi: 10.1177/014662167700100306. [DOI] [Google Scholar]
- Rogosch FA, Chicchetti D. Illustrating the interface of family and peer relations through the study of child maltreatment. Social Development. 1994;3:291–308. doi: 10.1111/j.1467-9507.1994.tb00046.x. [DOI] [Google Scholar]
- Rutter M. Resilience: Some conceptual considerations. Journal of Adolescent Health. 1993;14:626–631. doi: 10.1016/1054-139X(93)90196-V. [DOI] [PubMed] [Google Scholar]
- Salomon K, Matthews KA, Allen MT. Patterns of sympathetic reactivity in a sample of children and adolescents. Psychophysiology. 2000;37:842–849. doi: 10.1111/1469-8986.3760842. [DOI] [PubMed] [Google Scholar]
- Schermerhorn AC, D’Onofrio BM, Turkheimer E, Ganiban JM, Spotts EL, Lichtenstein P, Neiderhiser JM. A genetically informed study of associations between family functioning and child psychosocial adjustment. Developmental Psychology. 2011;47:707–725. doi: 10.1037/a0021362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwarz JC, Barton-Henry ML, Pruzinsky T. Assessing child-rearing behaviors: A comparison of rating made by mother, father, child, and sibling on the CRPBI. Child Development. 1985;56:462–479. doi: 10.2307/1129734. [DOI] [PubMed] [Google Scholar]
- Shirtcliff EA, Vitacco MJ, Graf AR, Gostisha AJ, Merz JL, Zahn-Waxler C. Neurobiology of empathy and callousness: Implications for the development of antisocial behavior. Behavioral Sciences & the Law. 2009;27:137–171. doi: 10.1002/bsl.862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stark R, Wolf OT, Tabbert K, Kagerer S, Zimmermann M, Kirsch P, Vaitl D. Influence of the stress hormone cortisol on fear conditioning in humans: Evidence for sex differences in the response of the prefrontal cortex. NeuroImage. 2006;32:1290–1298. doi: 10.1016/j.neuroimage.2006.05.046. [DOI] [PubMed] [Google Scholar]
- Staton L, El-Sheikh M, Buckhalt JA. Respiratory sinus arrhythmia and cognitive functioning in children. Developmental Psychobiology. 2009;51:249–258. doi: 10.1002/dev.20361. [DOI] [PubMed] [Google Scholar]
- Sterba SK, Bauer DJ. Matching method with theory in person-oriented developmental psychopathology research. Development and Psychopathology. 2010;22:239–254. doi: 10.1017/S0954579410000015. [DOI] [PubMed] [Google Scholar]
- Straus MA. Child-report, adult-recall, and sibling versions of the Revised Conflict Tactics Scale. Durham, NC: Family Research Laboratory; 1999. [Google Scholar]
- Straus MA, Hamby SL, Boney-McCoy S, Sugarman DB. The Revised Conflict Tactics Scales (CTS-2): Development and preliminary psychometric data. Journal of Family Issues. 1996;17:283–316. doi: 10.1177/019251396017003001. [DOI] [Google Scholar]
- Stroud LR, Salovey P, Epel ES. Sex differences in stress responses: Social rejection versus achievement stress. Biological Psychiatry. 2002;52:318–327. doi: 10.1016/S0006-3223(02)01333-1. [DOI] [PubMed] [Google Scholar]
- Taylor SE, Klein LC, Lewis BP, Gruenewald TL, Gurung RA, Updegraff JA. Biobehavioral responses to stress in females: Tend-and-befriend, not fight-or-flight. Psychological Review. 2000;107:411–429. doi: 10.1037/0033-295X.107.3.411. [DOI] [PubMed] [Google Scholar]
- van den Bos R, Harteveld M, Stoop H. Stress and decisionmaking in humans: Performance is related to cortisol reactivity, albeit differently in men and women. Psychoneuroendocrinology. 2009;34:1449–1458. doi: 10.1016/j.psyneuen.2009.04.016. [DOI] [PubMed] [Google Scholar]
- van Goozen SHM, Fairchild G, Snoek H, Harold GT. The evidence for a neurobiological model of childhood antisocial behavior. Psychological Bulletin. 2007;133:149–182. doi: 10.1037/0033-2909.133.1.149. [DOI] [PubMed] [Google Scholar]
- Vaughn MG, Edens JF, Howard MO, Smith ST. An investigation of primary and secondary psychopathy in a statewide sample of incarcerated youth. Youth Violence and Juvenile Justice. 2009;7:172–188. doi: 10.1177/1541204009333792. [DOI] [Google Scholar]
- West-Eberhard MJ. Developmental plasticity and evolution. New York, NY: Oxford University Press; 2003. [Google Scholar]
- Windle M. Parental Alcohol Experiences Scale. Research Institute on Addictions; Buffalo, NY: 1997. Unpublished inventory. [Google Scholar]