

Abstract
Marine sponges are the oldest Metazoa, very often presenting a complex microbial consortium. Such is the case of the marine sponge Arenosclera brasiliensis, endemic to Rio de Janeiro State, Brazil. In this investigation we characterized the diversity of some of the culturable heterotrophic bacteria living in association with A. brasiliensis and determined their antimicrobial activity. The genera Endozoicomonas (N = 32), Bacillus (N = 26), Shewanella (N = 17), Pseudovibrio (N = 12), and Ruegeria (N = 8) were dominant among the recovered isolates, corresponding to 97% of all isolates. Approximately one third of the isolates living in association with A. brasiliensis produced antibiotics that inhibited the growth of Bacillus subtilis, suggesting that bacteria associated with this sponge play a role in its health.
Keywords: Symbiont, Antimicrobial activity, Bacteria, Porifera
Introduction
Marine sponges are the oldest Metazoans living on Earth, and are considered to harbor one of the richest microbial symbiont communities among marine invertebrates (Taylor et al., 2007). More than 47 bacterial phyla have already been detected in sponges (Reveillaud et al., 2014), some of which are almost exclusively found in these marine invertebrates (Taylor et al., 2012). For example, a new species of Shewanella was isolated from sponges (Lee et al., 2006) and a new candidate phyla, named Poribacteria, was proposed to occur almost exclusively in association with sponges (Fieseler et al., 2004; Taylor et al., 2012). In addition to the high taxonomic diversity of bacteria coexisting in a single sponge individual, experimental evidences show that sponge-associated microbial symbionts produce secondary metabolites (Noyer, Thomas & Becerro, 2011; Sacristán-Soriano et al., 2011; Schneemann et al., 2011; Blunt et al., 2012). Moreover, sponge microbial symbionts present high metabolic diversity, based on variable sources of carbon, nitrogen, oligoelements and other essential nutrients (Hoffmann et al., 2005; Weisz et al., 2007). The investigation of the symbiotic microbial community diversity constitutes an essential aspect to understand marine sponge ecology and possible biotechnological applications.
A particularly relevant example of association between microorganisms and sponges occurs between a Micromonospora strain, which produces a bioactive alkaloid, and a haplosclerid sponge (Taylor et al., 2007). The direct assignment of a specific sponge-associated isolate of biotechnological relevance is however a rare occurrence, even considering the increasing efforts towards the discovery of sponge associated bacteria, cyanobacteria and fungi capable of producing biologically active secondary metabolites (Wang, 2006; Öztürk et al., 2013).
Generally, the assignment of an organism as the producer of a given compound is not straightforward, since the production of the compound may be result of the cooperation between symbionts and the host or among the symbionts (Hentschel et al., 2012). Many sponge bioactive compounds are structurally similar to products of prokaryote polyketide synthases (PKS) and non-ribosomal peptide synthases (NRPS). These two enzyme families are involved in many biosynthetic pathways of natural products (Piel, 2009). Setting the bacterial origin of bioactive compounds offers the possibility for heterologous large-scale production (Graça et al., 2013).
The marine sponge Arenosclera brasiliensis Muricy & Ribeiro, 1999 (Haplosclerida: Callyspongiidae), is endemic from the upwelling region of Cabo Frio, Rio de Janeiro State, Brazil (Muricy et al., 2011), and the natural source of antibiotic and cytotoxic polycyclic alkaloids (Torres et al., 2000; Berlinck et al., 2004). Evidences suggesting that bacteria associated with A. brasiliensis are the source of bioactive compounds of pharmacological interest are numerous. Recent metagenomic studies uncovered the highly diverse microbial community of A. brasiliensis (Trindade-Silva et al., 2012). Metagenomic analysis of A. brasiliensis also showed a high diversity of bacteria-derived genes encoding for a ketosynthase (KS) domain of the multifunctional type I polyketide synthases (PKS-I) (Trindade-Silva et al., 2013). Such feature is of particular interest, since it has been hypothesized that alkaloids from haplosclerid sponges are assembled from polyketide precursors related to type I PKS enzymology (Fontana, 2006). Moreover, it has been shown that polyketide-derived secondary metabolites isolated from sponges and other marine invertebrates frequently are metabolic products of associated bacteria (Piel et al., 2004; Piel, 2006; Piel, 2009). Consequently, one could hypothesize that the biosynthetic pathway leading to the formation of A. brasiliensis alkaloids may be of bacterial origin. Since many bacterial PKS producers are culturable, the isolation and characterization of A. brasiliensis associated-bacteria is the first step to establish the identity of sponge microorganisms which may have biotechnological potential.
The purpose of the present investigation was the isolation and taxonomic identification of bacteria associated with A. brasiliensis and to evaluate the ability of the bacterial isolates to produce antibacterial metabolites.
Material and Methods
Sample collection and bacterial isolation
Specimens of A. brasiliensis were collected at approximately 10 m depth by Scuba, at João Fernandinho Beach, Búzios, state of Rio de Janeiro (22°44′49″S 41°52′54″W) in January 2011. The sampling site is characterized by tropical conditions with very low annual rainfall. Water temperature ranges from 19 to 27 °C and mean daily air temperatures range from 16 to 28 °C. This part of the coast is influenced by upwelling of cold, nutrient-rich waters and the phenomenon is associated with local wind regime, bathymetry and seasonality (Yoneshigue, 1985; Guimaraens & Coutinho, 1996). Sponge specimens were transported to the laboratory in 20 L containers under controlled temperature (24 °C) in aerated seawater for approximately 3 h. In the laboratory, two specimens were three times washed in 10 mL sterile seawater for removing unassociated micro-organisms. Then, these specimens were dried by gently pressing on sterile paper towels. A piece of sponge (approx. 1 g) was homogenized in 5 mL sterile saline solution (3% NaCl in distilled water). The homogenate was 10-fold serially diluted starting with 100 µL homogenate in 900 µL sterile water to obtain dilutions of 10−1; 10−2 and 10−3 of the initial concentration. A 100 µL aliquot of each dilution was plated onto each of two growth media: BD Difco™ Marine Agar 2216 and MacConkey Agar, both supplemented with Amphotericin B (1 µg/mL) to inhibit the growth of fungi. Isolated bacterial colonies from both media were selected and streaked again at least twice to obtain pure cultures. Pure cultures were preserved in glycerol 20% at −80 °C.
DNA extraction and 16S rRNA sequencing
Bacterial isolates were grown on the same media used for isolation and incubated for 2–3 days aerobically at 30 °C. Bacterial isolates were harvested from agar plates and suspended in 200 µL of ultrapure sterile water. The bacterial suspensions were subjected to boiling for 10 min and freezing for 3 min to lyse the cells. The lysates were centrifuged for 1 min (4 °C at 13,000 g) for pelleting the cells debris. The resulting supernatant DNA solutions were used for the Polymerase Chain Reaction (PCR) as it follows.
Partial 16S rRNA gene sequences were amplified using the primers 27F (AGA GTT TGA TCM TGG CTC AG) (Lane, 1991) and 1093R (GTT GCG CTC GTT GCG GGA CT) (Thompson et al., 2001). A single PCR contained 25 µL with 1 µL DNA (10–80 ng); 1.5 mM MgCl2; 200 µM of each deoxynucleoside triphosphate; 0.5 µM forward and reverse primers and 1 U goTaq DNA polymerase. PCR cycles consisted of initial denaturation at 95 °C for 5 min and cycles of 95 °C for 1.5 min, annealing at 55–51 °C (the temperature was decreased by 1 °C between consecutive steps) for 1 min and the extension at 72 °C for 2 min. The three first cycles containing the higher annealing temperatures (55–53) were repeated 2 times followed by 5 repeats of the cycle with annealing at 52 °C. Finally, 28 repeats of the cycle with annealing at 51 °C and final extension at 72 °C for 4 min. PCR amplicons were checked and the quantity was estimated by electrophoresis on 1% agarose gels and KODAK MI SE—Molecular Imaging Software.
The PCR products were purified using ExoSap-IT (USB Corporation, Cleveland, OH, USA) and sequenced for both the forward and reverse strands using ABI Big Dye chemistry on an ABI 3500 DNA sequencer (Applied Biosystems, Foster City, CA, USA).
Phylogenetic reconstruction
All sequences were edited using SEQMANII software (DNASTAR, Inc.) and aligned in Clustal X with MEGA5 (Tamura et al., 2011). The aligned sequences were visually inspected and edited when necessary.
Partial 16S rRNA gene sequences were analyzed first via BLASTn tool (nucleotide collection database) (Altschul et al., 1990) and RDP Naive Bayesian rRNA Classifier Version 2.6, Sep 2013 (Wang et al., 2007) to aid selection of most closely related reference sequences. The alignment containing isolates and reference sequences was used first for determining the substitution model of evolution using Modeltest (Posada & Crandall, 1998) for maximum likelihood (ML) and neighbor-joining (NJ) phylogenetic reconstruction using MEGA5. Model-test result was Tamura Nei model with Gamma distributed rates among sites (TrN + G). Both phylogenetic methods were run with 1,000 bootstrap replicates.
Gene sequences obtained in this study are available through the website TAXVIBRIO (http://www.taxvibrio.lncc.br/) and at GenBank database under the accession numbers KJ372433 to KJ372528.
Antimicrobial activity assays
Antimicrobial activity against Bacillus subtilis was assessed using an adapted top agar method (Brady, 2007). Briefly, bacterial isolates were grown at 28 °C in the center of squares (5.76 cm2) marked in 245 mm × 245 mm sterile plates (Corning) during 6 days. After this period, the plates containing isolates were exposed to 3 mL of chloroform, during 1 h in order to kill the isolates, and then the plates were left opened to ensure complete chloroform evaporation. An overlay with B. subtilis LMG 7135T was poured over the killed isolates. The soft agar medium containing B. subtilis was prepared as follows: firstly, B. subtilis was grown in 50 mL Luria Broth (LB) medium at 28 °C for 16 h. Then, 200 mL of LB soft agar (0.7% agar) at 45 °C was added to B. subtilis inoculum. This semi solid medium containing B. subtilis was then poured over those killed isolates. The antimicrobial activity was taken into account with the appearance of haloes in B. subtilis confluent growth after 24 and 48 h. Four replicates were performed for each isolate in order to confirm antimicrobial activity. Control tests corresponded to squares where no isolate was inoculated and complete growth of B. subtilis was expected.
Antimicrobial activity against Vibrio sinaloensis LMG 25238T was also screened by a modified double-layer method previously described (Westerdahl et al., 1991). Briefly, the isolates were individually cultivated in Trypic Soil Agar (TSA) (NaCl 3%) for 48 h at 30 °C, under aerobic conditions. Dilutions on sterile distilled water (NaCl 3.0%) were adjusted using a spectrophotometer to optical density of 0.08–0.1 at 625 nm, corresponding to approx. 108 cells mL−1 and seeded, using a steers replicator, on plates containing marine agar. After incubation for 48 h at 30 °C, under aerobic conditions, growing spots were observed. V. sinaloensis (108 cels mL−1) was incorporated into sterile fluid semisolid marine medium (0.5% agar) and spread over the plates containing the isolated bacterial spots. Plates containing the isolates and the test-strain were incubated for 48 h at 30 °C. The bacterial growth inhibition was observed by determining the presence of the inhibition zones. The assay was performed in triplicate.
Results
A total of 98 culturable bacterial isolates were obtained and their partial 16S rRNA gene sequences were compared with the GenBank and RDP databases in order to find the closest neighbors and the respective type-strain. Genera Endozoicomonas (N = 32), Bacillus (N = 26), Shewanella (N = 17), Pseudovibrio (N = 12), and Ruegeria (N = 8) were dominant among the culturable microbiome of A. brasiliensis, corresponding to approx. 97% of all isolates (Complete list of isolates is on http://www.taxvibrio.lncc.br/). The three remaining isolates were closely related to Paenibacillus (N = 2) and Micrococcus (N = 1). Besides the taxonomic identification through comparisons of 16S rRNA sequences with databases, the 16S rRNA sequences were grouped using phylogenetic methods. The Endozoicomonas isolates presented sequence with 97.5% similarity towards the type strain of E. montiporae (Fig. 1; Fig. S1). The Shewanella isolates clustered all together, having at maximum 98% 16S rRNA sequence similarity towards S. irciniae type strain (Fig. 1; Fig. S2). Ruegeria isolates had 100% similarity with Ruegeria atlantica type strain (Fig. 1; Fig. S3). Part of Pseudovibrio isolates had 100% similarity with the sequence of P. denitrificans type strain and the remainder had identical 16S rRNA sequences with the P. asceidicola type strain (Fig. 1; Fig. S4). Paenibacillus isolates showed 99.8% similarity with P. illinoiensis type strain. Bacillus isolates had 100% similarity with the B. pumillus, B. safensis, B. thuringiensis, B. cereus, B. barbaricus and B. arsenicus type-strains (Fig. 1; Figs. S5–S6).
Figure 1.

Phylogenetic tree of partial 16S rRNA sequences (302 bp) of A. brasiliensis isolates and type strains sequences. The phylogeny reconstruction was based on neighbor-joining method and 1,000 bootstrap replications.
Antimicrobial activity
Antimicrobial activity was detected in 28.5% of the A. brasiliensis bacterial isolates of different species (Table 1). Bacillus subtilis test strain was susceptible to the supernatant of different A. brasiliensis bacterial isolates. The antimicrobial activity was widespread through different taxonomic groups of bacteria. Vibrio sinaloensis was susceptible to four isolates: two isolates closely related to Shewanella spongiae (i.e. Ab 105 and Ab 216), and two isolates closely related to Pseudovibrio ascidiaceicola (i.e. Ab 133 and Ab134).
Table 1. Bacterial isolates of A. brasiliensis showed antimicrobial activity.
Results of antimicrobial activity tests. Isolates belonging to different species inhibit the growth of B. subtilis.
| B. subtilis was susceptible to the following isolates | |
|---|---|
| Genus | Isolates |
| Endozoicomonas montiporae | Ab107/Ab227MC/Ab112/Ab213 |
| Shewanella irciniae | Ab114/Ab 101/Ab105/Ab109/Ab202/Ab216 |
| Ruegeria atlantica | Ab137/Ab138 |
| Pseudovibrio ascidiaceicola | Ab126/Ab132 |
| Pseudovibrio denitrificans | Ab127/Ab133/Ab134 |
| Paenibacillus illinoiensis | Ab232 |
| Bacillus pumillus | Ab 147/Ab148/Ab153/Ab161/ Ab166 |
| Bacillus cereus | Ab158/Ab160/Ab164/Ab233 |
| Bacillus nanhaiensis | Ab238 |
Discussion
As primitive Metazoans, marine sponges have established long-term associations with an array of microorganisms, which mainly include bacteria and cyanobacteria (Taylor et al., 2007). In the present investigation, we observed that all bacterial genera found among the culturable heterotrophic community of A. brasiliensis have already been isolated from other marine sponges, but, the present combination of bacterial genera was never reported within a single marine sponge. Comparisons between culturable bacterial communities from marine sponges are difficult to address since differences in media and culture conditions can significantly influence the isolation and cultivability of distinct bacterial isolates (Sipkema et al., 2011). In spite of this, it seems that some genera are commonly found among marine sponges, such as Pseudovibrio (Enticknap et al., 2006; Mohamed et al., 2008; Menezes et al., 2010; Santos et al., 2010; Flemer et al., 2011; Bruck, Reed & McCarthy, 2012; Margassery et al., 2012), Bacillus (Hentschel et al., 2001; Webster et al., 2001; Pabel et al., 2003; Lafi, Garson & Fuerst, 2005; Zhu, Li & Wang, 2008; Bruck et al., 2010; Devi et al., 2010; Santos et al., 2010; Flemer et al., 2011; Phelan et al., 2011; Bruck, Reed & McCarthy, 2012; Margassery et al., 2012), and Ruegeria (Mohamed et al., 2008; Menezes et al., 2010; Bruck, Reed & McCarthy, 2012; Margassery et al., 2012; Esteves et al., 2013; Haber & Ilan, 2013). Such bacterial genera can be found in distinct marine sponge species collected at distant geographic areas and with low phylogenetic relationship. On the other hand, Endozoicomonas, usually referred to as Spongiobacter (Mohamed et al., 2008; Flemer et al., 2011), and Shewanella (Flemer et al., 2011; Margassery et al., 2012; Haber & Ilan, 2013) are bacterial genera less frequently found within marine sponges (Table S1).
Endozoicomonas isolates obtained in this investigation were closely related to those isolates from Australian sponges and corals. Similarly, Shewanella, Ruegeria, Pseudovibrio, Paenibacillus, Bacillus and Micrococcus isolates grouped with related isolates obtained from different sponge species collected at different geographic locations (Figs. S1–S7).
The diversity and abundance of the genera detected through culture-dependent and culture-independent means have limited overlap
The diversity of culturable bacterial isolates recovered from A. brasiliensis presented similarity to the A. brasiliensis metagenomes previously analyzed by us (Trindade-Silva et al., 2012). For example, Pseudovibrio (N = 5), Ruegeria (N = 23), Shewanella (N = 16), Paenibacillus (N = 1) and Bacillus (N = 1) were observed in the metagenomic sequences analyzed previously, whereas Endozoicomonas and Micrococcus were not detected in the metagenomes of A. brasiliensis. In addition, we identified Pseudovibrio, Ruegeria, Shewanella, Paenibacillus, Bacillus, and Micrococcus in sequence libraries of polyketide synthase (PKS) genes originated from A. brasiliensis (Trindade-Silva et al., 2013), indicating some overlap between the culture-dependent and culture-independent studies. It demonstrates that suitable culture conditions can recover some of the bacterial genera detected by metagenomic analysis. Comparisons between culture-dependent and culture-independent approaches suggest that novel culturing approaches are needed in order to cultivate the missing bacterial diversity (Lavy et al., 2014).
The culturable heterotrophic bacteria of A. brasiliensis comprise well known antibiotic producers
Bacterial isolates belonging to the genera Endozoicomonas, Pseudovibrio and Paenibacillus are known for their ability to produce antibiotics. Pseudovibrio isolates originated from marine invertebrates such as tunicates (Sertan-de Guzman et al., 2007; Riesenfeld, Murray & Baker, 2008), corals (Rypien, Ward & Azam, 2010) and sponges showed antimicrobial activity (Santos et al., 2010; O’ Halloran et al., 2011). Pseudovibrio isolates from corals have antagonist effect against V. coralliilyticus (Rypien, Ward & Azam, 2010). In our study, we demonstrate that Pseudovibrio isolates of A. brasiliensis showed antimicrobial activity against Bacillus subtilis and Vibrio sinaloensis.
Bourne et al. (2008) found a correlation between the presence of Endozoicomonas (= Spongiobacter) and the health of corals. The authors argue that Endozoicomonas may be excluding potential pathogenic microorganisms from the holobiont. Mohamed and coworkers (2008) showed that an Endozoicomonas-related isolate from sponges produce a repertoire of N-acyl homoserine lactones. These quorum-sensing signaling metabolites may help delineate and structure the microbial communities in sponges. In the present study, several Endozoicomonas isolates showed antimicrobial activity against B. subtilis (Table 1). Endozoicomonas may play an ecological role in marine holobionts, shaping the associated microbial community through antimicrobial substances and signaling molecules.
Paenibacillus isolates belong to the microbial community of sponges, although they are found less abundantly than Bacillus (Phelan et al., 2011). The physiological role of sporeformers within sponge tissues is still unknown (Phelan et al., 2011), but probiotic activity was verified in Bacillus cereus and Paenibacillus sp. against Vibrio spp., conferring high levels of survival of shrimp larvae (Ravi et al., 2007). Additionally, antimicrobial activity was already detected in extracts of Paenibacillus isolates (Romanenko et al., 2013).
Micrococcus was the only Actinobacteria represented in the culturable community of A. brasiliensis, with a single isolate. Actinobacteria is a bacterial phylum commonly observed in invertebrate-associated microbial communities. Moreover, Actinobacteria comprises some of the most ingenious producers of potent antibiotics (Sun et al., 2010). Previous studies showed more diverse and abundant Actinobacteria communities in marine invertebrates than we recovered from A. brasiliensis. However, specific Actinobacteria growth conditions and nutrient requirements were not applied in the present study, which are normally required for enriching the isolation of representative isolates of this taxonomic group (Jiang et al., 2008; Sun et al., 2010). The diversity of actinomycetes recovered from A. brasiliensis is possibly underestimated; however, the microbes associated with A. brasiliensis may be providing protection against pathogens and keeping a healthy interaction with the symbiont community.
Conclusions
The sponge A. brasiliensis yielded a diverse culturable microbial community, including Endozoicomonas, Shewanella, Ruegeria, Pseudovibrio, Bacillus, and Paenibacillus isolates with high potential for antibiotic production. The compounds with antibiotic activity that can be explored as sources of natural products can also be those delineating the taxonomic composition of the associated microbial community. Genomic and post-genomic features of the antibiotic bacterial producers will enable to unveil the genes involved in the antibiotic production and define the chemical nature of the antibiotics. In addition, future studies will investigate whether antibiotics produced by microbes associated with A. brasiliensis may serve as chemical defenses in order to control the diversity and abundance of potential environmental pathogens.
Supplemental Information
Phylogenetic tree of partial 16S rRNA sequences of Endozoicomonas isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 202.
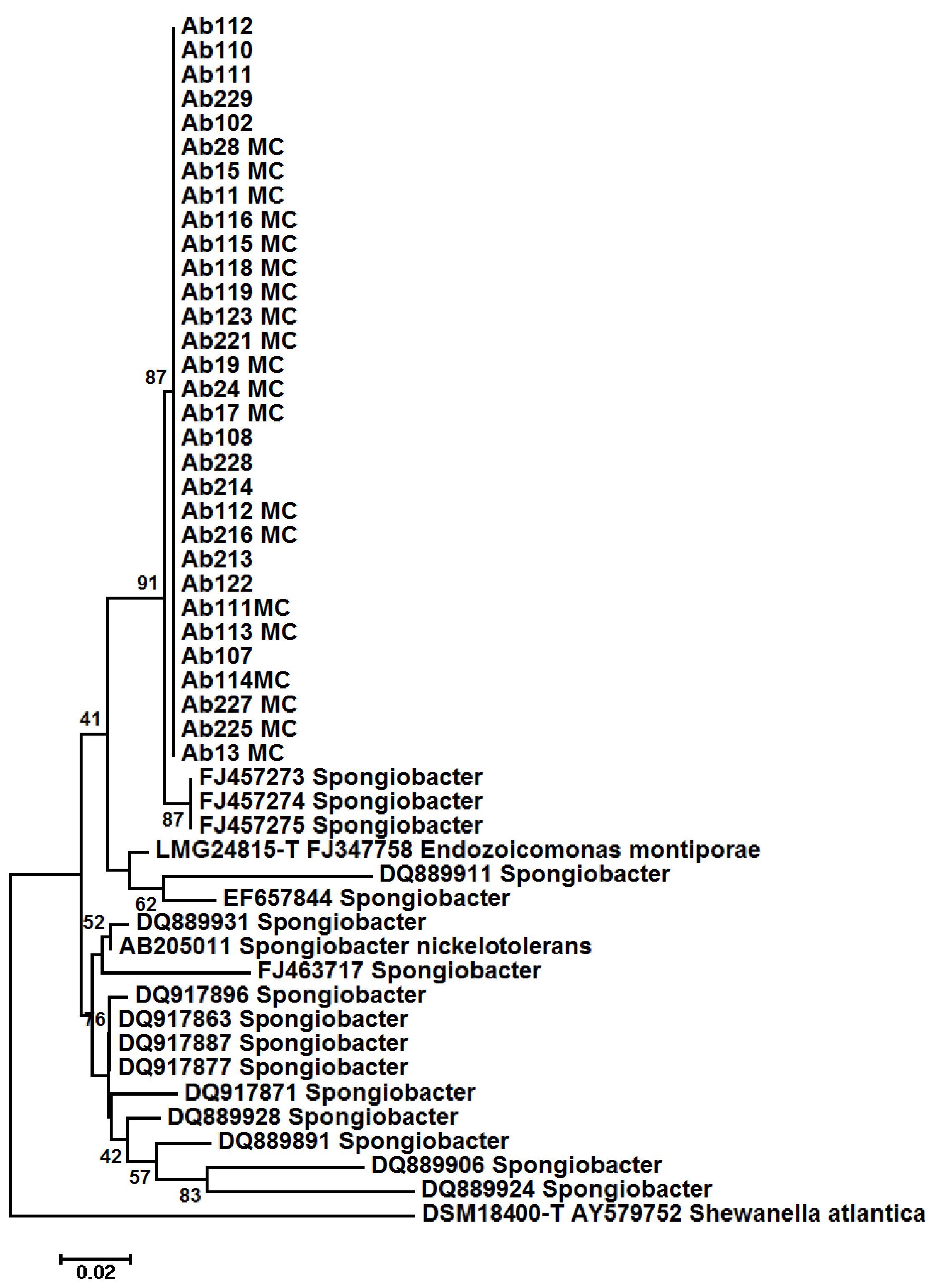{kind=link}
Phylogenetic tree of partial 16S rRNA sequences of Shewanella isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 404.
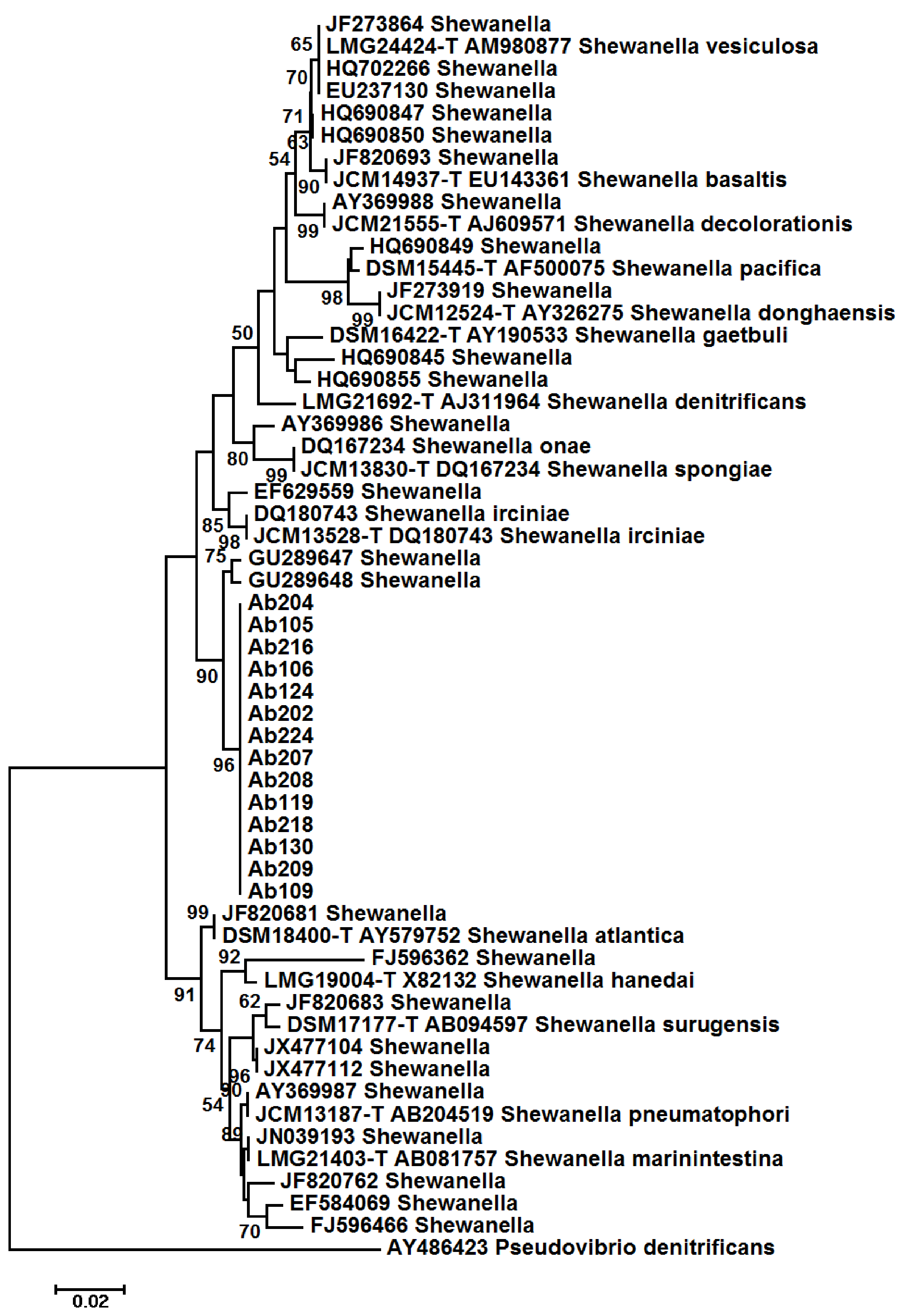{kind=link}
Phylogenetic tree of partial 16S rRNA sequences of Ruegeria isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 284.
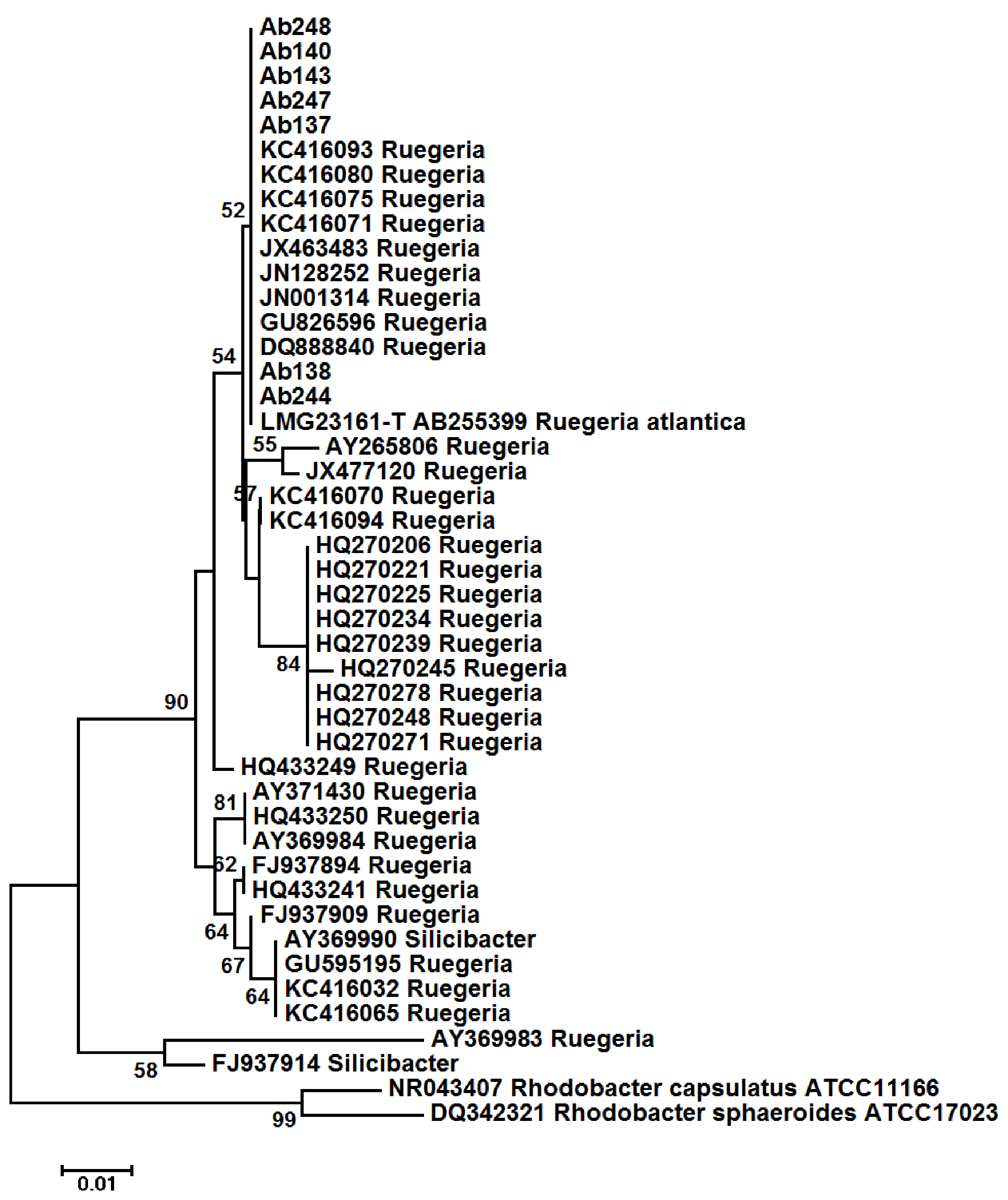{kind=link}
Phylogenetic tree of partial 16S rRNA sequences of Pseudovibrio isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 365.
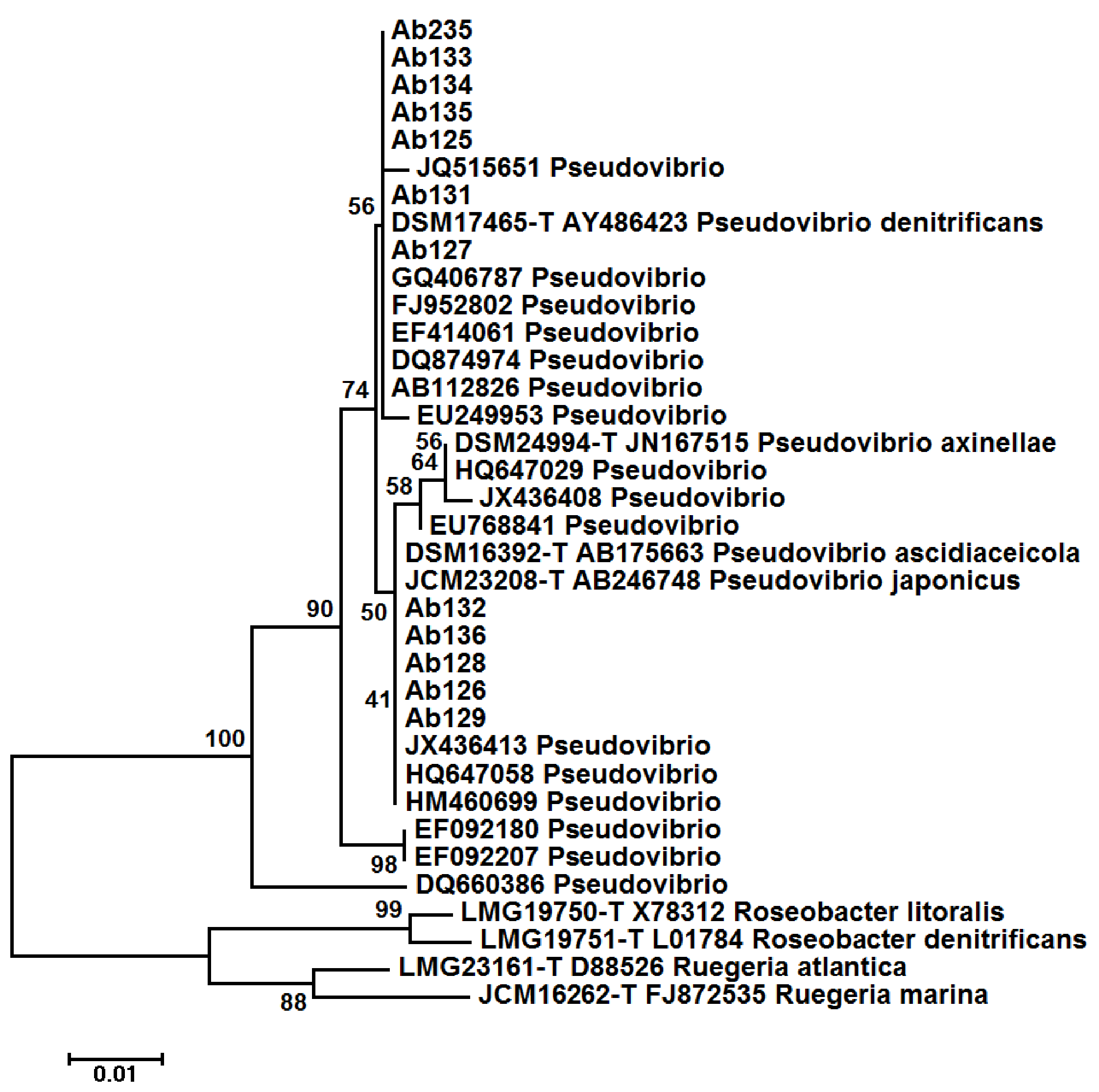{kind=link}
Phylogenetic tree of partial 16S rRNA sequences of Paenibacillus isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 533.
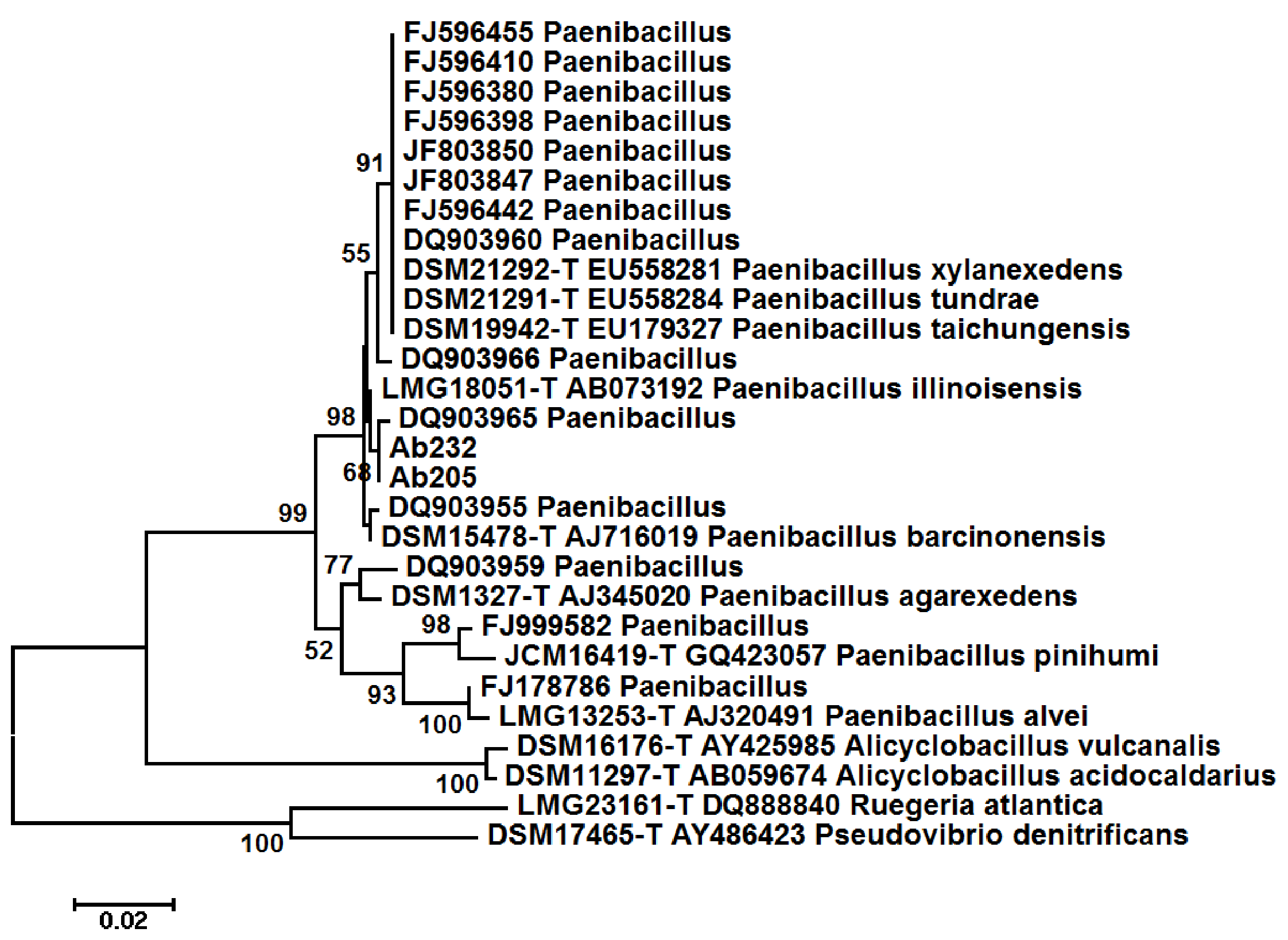{kind=link}
Phylogenetic tree of partial 16S rRNA sequences of Bacillus isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 407.
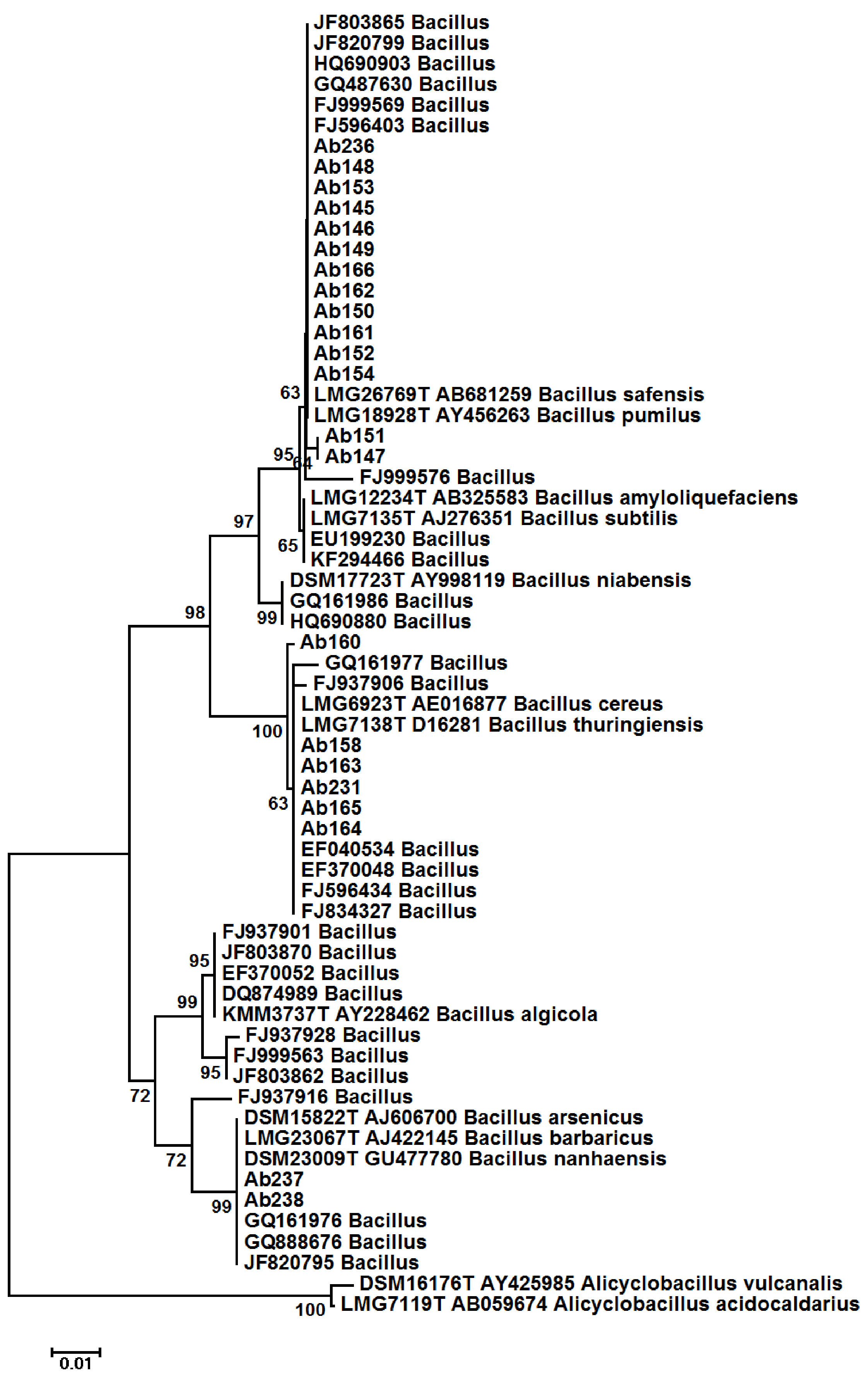{kind=link}
Phylogenetic tree of partial 16S rRNA sequences of Micrococcus isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 427.
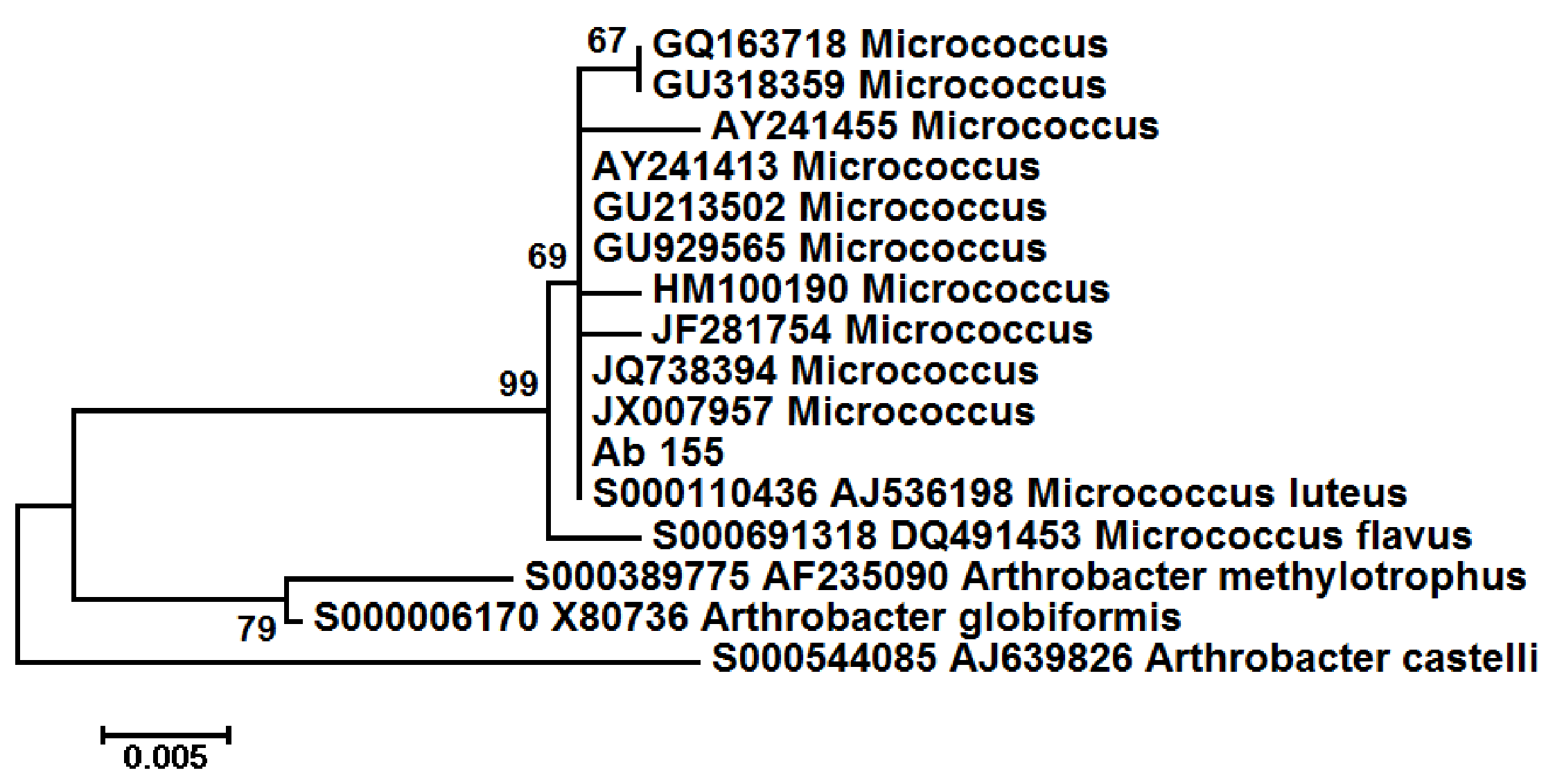{kind=link}
Genera of bacteria found in A. brasiliensis. Other sponge hosts where these genera were also found are presented with their references and locality of sample collection.
Raw data
Funding Statement
We received financial support from CAPES, CNPq (post-doctoral scholarship PDJ160605/2012-1), FAPERJ and FAPESP (grant 2010/50190-2 to RGSB and a post-doctoral scholarship 2009/11612-1). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
Fabiano Thompson is an Academic Editor for PeerJ. The authors declare there are no competing interests.
Author Contributions
Cintia P.J. Rua conceived and designed the experiments, performed the experiments, analyzed the data, wrote the paper, prepared figures and/or tables.
Amaro E. Trindade-Silva conceived and designed the experiments, performed the experiments, reviewed drafts of the paper.
Luciana R. Appolinario and Tainá M. Venas performed the experiments.
Gizele D. Garcia conceived and designed the experiments, performed the experiments.
Lucas S. Carvalho and Alinne Lima performed the experiments, analyzed the data.
Ricardo Kruger contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Renato C. Pereira and Roberto G.S. Berlinck reviewed drafts of the paper.
Rogério A.B. Valle and Cristiane C. Thompson analyzed the data.
Fabiano Thompson conceived and designed the experiments, analyzed the data, contributed reagents/materials/analysis tools, reviewed drafts of the paper.
DNA Deposition
References
- Altschul et al. (1990).Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. Journal of Molecular Biology. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- Berlinck et al. (2004).Berlinck RGS, Hajdu E, da Rocha RM, de Oliveira J, Hernandez ILC, Seleghim MHR, Granato AC, de Almeida EVR, Nunez CV, Muricy G, Peixinho S, Pessoa C, Moraes MO, Cavalcanti BC, Nascimento GGF, Thiemann O, Silva M, Souza AO, Silva CL, Minarini PRR. Challenges and rewards of research in marine natural products chemistry in Brazil. Journal of Natural Products. 2004;67:510–522. doi: 10.1021/np0304316. [DOI] [PubMed] [Google Scholar]
- Blunt et al. (2012).Blunt JW, Copp BR, Keyzers RA, Munro MHG, Prinsep MR. Marine natural products. Natural Product Reports. 2012;29:144–222. doi: 10.1039/c2np00090c. [DOI] [PubMed] [Google Scholar]
- Bourne et al. (2008).Bourne D, Iida Y, Uthicke S, Carolyn Smith-Keune C. Changes in coral-associated microbial communities during a bleaching event. The ISME Journal. 2008;2:350–363. doi: 10.1038/ismej.2007.112. [DOI] [PubMed] [Google Scholar]
- Brady (2007).Brady SF. Construction of soil environmental DNA cosmid libraries and screening for clones that produce biologically active small molecules. Nature Protocols. 2007;2:1297–1305. doi: 10.1038/nprot.2007.195. [DOI] [PubMed] [Google Scholar]
- Bruck et al. (2010).Bruck WM, Bruck TB, Self WT, Reed JK, Nitecki SS, McCarthy PJ. Comparison of the anaerobic microbiota of deep-water Geodia spp. and sandy sediments in the Straits of Florida. The ISME Journal. 2010;4:686–699. doi: 10.1038/ismej.2009.149. [DOI] [PubMed] [Google Scholar]
- Bruck, Reed & McCarthy (2012).Bruck WM, Reed JK, McCarthy PJ. The bacterial community of the lithistid sponge Discodermia spp. as determined by cultivation and culture-independent methods. Marine Biotechnology. 2012;14:762–773. doi: 10.1007/s10126-012-9443-6. [DOI] [PubMed] [Google Scholar]
- Devi et al. (2010).Devi P, Wahidullah S, Rodrigues C, Souza LD. The sponge-associated bacterium Bacillus licheniformis SAB1: a source of antimicrobial compounds. Marine Drugs. 2010;8:1203–1212. doi: 10.3390/md8041203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enticknap et al. (2006).Enticknap JJ, Kelly M, Peraud O, Hill RT. Characterization of a culturable alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. Applied and Environmental Microbiology. 2006;72:3724–3732. doi: 10.1128/AEM.72.5.3724-3732.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Esteves et al. (2013).Esteves AI, Hardoim CC, Xavier JR, Goncalves JM, Costa R. Molecular richness and biotechnological potential of bacteria cultured from Irciniidae sponges in the north-east Atlantic. FEMS Microbiology Ecology. 2013;85:519–536. doi: 10.1111/1574-6941.12140. [DOI] [PubMed] [Google Scholar]
- Fieseler et al. (2004).Fieseler L, Horn M, Wagner M, Hentschel U. Discovery of the novel candidate phylum “Poribacteria” in marine sponges. Applied and Environmental Microbiology. 2004;70:3724–3732. doi: 10.1128/AEM.70.6.3724-3732.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flemer et al. (2011).Flemer B, Kennedy J, Margassery LM, Morrissey JP, O’Gara F, Dobson AD. Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. Journal of Applied Microbiology. 2011;112:289–301. doi: 10.1111/j.1365-2672.2011.05211.x. [DOI] [PubMed] [Google Scholar]
- Fontana (2006).Fontana A. Biogenetic proposals and biosynthetic studies on secondary metabolites of opisthobranch molluscs. In: Cimino G, Gavagnin M, editors. Molluscs from chemo-ecological study to biotechnological application. Berlin, Heidelberg: Springer-Verlag; 2006. [Google Scholar]
- Graça et al. (2013).Graça AP, Bondoso J, Gaspar H, Xavier JR, Monteiro MC, de la Cruz M, Oves-Costales D, Vicente F, Lage OM. Antimicrobial activity of heterotrophic bacterial communities from the marine sponge Erylus discophorus (Astrophorida, Geodiidae) PLoS ONE. 2013;8:e419. doi: 10.1371/journal.pone.0078992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guimaraens & Coutinho (1996).Guimaraens MA, Coutinho R. Spatial and temporal variation of benthic marine algae at the Cabo Frio upwelling region, Rio de Janeiro, Brazil. Aquatic Botany. 1996;52:283–299. doi: 10.1016/0304-3770(95)00511-0. [DOI] [Google Scholar]
- Haber & Ilan (2013).Haber M, Ilan M. Diversity and antibacterial activity of bacteria cultured from Mediterranean Axinella spp. sponges. Journal of Applied Microbiology. 2013;116:519–532. doi: 10.1111/jam.12401. [DOI] [PubMed] [Google Scholar]
- Hentschel et al. (2012).Hentschel U, Piel J, Degnan SM, Taylor MW. Genomic insights into the marine sponge microbiome. Nature Reviews Microbiology. 2012;10:641–654. doi: 10.1038/nrmicro2839. [DOI] [PubMed] [Google Scholar]
- Hentschel et al. (2001).Hentschel U, Schmid M, Wagner M, Fieseler L, Gernert C, Hacker J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiology Ecology. 2001;35:305–312. doi: 10.1111/j.1574-6941.2001.tb00816.x. [DOI] [PubMed] [Google Scholar]
- Hoffmann et al. (2005).Hoffmann F, Larsen O, Thiel V, Rapp HT, Pape T, Michaelis W, Reitner J. An anaerobic world in sponges. Geomicrobiology Journal. 2005;22:1–10. doi: 10.1080/01490450590922505. [DOI] [Google Scholar]
- Jiang et al. (2008).Jiang S, Li X, Zhang L, Sun W, Dai S, Xie L, Liu Y, Lee KJ. Culturable actinobacteria isolated from marine sponge Iotrochota sp. Marine Biology. 2008;153:945–952. doi: 10.1007/s00227-007-0866-y. [DOI] [Google Scholar]
- Lafi, Garson & Fuerst (2005).Lafi FF, Garson MJ, Fuerst JA. Culturable bacterial symbionts isolated from two distinct sponge species (Pseudoceratina clavata and Rhabdastrella globostellata) from the Great Barrier Reef display similar phylogenetic diversity. Microbial Ecology. 2005;50:213–220. doi: 10.1007/s00248-004-0202-8. [DOI] [PubMed] [Google Scholar]
- Lane (1991).Lane DJ. 16S/23S rRNA sequencing. In: Stackebrandt EGM, editor. Nucleic acid techniques in bacterial systematics. New York: John Wiley & Sons; 1991. pp. 115–175. [Google Scholar]
- Lavy et al. (2014).Lavy A, Keren R, Haber M, Schwartz I, Ilan M. Implementing sponge physiological and genomic information to enhance the diversity of its culturable associated bacteria. FEMS Microbiology Ecology. 2014;87:486–502. doi: 10.1111/1574-6941.12240. [DOI] [PubMed] [Google Scholar]
- Lee et al. (2006).Lee OO, Lau SC, Tsoi MM, Li X, Plakhotnikova I, Dobretsov S, Wu MC, Wong PK, Weinbauer M, Qian PY. Shewanella irciniae sp. nov., a novel member of the family Shewanellaceae, isolated from the marine sponge Ircinia dendroides in the Bay of Villefranche, Mediterranean Sea. International Journal of Systematic and Evolutionary Microbiology. 2006;56:2871–2877. doi: 10.1099/ijs.0.64562-0. [DOI] [PubMed] [Google Scholar]
- Margassery et al. (2012).Margassery LM, Kennedy J, O’Gara F, Dobson AD, Morrissey JP. Diversity and antibacterial activity of bacteria isolated from the coastal marine sponges Amphilectus fucorum and Eurypon major. Letters in Applied Microbiology. 2012;55:2–8. doi: 10.1111/j.1472-765X.2012.03256.x. [DOI] [PubMed] [Google Scholar]
- Menezes et al. (2010).Menezes CB, Bonugli-Santos RC, Miqueletto PB, Passarini MR, Silva CH, Justo MR, Leal RR, Fantinatti-Garboggini F, Oliveira VM, Berlinck RG, Sette LD. Microbial diversity associated with algae, ascidians and sponges from the north coast of Sao Paulo state, Brazil. Microbiology Research. 2010;165:466–482. doi: 10.1016/j.micres.2009.09.005. [DOI] [PubMed] [Google Scholar]
- Mohamed et al. (2008).Mohamed NM, Cicirelli EM, Kan J, Chen F, Fuqua C, Hill RT. Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environmental Microbiology. 2008;10:75–86. doi: 10.1111/j.1462-2920.2008.01704.x. [DOI] [PubMed] [Google Scholar]
- Muricy et al. (2011).Muricy G, Lopes DA, Hajdu E, Carvalho MS, Moraes FC, Klautau M, Menegola C, Pinheiro U. Catalogue of Brazilian porifera. Rio de Janeiro: Museu Nacional; 2011. [Google Scholar]
- Noyer, Thomas & Becerro (2011).Noyer C, Thomas OP, Becerro MA. Patterns of chemical diversity in the Mediterranean sponge Spongia lamella. PLoS ONE. 2011;6:e419. doi: 10.1371/journal.pone.0020844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’ Halloran et al. (2011).O’ Halloran JA, Barbosa TM, Morrissey JP, Kennedy J, O’ Gara F, Dobson ADW. Diversity and antimicrobial activity of Pseudovibrio spp. from Irish marine sponges. Applied Microbiology. 2011;110:1495–1508. doi: 10.1111/j.1365-2672.2011.05008.x. [DOI] [PubMed] [Google Scholar]
- Öztürk et al. (2013).Öztürk B, de Jaeger L, Smidt H, Sipkema D. Culture-dependent and independent approaches for identifying novel halogenases encoded by Crambe crambe (marine sponge) microbiota. Scientific Reports. 2013;3 doi: 10.1038/srep02780. Article 2780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pabel et al. (2003).Pabel CT, Vater J, Wilde C, Franke P, Hofemeister J, Adler B, Bringmann G, Hacker J, Hentschel U. Antimicrobial activities and matrix-assisted laser desorption/ionization mass spectrometry of Bacillus isolates from the marine sponge Aplysina aerophoba. Marine Biotechnology. 2003;5:424–434. doi: 10.1007/s10126-002-0088-8. [DOI] [PubMed] [Google Scholar]
- Phelan et al. (2011).Phelan RW, O’Halloran JA, Kennedy J, Morrissey JP, Dobson AD, O’Gara F, Barbosa TM. Diversity and bioactive potential of endospore-forming bacteria cultured from the marine sponge Haliclona simulans. Journal of Applied Microbiology. 2011;112:65–78. doi: 10.1111/j.1365-2672.2011.05173.x. [DOI] [PubMed] [Google Scholar]
- Piel (2006).Piel J. Bacterial symbionts: prospects for the sustainable production of invertebrate-derived pharmaceuticals. Current Medicinal Chemistry. 2006;13:39–50. doi: 10.2174/092986706775197944. [DOI] [PubMed] [Google Scholar]
- Piel (2009).Piel J. Metabolites from symbiotic bacteria. Natural Product Reports. 2009;26:338–362. doi: 10.1039/b703499g. [DOI] [PubMed] [Google Scholar]
- Piel et al. (2004).Piel J, Hui DQ, Wen GP, Butzke D, Platzer M, Fusetani N, Matsunaga S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:16222–16227. doi: 10.1073/pnas.0405976101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Posada & Crandall (1998).Posada D, Crandall KA. MODELTEST: testing the model of DNA substitution. Bioinformatics. 1998;14:817–818. doi: 10.1093/bioinformatics/14.9.817. [DOI] [PubMed] [Google Scholar]
- Ravi et al. (2007).Ravi AV, Musthafa KS, Jegathammbal G, Kathiresan K, Pandian SK. Screening and evaluation of probiotics as a biocontrol agent against pathogenic vibrios in marine aquaculture. Letters in Applied Microbiology. 2007;45:219–223. doi: 10.1111/j.1472-765X.2007.02180.x. [DOI] [PubMed] [Google Scholar]
- Reveillaud et al. (2014).Reveillaud J, Maignien L, Eren AM, Huber JA, Apprill A, Sogin ML, Vanreusel A. Host-specificity among abundant and rare taxa in the sponge microbiome. The ISME Journal. 2014;8:1198–1209. doi: 10.1038/ismej.2013.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riesenfeld, Murray & Baker (2008).Riesenfeld CS, Murray AE, Baker BJ. Characterization of the microbial community and polyketide biosynthetic potential in the palmerolide-producing tunicate Synoicum adareanum. Journal of Natural Products. 2008;71:1812–1818. doi: 10.1021/np800287n. [DOI] [PubMed] [Google Scholar]
- Romanenko et al. (2013).Romanenko L, Tanaka N, Kalinovskaya N, Mikhailov V. Antimicrobial potential of deep surface sediment associated bacteria from the Sea of Japan. World Journal of Microbiology and Biotechnology. 2013;29:1169–1177. doi: 10.1007/s11274-013-1276-6. [DOI] [PubMed] [Google Scholar]
- Rypien, Ward & Azam (2010).Rypien KL, Ward JR, Azam F. Antagonistic interactions among coral-associated bacteria. Environmental Microbiology. 2010;12:28–39. doi: 10.1111/j.1462-2920.2009.02027.x. [DOI] [PubMed] [Google Scholar]
- Sacristán-Soriano et al. (2011).Sacristán-Soriano O, Banaigs B, Casamayor EO, Becerro MA. Exploring the links between natural products and bacterial assemblages in the sponge Aplysina aerophoba. Applied and Environmental Microbiology. 2011;77:862–870. doi: 10.1128/AEM.00100-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos et al. (2010).Santos OCS, Pontes PVML, Santos JFM, Muricy G, Giambiagi-deMarval M, Laport MS. Isolation, characterization and phylogeny of sponge-associated bacteria with antimicrobial activities from Brazil. Research in Microbiology. 2010;161:604–612. doi: 10.1016/j.resmic.2010.05.013. [DOI] [PubMed] [Google Scholar]
- Schneemann et al. (2011).Schneemann I, Wiese J, Kunz AL, Imhoff JF. Genetic approach for the fast discovery of phenazine producing bacteria. Marine Drugs. 2011;9:772–789. doi: 10.3390/md9050772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sertan-de Guzman et al. (2007).Sertan-de Guzman AA, Predicala RZ, Bernardo EB, Neilan BA, Elardo SP, Mangalindan GC, Tasdemir D, Ireland CM, Barraquio WL, Concepcion GP. Pseudovibrio denitrificans strain Z143-1, a heptylprodigiosin-producing bacterium isolated from a Philippine tunicate. FEMS Microbiology Letters. 2007;277:188–196. doi: 10.1111/j.1574-6968.2007.00950.x. [DOI] [PubMed] [Google Scholar]
- Sipkema et al. (2011).Sipkema D, Schippers K, Maalcke WJ, Yang Y, Salim S, Blanch HW. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (gellius) sp. Applied and Environmental Microbiology. 2011;77:2130–2140. doi: 10.1128/AEM.01203-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun et al. (2010).Sun W, Dai S, Jiang S, Wang G, Liu G, Wu H, Li X. Culture-dependent and culture-independent diversity of Actinobacteria associated with the marine sponge Hymeniacidon perleve from the South China Sea. Antonie van Leeuwenhoek. 2010;98:65–75. doi: 10.1007/s10482-010-9430-8. [DOI] [PubMed] [Google Scholar]
- Tamura et al. (2011).Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution. 2011;28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor et al. (2007).Taylor MW, Radax R, Steger D, Wagner M. Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiology and Molecular Biology Reviews. 2007;71:295–347. doi: 10.1128/MMBR.00040-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor et al. (2012).Taylor MW, Tsai P, Simister RL, Deines P, Botte E, Ericson G, Schmitt S, Webster NS. Sponge-specific’ bacteria are widespread (but rare) in diverse marine environments. The ISME Journal. 2012;7:438–443. doi: 10.1038/ismej.2012.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson et al. (2001).Thompson FL, Hoste B, Vandemeulebroecke K, Swings J. Genomic diversity amongst Vibrio isolates from different sources determined by fluorescent amplified fragment length polymorphism. Systematic and Applied Microbiology. 2001;24:520–538. doi: 10.1078/0723-2020-00067. [DOI] [PubMed] [Google Scholar]
- Torres et al. (2000).Torres YR, Berlinck RGS, Magalhaes A, Schefer AB, Ferreira AG, Hajdu E, Muricy G. Arenosclerins A-C and haliclonacyclamine E, new tetracyclic alkaloids from a Brazilian endemic haplosclerid sponge Arenosclera brasiliensis. Journal of Natural Products. 2000;63:1098–1105. doi: 10.1021/np9905618. [DOI] [PubMed] [Google Scholar]
- Trindade-Silva et al. (2012).Trindade-Silva AE, Rua C, Silva GGZ, Dutilh BE, Moreira APB, Edwards RA, Hajdu E, Lobo-Hajdu G, Vasconcelos AT, Berlinck RGS, Thompson FL. Taxonomic and functional microbial signatures of the endemic marine sponge Arenosclera brasiliensis. PLoS ONE. 2012;7:e419. doi: 10.1371/journal.pone.0039905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trindade-Silva et al. (2013).Trindade-Silva AE, Rua CPJ, Andrade BGN, Vicente ACP, Silva GGZ, Berlinck RGS, Thompson F. Polyketide synthase gene diversity within the microbiome of the sponge Arenosclera brasiliensis, endemic to the southern Atlantic Ocean. Applied and Environmental Microbiology. 2013;5:1598–1605. doi: 10.1128/AEM.03354-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang (2006).Wang G. Diversity and biotechnological potential of the sponge-associated microbial consortia. Journal of Industrial Microbiology and Biotechnology. 2006;33:545–551. doi: 10.1007/s10295-006-0123-2. [DOI] [PubMed] [Google Scholar]
- Wang et al. (2007).Wang Q, Garrity GM, Tiedje JM, Cole JR. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Applied and Environmental Microbiology. 2007;73:5261–5267. doi: 10.1128/AEM.00062-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster et al. (2001).Webster NS, Wilson KJ, Blackall LL, Hill RT. Phylogenetic diversity of bacteria associated with the marine sponge Rhopaloeides odorabile. Applied and Environmental Microbiology. 2001;67:434–444. doi: 10.1128/AEM.67.1.434-444.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weisz et al. (2007).Weisz J, Hentschel U, Lindquist N, Martens C. Linking abundance and diversity of sponge-associated microbial communities to metabolic differences in host sponges. Marine Biology. 2007;152:475–483. doi: 10.1007/s00227-007-0708-y. [DOI] [Google Scholar]
- Westerdahl et al. (1991).Westerdahl A, Olsson JC, Kjelleberg S, Conway PL. Isolation and characterization of turbot (Scophtalmus maximus)-associated bacteria with inhibitory effects against Vibrio anguillarum. Applied and Environmental Microbiology. 1991;57:2223–2228. doi: 10.1128/aem.57.8.2223-2228.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoneshigue (1985).Yoneshigue Y. Ph.D. thesis. 1985. Taxonomie et écologie des algues marines dans la region de Cabo Frio (Rio de Janeiro, Brésil) [Google Scholar]
- Zhu, Li & Wang (2008).Zhu P, Li QZ, Wang GY. Unique microbial signatures of the alien Hawaiian marine sponge Suberites zeteki. Microbial Ecology. 2008;55:406–414. doi: 10.1007/s00248-007-9285-3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Phylogenetic tree of partial 16S rRNA sequences of Endozoicomonas isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 202.
Phylogenetic tree of partial 16S rRNA sequences of Shewanella isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 404.
Phylogenetic tree of partial 16S rRNA sequences of Ruegeria isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 284.
Phylogenetic tree of partial 16S rRNA sequences of Pseudovibrio isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 365.
Phylogenetic tree of partial 16S rRNA sequences of Paenibacillus isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 533.
Phylogenetic tree of partial 16S rRNA sequences of Bacillus isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 407.
Phylogenetic tree of partial 16S rRNA sequences of Micrococcus isolates, type strains sequences and database sequences of bacterial strains isolated from marine invertebrates. The numbers of sites used in the phylogenetic reconstructions were 427.
Genera of bacteria found in A. brasiliensis. Other sponge hosts where these genera were also found are presented with their references and locality of sample collection.
Raw data