

Abstract
PrBP/δ, encoded by the Pde6d gene, is an isoprenyl-binding protein that regulates trafficking of isoprenylated proteins, such as PDE6 and GRK1, from photoreceptor inner segments to outer segments. Trafficking of PDE6 and GRK1 to photoreceptor outer segments is impeded in Pde6d knockout mice. In Pde6d-/-cones, PDE6 and GRK1 are nearly undetectable and the b-wave amplitudes of photopic ERGs in Pde6d-/- mice are reduced by over 50%. We reported recently that UNC119, a homolog of PrBP/δ highly expressed in photoreceptors, functions as an acyl-binding protein and regulates transport of G-proteins in sensory neurons. Since both PrBP/δ and UNC119 regulate peripheral protein trafficking in photoreceptors, we generated Pde6d;Unc119 double knockout mice in order to study how PrBP/δ and UNC119 may interact. Surprisingly, knockout of Unc119 partially reversed the transport defect of GRK1 in cone photoreceptors caused by deletion of Pde6d, and the b-wave amplitudes of photopic ERGs in the double knockout mice were significantly higher than those in the Pde6d-/- mice. These results suggest that cone transport of isoprenylated and acylated proteins is interdependent.
Keywords: PrBP/δ, Pde6d, UNC119, photoreceptors, GRK1, PDE6, ERG
62.1 Introduction
PrBP/δ, encoded by the Pde6d gene, is a small polypeptide consisting of 150 amino acids in mammals with an apparent molecular weight of ∼17kD [1]. As PrBP/δ co-purified with photoreceptor PDE6, it was thought initially to be a fourth subunit of rod PDE6 [2]. Many more PrBP/δ-interacting proteins, identified by yeast-two hybrid screening (e.g., GRK1, Ras, and other small GTPases in the Ras family [3-5]), suggested a more general function of PrBP/δ. Close examination of PrBP/δ-interacting proteins revealed that most share a common feature, a C-terminal isoprenyl group. The FRET assay shows that PrBP/δ has distinct affinities for farnesyl and geranylgeranyl moieties, binding farnesyl (kd = 0.7 μM) more tightly than geranylgeranyl (kd = 19 μM) [5]. Transport of isoprenylated proteins (GRK1, rod PDE6, and cone PDE6) to photoreceptor outer segments was impeded in Pde6d knockout photoreceptors, resulting in a slow degeneration of cones followed by degeneration of rods, a phenotype similar to human patients with cone-rod dystrophy [6].
UNC119, first discovered in C.elegans, shares 30-40% sequence similarity with PrBP/δ [7]. UNC119 is highly expressed in the synaptic termini of photoreceptors, and to a less extent in the inner segments [8]. Knockout of the Unc119 gene in mouse led to slow retinal degeneration starting at 6 months postnatally, a phenotype similar to human RP (Retinitis Pigmentosa) [8], although the mechanism of degeneration is unclear. We recently proposed that UNC119 is an acyl-binding protein. UNC119 binds the acylated N-terminal peptide of transducin α with a kd of 0.5 μM [9]. UNC119 can extract transducin from retina membranes in the presence of GTP, and is required for efficient return of transducin to photoreceptor outer segments after light-induced translocation of transducin from outer segments to inner segments during light adaptation, suggesting that UNC119 regulates trafficking of transducin in photoreceptors [9].
In this study, we generated PrBP/δ and UNC119 double knockout mice and observed that GRK1 is up-regulated in the mutant cone photoreceptors. With more GRK1 present in the cone photoreceptor cells, photopic ERGs reflecting cone function exhibit higher responses in the double knockout mice compared with those in the Pde6d single knockout mice.
62.2 Materials and methods
62.2.1. Mouse breeding genotyping
Procedures for the animal experiments were approved by the University of Utah IACUC and conformed to recommendations of the Association of Research for Vision and Ophthalmology. Animals were maintained in cyclic light (12 h light/12 h dark) conditions. Unc119-/- mice were provided by Dr. George Inana at the University of Miami. Pde6d;Unc119 double knockout mice were generated by mating Pde6d-/- mice to Unc119-/- mice. Genotyping of Pde6d-/- and Unc119-/- mice was described previously [6, 8].
62.2.2. Immunohistochemistry
Immunostaining of mouse retina sections was performed as described [10]. Retina cryosections were cut and immunolabeled using mouse monoclonal GRK1 antibody (G8, 1:1000, from Dr. Kris Palczewski, Case Western Reserve University) or rabbit polyclonal cone PDE6 antibody (!:500, from Dr. Tiansen Li, NEI/NIH).
62.2.3. Photopic Electroretinogram (ERG)
ERGs were recorded with a UTAS E-3000 (LKC Technologies, Inc.) as described [6, 11]. For photopic ERGs to record cone function, the mice were light-adapted under background light of 10 db (1.48 logcds·m−2) for 15 minutes. Single-flash responses were usually recorded at stimulus intensities of −4 db (−0.6 logcds·m−2) to 15 db (1.86 logcds·m−2).
62.3 Results
62.3.1. Knockout of the Unc119 gene in Pde6d-/- mice increases expression of GRK1 in photoreceptor outer segments
PrBP/δ and UNC119 each regulate transport of lipid-conjugated peripheral proteins to photoreceptor outer segments. Both, in turn, are regulated by the small GTPase ARL3, which controls cargo release from PrBP/δ and UNC119 [12, 13]. It remains unclear whether PrBP/δ and UNC119 coordinately regulate the transport of isoprenylated proteins and acylated protein to photoreceptor outer segments. We were interested to define the functional relationship of these two proteins by generating Pde6d and Unc119 double knockout mice. In the Pde6d-/-Unc119-/- mice, cone PDE6 is nearly undetectable, which is similar in the Pde6d single knockout (Fig. 1B). Expression of GRK1 in the Pde6d-/- and Pde6d-/-Unc119-/- mice is almost absent throughout the rods (Fig. 1A). However, expression of GRK1 is surprisingly increased in the cone outer segments, as compared to the Pde6d-/- cones with little GRK1 in COS (Fig. 1A).
Fig. 62.1.
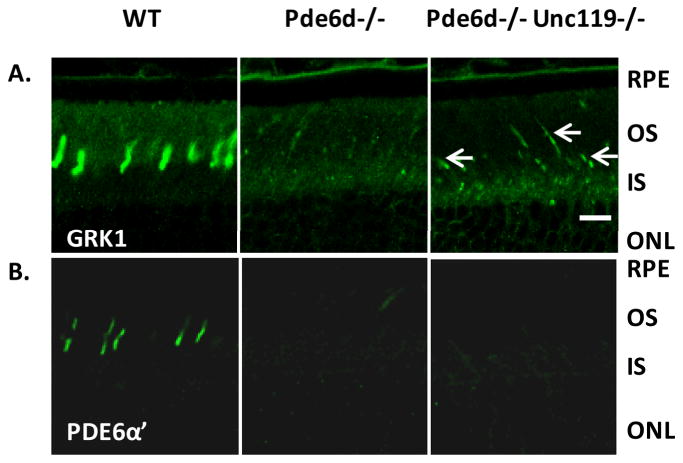
Immunolocalization of GRK1 and PDE6α′ in WT, Pde6d-/-, and Pde6d-/-Unc119-/- retinas. The eyes were co-embedded, and central retina sections were probed simultaneously using anti-GRK1 (A) and PDE6α′ (B). RPE, Retinal pigment epithelium; OS, outer segments; IS, inner segments; ONL, outer nuclear layer. Arrows points cone outer segments. Scale bar, 10 μm.
62.3.2. Knockout of the Unc119 gene in Pde6d-/- background increases cone photoresponses
GRK1 is a key enzyme to shut down the phototransduction cascade. GRK1 phosphorylates rhodopsin and cone opsins, thereby desensitizing rhodopsin and cone opsins following activation. In GRK1 gene knockout mice, both rod and cone photoreceptors showed slower recovery from photoactivation [14]. In Pde6d knockout mice, expression of GRK1 is significantly down-regulated and photoreceptors exhibit a phenotype of slower dark-adaption [6]. Lowered GRK1 in Pde6d-/- mice essentially delays desensitization of activated cone opsins and increase basal level of photoresponses under constant light, resulting in lower amplitude of photopic ERGs. In the Pde6d-/-Unc119-/- mice, the expression of GRK1 increases relative to that observed in Pde6d knockout mice. As expected, the amplitude of photopic ERGs in the Pde6d-/-Unc119-/- is higher than that in the Pde6d-/- mice; however, it is still lower than that in wild type mice (Fig. 2), which may be due to partial up-regulation of GRK1 and lower cone PDE6 expression in the double knockout.
Fig. 62.2.
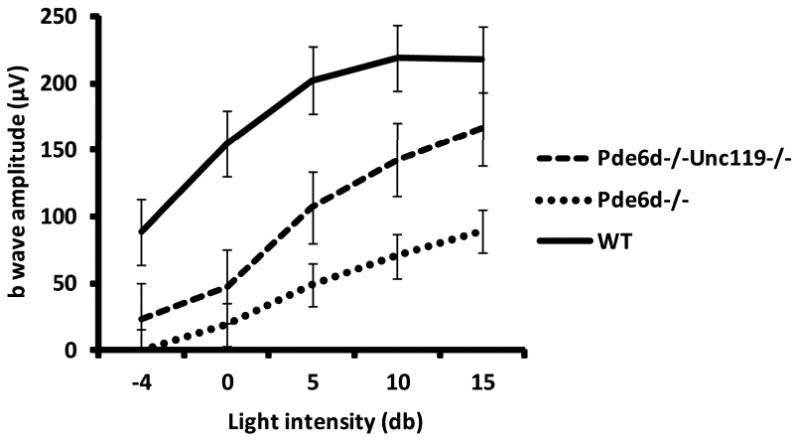
Photopic ERGs of WT, Pde6d-/-, and Pde6d-/-Unc119-/- mice. Photopic b-wave amplitudes are plotted as a function of light intensities. Error bars represent mean ± SD (n = 3).
62.4 Discussion
We show that deletion of the Unc119 gene in the Pde6d-/- background produced up-regulated expression of GRK1 in cone photoreceptors and consequently, cone b-wave photoresponses increased relative to that observed in Pde6d single knockout mice (Fig. 2). GRK1 is a peripheral membrane protein, and undergoes posttranslational isoprenylation at its C-terminus. PrBP/δ solubilizes GRK1 from membranes by binding the isoprenyl tail of GRK1, and forming a soluble complex to facilitate transport of GRK1. PrBP/δ also interacts with a small GTPase ARL3 [15], and GTP-bound ARL3 likely promotes release of GRK1 from PrBP/δ at the target membrane, analogous to release of farnesylated Rheb from PrBP/δ [12]. In contrast, UNC119 is an acyl-binding protein. Deletion of the UNC119 gene does not interfere with transport of GRK1 and other isoprenylated proteins [9]. However, UNC119 also interacts with ARL3, and GTP-bound ARL3 facilitates the release of acylated protein cargo from UNC119 [13]. Our result that the UNC119;Pde6d double knockout improves transport of GRK1 in cones is counter-intuitive, because ARL3 is not involved in regulating PrBP/δ cargo release when PrBP/δ is absent. However, in Pde6d knockout cones, residual GRK1 is present in cone outer segments, suggesting that GRK1 transport may be facilitated via alternative mechanisms. One possible mechanism is that other isoprenyl binding proteins may play a minor role in transport of GRK1 to outer segments, and one ‘candidate isoprenyl binding protein’ is RhoGDI that binds geranylgeranylated CDC42. Although RhoGDI is a possible candidate that may enhance transport of GRK1 to cone outer segments, we cannot exclude other isoprenyl-binding proteins as possible candidates, such as RabGDI; nor could we exclude possible common factors that play a role in transport pathways of both isoprenylated proteins and acylated proteins.
PrBP/δ and RhoGDI each contain a hydrophobic pocket sandwiched by two β-sheets and each can bind isoprenyl groups. It is possible that RhoGDI can accommodate isoprenyl tail of GRK1. Expression of Rho GTPase proteins in photoreceptors have been documented [16], suggesting that RhoGDI, an essential functional partner for Rho GTPase, is also expressed in photoreceptors. When PrBP/δ/GRK1 complex reaches its destination membrane, release of GRK1 requires interaction with GTP-bound ARL3. Similarly, RhoGDI/GRK1 complex may require ARL3 to discharge GRK1 from RhoGDI upon their arrival to the targeted membrane. ARL3 not only traffics isoprenylated proteins but also traffics acylated proteins, a process which is modulated by UNC119. Other factors besides ARL3 may share transport of isoprenylated and acylated proteins. Therefore, knockout of Unc119 gene will free at least some ARL3 and other unknown factors from the pathway of trafficking acylated proteins and enhance transport of GRK1 by the alternative transport pathway.
Future investigations will determine if rescue of GRK1 transport in the Pde6d;Unc119 double knockout mice would slow down the degeneration of cone photoreceptors. We will also investigate the expression profile of photoreceptor RhoGDI and test the binding affinity of RhoGDI to GRK1 and PDE6. This will address the role of RhoGDI in the observed rescue of GRK1 transport in Unc119;Pde6d double knockout mice.
Abbreviations
- RhoGDI
Rho GTPase Guanine nucleotide Dissociation Inhibitor
- ERG
electroretinogram
- COS
cone outer segment
References
- 1.Zhang H, Constantine R, Frederick JM, Baehr W. The prenyl-binding protein PrBP/δ: A chaperone participating in intracellular trafficking. Vision Res. 2012;75:19–25. doi: 10.1016/j.visres.2012.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gillespie PG, Prusti RK, Apel ED, Beavo JA. A soluble form of bovine rod photoreceptor phosphodiesterase has a novel 15-kDa subunit. J Biol Chem. 1989;264(21):12187–93. [PubMed] [Google Scholar]
- 3.Hillig RC, Hanzal-Bayer M, Linari M, Becker J, Wittinghofer A, Renault L. Structural and biochemical properties show ARL3-GDP as a distinct GTP binding protein. Structure. 2000;8(12):1239–45. doi: 10.1016/s0969-2126(00)00531-1. [DOI] [PubMed] [Google Scholar]
- 4.Hanzal-Bayer M, Renault L, Roversi P, Wittinghofer A, Hillig RC. The complex of Arl2-GTP and PDE delta: from structure to function. EMBO J. 2002;21(9):2095–106. doi: 10.1093/emboj/21.9.2095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zhang H, Liu XH, Zhang K, Chen CK, Frederick JM, Prestwich GD, Baehr W. Photoreceptor cGMP phosphodiesterase delta subunit (PDEdelta) functions as a prenyl-binding protein. J Biol Chem. 2004;279(1):407–13. doi: 10.1074/jbc.M306559200. [DOI] [PubMed] [Google Scholar]
- 6.Zhang H, Li S, Doan T, Rieke F, Detwiler PB, Frederick JM, Baehr W. Deletion of PrBP/delta impedes transport of GRK1 and PDE6 catalytic subunits to photoreceptor outer segments. Proc Natl Acad Sci U S A. 2007;104(21):8857–62. doi: 10.1073/pnas.0701681104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Maduro M, Pilgrim D. Identification and cloning of unc-119, a gene expressed in the Caenorhabditis elegans nervous system. Genetics. 1995;141(3):977–88. doi: 10.1093/genetics/141.3.977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ishiba Y, Higashide T, Mori N, Kobayashi A, Kubota S, McLaren MJ, Satoh H, Wong F, Inana G. Targeted inactivation of synaptic HRG4 (UNC119) causes dysfunction in the distal photoreceptor and slow retinal degeneration, revealing a new function. Exp Eye Res. 2007;84(3):473–85. doi: 10.1016/j.exer.2006.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhang H, Constantine R, Vorobiev S, Chen Y, Seetharaman J, Huang YJ, Xiao R, Montelione GT, Gerstner CD, Davis MW, Inana G, Whitby FG, Jorgensen EM, Hill CP, Tong L, Baehr W. UNC119 is required for G protein trafficking in sensory neurons. Nat Neurosci. 2011;14(7):874–80. doi: 10.1038/nn.2835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhang H, Fan J, Li S, Karan S, Rohrer B, Palczewski K, Frederick JM, Crouch RK, Baehr W. Trafficking of membrane-associated proteins to cone photoreceptor outer segments requires the chromophore 11-cis-retinal. J Neurosci. 2008;28(15):4008–14. doi: 10.1523/JNEUROSCI.0317-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jiang L, Zhang H, Dizhoor AM, Boye SE, Hauswirth WW, Frederick JM, Baehr W. Long-term RNA interference gene therapy in a dominant retinitis pigmentosa mouse model. Proc Natl Acad Sci U S A. 2011;108(45):18476–81. doi: 10.1073/pnas.1112758108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ismail SA, Chen YX, Rusinova A, Chandra A, Bierbaum M, Gremer L, Triola G, Waldmann H, Bastiaens PI, Wittinghofer A. Arl2-GTP and Arl3-GTP regulate a GDI-like transport system for farnesylated cargo. Nat Chem Biol. 2011;7(12):942–9. doi: 10.1038/nchembio.686. [DOI] [PubMed] [Google Scholar]
- 13.Wright KJ, Baye LM, Olivier-Mason A, Mukhopadhyay S, Sang L, Kwong M, Wang W, Pretorius PR, Sheffield VC, Sengupta P, Slusarski DC, Jackson PK. An ARL3-UNC119-RP2 GTPase cycle targets myristoylated NPHP3 to the primary cilium. Genes Dev. 2011;25(22):2347–60. doi: 10.1101/gad.173443.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chen CK, Burns ME, Spencer M, Niemi GA, Chen J, Hurley JB, Baylor DA, Simon MI. Abnormal photoresponses and light-induced apoptosis in rods lacking rhodopsin kinase. Proc Natl Acad Sci U S A. 1999;96(7):3718–22. doi: 10.1073/pnas.96.7.3718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Linari M, Hanzal-Bayer M, Becker J. The delta subunit of rod specific cyclic GMP phosphodiesterase, PDE delta, interacts with the Arf-like protein Arl3 in a GTP specific manner. FEBS Lett. 1999;458(1):55–9. doi: 10.1016/s0014-5793(99)01117-5. [DOI] [PubMed] [Google Scholar]
- 16.Fontainhas AM, Townes-Anderson E. RhoA and its role in synaptic structural plasticity of isolated salamander photoreceptors. Invest Ophthalmol Vis Sci. 2008;49(9):4177–87. doi: 10.1167/iovs.07-1580. [DOI] [PubMed] [Google Scholar]