

Abstract
Atopic allergy is a genetically determined immunodisorder that affects almost 20% of the population worldwide. Immediate symptoms of type I allergy are caused by the release of biologic mediators from effector cells induced by IgE-allergen complexes that cross-link the high-affinity receptor for IgE (FcεRI). Chronic disease manifestations result from allergen-specific T-cell activation, a process that is enhanced when allergens are presented via FcεRI-bound IgE. We report the baculovirus expression, as soluble recombinant proteins, of the minimal units required for human IgE and FcεRI interaction: Cε3 represents the third constant domain of the IgE heavy chain, and α2 is the membrane-proximal Ig-like module from FcεRIα. Native overlay experiments showed binding of human FcεRIα to recombinant Cε3 and of natural or recombinant human IgE to recombinant α2. Moreover, recombinant Cε3 inhibited binding of natural IgE antibodies to α2, and preincubation of human IgE with α2 inhibited anti-IgE–triggered histamine release from human basophils. Isolated Cε3 and α2 can now be used for the molecular and structural analysis of the IgE-FcεRI interaction, as well as for diagnostic and therapeutic applications.
Introduction
Atopic allergy, a hypersensitivity disease determined by genetic and environmental factors, affects almost 20% of the population worldwide (1). Allergic patients mount IgE antibodies against per se innocuous antigens (allergens), derived mostly from airborne particles, i.e., pollen, mite feces, animal hair/dander, moulds. Immediate-type symptoms of type I allergy (allergic rhinitis, conjunctivitis, dermatitis, asthma, and anaphylactic shock) result from the release of biologic mediators (e.g., histamine, leukotrienes) induced by cross-linking of high-affinity Fcε receptors (FcεRI) present on allergic effector cells (e.g., mast cells, basophils), after formation of IgE-allergen complexes (2). There is also evidence for the presence of FcεRI on cells of the late allergic response, such as eosinophils (3). The recent demonstration that allergen-specific T-cell activation is greatly enhanced when allergens are presented via FcεRI-bound IgE by professional antigen-presenting cells (e.g., monocytes, dendritic cells) points to the critical involvement of the IgE-FcεRI interaction in the elicitation and maintenance of the chronic manifestations of atopy, such as atopic dermatitis and chronic asthma (4, 5).
Because of their central role in atopic allergy, great emphasis has been placed on the characterization of human IgE antibodies and FcεRI (6). IgE, the least abundant immunoglobulin class, contains 4 heavy-chain constant immunoglobulin domains (Cε1–Cε4). The divergence in studies regarding the FcεRI-binding site on human IgE may be due to the lack of native conformation of Escherichia coli–expressed fragments of IgE. Evidence was provided that the FcεRI-binding site may reside at the junction of the Cε2-Cε3 domains (7). On the other hand, a construct containing Cε3-Cε4 (8) seemed to be involved in the binding of FcεRI. These investigations were probably hampered by the lack of native-like structure of the E. coli–derived proteins. Conversely, soluble functional fragments of IgE Fc and mutant chimeric IgE have been obtained in mammalian cells’ expression systems (9, 10).
Human FcεRI occurs in 2 major variants. On allergic effector cells (e.g., mast cells, basophils), FcεRI is known to consist of a single α chain, 1 β chain, and 2 γ chains. On antigen-presenting cells (e.g., monocytes, dendritic cells) the β chain is absent (5). The extracellular portion of FcεRIα consists of 2 Ig-like modules that bind IgE with high affinity, even in the absence of the β and γ subunits. Chimeric receptor studies indicate that α2, the human membrane-proximal Ig-like module of FcεRIα, confers a major contribution to the binding of IgE (11). Nevertheless, no functional isolated α2 module has been reported.
The aim of the present study was to express soluble recombinant protein modules that contain minimal units required for the interaction of human IgE and human FcεRI. Baculovirus expression of 3 IgE Cε3 constructs, 1 FcεRIα1-2, and 1 FcεRIα2 construct is reported. Recombinant protein modules were tested for their solubility and capacity to interact with complete native human IgE and a complete baculovirus–expressed α subunit of FcεRI in overlays performed under native conditions. We further investigated the ability of the recombinant protein modules to inhibit the interaction of human IgE and FcεRI by overlay experiments. Using cultured human basophils, recombinant α-chain constructs were tested for their ability to inhibit IgE binding to living effector cells and thus prevent anti-IgE–induced basophil histamine release. Results are discussed in view of a future application of the recombinant protein modules for the structural analysis of the IgE-FcεRI interaction, as well as for diagnostic and therapeutic purposes.
Methods
Materials.
The Sf9 cell line was purchased from the German Collection of Microorganisms and Cell Cultures (Braunschweig, Germany). Sera were collected from allergic patients who were diagnosed on the basis of a case history indicative of type I allergy, skin prick testing, and the demonstration of allergen-specific serum IgE antibodies by radioallergosorbent test (RAST; Pharmacia & Upjohn Diagnostics, Uppsala, Sweden) (12). The human IgE–secreting myeloma cell line U266 was obtained from American Type Culture Collection (Rockville, Maryland, USA) (13). Recombinant baculovirus–expressed human α chain of the high-affinity Fcε receptor was a kind gift from F. Kricek (Novartis Research Institute, Vienna, Austria). Recombinant chimeric IgE mAb (Bip 1-IgE) with specificity for the major birch pollen allergen Bet v 1 (14) was generated by fusion of the variable regions of the mouse monoclonal IgG1 antibody Bip 1 (15) with the constant region of human IgE (16). Antibody 12 represents a mouse monoclonal IgG1 anti-human IgE antibody that is able to bind to α chain– and basophil-bound human IgE antibodies (16). Mouse monoclonal anti-human IgA was purchased from the Central Laboratory of the Netherlands Red Cross Blood Transfusion Service (Amsterdam, the Netherlands). 125I-labeled anti-human IgE RAST antibodies were purchased from Pharmacia & Upjohn Diagnostics.
Multiple alignments and molecular graphics.
α2 sequence comparison with sequences deposited in the database was made using the European Molecular Biology Laboratory (EMBL) Advanced BLAST2 Search Server. Comparison of sequences was made using the FastA program (17) of the GCG package (18). Multiple sequence alignments were produced with CLUSTALX (19). The alignments were visually inspected and, when necessary, manually corrected. The Genetic Data Environment sequence editor (S. Smith, Harvard University, Cambridge, Massachusetts, USA) and COLORMASK (J. Thompson, EMBL, Heidelberg, Germany) were used to color conserved residues with related properties. Protein secondary structure and solvent accessibility predictions were made using the PHD program on the EMBL PredictProtein Server (20).
Cε3, α1-2, and α2 cDNA amplification and plasmid construction.
A 311-bp cDNA coding for α2 was PCR amplified from human genomic DNA using the primers containing Eco47III (5′-ggaattcatgAGCGCTGACTGGCTGCTCCTT) and EcoRI (5′-cgGAATTCTTATTAATGGTGATGATGGTGATGTGCAGCTTTTATTACAGTAAT). The α2-encoding PCR fragment was Eco47III-EcoRI digested and cloned into the Eco105I-EcoRI sites of p372ΔNotHGH-mellead (L. Mauch and G. Stahnke, unpublished data), a pBluescriptSK(+) (Stratagene, La Jolla, California, USA)–derived plasmid containing the melittin leader sequence for endoplasmic reticulum (ER) localization (21). BglII-EcoRI digestion of the resulting construct liberated a 358-bp NH2-terminal melittin leader α2 version, which was inserted into the BglII-EcoRI sites of pVL1392 (Invitrogen BV, Leek, the Netherlands). The cDNA coding for α1-2 was PCR amplified from pSVL-FcεRIα (22) using the primers containing BglII (5′-cggcgagctcAGATCTAACCCTATAAATATGGCTCCTGCCATG) and EcoRI (5′-cgGAATTCATCAATGATATGATGATGATGAGCTTTTATTACAGT), respectively, and was subcloned into the BglII-EcoRI sites of pVL1392. cDNAs coding for Cε3I and Cε3II were PCR amplified from pεExpress, a vector containing the complete human Cε-chain gene (H. Grönlund, unpublished observations). They were cloned into the SacII-SpeI sites of pBluescriptIIKS(+) (Stratagene) after PCR amplification with the sense primers containing SacII (5′-tccCCGCGGCCATGGCAGATTCCAACCCGAGA) and SacII (5′-tccCCGCGGCCATGGCAAGAGGGGTGAGCGCC), respectively, and the antisense primer containing SpeI (5′-ggACTAGTACGCGTTCAGCTGGTCTTGGTCGTGGA). NcoI digestion, followed by mung bean nuclease treatment and digestion with EcoRI, enabled insertion of Cε3I and Cε3II into the Eco105I-EcoRI sites of p372ΔNotHGH-mellead. Subsequent BglII-EcoRI digestion and ligation into pVL1392 enabled cloning of NH2-terminal melittin leader Cε3I and Cε3II. A cDNA coding for Cε3III was PCR amplified from pεExpress, using the primers containing NcoI (5′-atgCCATGGCCAACCCGAGAGGGGTGAGC) and EcoRI (5′-cgGAATTCATTAACGCGGGCCACTAGTCTT). NcoI digestion, mung bean nuclease treatment, and subsequent EcoRI digestion enabled cloning into the Eco105I-EcoRI sites of p372ΔNotHGH-mellead. BglII-EcoRI digestion enabled cloning into the corresponding sites of pVL1392 for the NH2-terminal melittin leader version of Cε3III. All constructs were confirmed by DNA sequencing, and molecular biologic manipulations followed established protocols (23).
Recombinant baculovirus selection.
Sf9 cells (24) were cotransfected with a mixture of the pVL1392 constructs and pAk Bac baculovirus DNA (Promega Corp., Madison, Wisconsin, USA). The clones with highest level of protein secretion were chosen by Western blotting for virus amplification. For each construct, 1.6 × 109 Sf9 cells were infected by recombinant baculovirus at an moi of 1 and cultured in a 3-L spinner/flask in Insect-Xpress medium (BioWhittaker Inc., Walkersville, Maryland, USA) without FCS.
Western blot analysis of FcεRI α1-2, α2, and Cε3 expression.
Supernatants from nonrecombinant Sf9 cells, as well as from Sf9 cells expressing α1-2, α2, and Cε3 constructs, were subjected to ultracentrifugation (Beckman Ti45; 40,000 rpm, 40 minutes, 4°C). Equal amounts of each supernatant were separated by 15% SDS-PAGE (25) and transferred to Immobilon-P (Millipore Corp., Bedford, Massachusetts, USA) (26). Membranes were incubated twice for 5 minutes and once for 30 minutes in buffer A (50 mM sodium phosphate [pH 7.5] 0.5% vol/vol Tween-20, 0.5% wt/vol BSA, 0.05% wt/vol sodium azide). Membranes containing α1-2 and α2 were probed with an anti–COOH-terminal hexahistidine mouse mAb (Invitrogen BV). Bound mouse mAb’s were detected with an alkaline phosphatase–conjugated rabbit anti-mouse antiserum (Jackson ImmunoResearch Laboratories Inc., West Grove, Pennsylvania, USA). Blotted recombinant Cε3 was detected with 125I-labeled anti-human IgE antibodies (RAST).
125I labeling of purified proteins, blot overlays, and overlay inhibition experiments.
Purified proteins were labeled with 125I using the chloramine-T method (23). Binding of natural human IgE to recombinant α1-2 and α2 was demonstrated as follows. One hundred–microliter aliquots of recombinant α1-2– and α2-containing ultracentrifuged supernatants were transferred onto nitrocellulose (Schleicher & Schuell GmbH, Dassel, Germany) with a soak-blot chamber (Hoefer Scientific Instruments, San Francisco, California, USA). Nitrocellulose blots were washed with buffer A and were incubated with serum or with the chimeric IgE antibody supernatant (diluted 1:2 in buffer A), or with U266 supernatant, overnight at 4°C. Bound IgE antibodies were detected with 125I-labeled anti-human IgE antibodies (1:5 diluted in buffer A; RAST, 200,000 cpm/mL) or with 125I-labeled anti-human IgE antibody 12. For control purposes, 125I-labeled α chain (200,000 cpm/mL) was used. The ability of nitrocellulose-bound Cε3 to interact with complete recombinant α chain was shown as follows. One hundred–microliter aliquots of recombinant Cε3–containing supernatant were dotted onto nitrocellulose. Nitrocellulose blots were washed with buffer A, incubated with 125I-labeled α chain or, for control purposes, with 125I-labeled rBet v 1, overnight at room temperature (200,000 cpm/mL; diluted in buffer A), and viewed by autoradiography. The ability of recombinant Cε3 constructs to inhibit binding of complete natural human IgE to nitrocellulose-bound recombinant α1-2 or α2 was demonstrated as follows. Twenty-microliter aliquots of α1-2– or α2-containing supernatant were dotted onto nitrocellulose. Nitrocellulose membranes were blocked with buffer A and preincubated with Cε3I-containing supernatants or, for control purposes, with supernatant from nonrecombinant Sf9 cells (both diluted 1:1 in buffer A), overnight at 4°C. After washing in buffer A, both membranes were incubated with IgE-containing serum from an atopic patient (diluted 1:1 in buffer A) overnight at 4°C. Bound human IgE antibodies were detected with 125I-labeled anti-human IgE antibodies (RAST).
Isolation and culture of basophils, and histamine release experiments.
Peripheral blood was obtained from a patient with untreated chronic-phase chronic myeloid leukemia (CML) after informed consent was obtained. Mononuclear cells (MNCs) were isolated using Ficoll (Pharmacia & Upjohn Diagnostics). After washing in PBS, MNCs were cultured in RPMI-1640 (Proteine Antigene Antikorper, Linz, Austria) medium supplemented with 10% FCS (GIBCO BRL, Gaithersburg, Maryland, USA) and 100U/mL of recombinant human (rh) IL-3 (PeproTech Inc., Rocky Hill, New York) in 5% CO2 at 37°C for 2 weeks. After 2 weeks, the percentage of basophils amounted to 27%, as assessed by Giemsa staining. The ability of recombinant α-chain constructs to prevent IgE binding to human basophils was investigated by preincubating human myeloma IgE (Chemicon International, Temecula, California, USA) with either 0.9% NaCl (control), FcεRIα diluted in 0.9% NaCl to a concentration of 10 μg/mL, or α1-2– or α2-containing supernatant diluted 1:2 in 0.9% NaCl for 1 hour at 4°C. Equal numbers of basophils were then exposed to the IgE preparations for 4 hours at 4°C. Thereafter, cells were washed in PBS, and histamine release was induced with various concentrations (0.1–10 μg/mL) of mouse monoclonal anti-human IgE antibody E-124-2-8 (Immunotech, Marseilles, France) for 30 minutes at 37°C (27). Cells were then centrifuged at 4°C, and the cell-free supernatants and cell lysates were recovered. Liberated histamine (supernatants) was measured by radioimmunoassay (Immunotech) and is expressed as a percentage of total histamine release (27). All determinations were carried out in triplicate.
Results
Selection of module boundaries by multiple sequence alignment and structure prediction.
Cε3 and the 2 α modules are expected to adopt a typical immunoglobulin fold formed by β sandwich (28). Within the IgSF, the proteins have been reported to belong to the C1 and C2 set, respectively (29). To select the domain boundaries that should lead to correctly folded proteins, a BLAST search was performed, and the highest hits were used to prepare 2 separated multiple alignments (Figure 1, a and b). The presence of a conserved pattern of hydrophobics in the multiple alignments suggested the position of the β strands and, therefore, position of the regions necessary to retain the tertiary fold. The sequence of Cε3 was further submitted to the BLAST program available under the structural classification of proteins (SCOP) web page that identifies sequences for which three-dimensional structures exist (30). The highest hit found in this way is the 1fc1 structure, a constant domain from a human IgG (31). The region from 238 to 343 of 1fc1, which corresponds to the whole constant domain, aligns with the sequence of Cε3 with a 49.5% similarity and a 32.3% identity. The coordinates of 1fc1 were therefore used to model Cε3 that is shown in Figure 1c, also given as a reference for the immunoglobulin fold. Three constructs were chosen for Cε3 with slightly different lengths. This was dictated by the observation that when working with isolated domains, addition or deletion of even a few NH2-terminal and/or COOH-terminal residues may have strong influence on the domain stability (32). The NH2-terminus of Cε3 was extended by 4 (Cε3II), 6 (Cε3III), and 8 (Cε3I) residues, respectively, from the first residue of Cε3 that aligns with 1fc1 (ser337 in Cε3 with ser239 in 1fc1). In this way, the NH2-terminus may pack against the 2 loops in close spatial proximity (Figure 1c). Two choices were made for the COOH-terminus differing by 3 residues. α1-2 boundaries were selected according to previous reports (33, 34). For α2, only 1 construct was prepared, in agreement with the model reported (35).
Figure 1.

(a) FcεRIα1-2 and α2 multiple sequence alignment, secondary structure, and solvent accessibility prediction. The top 3 lines represent α2, α1-2, and α-chain constructs, respectively. The consensus line derives from a Gonnet pam250 matrix in which asterisks represent fully conserved residues, and columns and dots represent, respectively, strong and weak groups of conserved residues (defined in ref. 45); after the double virgule (//), FCE2_RAT is not considered for the consensus. Each sequence is preceded by its SwissProt database’s identification entry name. Hyphens in the sequences indicate gaps. Sequences are colored to illustrate conservation of features in addition to amino acid identity. All glycines and prolines are colored in orange and yellow, respectively. Other residues are colored according to conservation of their physicochemical properties (purple, acidic; blue, hydrophobic; light blue, hydrophobic tendency; red, basic; green, hydrophilic; white, unconserved). The last 3 lines indicate the predicted secondary structure (SecStrPred), the predicted solvent accessibility (accessibility), both according to the PHD program and a numbering reference (ruler). In the secondary structure prediction, uppercase letters are used for positions where accuracy exceeds 86%, and the overall accuracy of the prediction is 72%. E indicates residue in predicted β strand. In the solvent accessibility prediction, uppercase letters are used for residues where accuracy exceeds 69%, and the overall accuracy of the prediction is 54%. A and B indicate exposed and buried residues, respectively. (b) Cε3 multiple sequence alignment, secondary structure, and solvent accessibility prediction. The top 3 lines represent Cε3I, Cε3II, and Cε3III constructs, respectively. Each sequence is preceded by its SwissProt or SPTREMBLNEW database’s identification entry name. SecCgamma2 line represents the secondary structure content of the Cγ2 module of IgG1, for which the three-dimensional structure is known (31). E indicates residue in β strand. Consensus, SecStrPred, accessibility and ruler lines, hyphens, and color code are as in a. (c) Ribbon representation of the immunoglobulin fold as modeled for Cε3 on the coordinates of 1fc1. The fold belongs to the C1 immunoglobulin subfamily. The 2 sheets are colored differently. The NH2- and COOH-termini (N and C, respectively) are indicated near the corresponding strand.
Recombinant α1-2, α2, and Cε3 are secreted as soluble proteins of correct molecular weight.
A monoclonal anti-hexahistidine antibody detected recombinant α1-2 as a double band with a molecular weight of about 35 kDa, likely corresponding to different glycosylation states of the protein (Figure 2a, lane 2). Similar results were observed previously for a slightly different Sf9-expressed FcεRIα1-2 construct (34). Recombinant α2 was detected in the Sf9 supernatant as single band with a molecular weight of about 16 kDa by the anti-hexahistidine antibody (Figure 2a, lane 3), whereas no reaction was observed with nitrocellulose-blotted nonrecombinant Sf9 cell supernatant (Figure 2a, lane 1). When equal amounts of α1-2– and α2-containing supernatants were dot-blotted and probed with the anti-hexahistidine antibody, we found that α2 was secreted at considerably lower levels than α1-2 (data not shown).
Figure 2.

(a) Detection of nitrocellulose-blotted recombinant α1-2 and α2. Supernatants from Sf9 cells infected with nonrecombinant baculovirus (lane 1) and from Sf9 cells infected with α1-2 (lane 2) and α2 (lane 3) recombinant baculoviruses were separated by SDS-PAGE and blotted onto nitrocellulose. Recombinant α1-2 (35 kDa) and α2 (16 kDa) are recognized by an anti-hexahistidine mAb in lanes 2 and 3, respectively. (b) Detection of nitrocellulose-blotted recombinant Cε3 with anti-human IgE antibodies. Nitrocellulose-blotted supernatants from Sf9 insect cells infected with nonrecombinant (lane 1) and Cε3III-expressing (lane 2) baculoviruses were exposed to 125I-labeled anti-human IgE antibodies.
As exemplified for recombinant Cε3III, all Cε3 constructs (Cε3I–III) were detected as soluble proteins in nitrocellulose-blotted Sf9 cell supernatants by anti-human IgE antibodies (Figure 2b, lane 2). Anti-human IgE antibodies failed to react with supernatant from nonrecombinant Sf9 cells (Figure 2b, lane 1).
Nitrocellulose-bound recombinant α1-2 and α2 bind natural human IgE of various sources.
Recombinant α1-2 (Figure 3a) or α2 (Figure 3b) were immobilized to nitrocellulose under nondenaturing conditions and exposed to serum IgE from an atopic patient (Figure 3, lanes 1, 4, and 5), a chimeric IgE antibody (Bip 1) consisting of the human IgE constant region and a mouse variable region (Figure 3, lane 2), or human myeloma cell–derived IgE (Figure 3, lane 3). Complete human IgE antibodies bound to the recombinant α1-2 and α2 domains were then viewed with 125I-labeled anti-human IgE antibodies, which can recognize FceRI-bound IgE (RAST anti-IgE: Figure 3, lanes 1–3; monoclonal anti-human IgE 12: Figure 3, lane 4). Results obtained showed that recombinant α1-2 and, to a lesser extent, recombinant α2, bound natural human IgE antibodies from all 3 sources. When 125I-labeled complete recombinant α chain was incubated with α1-2– or α2-bound natural human IgE, no reactivity was observed (Figure 3, lane 5).
Figure 3.

IgE-binding capacity of nitrocellulose-dotted, baculovirus-expressed α-chain constructs. Sf9 cell supernatants containing recombinant α1-2 (a) or α2 (b) construct were dotted on nitrocellulose. Filter-bound proteins were incubated with serum from an atopic patient containing high IgE levels (lanes 1, 4, and 5), with chimeric monoclonal IgE (2), and with a human IgE–containing myeloma supernatant (3), and were detected with 125I-labeled anti-human IgE RAST antibodies (1-3), a 125I-labeled mouse monoclonal anti-human IgE antibody 12 (4), and, for control purposes, a 125I-labeled human α chain (5).
Recombinant Cε3 constructs bind complete recombinant α chain and inhibit binding of natural human IgE to recombinant α1-2 and α2. As exemplified for recombinant Cε3II (Figure 4), all Cε3 constructs bound 125I-labeled complete recombinant a chain (Figure 4, panel 1) but not a 125I-labeled control protein (recombinant Bet v 1; Figure 4, panel 2). Nitrocellulose-bound recombinant α1-2 (Figure 5, lane a) or α2 (Figure 5, lane b) was preincubated with Sf9 supernatants without (Figure 5, panel 1) or with (Figure 5, panel 2) recombinant Cε3I and then exposed to equal amounts of natural human serum IgE. Preincubation with recombinant Cε3I resulted in a great reduction of human IgE binding to the α1-2 construct and in a complete abolishment of IgE reactivity to recombinant α2 (Figure 5). Similar results were obtained after preincubation with recombinant Cε3II and Cε3III (data not shown).
Figure 4.
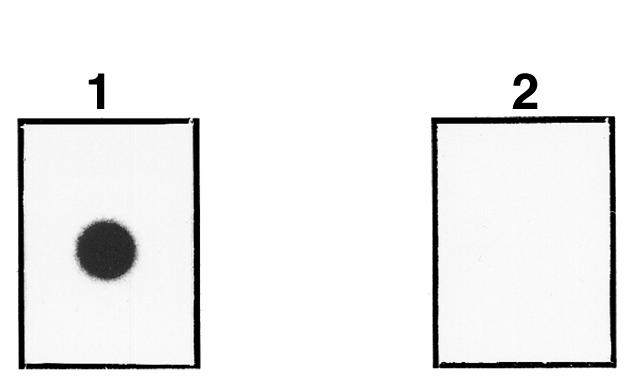
Nitrocellulose-dotted recombinant Cε3 binds recombinant FcεRIα. Nitrocellulose-dotted Sf9 cell supernatant containing recombinant Cε3II was incubated with 125I-labeled α chain (panel 1) or 125I-labeled rBet v 1 (panel 2).
Figure 5.

Recombinant Cε3 inhibits IgE binding to recombinant α-chain constructs. Sf9 cell supernatants containing recombinant α1-2 (lane a) and recombinant α2 (lane b) were dotted onto nitrocellulose and preincubated either with Sf9 cell supernatants without (panel 1) or with (panel 2) recombinant Cε3. After washing, membranes were incubated with equal amounts of human serum IgE, and bound complete natural IgE antibodies were detected with 125I-labeled anti-human IgE antibodies.
Recombinant α1-2 and α2 inhibit IgE binding to human basophils and anti-IgE–induced histamine release.
To analyze the biologic activity of recombinant α1-2 and α2, we investigated whether the recombinant α-chain constructs can sequester natural human IgE and, by this, prevent histamine release from cultured human CML basophils. Figure 6 shows that preincubation of IgE with complete a chain and the α1-2 construct completely inhibited IgE binding to the cells, as no histamine release could be subsequently induced with anti-IgE antibodies (0.1–10 μg/mL). Preabsorption of IgE with recombinant α2 leads to an approximately 50% reduction of histamine release, compared with the histamine release induced by saline-treated IgE, at all 3 concentrations of anti-human IgE (Figure 6).
Figure 6.

Recombinant α1-2 and α2 prevent IgE binding to human basophils and anti-IgE–induced histamine release. Purified human IgE was preincubated with 0.9% NaCl (control), complete recombinant FcεRIα (FcεRI α chain), recombinant α1-2 (α1-2), or α2 (α2), and then exposed to human basophils. After washing, cells were incubated with various concentrations of anti-human IgE antibodies (x axis). Histamine released in the cell-free supernatant is expressed as a percentage of total histamine release (y axis). All values are expressed as the mean ± SD of triplicate determinations.
Discussion
The interaction of IgE antibodies with FcεRI on effector cells (e.g., mast cells, basophils, eosinophils) (2, 3) and inducer cells (monocytes, dendritic cells) (4, 5) of the allergic reaction represents the key event responsible for the acute and chronic manifestations of type I allergy. Despite the central role of the IgE-FcεRI interaction in the pathogenesis of atopy, the precise mode of the IgE-FcεRI interaction is not fully understood, and the minimal domains required for this interaction have not yet been produced as active recombinant proteins. As we failed to produce soluble α1-2, α2, and Cε3 in E. coli, we expressed these proteins in insect cells. All recombinant proteins (α1-2, α2, Cε3I-III) were secreted by baculovirus-infected insect cells as soluble proteins of correct molecular weight in the cell culture supernatants. Evidence for the correct folding and functional activity of recombinant Cε3 constructs comes from our demonstration that in nondenaturing overlay experiments, recombinant Cε3 bound the complete recombinant α chain and also inhibited binding of natural IgE antibodies to the α1-2 and α2 constructs. The incomplete inhibition of the binding of complete natural IgE to the α1-2 construct may result from a lower affinity of the insect cell–derived Cε3 domain or the lacking accessory activity of Cε2 and/or Cε4. The strong inhibition of IgE binding to α1-2 by our recombinant insect cell–derived Cε3 suggests, however, that it can compete with complete IgE for its high-affinity receptor and thus may be considered as a potential tool for therapy of type I allergy. In this context, it will be interesting to see whether administration of Cε3 can downregulate IgE production or may be useful to prevent potentially increased production of IgE in response to novel therapy forms of atopy that aim at a depletion of IgE antibodies (36).
Regarding the recombinant α-chain constructs α1-2 and α2, we found that both proteins were able to bind human IgE antibodies. The biologic activity of recombinant α1-2 and α2 was demonstrated by their ability to prevent IgE binding to human basophils and subsequent anti-IgE–induced histamine release. The weaker IgE-binding capacity of recombinant α2 in the overlay and basophil experiments could be due to lower affinity or the lower amounts of secreted α2 compared with α1-2. Although we have no evidence that the isolated α1 domain can bind IgE, and although the structural analysis of the complete α chain (37) points against a direct interaction of α1 with IgE, it is possible that α1 may be involved in the interaction with IgE.
Nevertheless, our data provide clear evidence that isolated recombinant α2 alone can bind IgE antibodies, and we thus identify it as the minimal structural requirement for IgE binding. The latter result is in contrast to a previous study reporting that human recombinant α2 expressed in the membranes of COS-7 cells failed to bind IgE antibodies (11). Several explanations for this discrepancy are conceivable. First, it is possible that the COS cell–expressed α2, although detected by an anti-FcεRIα peptide (amino acids 160–197) antiserum, was partly buried in the cell membrane and thus not accessible for IgE. The second possibility, that the COS cell–expressed α2 was not properly folded, is supported by the observation that this α2 was not recognized by anti-FcεRIα mAb’s that inhibited IgE binding to the receptor and thus presumptively recognized folded α2 (11). Finally, it is possible that a differential folding ability of soluble or membrane-inserted α2 could be due to alternative processing of the protein in the ER-Golgi districts.
That α2 bound IgE antibodies and inhibited IgE binding to human basophils suggests, however, that α2 can be used for diagnostic as well as therapeutic purposes. Regarding in vitro allergy diagnosis, labeled α2 might be used to discriminate IgE fractions that may contain bound soluble forms of FcεRI (38) and thus lack capacity to sensitize effector and inducer cells of atopy. As with the recently described anti-human IgE antibodies (36), we may consider therapeutic administration of α2 to remove IgE antibodies from the circulation and to prevent IgE binding to effector and inducer cells of the allergic reaction. In fact, it was shown in murine models, as well as in humans, that elevated levels of IgE enhanced FcεRI expression on mast cells and basophils (39–41) and that treatment with anti-human IgE antibodies suppressed the early- and late-phase responses to inhaled allergens in allergic asthmatic subjects (42). It has also been demonstrated that complete recombinant α chain inhibited IgE production in vitro and prevented anaphylactic shock in a murine model (43, 44). If the small α2 domain is used for therapeutic removal of circulating IgE instead of anti-human IgE antibodies or complete α chain, the risk in inducing antiantibodies or anti-FcεRI autoantibodies might be reduced.
In conclusion, we expressed minimal domains of the IgE-FcεRI interaction, α2 and Cε3, as soluble and functional proteins in insect cells. Larger amounts of purified recombinant α2 and Cε3 can now be produced to investigate the mode of IgE binding to FcεRI by structural methods (nuclear magnetic resonance, x-ray crystallography) as well as by kinetic measurements. Both recombinant protein modules may also be used to study their influence on the regulation of IgE and FcεRI expression, using in vitro and in vivo systems that provide information as to whether the administration of Cε3 or α2 may represent strategies for the therapy of atopic diseases.
Acknowledgments
We thank Gisela Stahnke for kindly providing the plasmid p372ΔNotHGH-mellead and for invaluable help on cloning strategy; Henry Metzger for the kind gift of plasmid pSVL-FcεRIα; Heinz Haubruck and Jacky Schmitt for useful discussions and help in the baculovirus expression strategy; Alberto Bianco for helpful discussions; Annarita Berlese for human genomic DNA purification and suggestions; and Dietrich Kraft for his support. We thank F. Kricek for purified baculovirus–expressed FcεRIα. This project was supported in part by grants Y078GEN and F0506 of the Austrian Science Fund, by the Interdisziplinäres Cooperations Projekt program of the Austrian Ministry of Research and Transports, and by a grant from Pharmacia & Upjohn Diagnostics. L. Vangelista was supported by an EMBL Predoc Fellowship.
Footnotes
Luca Vangelista’s present address is: Molecular Immunology Group, International Center for Genetic Engineering and Biotechnology, 34012 Trieste, Italy.
References
- 1.Romagnani S. Atopic allergy and other hypersensitivities: interactions between genetic susceptibility, innocuous and/or microbial antigens and the immune system. Curr Opin Immunol. 1997;9:773–775. doi: 10.1016/s0952-7915(97)80176-8. [DOI] [PubMed] [Google Scholar]
- 2.Beaven MA, Metzger H. Signal transduction by Fc receptors: the Fc epsilon RI case. Immunol Today. 1993;14:222–226. doi: 10.1016/0167-5699(93)90167-j. [DOI] [PubMed] [Google Scholar]
- 3.Gounni AS, et al. High-affinity IgE receptor on eosinophils is involved in defence against parasites. Nature. 1994;367:183–186. doi: 10.1038/367183a0. [DOI] [PubMed] [Google Scholar]
- 4.Bieber T. FcεRI on antigen-presenting cells. Curr Opin Immunol. 1996;8:773–777. doi: 10.1016/s0952-7915(96)80003-3. [DOI] [PubMed] [Google Scholar]
- 5.Stingl G, Maurer D. IgE-mediated allergen presentation via Fc epsilon RI on antigen-presenting cells. Int Arch Allergy Immunol. 1997;113:24–29. doi: 10.1159/000237499. [DOI] [PubMed] [Google Scholar]
- 6.Sutton BJ, Gould HJ. The human IgE network. Nature. 1993;366:421–428. doi: 10.1038/366421a0. [DOI] [PubMed] [Google Scholar]
- 7.Helm B, et al. The mast cell binding site on human immunoglobulin E. Nature. 1988;331:180–183. doi: 10.1038/331180a0. [DOI] [PubMed] [Google Scholar]
- 8.Helm B, et al. Identification of the high affinity receptor binding region in human immunoglobulin E. J Biol Chem. 1996;271:7494–7500. doi: 10.1074/jbc.271.13.7494. [DOI] [PubMed] [Google Scholar]
- 9.Basu M, et al. Purification and characterization of human recombinant IgE-Fc fragments that bind to the human high affinity IgE receptor. J Biol Chem. 1993;268:13118–13127. [PubMed] [Google Scholar]
- 10.Presta L, et al. The binding site on human immunoglobulin E for its high affinity receptor. J Biol Chem. 1994;269:26368–26373. [PubMed] [Google Scholar]
- 11.Mallamaci MA, et al. Identification of sites on the human FcεRIα subunit which are involved in binding human and rat IgE. J Biol Chem. 1993;268:22076–22083. [PubMed] [Google Scholar]
- 12.Valenta R, Vrtala S, Ebner C, Kraft D, Scheiner O. Diagnosis of grass pollen allergy with recombinant timothy grass (Phleum pratense) pollen allergens. Int Arch Allergy Immunol. 1992;97:287–294. doi: 10.1159/000236135. [DOI] [PubMed] [Google Scholar]
- 13.Nilsson K, Bennich H, Johansson SG, Ponten J. Established immunoglobulin producing myeloma (IgE) and lymphoblastoid (IgG) cell lines from an IgE myeloma patient. Clin Exp Immunol. 1970;7:477–489. [PMC free article] [PubMed] [Google Scholar]
- 14.Breiteneder H, et al. The gene coding for the major birch pollen allergen Bet v I, is highly homologous to a pea disease resistance response gene. EMBO J. 1989;8:1935–1938. doi: 10.1002/j.1460-2075.1989.tb03597.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Laffer S, et al. Molecular characterization of Bip 1, a monoclonal antibody that modulates IgE binding to birch pollen allergen, Bet v 1. J Immunol. 1996;157:4953–4962. [PubMed] [Google Scholar]
- 16.Laffer S, et al. An in vitro model for the allergen-IgE-FcεRI interaction. Int Arch Allergy Immunol. 1999;118:116–118. doi: 10.1159/000024044. [DOI] [PubMed] [Google Scholar]
- 17.Pearson WR, Lipman DJ. Improved tools for biological sequence comparison. Proc Natl Acad Sci USA. 1988;85:2444–2448. doi: 10.1073/pnas.85.8.2444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Devereux J, Haeberli P, Smithies O. A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res. 1984;12:387–395. doi: 10.1093/nar/12.1part1.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;25:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rost B, Sander C. Combining evolutionary information and neural networks to predict protein secondary structure. Proteins. 1994;20:216–226. doi: 10.1002/prot.340190108. [DOI] [PubMed] [Google Scholar]
- 21.Tessier DC, Thomas DY, Khouri HE, Laliberte F, Vernet T. Enhanced secretion from insect cells of a foreign protein fused to the honeybee melittin signal peptide. Gene. 1991;98:177–183. doi: 10.1016/0378-1119(91)90171-7. [DOI] [PubMed] [Google Scholar]
- 22.Miller L, Blank U, Metzger H, Kinet JP. Expression of high-affinity binding of human immunoglobulin E by transfected cells. Science. 1989;244:334–337. doi: 10.1126/science.2523561. [DOI] [PubMed] [Google Scholar]
- 23.Sambrook, J., Fritsch, E.F., and Maniatis, T. 1989. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press. Cold Spring Harbor, NY.
- 24.Vaughn JL, Goodwin RH, Thompkins GJ, McCawley P. The establishment of two insect cell lines from the insect Spodoptera frugiperda (Lepidoptera: Noctuidae) In vitro. 1977;13:213–217. doi: 10.1007/BF02615077. [DOI] [PubMed] [Google Scholar]
- 25.Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 26.Towbin H, Staehelin T, Gordon T. Electrophoretic transfers of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 1979;76:4350–4354. doi: 10.1073/pnas.76.9.4350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Valent P, et al. Interleukin 3 activates human blood basophils via high-affinity binding sites. Proc Natl Acad Sci USA. 1989;86:5542–5546. doi: 10.1073/pnas.86.14.5542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bork P, Holm L, Sander C. The immunoglobulin fold. Structural classification, sequence patterns and common core. J Mol Biol. 1994;242:309–320. doi: 10.1006/jmbi.1994.1582. [DOI] [PubMed] [Google Scholar]
- 29.Harpaz Y, Chothia C. Many of the immunoglobulin superfamily domains in cell adhesion molecules and surface receptors belong to a new structural set which is close to that containing variable domains. J Mol Biol. 1994;238:528–539. doi: 10.1006/jmbi.1994.1312. [DOI] [PubMed] [Google Scholar]
- 30.Murzin AG, Brenner SE, Hubbard T, Chothia C. SCOP: a structural classification of proteins database for the investigation of sequences and structures. J Mol Biol. 1995;247:536–540. doi: 10.1006/jmbi.1995.0159. [DOI] [PubMed] [Google Scholar]
- 31.Deisenhofer J. Crystallographic refinement and atomic models of a human Fc fragment and its complex with fragment B of protein A from Staphylococcus aureus at 2.9- and 2.8-A resolution. Biochemistry. 1981;20:2361–2370. [PubMed] [Google Scholar]
- 32.Pfuhl M, Improta S, Politou AS, Pastore A. When a module is also a domain: the role of the N terminus in the stability and the dynamics of immunoglobulin domains from titin. J Mol Biol. 1997;265:242–256. doi: 10.1006/jmbi.1996.0725. [DOI] [PubMed] [Google Scholar]
- 33.Blank U, Ra C, Kinet JP. Characterization of truncated α chain products from human, rat and mouse high affinity receptor for immunoglobulin E. J Biol Chem. 1991;266:2639–2646. [PubMed] [Google Scholar]
- 34.Yagi S, et al. High-level expression of the truncated alpha chain of human high-affinity receptor for IgE as a soluble form by baculovirus-infected insect cells. Biochemical characterization of the recombinant product. Eur J Biochem. 1994;220:593–598. doi: 10.1111/j.1432-1033.1994.tb18660.x. [DOI] [PubMed] [Google Scholar]
- 35.Padlan EA, Helm BA. A modeling study of the alpha-subunit of human high-affinity receptor for immunoglobulin-E. Receptor. 1992;2:129–144. [PubMed] [Google Scholar]
- 36.Heusser C, Jardieu P. Therapeutic potential of anti-IgE antibodies. Curr Opin Immunol. 1997;9:805–813. doi: 10.1016/s0952-7915(97)80182-3. [DOI] [PubMed] [Google Scholar]
- 37.Garman SC, Kinet JP, Jardetzky TS. Crystal structure of the human high-affinity IgE receptor. Cell. 1998;95:951–961. doi: 10.1016/s0092-8674(00)81719-5. [DOI] [PubMed] [Google Scholar]
- 38.Bjerke T, et al. Soluble FcεRI α-subunit in serum from allergic and non-allergic patients with asthma: Immunopurification and correlation to disease activity. J Allergy Clin Immunol. 1996;97:279. [Google Scholar]
- 39.Yamaguchi M, et al. IgE enhances mouse mast cell Fc epsilon RI expression in vitro and in vivo: evidence for a novel amplification mechanism in IgE-dependent reactions. J Exp Med. 1997;185:663–672. doi: 10.1084/jem.185.4.663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.MacGlashan D, et al. In vitro regulation of Fc epsilon RI alpha expression on human basophils by IgE antibody. Blood. 1998;91:1633–1643. [PubMed] [Google Scholar]
- 41.Shan D, Ledbetter JA, Press OW. In vitro regulation of Fc epsilon RI alpha expression on human basophils by IgE antibody. Blood. 1998;91:1644–1652. [Google Scholar]
- 42.Fahy JV, et al. The effect of an anti-IgE monoclonal antibody on the early- and late-phase responses to allergen inhalation in asthmatic subjects. Am J Respir Crit Care Med. 1997;155:1828–1834. doi: 10.1164/ajrccm.155.6.9196082. [DOI] [PubMed] [Google Scholar]
- 43.Yanagihara Y, et al. Recombinant soluble form of the human high-affinity immunoglobulin E (IgE) receptor inhibits IgE production through its specific binding to IgE-bearing B cells. J Clin Invest. 1994;94:2162–2165. doi: 10.1172/JCI117574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Naito K, Hirama M, Okumura K, Ra C. Recombinant soluble form of the human high-affinity receptor for IgE prevents anaphylactic shock in mice. J Allergy Clin Immunol. 1996;97:773–780. doi: 10.1016/s0091-6749(96)80155-2. [DOI] [PubMed] [Google Scholar]
- 45.Benner SA, Cohen MA, Gonnet GH. Amino acid substitution during functionally constrained divergent evolution of protein sequences. Protein Eng. 1994;7:1323–1332. doi: 10.1093/protein/7.11.1323. [DOI] [PubMed] [Google Scholar]