

Abstract
Stress is associated with the onset and severity of several psychiatric disorders that occur more frequently in women than men, including posttraumatic stress disorder (PTSD) and depression. Patients with these disorders present with dysregulation of several stress response systems, including the neuroendocrine response to stress, corticolimbic responses to negatively valenced stimuli, and hyperarousal. Thus, sex differences within their underlying circuitry may explain sex biases in disease prevalence. This review describes clinical studies that identify sex differences within the activity of these circuits, as well as preclinical studies that demonstrate cellular and molecular sex differences in stress responses systems. These studies reveal sex differences from the molecular to the systems level that increase endocrine, emotional, and arousal responses to stress in females. Exploring these sex differences is critical because this research can reveal the neurobiological underpinnings of vulnerability to stress-related psychiatric disorders and guide the development of novel pharmacotherapies.
Keywords: Sex difference, Stress, Depression, Posttraumatic stress disorder, Arousal, Hypothalamic pituitary adrenal axis, Emotion, Corticotropin releasing factor, Locus coeruleus
1. Introduction
Stressor exposure initiates a complex set of neuronal, endocrine, and behavioral responses that prepare an organism to cope with this perturbation in homeostasis. Although initiation of these stress responses is typically adaptive, their persistent or inappropriate activation is linked to the pathophysiology of several medical and psychiatric disorders. Despite the fact that most people will experience a large number of stressful events during their lifetime, only a small percentage go on to develop the dysregulated stress responses that characterize these diseases. Thus, a major challenge of modern medicine is to determine what factors confer vulnerability or resilience to stress. Several of these factors already have been identified. For example, early life stress can increase the vulnerability to develop certain psychiatric disorders in adulthood [1; 2; 3]. Additionally, coping strategy is a determinant of the development of certain diseases, such that a passive coping strategy is a risk factor for major depression, while a proactive coping strategy is a risk factor for cardiovascular disease [4; 5; 6; 7; 8; 9]. Another factor associated with stress vulnerability is biological sex. Although historically sex differences in medical and psychiatric disorders were largely ignored, more recent research has focused on the neurobiological underpinnings of sex differences in vulnerability and resilience to stress and its related disorders.
The interest in sex as a moderating factor of disease vulnerability comes, in part, from epidemiological data that reveal sex differences in the prevalence of many disorders that are exacerbated by stress (Table 1). For example, men are more likely to suffer from substance-related disorders, such as alcohol and drug abuse [10; 11]. In contrast, women are roughly twice as likely to suffer from anxiety disorders, such as panic disorder, and trauma-related disorders, such as posttraumatic stress disorder (PTSD)[12; 13; 14; 15]. Women also have higher rates of major depression than men [16; 17; 18]. Medical disorders that are often comorbid with depression and anxiety, such as migraines, insomnia, and irritable bowel syndrome, are reported more frequently in women, perhaps suggesting some common underlying pathology [19; 20; 21; 22; 23; 24]. These epidemiological data detail sex differences in many stress-related disorders, but population-based studies of disease prevalence can fail to capture nuances in presentation of these diseases. For instance, although fewer women experiment with drugs, when women are exposed to addictive drugs they develop substance abuse faster than men [25; 26]. Additionally, a recent study on depression found that when additional symptoms related to the disorder, such as anger attacks/aggression, substance abuse, and risk taking, were included in the diagnosis, the sex disparities in disease prevalence were eliminated [27]. Even while bearing these caveats in mind, the sex differences in the prevalence and presentation of stress-related disorders suggest sex differences in their underlying biology.
Table 1.
Sex differences in prevalence of stress-related disorders.
| Lifetime Prevalence | Ratio | |||
|---|---|---|---|---|
| Female | Males | Female:Male | Citation | |
| Panic | 6.2% | 3.1% | 2.0 | [336] |
| Generalized Anxiety | 7.1% | 4.2% | 1.7 | [336] |
| Any Anxiety Disorder | 36.4% | 25.4% | 1.4 | [336] |
| PTSD | 9.7% | 3.6% | 2.7 | [336] |
| Major Depression | 20.2% | 13.2% | 1.5 | [336] |
| Any Affective Disorder | 24.4% | 17.5% | 1.4 | [336] |
| Alcohol Abuse | 7.5% | 19.6% | 0.4 | [336] |
| Drug Abuse | 4.8% | 11.6% | 0.4 | [336] |
| Migraine | 18.2% | 6.5% | 2.8 | [22] |
| Insomnia | 12.9% | 6.2% | 2.1 | [23] |
| Irritable Bowel Syndrome | 14.5% | 7.7% | 1.9 | [24] |
Given the diversity of psychiatric and medical disorders related to stress that have a sex bias, exploring the neurobiological mechanisms that contribute sex disparities in all of these disorders is beyond the scope of this review. Instead, we will focus on several disorders with a sex bias that share common pathophysiology as prototypical examples. These examples will illustrate how sex differences at the molecular and cellular level can contribute to sex differences in disease vulnerability and severity. The disorders that are the focus of the review include trauma-related disorders and major depression. Not only are these stress-related psychiatric disorders more prevalent in women, but they share three other features: dysregulation of the hypothalamic-pituitary-adrenal (HPA) axis, heightened reactivity to emotional stimuli with a negative valence, and hyperarousal. The goal is, when possible, to link sex differences in clinical features to sex differences in the circuitry that mediates these features (Fig. 1). Additionally, preclinical studies that associate sex specific cellular and molecular alterations with sex differences in stress hormone levels, emotions, and arousal will be highlighted.
Figure 1.
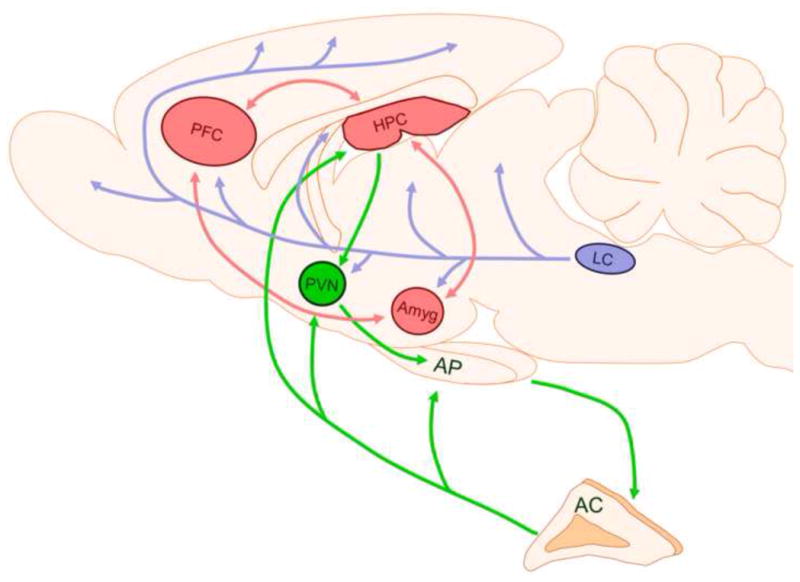
Schematic representing the circuitry primarily responsible for mediating symptoms of PTSD and depression. The HPA axis that regulates the neuroendocrine responses to stress is shown in green. The corticolimbic circuitry that mediates emotional responses to negatively valenced stimuli is shown in red. The LC-norepinephrine system that initiates arousal responses to stress is shown in blue. AC: adrenal cortex; Amyg: amygdala; AP: anterior pituitary; HPC: hippocampus; LC: locus coeruleus; PFC: prefrontal cortex; PVN: paraventricular nucleus
2. Sex differences in neuroendocrine responses to stress
2.1 The HPA axis
One of the hallmarks of the stress response is activation of the HPA axis. Activation of this neuroendocrine response to stress results in the release of glucocorticoids. These hormones prepare the organism to deal with threatening stimuli by increasing energy though glucose metabolism, lipolysis, and proteolysis, while suppressing growth, reproduction, and the immune system [28; 29; 30]. There are several brain regions and endocrine glands that work in concert to modulate HPA axis activity in response to stress. Initiation of this response following stressor exposure occurs when hypophysiotrophic neurons in the paraventricular nucleus of the hypothalamus (PVN) release corticotropin releasing factor (CRF) and vasopressin [31; 32]. These neuropeptides stimulate the pituitary gland to secrete adrenocorticotropic hormone (ACTH) into the blood stream. In turn, ACTH acts on the cortex of the adrenal glands to induce the production of glucocorticoids. In humans the primary glucocorticoid is cortisol, while in rats and mice it is corticosterone [33; 34]. These glucocorticoids then “feedback” on glucocorticoid receptors (GRs) in the PVN and pituitary to limit activation of this system [35; 36; 37]. There are two types of feedback, fast and delayed, and both are thought to be mediated by GRs. In fast feedback, membrane GRs induce endocannabinoid suppression of the hypothalamus [38; 39]. In delayed feedback, GRs are shuttled from the cytosol to the nucleus where they can repress gene transcription of CRF, CRF1 receptors, and precursors of ACTH [40; 41; 42; 43; 44; 45]. In addition to their role in mediating negative feedback at the level of the pituitary and hypothalamus, GRs are also located in many other brain regions, including the hippocampus, septum, amygdala, hypothalamus, and nucleus of the solitary tract [35; 46; 47]. Glucocorticoids also can indirectly regulate negative feedback at the level of the PVN through activation of GRs in the hippocampus, which can inhibit this neuroendocrine response via connections to the PVN [29; 48]. Appropriate glucocorticoid negative feedback is critical because the metabolic and immune changes induced by glucocorticoid release that are adaptive in the short term, are detrimental to health if they endure [29; 49; 50].
Cortisol levels that are either too low or too high characterize certain psychiatric disorders. For example, in PTSD there is a complex dysregulation of cortisol that typically manifests as low waking and evening cortisol, but higher cortisol responses to trauma-related cues [51; 52; 53; 54]. Many patients with depression, in contrast, have high levels of cortisol [55; 56; 57; 58]. These alterations in cortisol levels are thought to precede the development of both PTSD and depression, and when symptoms of these disorders remit, cortisol levels normalize [55; 57; 59; 60]. Thus, dysregulation of the neuroendocrine response to stress is likely a risk factor for developing these stress-related psychiatric disorders.
2.2 Clinical studies demonstrating sex differences in HPA axis activity
Because women are more likely than men to suffer from disorders PTSD and depression, which are linked to HPA axis dysregulation, sex differences in the HPA axis have been evaluated. In healthy participants, baseline cortisol levels are typically comparable between men and women [61; 62] In contrast, sex differences in cortisol levels following stress have been reported, but the direction of these effects is inconsistent. Some studies find higher stress-induced cortisol levels in women, while others find higher stress-induced cortisol levels in men [61; 62; 63; 64; 65; 66]. These discrepant findings are thought to relate to the different stressors used in these studies, or other characteristics of the participants, such as their age or hormonal status [61; 63; 65]. Given these conflicting findings, it does not appear that sex differences in the neuroendocrine response to stress predispose women to stress-related psychiatric disorders. However, no longitudinal studies that are statistically designed (i.e., sufficiently powered) to detect sex differences have investigated whether sex differences in neuroendocrine responses to stress predict female vulnerability to PTSD and depression. Thus, this possibility cannot yet be ruled out.
In patient populations, sex differences in cortisol levels have been reported in PTSD and depression. Some have reported that women with PTSD have lower cortisol levels than their healthy counterparts, an effect not observed in men [67; 68]. Although not always replicated, these results do suggest that, in some cases, low cortisol levels distinguish women with PTSD from those without [69; 70]. Results from studies investigating sex differences in depression are more consistent. Depressed women typically have higher cortisol levels than depressed men [71; 72]. This sex difference is particularly pronounced following stressful and other negative life events [73; 74]. Together, these studies suggest greater HPA axis dysregulation in women than men with stress-related psychiatric disorders.
Elevated cortisol levels associated with depression have been attributed to impaired glucocorticoid negative feedback. This link is based, in part, from findings using dexamethasone suppression as in the dexamethasone suppression test (DST). Dexamethasone is a synthetic glucocorticoid that binds with high affinity to GRs in the brain and pituitary to suppress the release of cortisol in healthy people [75]. However, dexamethasone fails to suppress cortisol in 20–60% of patients with depression [76; 77; 78]. A modified version of this test, called the combined DST/CRF test, identifies roughly 80% of patients with depression [79; 80; 81]. In the combined DST/CRF test, dexamethasone treatment is followed by administration of CRF. When administrated alone, CRF acts at the level of the pituitary to increase ACTH and cortisol. However, if this CRF administration is preceded by dexamethasone treatment, as it is in the DST/CRF test, dexamethasone suppresses the ability of CRF to increase ACTH and cortisol levels, at least in healthy subjects. In contrast, in the majority of depressed patients, pretreatment with dexamethasone fails to suppress CRF-induced ACTH and cortisol release [79; 80; 81].
Sex differences in the DST and combined DST/CRF test have been evaluated. Although dexamethasone suppression is influenced by menstrual cycle phase, sex differences in cortisol release when dexamethasone is administered alone are not typically reported [82; 83; 84; 85]. However, the hormonal response to CRF following dexamethasone pretreatment is greater in both healthy and depressed women than in their male counterparts [86; 87]. This suggests that, while there may not be striking sex differences in glucocorticoid negative feedback, there are sex differences in CRF sensitivity. In support of this idea, the administration of CRF alone to healthy individuals increases the neuroendocrine response to stress to a greater degree in women than men [71; 88]. This effect appears to be mediated by the ovarian hormone surge during puberty, because cortisol output in response to CRF increases with pubertal stage in girls but not boys [89]. When considered together, these studies suggest that pubertal surge in ovarian hormones increases CRF sensitivity at the level of the pituitary in women.
Evidence suggests that CRF is elevated in brains of patients with depression and PTSD, perhaps as a result of CRF hypersecretion [90; 91; 92]. For example, patients with these disorders have higher levels of CRF in their cerebrospinal fluid, which are thought to reflect high levels of central CRF release [92; 93; 94; 95]. Postmortem studies have confirmed increased CRF expression in the brains of depressed patients [96; 97; 98; 99]. These studies also have revealed alterations in the expression of the CRF1 receptor subtype, which is the subtype that mediates the HPA axis and anxiety-related behavior [96; 100; 101; 102]. Additionally, single nucleotide polymorphisms on the CRF1 gene are found in patients with stress-related psychiatric disorders [103; 104; 105; 106]. Together these results suggest widespread dysregulation of the central CRF system in certain psychiatric diseases. This central dysregulation could alter the neuroendocrine response to stress. Given the evidence that CRF dysregulation is characteristic of these disorders and they occur more frequently in women, it is somewhat surprising that few clinical and postmortem studies looked for specifically for sex differences in the CRF system (but see [107]). However, preclinical models, detailed below, have identified sex differences in CRF function, as well as in molecular and hormonal regulation of the neuroendocrine response to stress. If confirmed in humans, these sex differences could help explain female vulnerability to stress-related psychiatric disorders.
2.3 Sex differences in HPA axis activity in preclinical models
Over 50 years ago Kitay (1961) reported sex differences in corticosterone levels in rats [108]. Since then many investigators have confirmed that female rats have higher basal and stress-induced corticosterone levels than male rats (e.g., [109; 110]. For example, compared to male rats, higher diurnal corticosterone peaks are observed in female rats [111]. Additionally, stressor exposure causes a greater increase in ACTH and corticosterone in female than male rats [108; 109; 110; 112; 113; 114; 115; 116; 117]. These neuroendocrine sex differences are established, in part, by ovarian hormones. Females in proestrus (the phase of the estrous cycle when estrogen and progesterone levels are elevated) have higher corticosterone levels than females in diestrus (when estrogen and progesterone levels are low) or males [118; 119; 120]. Collectively, these studies demonstrate that, unlike the equivocal findings in humans, reliable sex differences in HPA axis activity are found in rodents. The reason for the discrepancy between the human and rodent literature is unclear. If it were possible to control for the same number of variables in humans as it is in rodents (e.g., diet, housing conditions, etc.), perhaps consistently higher levels of cortisol would be observed in women compared to men. Alternatively, the neuroendocrine response in female rodents may simply be different than it is in women. Even if this is the case, the elevated glucocorticoids found in female relative to male rats appear analogous to the hormonal conditions found in depressed women (i.e., increased basal and stress-induced glucocorticoid release compared to depressed men). Thus, exploring the molecular mechanisms underlying this sex difference in rodents could prove to be clinically relevant.
Given that the HPA axis is regulated by a complex interplay of neural and peripheral tissues, there are many sites at which sex differences can occur. The pituitary response to CRF, for example, is greater in female than male rats, much like what is observed in humans [71; 88; 113]. At the level of the adrenals, estradiol, the major female estrogen, can enhance ACTH sensitivity, an effect that could contribute to greater corticosterone release in female compared to male rodents [121]. Sex differences also occur centrally that can alter the neuroendocrine response to stress. As noted, both CRF and vasopressin stimulate ACTH release. CRF expression in the PVN is often found to be higher in females than in male rodents [109; 114; 122] but see [123]. This effect is regulated by gonadal hormones, because proestrus levels of ovarian hormones enhance CRF expression, while androgens suppress CRF expression [114; 124; 125; 126]. Vasopressin expression in the PVN is also affected by gonadal hormones. Following stress, vasopressin expression is increased by treatment with estradiol but decreased by testosterone treatment [125; 127]. When taken together, these studies reveal sex differences throughout the HPA axis (Fig. 2). These sex differences are established, at least in part, by gonadal hormones. Thus, it is not surprising that both androgen and estrogen receptors are found in the PVN, pituitary, and adrenal glands [128; 129; 130; 131; 132; 133; 134; 135]. Exactly how these hormones modulate hypothalamic, pituitary, and adrenal activity remains largely unknown. However, direct regulation of vasopressin and CRF gene expression by estrogen is possible because promoter regions on both genes have estrogen responsive elements [136; 137]. The CRF promoter also has an androgen responsive element, which implicates testosterone in the regulation of CRF [138].
Figure 2.
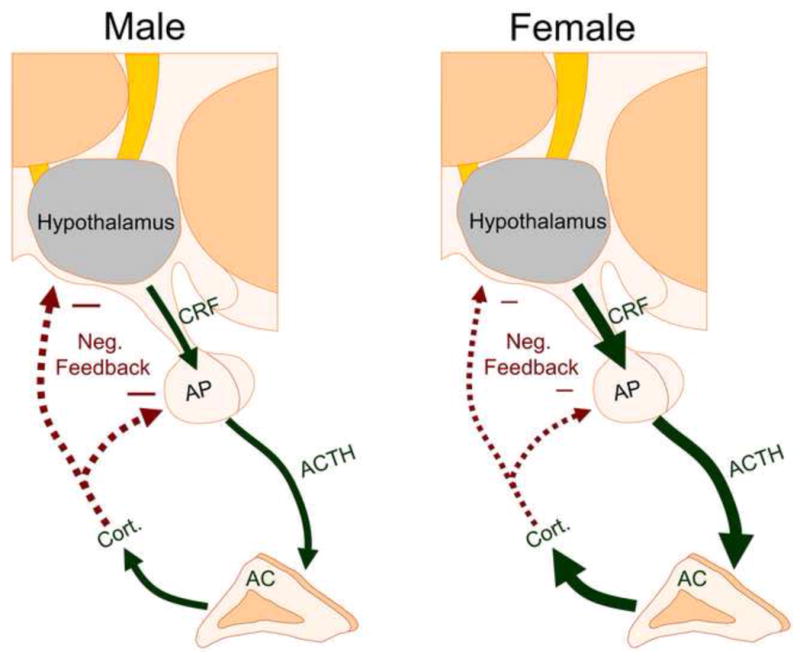
Schematic representing sex differences in the HPA axis response to stress in rodents. Compared to male rats (left panel), female rats (right panel) have greater stress-induced release of CRF, ACTH, and corticosterone (cort.) due to sex differences in the hypothalamus, anterior pituitary (AP), and the cortex of the adrenal gland (AC). Negative feedback (shown with the red arrows) is also decreased in females is thought to be due to sex differences in GR expression, GR translocation, and GABAergic inhibition. Reduced negative feedback in females can further increase the release of stress hormones.
2.4 Sex differences in glucocorticoid negative feedback in preclinical models
In addition to the sex differences that can potentiate corticosterone release in females, there are also sex differences in glucocorticoid negative feedback in rats (Fig. 2). Studies investigating the time course of stress-induced corticosterone release have revealed that it takes longer for corticosterone to return to baseline levels in female than male rats, suggesting slower feedback in females [108; 110; 115]. Estrogen mediates this effect because replacement with estradiol prolongs stress-induced corticosterone release and impairs dexamethasone suppression [119; 139; 140]. Glucocorticoid negative feedback is primarily mediated by GRs [35; 38; 41; 141]. Therefore, decreased GR expression in females compared to males could explain their slower feedback. Evidence for fewer GRs in females derives from studies showing that hypothalamic glucocorticoid binding and pituitary GRs are lower in female compared to male rats [142; 143]. Consistent with its role in modulating feedback, estrogen treatment downregulates GR expression in the hippocampus, hypothalamus, and pituitary [142; 144]. Therefore, slower glucocorticoid negative feedback in females may result from an estrogen-induced reduction of GR expression in the brain and pituitary. Another mechanism that can establish sex differences in glucocorticoid negative feedback involves GR translocation, the process by which GRs move from the cytosol to the nucleus to repress the transcription of genes (e.g., the CRF gene) to terminate the neuroendocrine response to stress [40; 43; 44]. During translocation, GRs are shuttled by co-chaperone proteins that either promote or inhibit their movement into the nucleus. Bourke et al. (2003) found that chronic stressor exposure during adolescence upregulates co-chaperones that inhibit GR translocation and impairs glucocorticoid negative feedback in female but not male rats [145]. This study highlights how sex specific molecular regulation of receptors can affect the neuroendocrine response to stress. Finally, other mechanisms that do not involve GRs also may differentially affect feedback in males and females. For example, estrogen purportedly reduces GABAergic inhibition of the PVN, an effect that could translate into reduced feedback in females [140]. The aforementioned findings were conducted by different groups using a variety of stress and endocrine manipulations, so the different mechanisms (e.g., GR expression, GR translocation, and GABAergic inhibition) were independently identified, but it is possible that all of these sex differences operate collectively to prolong the corticosterone release following stressor exposure in female rats.
Dysergulation of the neuroendocrine response to stress is probably the most studied factor implicated in the pathophysiological of PTSD and depression. This is, in part, because stress is an important etiological contributor to these disorders, and, in part, because cortisol levels are easily assessed using non-invasive techniques (e.g., saliva or urine collection) in humans. This focus on the peripheral levels of hormones involved in the HPA axis has revealed sex differences in cortisol in both PTSD and depression. However, clearly more work is needed to identify the central mechanisms that contribute these sex differences in humans. Perhaps when new technologies, such as positron emission tomography ligands to noninvasively study glucocorticoid receptors are developed [146], they will reveal underlying sex differences that help explain the female bias in these disorders. Until then, we must rely on preclinical data. The reviewed rodent studies suggest sex differences in a variety of molecular mechanisms, from receptor translocation to peptide expression, that can both potentiate stress-induced glucocorticoid release and slow negative feedback in female compared to male rats. Given the link between chronic stress, high glucocorticoid levels, and depression, if these mechanisms are confirmed in humans, they would help explain female vulnerability to this disease.
3. Sex differences in negative valence
3.1 Sex differences in the circuitry activated by stimuli with a negative valence
Stressful events not only initiate hormonal responses but they also trigger negative emotions, such as fear and anxiety. Typically these emotions activate adaptive cognitive and behavioral responses aimed at coping with the stressor [147; 148]. However, overactivation or dysregulation of these negative emotions increases susceptibility to stress-related psychiatric disorders [149; 150]. Anecdotally, it is often remarked that women are more emotional than men. Although this is overstated by the lay community, there is scientific evidence that women experience emotions, particularly those with a negative valence (e.g., fear, anger, sadness), with greater intensity than men [151; 152; 153]. Additionally, women often engage in emotion-focused coping strategies and report higher negative affect than men, characteristics that are predictive of anxiety and depressive symptoms [154; 155; 156]. Thus, some researchers have attributed sex differences in the rates of stress-related disorders to sex differences in emotional responses to stress [157; 158].
Sex differences in emotional reactivity may stem from sex differences in the neural circuits underlying emotional expression. The brain regions that respond to emotional stimuli with a negative valence include, among other areas, corticolimbic circuitry comprised of the amygdala, prefrontal cortex, and hippocampus [159; 160; 161; 162]. Several functional neuroimaging studies have identified sex differences in the magnitude of corticolimbic responses to emotional stimuli. For example, aversive stimuli and fear conditioning increased activity in the amygdala and certain cortical regions more in women than men [163; 164]. A recent meta-analysis similarly revealed that, compared to men, negative emotions in women elicit greater activation in regions including the left amygdala, anterior cingulate, and medial prefrontal cortex [165]. Additionally, women have greater hippocampal activation than men when encoding emotional words [166]. Together these studies reveal greater activation of corticolimbic circuits in women than men to negatively valenced emotional stimuli. Other studies have demonstrated that certain emotional stimuli engage different circuits in men and women. Negatively valenced words, for example, activated the left perirhinal cortex and hippocampus in women, but the right supramarginal gyrus in men [167]. Still other lines of work reveal that, in some cases, emotional stimuli activate the same structure in men and women, but the degree of this activation is distinguished by lateralization. Specifically, Cahill and others have demonstrated that memory tasks involving emotionally provocative stimuli activate the left amygdala in women but the right amygdala in men [168; 169; 170; 171]. When taken together, these findings suggest sex differences in brain activity in response to negative emotions. However, it is clear from the aforementioned studies that not all negatively valenced stimuli elicit the same pattern of sexually differentiated brain activation. These disparate results are likely attributable to differences in the type of negative stimuli used or other characteristics of the participants (e.g., participant age, hormonal status, etc.).
3.2 Sex differences in the corticolimbic circuitry in depression and PTSD
Because negatively valenced stimuli differentially activate corticolimbic circuitry more in in women than in men, traumatic events or chronic stressor exposure could have a greater impact on the female brain, perhaps by initiating a neuroplasticity that increases female vulnerability to stress-related psychiatric disorders. While subtle changes in neuronal plasticity (e.g., alteration in dendritic arborizations) are currently impossible to image in humans, large scale volumetric changes in the hippocampus, amygdala, and cortex can be observed, and some sex differences in these regions have been reported in patients with stress-related psychiatric disorders. One of the most reliably reported structural changes in psychiatry is the reduced hippocampal size observed in depression [172; 173; 174]. Although there is no sex difference in the overall hippocampal volume of depressed patients [175], women that fail to respond to antidepressant medications have smaller hippocampi than women that respond to treatment, an effect not observed in men [176]. Additionally, a recent paper found that depressed men have abnormities in prefrontal-striatal circuits, but women have abnormities in prefrontal-limbic circuits, which, as noted, processes negative emotions [177]. Volumetric changes in corticolimbic circuits have also been assessed in patients with PTSD, but structural sex differences have not observed [178; 179].
Although widespread sex differences in corticolimbic structure are not reported in all stress-related disorders, the volumetric reductions that have been observed in some women do suggest greater alterations in their underlying circuitry. However, because the vast majority of imaging studies employ cross-sectional designs, it is difficult to determine the causal relationship between structural sex differences and disease prevalence. If, for example, volumetric reductions occur in women following disease onset, it would indicate that stress-related psychiatric disorders result in greater corticolimbic remodeling of the female brain. Alternatively, if these sex differences precede the onset of depression and PTSD, then this would suggest that these preexisting factors can increase female vulnerability to these diseases. Although determining causality is difficult, studies of one group of monozygotic twins discordant for combat exposure revealed that a smaller hippocampus was a risk factor for developing PTSD, while reduced gray matter density in pregenual anterior cingulate cortex occurred as a result of the disorder [180; 181]. However, this cohort only included males, so these results do not address whether sex differences in corticolimbic circuitry predict sex difference in disease incidence. Because causality is difficult to determine in clinical populations, scientists have turned to preclinical models to begin to address the nature of these relationships, as well as to investigate cellular sex differences that cannot be evaluated with current approaches in humans.
3.3 Sex differences in neuronal morphology
Non-human animal models investigating sex differences in corticolimbic circuitry typically focus on identifying sex differences in neuronal morphology rather than evaluating volumetric modifications. The most studied morphological changes include alterations in dendritic branches and spines, small protrusions on branches that are the site of excitatory synapses. Higher numbers of branches and spines translate into increased connectivity between neurons, thus these morphological changes are thought to be functionally relevant (for review see [182; 183; 184; 185]). Branches and spines are dynamically regulated in an activity-dependent manner [186; 187; 188]. Changes in the environment can rapidly, within an hour, alter spine density [186]. Yet modifications of spines and branches can persist for weeks and, in some cases, even throughout the lifetime of the animal [189; 190].
The majority of studies investigating neuronal morphology use only male subjects. However, when females are included, sex differences in spine density and dendritic morphology in corticolimbic regions have been observed [191; 192; 193]. The seminal work of Gould and Woolley sparked interest in spine alterations in the hippocampus, when they demonstrated that proestrous levels of estrogen increased the density of spines on apical dendrites of pyramidal neurons in the CA1 region in the female rat [194; 195]. Given that males lack high circulating estrogen levels, it is not surprising that the spine density of male rats is lower relative to that of females in proestrus [192]. The morphology of neurons in the amygdala and prefrontal cortex is also sexually differentiated. Compared to female rats, males have longer dendrites and denser spines on neurons in the left posterodorsal subnucleus of the medial amygdala and basolateral nucleus of the amygdala, respectively [196; 197]. Male prefrontal cortical neurons also have more branches than those of females [193]. Despite the fact that these studies clearly demonstrate morphological sex differences throughout corticolimbic circuits, the functional significance of these sex differences remain unknown. It has been proposed that sex differences in hippocampal dendritic spine density underlie sex differences in the modulation of learning, while sex differences in the medial amygdala establish sex-specific social behaviors, but no studies have causally established these relationships [197; 198]. Nevertheless, given that a dysregulation of corticolimbic circuits is observed in sex biased stress-related disorders, the possibility remains that these cellular sex differences predispose females to stressful events, an idea that clearly warrants further investigation.
3.4 Stress-induced sex differences in neuronal morphology in preclinical models
The relationship between stress, psychiatric disorders, and corticolimbic circuitry has prompted many investigators to evaluate stress-induced modifications of neuronal morphology in the hippocampus, amygdala, and cortex in rodent models. Disappointingly, few of these studies include females. For example, chronic stress induces hypertrophy of dendrites in the amygdala in males, an effect thought to be relevant to enhanced emotionality [190; 199]. Surprisingly however, this has not been investigated in females [200]. Fortunately, several studies have examined stress-induced alterations in neuronal morphology in the prefrontal cortex and hippocampus in both males and females. As detailed below, these studies identified cellular sex differences that may be relevant for understanding sex differences in the etiology of certain psychiatric disorders.
Sex differences in the remodeling of dendrites in the prefrontal cortex are observed following chronic stress. Specifically, Garret and Wellman (2009) demonstrated that chronic stress induces dendritic atrophy of neurons in the medial prefrontal cortex in male rats. However, exposure to the same chronic stressor in female rats results in the opposite effect (i.e., stress induces dendritic hypertrophy of female cortical neurons). These stress-induced morphological sex differences are modulated by ovarian hormones, as they are not observed in ovariectomized female rats [193]. Because the prefrontal cortex regulates fear and anxiety via its interconnections with the amygdala, investigating sex differences within this circuit may be clinically relevant [201; 202]. Thus, Shansky and colleagues (2009, 2010) compared the morphology of prefrontal neurons that project to the basolateral amygdala to those that project elsewhere [203; 204]. In males, the cortical neurons that project to the basolateral amygdala are spared from the dendritic retraction induced by chronic stress [203]. In contrast, it is precisely these basolateral amygdala projecting neurons that have longer and more complex dendrites in stressed females that are ovariectomized with estradiol replacement compared to their ovariectomized counterparts that were not treated with estradiol and unstressed controls [204]. Although males and females were not directly compared in these studies, these data do suggest that chronic stress induces circuit-specific sex differences. It is unclear exactly how these morphological sex differences translate into sex differences in responses to negatively valenced stimuli, because these modifications have yet to be linked to behavioral endpoints. However, it has been hypothesized that the retraction of dendrites observed in males following chronic stressor exposure may be a compensatory response aimed at protecting their neurons from overactivation [205]. If supported by future studies, this would indicate that female neurons, which instead sprout dendrites in response to stress, would be more vulnerable to overactivation and perhaps excitotoxicity.
Neurons in hippocampus are also sensitive to the effects of stress. Chronic stress induces retraction of dendrites in the CA3 region of the hippocampus in male rats [206; 207]. Exposure of males to the same chronic stressor also impairs spatial learning, which requires an intact hippocampus [208; 209; 210; 211]. Pharmacological manipulations that prevent the stress-induced dendritic retraction in males also prevent stress-induced spatial learning deficits [212; 213; 214]. Therefore, it has been proposed that the disruptions in spatial learning following stress are the result of dendritic remodeling [212; 215]. Interestingly, these effects are not observed in female rats [206]. Specifically, chronic stressor exposure in female rats induces either a mild dendritic retraction in CA3 or has no effect on dendritic morphology [206; 216]. Spatial learning in females also is unaffected, or in some cases improved, by chronic stress [216; 217; 218]. These studies highlight the fact that females are not always more adversely impacted by stress, but rather that stress affects males and females differently. It is these differential effects of stress that likely contribute to sex biases in many disorders, including those that occur more frequently in males.
In addition to the aforementioned effects of chronic stress on the hippocampus, acute stressor exposure also can modify hippocampal neurons. For example, Shors and colleagues (2001) demonstrated that exposure to an acute stressor (30 min restraint plus periodic tail shock) alters the density of dendritic spines on hippocampal neurons in a sex-specific manner [192]. Specifically, acute stress increases the density of spines on apical dendrites of pyramidal neurons within the CA1 region in male rats. Conversely, in female rats, acute stress decreases spine density within the same area. These spine changes correlate with sex differences in a classical eyeblink conditioning task, with the same acute stressor that modifies spines enhancing learning on this task in male rats, but impairing learning in female rats [219; 220]. Although it has yet to be determined whether sex differences in spine density directly cause sex differences in conditioning, both effects are organized by the perinatal testosterone surge and require NMDA receptors, suggesting that there is a common underlying mechanism [183; 221; 222; 223; 224].
The reviewed results indicate that the hippocampus, amygdala, and cortex contain sexually differentiated dendritic morphology and/or spine density. These morphological differences likely result from sex differences in intracellular signaling events that induce cytoskeletal remodeling. Although the precise signaling mechanisms responsible for morphological sex differences remain largely unknown, many studies have linked cellular remodeling to circulating estrogen in females and to glucocorticoids in males. In females, estradiol treatment is thought to increase spines in the hippocampus via activation of a signaling pathway involving RhoA, LIM kinase, and cofilin, although other signaling molecules (e.g., Akt) may also play a role [225; 226; 227]. The signaling pathways underlying estrogen modulation of morphology in other regions are yet to be determined. In males, glucocorticoids critically mediate stress-induced dendritic retraction in the hippocampus and prefrontal cortex [228; 229]. In addition to glucocorticoids, neurotransmitters including glutamate, dopamine, and serotonin also have been implicated in dendritic remodeling in males [229; 230; 231; 232; 233]. Although the downstream cellular events initiated by glucocorticoids and neurotransmitters that underlie stress-induced plasticity in males remain to be determined, the wide variety of hormones and neurotransmitters identified thus far suggest that multiple intracellular mechanisms can lead to morphological alterations.
It is clear from the reviewed clinical and preclinical studies that sex differences in corticolimbic circuits occur from the cellular to the systems level. However, more research is needed to integrate these levels of analysis to truly understand how cellular sex differences in the hippocampus, amygdala, and cortex translate into sex differences in brain structure and function in patient populations. In preclinical studies, for example, tools to induce morphological changes should be developed and employed to determine whether sex differences in morphology actually cause sex differences in stress reactivity or anxiety-related behavior. In the clinic, a prospective study tracking whether greater feminization of corticolimbic responses to emotional stimuli predict disease onset would reveal whether known neuronal sex differences in healthy subjects are actually risk factors for disorders, such as depression and PTSD. Despite the fact that future studies should employ a more integrative approach, based on the existing data one can posit that molecular sex differences can lead to increased corticolimbic activation to negatively valenced stimuli present during stressor exposure in females. This effect would in turn increase female vulnerability for developing stress-related psychiatric disorders.
4. Sex differences in hyperarousal
4.1 Sex differences in the hyperarousal symptoms of depression and PTSD
A core feature of stress-related psychiatric disorders is hyperarousal, a maladaptive state that leads to agitation, restlessness, lack of concentration, and cognitive disruptions. Interestingly, sex differences in hyperarousal have been identified. For example, heightened arousal defines one of the symptom clusters of PTSD, and these hyperarousal symptoms are often more pronounced in women than men [234; 235; 236]. In both PTSD and depression, female patients typically report more sleep disruptions than men, an effect thought to reflect high levels of arousal [237; 238; 239; 240; 241] but see [242]. Additionally, depressed women ruminate (i.e., have recurrent negative thoughts) more than men [243; 244]. These ruminations predict depressive symptoms and are associated with heightened arousal [245; 246; 247; 248; 249]. These reported sex differences in hyperarousal most likely stem from sex differences in brain arousal centers.
The brain system thought to mediate the hyperarousal observed in patients with stress-related psychiatric disorders is the locus coeruleus (LC)-norepinephrine system [91; 250; 251; 252; 253; 254]. The LC is a compact pontine nucleus that provides norepinephrine throughout the brain via its widespread projection system [255]. The LC innervates all levels of the neuroaxis. It the primary source of norepinephrine for the forebrain and the sole source for the hippocampus and cortex[256]. Typically activation of this system releases moderate amounts of norepinephrine to coordinate arousal, attention, and vigilance [257; 258; 259]. However, overactivation of the LC system leads to excessive norepinephrine release, an effect that can result in hyperarousal [260; 261]. Because hyperarousal characterizes symptoms of PTSD and depression, it is not surprising that high concentrations of norepinephrine in the cerebrospinal fluid are reported in patients with these disorders [91; 262; 263]. Cerebrospinal fluid levels are thought to reflect central not peripheral norepinephrine release, so these results indicate a hypernoradrenergic state within the brains of some patients [91]. Sex differences in cerebrospinal fluid levels of norepinephrine have not been specifically investigated. However, if evidence of greater central hypersecretion of norepinephrine was found in women than men, this effect could account for their prominent hyperarousal symptoms.
4.2 Sex differences in locus coeruleus structure
Several preclinical studies have evaluated sex differences in the LC-norepinephrine system. For example, on a structural level, the LC is comprised of more neurons in female than male rats of the Wistar strain, however this does not appear to be the case for all rat strains [264; 265; 266]. In addition to neuron number, sex differences in LC dendritic morphology have been identified. Specifically the dendrites of LC neurons are longer and more complex (i.e., have more branches and ends) in female rats and mice compared to their male counterparts [267; 268]. Although LC neurons do not have spines, synapse density can be gauged using immunoreactivity for synaptophysin, a synaptic vesicle protein. This approach revealed that the pericoerulear regions (peri-LC) into which LC dendrites extend had denser synaptophysin labeling in female than male rats, suggesting that the dendrites in females receive greater synaptic input [268]. These sex differences in dendritic morphology could bias the type of afferent information that the LC receives in females, because there is a topographical pattern of LC afferents, such that some terminate near the nucleus, while others terminate on dendrites that extend for hundreds of microns into the peri-LC region [269]. The shorter dendrites of males would still be able to easily receive inputs that terminate in the nuclear LC, such as those the project from the nucleus paragigantoceullaris, the nucleus prepositus hypoglossi, and Barrington’s nucleus [269; 270; 271]. However, the longer and more complex dendrites of females would likely make more connections with axons that terminate in the peri-LC regions (Fig. 3). The peri-LC region receives CRF innervation from the PVN and input from limbic regions, including the central nucleus of the amygdala and bed nucleus of the stria terminalis [272; 273]. Thus, compared to males, the LC of female rodents appears to be poised to receive and process more emotion-related information. This structure could support greater arousal responses to emotional stimuli present during stressor exposure in females.
Figure 3.
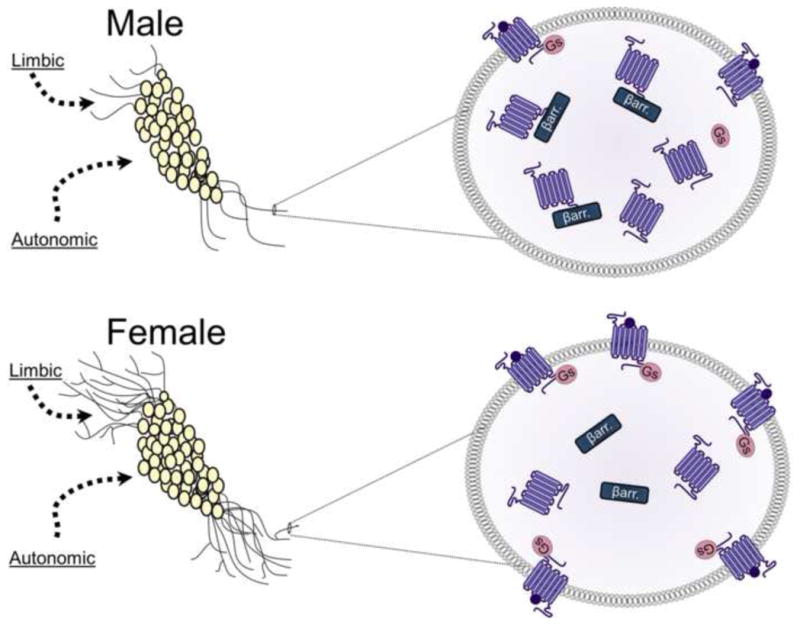
Schematic depicting sex differences in the LC–arousal system in rodents. The images on the left depict LC neurons with dendrites that are longer and more complex in females (bottom left panel) compared to males (top left panel). The longer dendrites of females are more likely to receive limbic afferents that terminate in peri-LC region. However, males and females would likely receive comparable amounts of input from autonomic regions that synapse near the cell bodies. The images on the right depict magnified views of LC dendrites to illustrate sex differences in the CRF1 receptor coupling and trafficking. The top right panel shows how CRF1 receptors (purple) of males associate with βarrestin2 (βarr.) and internalize following stressor exposure. In contrast, the bottom right panel shows how CRF1 receptor of females couple to Gs and traffic to the plasma membrane following stressor exposure.
4.3 Sex differences in the modulation of the locus coeruleus by stress
The LC arousal system was first linked to arousal and vigilance based on the electrophysiological properties of LC neurons. These neurons have two modes of firing: tonic and phasic. Switching between tonic and phasic modes of firing is thought to facilitate shifts in arousal and attention that can help the organism respond to a changing environment. Tonic firing is positively correlated with EEG and behavioral measures of arousal and is associated with scanning the environment, cognitive flexibility, and liable attention [257; 258; 274; 275]. In contrast, phasic firing, which is synchronized in response to sensory stimuli, is associated with focusing attention toward discrete stimuli [275; 276; 277].
Given that the electrophysiological properties of LC neurons mediate different states of arousal and that hyperarousal symptoms are more prominent in some women, sex differences in LC neuronal physiology were evaluated [278]. On most physiological measures, male and female neurons are comparable. However, LC neuronal responses were distinguished by stress. Hypotensive stress causes a moderate increase in tonic firing in male rats [278; 279]. In females, though, exposure to the same stressor causes a large increase in LC neuronal firing, such that their neurons fired much faster than those of stressed males [278]. This enhanced neuronal sensitivity to stress in females suggests sex differences in the mechanisms underlying stress activation of LC neurons.
Studies conducted in male rats have determined that stress activates LC neurons via CRF. This link is based, in part, on the fact that local infusions of CRF into the LC increase tonic firing and reduce phasic firing to sensory stimuli, thereby mimicking the electrophysiological effects of stress [280; 281]. Stress also alters downstream endpoints of LC activation, such as heightened EEG and cortical norepinephrine release, via CRF-dependent mechanisms [282; 283; 284]. Moreover, effects of hypotensive stress on LC physiology are blocked by CRF1 receptor antagonism [285; 286; 287]. Collectively, these findings suggest that stressor exposure causes CRF to be released into the LC, which shifts the physiological properties of LC neurons into a high tonic-low phasic mode that alters arousal and attention by increasing norepinephrine release in target regions. In response to acute or moderate stressors, the increase in tonic activation is thought to be adaptive as it promotes scanning the environment for danger and increasing cognitive flexibility. Indeed, male rats given moderate amounts of CRF into the LC performed better on the attentional set shifting task of cognitive flexibility than their vehicle infused counterparts [288]. However, if this system was activated inappropriately or persistently, it could lead to a high arousal state that would disrupt focused attention and eventually become maladaptive.
Because the effects of stress on LC neuronal physiology are mediated by CRF, the finding that LC neurons of females are more responsive to hypotensive stress than those of males could be explained by sex differences in CRF sensitivity. In support of this idea, when sex differences in LC neuronal firing following local infusions of CRF were assessed, it was determined that the CRF dose response curve for LC activation is shifted to the left in female compared to male rats [278]. In other words, a dose of CRF that failed to activate neurons in males causes a significant increase in tonic firing in females. It should be noted that these effects are not mediated by adult hormonal status in females, suggesting other mechanisms drive this sexual difference [278]. These data confirm that female LC neurons are more sensitive to CRF. Thus, stressful events could more easily shift LC neuronal firing into a high tonic mode in females, thereby heightening arousal and disrupting focused attention.
Sex differences in CRF modulation of LC neurons are also observed in rats with a history of stress [278]. In male rats, it has been demonstrated that footshock or swim stress given 24 h prior to CRF administration produces a complex change in the CRF dose–response curve for LC activation, such that the linear portion of the curve is shifted to the left (indicative of sensitization), while the maximum is decreased relative to that of unstressed males [278; 289; 290]. Interestingly, the effect of prior stress is not observed in females, as their CRF-dose response curve is similar to that of unstressed females, perhaps because unstressed females are already at ceiling levels of CRF activation [278]. Taken together these electrophysiological studies reveal a complex pattern of CRF modulation of LC neurons that is sex specific.
4.4 Sex differences in the CRF1 receptor coupling and signaling
Sex differences in LC physiology are linked to sex differences in the CRF1 receptor, which mediates the effects of CRF on LC neurons [291; 292]. CRF receptors are members of the B1 group of the G protein-coupled receptor (GPCR) superfamily, and as such, CRF receptors bind/couple to the GTP-binding proteins [100; 293]. The CRF1 receptor preferentially couples to Gs, so when it is activated, the CRF1 receptor signals though the cyclic adenosine monophosphate (cAMP) and protein kinase A (PKA) second messenger cascade [294; 295]. This signaling pathway mediates the activation of LC neurons by CRF, purportedly by phosphorylating potassium channels [291]. Given that CRF activation of cAMP signaling drives LC firing, it was not surprising that increased LC neuronal responses to CRF observed in unstressed females are attributable to their greater cAMP signaling [296]. Specifically, local infusion of a cAMP antagonist into the LC revealed that, while only about 50% of LC response to CRF is cAMP mediated in unstressed males, nearly all of the LC response to CRF is cAMP mediated in unstressed females.
Sex differences in cAMP-PKA signaling are thought to be mediated by sex differences in Gs association with the CRF1 receptor [296]. Specifically, immunoprecipitation of the CRF1 receptor from cortical tissue—which was used because of the high levels of CRF1 protein required for this technique—revealed that more Gs protein was pulled down with the CRF1 receptor in female than male rats. This indicated greater CRF1-Gs coupling in females than in males (Fig. 3). This increased coupling was observed in females, regardless of adult hormonal status. Following swim stress, CRF1–Gs coupling increased in males to a level similar to that observed in females, but stress did not alter coupling in females [296]. The sex differences in cortical CRF1–Gs coupling mirrored sex differences in LC electrophysiology. These data suggest that greater CRF1 receptor binding to Gs in females facilitates greater cAMP signaling, resulting in increased LC firing relative to unstressed males. Notably, these were the first studies to link sex differences in physiology to sex differences in the coupling and signaling of a stress-related receptor.
4.5 Sex differences in the CRF1 receptor trafficking
Sex differences in LC-mediated arousal are also linked to sex differences in CRF1 receptor internalization [296]. Like most G-protein coupled receptors, CRF1 receptors are thought to desensitize and internalize in response to excessive ligand or agonist binding. This process is initiated when G-protein-coupled receptor kinase 3 phosphorylates the receptor, thereby recruiting the βarrestin2 protein [297; 298]. βarrestin2 then traffics the receptor from the membrane into clathrin-coated pits for endocytosis [100; 297; 299]. These internalized receptors are ultimately recycled back to the membrane or degraded. This standard model of CRF1 receptor internalization appears to accurately reflect what happens in male rats. Immunoelectron microscopy studies revealed a higher proportion of cytosolic receptors in male rats exposed to either a local infusion of CRF or swim stress than in controls, indicating that these manipulations induce CRF1 receptor internalization [300; 301]. However, exposure to the same swim stressor in females fails to induce internalization [296]. In fact, there are a greater proportion of CRF1 receptors on the plasma membrane of LC neurons in stressed than unstressed females, suggesting that stress induces trafficking in the opposite direction in females than it does in male rats (Fig. 3). These reported sex differences in internalization may result from sex differences in CRF1 receptor association with βarrestin2. An immunoprecipitation study of rat cortical tissue revealed that swim stress increases βarrestin2 association with the CRF1 receptor in male, but not female rats [296]. If this also occurs in the LC, it would explain a lack of receptor internalization in females.
Receptor internalization is thought to be a compensatory mechanism aimed at attenuating neuronal responding under conditions of excessive ligand release [302; 303; 304]. Therefore, the lack of CRF1 receptor internalization in females could make them more vulnerable to hyperarousal under conditions of CRF hypersecretion, as can occur in chronic stress, PTSD, and depression. To test sex differences in the electrophysiological properties of LC neurons under conditions of CRF hypersecretion, CRF overexpressing mice were used and their physiology was compared to that of wild type animals [267]. LC neurons of male and female wild type mice are comparable on most measures, much like what was observed in unstressed rats [267; 278]. In female CRF overexpressing mice, however, LC neurons fire roughly 3 times faster than wild type controls. This result was not surprising because CRF activates LC neurons and these mice overexpress CRF. However, it was surprising to find that the tonic firing of LC neurons in male CRF overexpressing mice is similar to that of wild type animals [267]. This suggested that a compensatory mechanism in male CRF overexpressing mice keeps their firing at normal levels. Male and female overexpressing mice have similarly high levels of CRF expression in the LC (much higher than wild type animals), so sex differences in physiology are not attributable sex differences in CRF expression [267]. Instead, these sex differences are associated with sex differences in CRF1 receptor internalization. Specifically, an immunoelectron microscopy study revealed a high proportion of CRF1 receptors in the cytosol of male CRF overexpressing mice, but a high proportion of receptors on the plasma membrane of female CRF overexpressing mice [267]. Thus, CRF1 receptors internalize in male, but not female CRF overexpressing mice. This pattern mirrored what was observed in stressed rats [267; 296]. These sex differences in CRF1 receptor internalization explain the sex differences in LC physiology observed in CRF overexpressing mice. Unlike male CRF overexpressing mice that likely normalize their firing via internalization, female CRF overexpressing lack internalization and thus their LC neurons can be continually activated by CRF. This effect would likely lead to a state of heightened arousal in female CRF overexpressing mice.
Together, this body of literature suggests that the CRF1 receptor is very different with respect to the way it binds proteins in males and females. The preferential coupling of the female CRF1 receptor to Gs leads to more activation of the cAMP-PKA signaling cascade, while the preferentially binding of the male CRF1 receptor to βarrestin2 is linked to increased internalization. It is important to note that these sexually distinct protein interactions may have even broader implications. βarrestin proteins are not only critical for receptor internalization, but their activation can initiate signaling cascades, including mitogen-activated protein kinases (e.g., ERK2, JNK3, and p38), tyrosine kinases (e.g., c-SRC, Hck), and small GTPases (e.g., RhoA) (for review see [305; 306; 307; 308]). These data have led us to hypothesize that the signaling of the CRF1 receptor is sex biased, such that it signals more through βarrestin mediated pathways in males and Gs mediated pathways in females [309; 310; 311]. Given that Gs and βarrestin typically activate distinct signaling cascades, activation of the CRF1 receptor would lead to different downstream cellular events in males compared to females. Thus, this sex biased signaling could translate into sex specific responses to stress. In females, the link between Gs biased signaling of the CRF1 receptors and hyperarousal can potentially predispose them to stress-related psychiatric disorders. However, it is important to note the male biased βarrestin signaling of CRF1 receptors, could underlie stress-related disorders that are more prevalent in man. Thus, sex biased signaling may be an important determinant of sex differences in disease vulnerability.
5. Interactions between neuroendocrine, corticolimbic, and noradrenergic arousal systems
It is clear that PTSD and depression share a common pathophysiology which includes dysregulation of the HPA axis, corticolimbic circuits, and arousal centers. Sex differences from the molecular to the systems level that occur within these circuits can increase vulnerability to these disorders and exacerbate their presentation in females. It is possible that symptoms of stress-related psychiatric disorders in some women are attributable only to sex differences within one circuit. For example, sex differences in amygdala activation to negatively valenced stimuli may increase emotional responses to stress and drive depressive symptoms in women. However, another possibility is that these sex differences work in concert to increase pathology in women. This scenario is possible because the hypothalamus, hippocampus, amygdala, prefrontal cortex, and locus coeruleus are interconnected and can influence one another. Thus, perturbations in one circuit could, in turn, perturb another circuit. For example, greater emotional arousal in females could result from the combined effects of sex differences in amygdala activation and sex differences in LC dendritic morphology. Because negatively valenced emotional stimuli cause greater activation of the amygdala in females, a stimulus that is subthreshold for amygdala activation in males could have an effect in females. Even this small increase in amygdala activation in females could lead to greater emotional arousal because their amygdala response would be amplified by their greater amygdala-LC connectivity. Thus, in response to traumatic events that trigger extreme emotional reactions, this combination of sex differences would increase the likelihood that females would shift into the dysregulated state of hyperarousal. Taken together then, the pervasive sex differences in stress response systems have the potential to create multiple substrates that can, via independent or synergistic action, contribute to the higher rates of depression and PTSD in women.
6. The role of gonadal hormones and sex chromosome compliment in PTSD and depression
Sex differences in the brain are typically established by activational effects of gonadal steroid hormones, organizational effects of gonadal steroids, or sex chromosome effects [312; 313; 314; 315]. Activational effects of hormones occur when circulating gonadal hormones act on brain structures to alter behavior. These activational effects disappear when the circulating hormones are removed. In contrast, organizational effects of hormones occur when gonadal hormone surges early in development (e.g., prenatal) or later in development (e.g., puberty) permanently alter brain structures [313; 314; 315; 316]. In addition to hormonal effects, sex differences in the brain can also be established by the different complement of sex chromosome genes present on XX (genetically female) versus XY (genetically male) chromosome pairs. For the majority of studies discussed in this review, the way in which sex differences arise remains unknown. However, as detailed below, there is some evidence that all three mechanisms can contribute to sexual differentiation of the circuitry that mediates stress-related psychiatric disorders, at least in preclinical models.
Epidemiological data support a role for ovarian hormones in depression. The increased rates of depression in women compared to men emerge following puberty (when ovarian hormone circulation increases) and remain high until menopause (when levels of ovarian hormones drop), which suggests that activational effects of ovarian hormones may be critical [17; 18; 317]. However, it is not simply that high levels of ovarian hormones are a risk factor for depression because in some women depressive symptoms are precipitated by a drop in estrogen, as can occur during the luteal phase of the menstrual cycle, the postpartum period, or menopause [318; 319; 320; 321]. It has therefore been proposed that sudden changes in estrogen can trigger depression [321; 322; 323]. There is evidence that the depressive symptoms induced by the decreased estrogen levels associated with menopause can be ameliorated by estrogen treatment, again indicating that this effect is activational in nature [324; 325]. In addition to its role in depression, estrogen also has been linked to PTSD. In particular, low estrogen levels are thought to increase vulnerability to PTSD in women with a history of trauma exposure [326]. This result suggests the exciting possibility that estrogen replacement therapy could ameliorate PTSD symptoms in these women. If true, this would suggest an activational role for estrogen in PTSD as well. As previously detailed, activational effects of estrogen can alter levels of stress-related neuropeptides and neuronal morphology within stress circuits in preclinical studies [136; 137; 192; 204]. Thus, if these findings hold true in humans, activational effects of estrogen on neuropeptides and morphology could contribute to the sex bias in depression and PTSD.
Circulating estrogen is not the only hormonal factor that can contribute to sexual differentiation. As noted, testosterone can regulate CRF expression, so the role of circulating androgens in masculinizing the stress response should also be considered [138]. In addition, the perinatal surge of testosterone that leads to brain masculinization in males also appears critical. This surge establishes sex differences in stress effects on dendritic spines in the hippocampus and classical conditioning observed in adult rats, and likely has other similarly striking effects on stress response circuitry [223; 224; 327]. In addition to organizational effects of early testosterone in males, the pubertal surge of gonadal hormones in both males and females can organize certain sexually dimorphic behaviors [314; 316]. Thus, it is conceivable that some sex differences in stress systems occur as a result of puberty. Because sex differences in CRF1 receptor function are unaffected by circulating hormones, it is likely that this particular mechanism is established by organizational effects of gonadal hormones.
As noted, an alternative possibility to gonadal hormone driven sexual differentiation is that genes on the sex chromosomes themselves differentially influence stress responses in males versus females. Although this possibility is, at this point in time, underexplored, sex chromosome complement independent of gonadal steroid exposure has been shown to contribute to sex differences in certain anxiety-related behavior in mice in one paper [328]. It is still unclear exactly how genes on these chromosomes result in changes in anxiety, but this study does underscore that sex chromosome complement should also be considered when trying to elucidate sex and stress interactions.
7. Implications for treatment
Investigating sex differences in the mechanisms that contribute to the sex bias in stress-related psychiatric disease is not only important for understanding disease vulnerability, but it is also critical for developing better treatments for these disorders. Developing new treatments for PTSD and depression is vital, because a large portion of patients with these disorders are treatment-resistant [329; 330]. The preclinical data reviewed here reveal several potential targets for new pharmacotherapies with greater specificity for females. For example, a reduction in glucocorticoid levels could be achieved by targeting co-chaperones that facilitate GR translocation. If developed, these compounds could facilitate glucocorticoid negative feedback, thereby normalizing the high levels of cortisol that are prominent in depressed women. The data suggesting a sex bias in the signaling of the CRF1 receptor also have important implications for drug development. “Biased agonists” that shift signaling towards βarrestin-mediated pathways and away from Gs mediated pathways have been developed for the angiotensin II receptor and the β2-adrenergic receptor [331; 332; 333; 334]. If similar compounds were designed to bias the CRF1 receptor towards βarrestin, they could potentially make females more resilient to the hyperarousal induced by stress.
The studies reviewed here provide examples of a number of sex differences at the molecular level. Despite this body of work, the majority of preclinical studies exclusively use male subjects. This male bias is particularly egregious in the fields of neuroscience and pharmacology, where single-sex studies of male animals outnumber those of females roughly 5 to 1 [335]. Given that there are sex differences in potential molecular targets of therapeutics, it is possible that certain compounds will work better in females than males, or that some drugs would only be effective in females. If novel therapeutics continue to be screened exclusively in males, drugs that would work well in females may never be identified. Thus, the male bias in preclinical research may deny women important treatments for stress-related disorders.
8. Conclusions
This review explored sex differences in the neurobiological bases of PTSD and depression as a way to illustrate how sex differences in stress response systems can contribute to sex biases in psychiatric disorders. Preclinical data demonstrates sex differences in several cellular and molecular mechanisms, including cell signaling, peptide expression, hormone release, receptor trafficking, synaptogenesis, and dendritic remodeling. What is remarkable is that, at each level of analysis, sex differences exist that can be linked to increased endocrine, emotional, and/or arousal responses to stress in females compared to males. Additional work is needed to determine whether the mechanisms identified in preclinical studies are also apparent in clinical populations. However, at minimum, these data highlight the multitude of factors that have the potential to establish sex differences in disease prevalence and presentation. More broadly, the study of sex differences in stress response systems will likely impact the way disorders are treated in the future, as they reveal novel targets for the development of novel pharmacotherapies that can be specifically tailored to the physiology of women.
Highlights.
Psychiatric disorders that occur more often in women than men are related to stress
Sex differences in stress response systems contribute to this sex bias in disease
Examples of these sex difference from the molecular to systems level are detailed
Studying sex differences in stress systems can reveal novel pharmaceutical targets
Acknowledgments
Supported by PHS grants MH092438 and MH040008.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.McCrory E, De Brito SA, Viding E. The link between child abuse and psychopathology: a review of neurobiological and genetic research. Journal of the Royal Society of Medicine. 2012;105:151–6. doi: 10.1258/jrsm.2011.110222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bremne JD, Vermetten E. Stress and development: behavioral and biological consequences. Development and psychopathology. 2001;13:473–89. doi: 10.1017/s0954579401003042. [DOI] [PubMed] [Google Scholar]
- 3.Heim C, Binder EB. Current research trends in early life stress and depression: review of human studies on sensitive periods, gene-environment interactions, and epigenetics. Experimental neurology. 2012;233:102–11. doi: 10.1016/j.expneurol.2011.10.032. [DOI] [PubMed] [Google Scholar]
- 4.Koolhaas JM. Coping style and immunity in animals: making sense of individual variation. Brain Behav Immun. 2008;22:662–7. doi: 10.1016/j.bbi.2007.11.006. [DOI] [PubMed] [Google Scholar]
- 5.Mao WC, Bardwell WA, Major JM, Dimsdale JE. Coping strategies, hostility, and depressive symptoms: a path model. Int J Behav Med. 2003;10:331–42. doi: 10.1207/s15327558ijbm1004_4. [DOI] [PubMed] [Google Scholar]
- 6.Matheson K, Anisman H. Systems of coping associated with dysphoria, anxiety and depressive illness: a multivariate profile perspective. Stress. 2003;6:223–34. doi: 10.1080/10253890310001594487. [DOI] [PubMed] [Google Scholar]
- 7.Koolhaas JM, Korte SM, De Boer SF, Van Der Vegt BJ, Van Reenen CG, Hopster H, De Jong IC, Ruis MAW, Blokhuis HJ. Coping styles in animals: current status in behavior and stress-physiology. Neuroscience & Biobehavioral Reviews. 1999;23:925–935. doi: 10.1016/s0149-7634(99)00026-3. [DOI] [PubMed] [Google Scholar]
- 8.Manuck SB, Kaplan JR, Clarkson TB. Behaviorally induced heart rate reactivity and atherosclerosis in cynomolgus monkeys. Psychosomatic medicine. 1983;45:95–108. doi: 10.1097/00006842-198305000-00002. [DOI] [PubMed] [Google Scholar]
- 9.Sgoifo A, de Boer SF, Westenbroek C, Maes FW, Beldhuis H, Suzuki T, Koolhaas JM. Incidence of arrhythmias and heart rate variability in wild-type rats exposed to social stress. Am J Physiol. 1997;273:H1754–60. doi: 10.1152/ajpheart.1997.273.4.H1754. [DOI] [PubMed] [Google Scholar]
- 10.Grant BF, Dawson DA, Stinson FS, Chou SP, Dufour MC, Pickering RP. The 12-month prevalence and trends in DSM-IV alcohol abuse and dependence: United States, 1991–1992 and 2001–2002. Drug and Alcohol Dependence. 2004;74:223–234. doi: 10.1016/j.drugalcdep.2004.02.004. [DOI] [PubMed] [Google Scholar]
- 11.Johnson LD, O’Malley PM, Bachman JG, Schulenberg JE. U.o. Michigan, editor. Monitoring the Future national results on adolescent drug use: Overview of key findings, 2011. Ann Arbor: Institute for Social Research, Ann Arbor, Michigan; 2011. [Google Scholar]
- 12.Kessler RC, Sonnega A, Bromet E, Hughes M, Nelson CB. Posttraumatic stress disorder in the National Comorbidity Survey. Arch Gen Psychiatry. 1995;52:1048–60. doi: 10.1001/archpsyc.1995.03950240066012. [DOI] [PubMed] [Google Scholar]
- 13.Tolin DF, Foa EB. Sex differences in trauma and posttraumatic stress disorder: a quantitative review of 25 years of research. Psychol Bull. 2006;132:959–92. doi: 10.1037/0033-2909.132.6.959. [DOI] [PubMed] [Google Scholar]
- 14.Breslau N. Gender differences in trauma and posttraumatic stress disorder. J Gend Specif Med. 2002;5:34–40. [PubMed] [Google Scholar]
- 15.Sheikh JI, Leskin GA, Klein DF. Gender differences in panic disorder: findings from the National Comorbidity Survey. Am J Psychiatry. 2002;159:55–8. doi: 10.1176/appi.ajp.159.1.55. [DOI] [PubMed] [Google Scholar]
- 16.Kendler KS, Kessler RC, Walters EE, MacLean C, Neale MC, Heath AC, Eaves LJ. Stressful life events, genetic liability, and onset of an episode of major depression in women. Am J Psychiatry. 1995;152:833–42. doi: 10.1176/ajp.152.6.833. [DOI] [PubMed] [Google Scholar]
- 17.Kessler RC, McGonagle KA, Swartz M, Blazer DG, Nelson CB. Sex and depression in the National Comorbidity Survey. I: Lifetime prevalence, chronicity and recurrence. J Affect Disord. 1993;29:85–96. doi: 10.1016/0165-0327(93)90026-g. [DOI] [PubMed] [Google Scholar]
- 18.Kessler RC. Epidemiology of women and depression. J Affect Disord. 2003;74:5–13. doi: 10.1016/s0165-0327(02)00426-3. [DOI] [PubMed] [Google Scholar]
- 19.Lydiard RB. Irritable bowel syndrome, anxiety, and depression: What are the links? Journal of Clinical Psychiatry. 2001;62:38–45. [PubMed] [Google Scholar]
- 20.Beghi E, Allais G, Cortelli P, D’Amico D, De Simone R, d’Onofrio F, Genco S, Manzoni GC, Moschiano F, Tonini MC, Torelli P, Quartaroli M, Roncolato M, Salvi S, Bussone G. Headache and anxiety-depressive disorder comorbidity: the HADAS study. Neurol Sci. 2007;28(Suppl 2):S217–9. doi: 10.1007/s10072-007-0780-6. [DOI] [PubMed] [Google Scholar]
- 21.van Mill JG, Hoogendijk WJ, Vogelzangs N, van Dyck R, Penninx BW. Insomnia and sleep duration in a large cohort of patients with major depressive disorder and anxiety disorders. The Journal of clinical psychiatry. 2010;71:239–46. doi: 10.4088/JCP.09m05218gry. [DOI] [PubMed] [Google Scholar]
- 22.Lipton RB, Stewart WF, Diamond S, Diamond ML, Reed M. Prevalence and burden of migraine in the United States: data from the American Migraine Study II. Headache. 2001;41:646–57. doi: 10.1046/j.1526-4610.2001.041007646.x. [DOI] [PubMed] [Google Scholar]
- 23.Singareddy R, Vgontzas AN, Fernandez-Mendoza J, Liao D, Calhoun S, Shaffer ML, Bixler EO. Risk factors for incident chronic insomnia: a general population prospective study. Sleep medicine. 2012;13:346–53. doi: 10.1016/j.sleep.2011.10.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Drossman DA, Thompson WG, Talley NJ, Funch-Jensen P, Janssens J, Whitehead WE. Identification of sub-groups of functional gastrointestinal disorders. Gastroenterology International. 1990;3:159–172. [Google Scholar]
- 25.Becker JB, Perry AN, Westenbroek C. Sex differences in the neural mechanisms mediating addiction: a new synthesis and hypothesis. Biol Sex Differ. 2012;3:14. doi: 10.1186/2042-6410-3-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Fattore L, Melis M, Fadda P, Fratta W. Sex differences in addictive disorders. Frontiers in Neuroendocrinology. 2014 doi: 10.1016/j.yfrne.2014.04.003. In press. [DOI] [PubMed] [Google Scholar]
- 27.Martin LA, Neighbors HW, Griffith DM. The Experience of Symptoms of Depression in Men vs Women: Analysis of the National Comorbidity Survey Replication. JAMA Psychiatry. 2013 doi: 10.1001/jamapsychiatry.2013.1985. [DOI] [PubMed] [Google Scholar]
- 28.Munck A, Guyre PM, Holbrook NJ. Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocrine reviews. 1984;5:25–44. doi: 10.1210/edrv-5-1-25. [DOI] [PubMed] [Google Scholar]
- 29.McEwen BS, Gianaros PJ. Stress- and allostasis-induced brain plasticity. Annual review of medicine. 2011;62:431–45. doi: 10.1146/annurev-med-052209-100430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.McEwen BS, Seeman T. Protective and damaging effects of mediators of stress. Elaborating and testing the concepts of allostasis and allostatic load. Annals of the New York Academy of Sciences. 1999;896:30–47. doi: 10.1111/j.1749-6632.1999.tb08103.x. [DOI] [PubMed] [Google Scholar]
- 31.Vale W, Spiess J, Rivier C, Rivier J. Characterization of a 41-residue ovine hypothalamic peptide that stimulates secretion of corticotropin and beta-endorphin. Science. 1981;213:1394–7. doi: 10.1126/science.6267699. [DOI] [PubMed] [Google Scholar]
- 32.Holmes MC, Antoni FA, Aguilera G, Catt KJ. Magnocellular axons in passage through the median eminence release vasopressin. Nature. 1986;319:326–9. doi: 10.1038/319326a0. [DOI] [PubMed] [Google Scholar]
- 33.Bush IE, Sandberg AA. Adrenocortical hormones in human plasma. The Journal of biological chemistry. 1953;205:783–93. [PubMed] [Google Scholar]
- 34.Eleftheriou BE. Bound and Free Corticosteroid in the Plasma of Two Subspecies of Deer Mice (Peromyscus Maniculatus) after Exposure to a Low Ambient Temperature. The Journal of endocrinology. 1964;31:75–80. doi: 10.1677/joe.0.0310075. [DOI] [PubMed] [Google Scholar]
- 35.De Kloet ER, Vreugdenhil E, Oitzl MS, Joels M. Brain corticosteroid receptor balance in health and disease. Endocrine reviews. 1998;19:269–301. doi: 10.1210/edrv.19.3.0331. [DOI] [PubMed] [Google Scholar]
- 36.De Kloet ER, Reul JM. Feedback action and tonic influence of corticosteroids on brain function: a concept arising from the heterogeneity of brain receptor systems. Psychoneuroendocrinology. 1987;12:83–105. doi: 10.1016/0306-4530(87)90040-0. [DOI] [PubMed] [Google Scholar]
- 37.Dallman MF, Akana SF, Jacobson L, Levin N, Cascio CS, Shinsako J. Characterization of corticosterone feedback regulation of ACTH secretion. Annals of the New York Academy of Sciences. 1987;512:402–14. doi: 10.1111/j.1749-6632.1987.tb24976.x. [DOI] [PubMed] [Google Scholar]
- 38.Tasker JG, Herman JP. Mechanisms of rapid glucocorticoid feedback inhibition of the hypothalamic-pituitary-adrenal axis. Stress. 2011;14:398–406. doi: 10.3109/10253890.2011.586446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Di S, Malcher-Lopes R, Halmos KC, Tasker JG. Nongenomic glucocorticoid inhibition via endocannabinoid release in the hypothalamus: a fast feedback mechanism. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2003;23:4850–7. doi: 10.1523/JNEUROSCI.23-12-04850.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Charron J, Drouin J. Glucocorticoid inhibition of transcription from episomal proopiomelanocortin gene promoter. Proceedings of the National Academy of Sciences of the United States of America. 1986;83:8903–7. doi: 10.1073/pnas.83.23.8903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Newton R. Molecular mechanisms of glucocorticoid action: what is important? Thorax. 2000;55:603–13. doi: 10.1136/thorax.55.7.603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Jingami H, Matsukura S, Numa S, Imura H. Effects of adrenalectomy and dexamethasone administration on the level of prepro-corticotropin-releasing factor messenger ribonucleic acid (mRNA) in the hypothalamus and adrenocorticotropin/beta-lipotropin precursor mRNA in the pituitary in rats. Endocrinology. 1985;117:1314–20. doi: 10.1210/endo-117-4-1314. [DOI] [PubMed] [Google Scholar]
- 43.Kageyama K, Suda T. Regulatory mechanisms underlying corticotropin-releasing factor gene expression in the hypothalamus. Endocrine journal. 2009;56:335–44. doi: 10.1507/endocrj.k09e-075. [DOI] [PubMed] [Google Scholar]
- 44.Malkoski SP, Dorin RI. Composite glucocorticoid regulation at a functionally defined negative glucocorticoid response element of the human corticotropin-releasing hormone gene. Molecular endocrinology. 1999;13:1629–44. doi: 10.1210/mend.13.10.0351. [DOI] [PubMed] [Google Scholar]
- 45.Pozzoli G, Bilezikjian LM, Perrin MH, Blount AL, Vale WW. Corticotropin-releasing factor (CRF) and glucocorticoids modulate the expression of type 1 CRF receptor messenger ribonucleic acid in rat anterior pituitary cell cultures. Endocrinology. 1996;137:65–71. doi: 10.1210/endo.137.1.8536643. [DOI] [PubMed] [Google Scholar]
- 46.Reul JM, de Kloet ER. Two receptor systems for corticosterone in rat brain: microdistribution and differential occupation. Endocrinology. 1985;117:2505–11. doi: 10.1210/endo-117-6-2505. [DOI] [PubMed] [Google Scholar]
- 47.Veldhuis HD, Van Koppen C, Van Ittersum M, De Kloet ER. Specificity of the adrenal steroid receptor system in rat hippocampus. Endocrinology. 1982;110:2044–51. doi: 10.1210/endo-110-6-2044. [DOI] [PubMed] [Google Scholar]
- 48.Sapolsky RM, Meaney MJ, McEwen BS. The development of the glucocorticoid receptor system in the rat limbic brain. III. Negative-feedback regulation. Brain research. 1985;350:169–73. doi: 10.1016/0165-3806(85)90261-5. [DOI] [PubMed] [Google Scholar]
- 49.McEwen BS. Protective and damaging effects of stress mediators. The New England journal of medicine. 1998;338:171–9. doi: 10.1056/NEJM199801153380307. [DOI] [PubMed] [Google Scholar]
- 50.Sorrells SF, Caso JR, Munhoz CD, Sapolsky RM. The stressed CNS: when glucocorticoids aggravate inflammation. Neuron. 2009;64:33–9. doi: 10.1016/j.neuron.2009.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yehuda R, Teicher MH, Trestman RL, Levengood RA, Siever LJ. Cortisol regulation in posttraumatic stress disorder and major depression: a chronobiological analysis. Biol Psychiatry. 1996;40:79–88. doi: 10.1016/0006-3223(95)00451-3. [DOI] [PubMed] [Google Scholar]
- 52.Yehuda R, Golier JA, Kaufman S. Circadian rhythm of salivary cortisol in Holocaust survivors with and without PTSD. Am J Psychiatry. 2005;162:998–1000. doi: 10.1176/appi.ajp.162.5.998. [DOI] [PubMed] [Google Scholar]
- 53.Elzinga BM, Schmahl CG, Vermetten E, van Dyck R, Bremner JD. Higher cortisol levels following exposure to traumatic reminders in abuse-related PTSD. Neuropsychopharmacology. 2003;28:1656–65. doi: 10.1038/sj.npp.1300226. [DOI] [PubMed] [Google Scholar]
- 54.Liberzon I, Abelson JL, Flagel SB, Raz J, Young EA. Neuroendocrine and psychophysiologic responses in PTSD: a symptom provocation study. Neuropsychopharmacology. 1999;21:40–50. doi: 10.1016/S0893-133X(98)00128-6. [DOI] [PubMed] [Google Scholar]
- 55.Holsboer F. Stress, hypercortisolism and corticosteroid receptors in depression: implications for therapy. Journal of affective disorders. 2001;62:77–91. doi: 10.1016/s0165-0327(00)00352-9. [DOI] [PubMed] [Google Scholar]
- 56.Deuschle M, Schweiger U, Weber B, Gotthardt U, Korner A, Schmider J, Standhardt H, Lammers CH, Heuser I. Diurnal activity and pulsatility of the hypothalamus-pituitary-adrenal system in male depressed patients and healthy controls. The Journal of clinical endocrinology and metabolism. 1997;82:234–8. doi: 10.1210/jcem.82.1.3689. [DOI] [PubMed] [Google Scholar]
- 57.Nestler EJ, Barrot M, DiLeone RJ, Eisch AJ, Gold SJ, Monteggia LM. Neurobiology of depression. Neuron. 2002;34:13–25. doi: 10.1016/s0896-6273(02)00653-0. [DOI] [PubMed] [Google Scholar]
- 58.Plotsky PM, Owens MJ, Nemeroff CB. Psychoneuroendocrinology of depression. Hypothalamic-pituitary-adrenal axis. Psychiatr Clin North Am. 1998;21:293–307. doi: 10.1016/s0193-953x(05)70006-x. [DOI] [PubMed] [Google Scholar]
- 59.Barden N. Implication of the hypothalamic-pituitary-adrenal axis in the physiopathology of depression. Journal of psychiatry & neuroscience: JPN. 2004;29:185–93. [PMC free article] [PubMed] [Google Scholar]
- 60.Olff M, de Vries GJ, Guzelcan Y, Assies J, Gersons BP. Changes in cortisol and DHEA plasma levels after psychotherapy for PTSD. Psychoneuroendocrinology. 2007;32:619–26. doi: 10.1016/j.psyneuen.2007.04.001. [DOI] [PubMed] [Google Scholar]
- 61.Uhart M, Chong RY, Oswald L, Lin PI, Wand GS. Gender differences in hypothalamic-pituitary-adrenal (HPA) axis reactivity. Psychoneuroendocrinology. 2006;31:642–52. doi: 10.1016/j.psyneuen.2006.02.003. [DOI] [PubMed] [Google Scholar]
- 62.Kirschbaum C, Kudielka BM, Gaab J, Schommer NC, Hellhammer DH. Impact of gender, menstrual cycle phase, and oral contraceptives on the activity of the hypothalamus-pituitary-adrenal axis. Psychosom Med. 1999;61:154–62. doi: 10.1097/00006842-199903000-00006. [DOI] [PubMed] [Google Scholar]
- 63.Seeman TE, Singer B, Wilkinson CW, McEwen B. Gender differences in age-related changes in HPA axis reactivity. Psychoneuroendocrinology. 2001;26:225–40. doi: 10.1016/s0306-4530(00)00043-3. [DOI] [PubMed] [Google Scholar]
- 64.Kudielka BM, Hellhammer J, Hellhammer DH, Wolf OT, Pirke KM, Varadi E, Pilz J, Kirschbaum C. Sex differences in endocrine and psychological responses to psychosocial stress in healthy elderly subjects and the impact of a 2-week dehydroepiandrosterone treatment. J Clin Endocrinol Metab. 1998;83:1756–61. doi: 10.1210/jcem.83.5.4758. [DOI] [PubMed] [Google Scholar]
- 65.Kudielka BM, Kirschbaum C. Sex differences in HPA axis responses to stress: a review. Biol Psychol. 2005;69:113–32. doi: 10.1016/j.biopsycho.2004.11.009. [DOI] [PubMed] [Google Scholar]
- 66.Collins A, Frankenhaeuser M. Stress responses in male and female engineering students. J Human Stress. 1978;4:43–8. doi: 10.1080/0097840X.1978.9934986. [DOI] [PubMed] [Google Scholar]
- 67.Meewisse ML, Reitsma JB, de Vries GJ, Gersons BP, Olff M. Cortisol and post-traumatic stress disorder in adults: systematic review and meta-analysis. The British journal of psychiatry: the journal of mental science. 2007;191:387–92. doi: 10.1192/bjp.bp.106.024877. [DOI] [PubMed] [Google Scholar]
- 68.Freidenberg BM, Gusmano R, Hickling EJ, Blanchard EB, Bremner JD, Frye C. Women with PTSD have lower basal salivary cortisol levels later in the day than do men with PTSD: a preliminary study. Physiology & behavior. 2010;99:234–6. doi: 10.1016/j.physbeh.2009.06.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Paris JJ, Franco C, Sodano R, Freidenberg B, Gordis E, Anderson DA, Forsyth JP, Wulfert E, Frye CA. Sex differences in salivary cortisol in response to acute stressors among healthy participants, in recreational or pathological gamblers, and in those with posttraumatic stress disorder. Hormones and behavior. 2010;57:35–45. doi: 10.1016/j.yhbeh.2009.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Klaassens ER, Giltay EJ, Cuijpers P, van Veen T, Zitman FG. Adulthood trauma and HPA-axis functioning in healthy subjects and PTSD patients: a meta-analysis. Psychoneuroendocrinology. 2012;37:317–31. doi: 10.1016/j.psyneuen.2011.07.003. [DOI] [PubMed] [Google Scholar]
- 71.Young EA, Altemus M. Puberty, ovarian steroids, and stress. Ann N Y Acad Sci. 2004;1021:124–33. doi: 10.1196/annals.1308.013. [DOI] [PubMed] [Google Scholar]
- 72.Young EA. Glucocorticoid cascade hypothesis revisited: role of gonadal steroids. Depression. 1995;3:20–27. [Google Scholar]
- 73.Peeters F, Nicholson NA, Berkhof J. Cortisol responses to daily events in major depressive disorder. Psychosomatic medicine. 2003;65:836–41. doi: 10.1097/01.psy.0000088594.17747.2e. [DOI] [PubMed] [Google Scholar]
- 74.Chopra KK, Ravindran A, Kennedy SH, Mackenzie B, Matthews S, Anisman H, Bagby RM, Farvolden P, Levitan RD. Sex differences in hormonal responses to a social stressor in chronic major depression. Psychoneuroendocrinology. 2009;34:1235–41. doi: 10.1016/j.psyneuen.2009.03.014. [DOI] [PubMed] [Google Scholar]
- 75.Reul JM, van den Bosch FR, de Kloet ER. Relative occupation of type-I and type-II corticosteroid receptors in rat brain following stress and dexamethasone treatment: functional implications. The Journal of endocrinology. 1987;115:459–67. doi: 10.1677/joe.0.1150459. [DOI] [PubMed] [Google Scholar]
- 76.Arana GW, Baldessarini RJ, Ornsteen M. The dexamethasone suppression test for diagnosis and prognosis in psychiatry. Commentary and review. Archives of general psychiatry. 1985;42:1193–204. doi: 10.1001/archpsyc.1985.01790350067012. [DOI] [PubMed] [Google Scholar]
- 77.Arana GW. Dexamethasone suppression test in the diagnosis of depression. JAMA: the journal of the American Medical Association. 1991;265:2253–4. [PubMed] [Google Scholar]
- 78.Carroll BJ, Feinberg M, Greden JF, Tarika J, Albala AA, Haskett RF, James NM, Kronfol Z, Lohr N, Steiner M, de Vigne JP, Young E. A specific laboratory test for the diagnosis of melancholia. Standardization, validation, and clinical utility. Archives of general psychiatry. 1981;38:15–22. doi: 10.1001/archpsyc.1981.01780260017001. [DOI] [PubMed] [Google Scholar]
- 79.Watson S, Gallagher P, Smith MS, Ferrier IN, Young AH. The dex/CRH test--is it better than the DST? Psychoneuroendocrinology. 2006;31:889–94. doi: 10.1016/j.psyneuen.2006.03.001. [DOI] [PubMed] [Google Scholar]
- 80.Heuser I, Yassouridis A, Holsboer F. The combined dexamethasone/CRH test: a refined laboratory test for psychiatric disorders. Journal of psychiatric research. 1994;28:341–56. doi: 10.1016/0022-3956(94)90017-5. [DOI] [PubMed] [Google Scholar]
- 81.Ising M, Kunzel HE, Binder EB, Nickel T, Modell S, Holsboer F. The combined dexamethasone/CRH test as a potential surrogate marker in depression. Progress in neuro-psychopharmacology & biological psychiatry. 2005;29:1085–93. doi: 10.1016/j.pnpbp.2005.03.014. [DOI] [PubMed] [Google Scholar]
- 82.Altemus M, Redwine L, Leong YM, Yoshikawa T, Yehuda R, Detera-Wadleigh S, Murphy DL. Reduced sensitivity to glucocorticoid feedback and reduced glucocorticoid receptor mRNA expression in the luteal phase of the menstrual cycle. Neuropsychopharmacology. 1997;17:100–9. doi: 10.1016/S0893-133X(97)00039-0. [DOI] [PubMed] [Google Scholar]
- 83.Lopez-Duran NL, Kovacs M, George CJ. Hypothalamic-pituitary-adrenal axis dysregulation in depressed children and adolescents: a meta-analysis. Psychoneuroendocrinology. 2009;34:1272–83. doi: 10.1016/j.psyneuen.2009.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Huizenga NA, Koper JW, de Lange P, Pols HA, Stolk RP, Grobbee DE, de Jong FH, Lamberts SW. Interperson variability but intraperson stability of baseline plasma cortisol concentrations, and its relation to feedback sensitivity of the hypothalamo-pituitary-adrenal axis to a low dose of dexamethasone in elderly individuals. J Clin Endocrinol Metab. 1998;83:47–54. doi: 10.1210/jcem.83.1.4498. [DOI] [PubMed] [Google Scholar]
- 85.Coppen A, Abou-Saleh M, Milln P, Metcalfe M, Harwood J, Bailey J. Dexamethasone suppression test in depression and other psychiatric illness. The British journal of psychiatry: the journal of mental science. 1983;142:498–504. doi: 10.1192/bjp.142.5.498. [DOI] [PubMed] [Google Scholar]
- 86.Kunugi H, Ida I, Owashi T, Kimura M, Inoue Y, Nakagawa S, Yabana T, Urushibara T, Kanai R, Aihara M, Yuuki N, Otsubo T, Oshima A, Kudo K, Inoue T, Kitaichi Y, Shirakawa O, Isogawa K, Nagayama H, Kamijima K, Nanko S, Kanba S, Higuchi T, Mikuni M. Assessment of the dexamethasone/CRH test as a state-dependent marker for hypothalamic-pituitary-adrenal (HPA) axis abnormalities in major depressive episode: a Multicenter Study. Neuropsychopharmacology. 2006;31:212–20. doi: 10.1038/sj.npp.1300868. [DOI] [PubMed] [Google Scholar]
- 87.Heuser IJ, Gotthardt U, Schweiger U, Schmider J, Lammers CH, Dettling M, Holsboer F. Age-associated changes of pituitary-adrenocortical hormone regulation in humans: importance of gender. Neurobiol Aging. 1994;15:227–31. doi: 10.1016/0197-4580(94)90117-1. [DOI] [PubMed] [Google Scholar]
- 88.Gallucci WT, Baum A, Laue L, Rabin DS, Chrousos GP, Gold PW, Kling MA. Sex differences in sensitivity of the hypothalamic-pituitary-adrenal axis. Health Psychol. 1993;12:420–5. doi: 10.1037//0278-6133.12.5.420. [DOI] [PubMed] [Google Scholar]
- 89.Stroud LR, Papandonatos GD, Williamson DE, Dahl RE. Sex differences in cortisol response to corticotropin releasing hormone challenge over puberty: Pittsburgh Pediatric Neurobehavioral Studies. Psychoneuroendocrinology. 2011;36:1226–38. doi: 10.1016/j.psyneuen.2011.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Gold PW, Chrousos GP. Clinical studies with corticotropin releasing factor: implications for the diagnosis and pathophysiology of depression, Cushing’s disease, and adrenal insufficiency. Psychoneuroendocrinology. 1985;10:401–19. doi: 10.1016/0306-4530(85)90080-0. [DOI] [PubMed] [Google Scholar]
- 91.Gold PW, Chrousos GP. Organization of the stress system and its dysregulation in melancholic and atypical depression: high vs low CRH/NE states. Mol Psychiatry. 2002;7:254–75. doi: 10.1038/sj.mp.4001032. [DOI] [PubMed] [Google Scholar]
- 92.Bremner JD, Licinio J, Darnell A, Krystal JH, Owens MJ, Southwick SM, Nemeroff CB, Charney DS. Elevated CSF corticotropin-releasing factor concentrations in posttraumatic stress disorder. Am J Psychiatry. 1997;154:624–9. doi: 10.1176/ajp.154.5.624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Nemeroff CB, Widerlov E, Bissette G, Walleus H, Karlsson I, Eklund K, Kilts CD, Loosen PT, Vale W. Elevated concentrations of CSF corticotropin-releasing factor-like immunoreactivity in depressed patients. Science. 1984;226:1342–4. doi: 10.1126/science.6334362. [DOI] [PubMed] [Google Scholar]
- 94.Baker DG, West SA, Nicholson WE, Ekhator NN, Kasckow JW, Hill KK, Bruce AB, Orth DN, Geracioti TD., Jr Serial CSF corticotropin-releasing hormone levels and adrenocortical activity in combat veterans with posttraumatic stress disorder. Am J Psychiatry. 1999;156:585–8. doi: 10.1176/ajp.156.4.585. [DOI] [PubMed] [Google Scholar]
- 95.Banki CM, Karmacsi L, Bissette G, Nemeroff CB. CSF corticotropin-releasing hormone and somatostatin in major depression: response to antidepressant treatment and relapse. Eur Neuropsychopharmacol. 1992;2:107–13. doi: 10.1016/0924-977x(92)90019-5. [DOI] [PubMed] [Google Scholar]
- 96.Wang SS, Kamphuis W, Huitinga I, Zhou JN, Swaab DF. Gene expression analysis in the human hypothalamus in depression by laser microdissection and real-time PCR: the presence of multiple receptor imbalances. Mol Psychiatry. 2008;13:786–99. 741. doi: 10.1038/mp.2008.38. [DOI] [PubMed] [Google Scholar]
- 97.Raadsheer FC, Hoogendijk WJ, Stam FC, Tilders FJ, Swaab DF. Increased numbers of corticotropin-releasing hormone expressing neurons in the hypothalamic paraventricular nucleus of depressed patients. Neuroendocrinology. 1994;60:436–44. doi: 10.1159/000126778. [DOI] [PubMed] [Google Scholar]
- 98.Austin MC, Janosky JE, Murphy HA. Increased corticotropin-releasing hormone immunoreactivity in monoamine-containing pontine nuclei of depressed suicide men. Mol Psychiatry. 2003;8:324–32. doi: 10.1038/sj.mp.4001250. [DOI] [PubMed] [Google Scholar]
- 99.Bissette G, Klimek V, Pan J, Stockmeier C, Ordway G. Elevated concentrations of CRF in the locus coeruleus of depressed subjects. Neuropsychopharmacology. 2003;28:1328–35. doi: 10.1038/sj.npp.1300191. [DOI] [PubMed] [Google Scholar]
- 100.Hauger RL, Risbrough V, Oakley RH, Olivares-Reyes JA, Dautzenberg FM. Role of CRF receptor signaling in stress vulnerability, anxiety, and depression. Ann N Y Acad Sci. 2009;1179:120–43. doi: 10.1111/j.1749-6632.2009.05011.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Heinrichs SC, Lapsansky J, Lovenberg TW, De Souza EB, Chalmers DT. Corticotropin-releasing factor CRF1, but not CRF2, receptors mediate anxiogenic-like behavior. Regulatory peptides. 1997;71:15–21. doi: 10.1016/s0167-0115(97)01005-7. [DOI] [PubMed] [Google Scholar]
- 102.Smith GW, Aubry JM, Dellu F, Contarino A, Bilezikjian LM, Gold LH, Chen R, Marchuk Y, Hauser C, Bentley CA, Sawchenko PE, Koob GF, Vale W, Lee KF. Corticotropin releasing factor receptor 1-deficient mice display decreased anxiety, impaired stress response, and aberrant neuroendocrine development. Neuron. 1998;20:1093–102. doi: 10.1016/s0896-6273(00)80491-2. [DOI] [PubMed] [Google Scholar]
- 103.Amstadter AB, Nugent NR, Yang BZ, Miller A, Siburian R, Moorjani P, Haddad S, Basu A, Fagerness J, Saxe G, Smoller JW, Koenen KC. Corticotrophin-releasing hormone type 1 receptor gene (CRHR1) variants predict posttraumatic stress disorder onset and course in pediatric injury patients. Dis Markers. 2011;30:89–99. doi: 10.3233/DMA-2011-0761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Wasserman D, Sokolowski M, Rozanov V, Wasserman J. The CRHR1 gene: a marker for suicidality in depressed males exposed to low stress. Genes, brain, and behavior. 2008;7:14–9. doi: 10.1111/j.1601-183X.2007.00310.x. [DOI] [PubMed] [Google Scholar]
- 105.Liu Z, Zhu F, Wang G, Xiao Z, Wang H, Tang J, Wang X, Qiu D, Liu W, Cao Z, Li W. Association of corticotropin-releasing hormone receptor1 gene SNP and haplotype with major depression. Neuroscience letters. 2006;404:358–62. doi: 10.1016/j.neulet.2006.06.016. [DOI] [PubMed] [Google Scholar]
- 106.Polanczyk G, Caspi A, Williams B, Price TS, Danese A, Sugden K, Uher R, Poulton R, Moffitt TE. Protective effect of CRHR1 gene variants on the development of adult depression following childhood maltreatment: replication and extension. Archives of general psychiatry. 2009;66:978–85. doi: 10.1001/archgenpsychiatry.2009.114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Bao AM, Swaab DF. Gender difference in age-related number of corticotropin-releasing hormone-expressing neurons in the human hypothalamic paraventricular nucleus and the role of sex hormones. Neuroendocrinology. 2007;85:27–36. doi: 10.1159/000099832. [DOI] [PubMed] [Google Scholar]
- 108.Kitay JI. Sex differences in adrenal cortical secretion in the rat. Endocrinology. 1961;68:818–24. doi: 10.1210/endo-68-5-818. [DOI] [PubMed] [Google Scholar]
- 109.Viau V, Bingham B, Davis J, Lee P, Wong M. Gender and puberty interact on the stress-induced activation of parvocellular neurosecretory neurons and corticotropin-releasing hormone messenger ribonucleic acid expression in the rat. Endocrinology. 2005;146:137–46. doi: 10.1210/en.2004-0846. [DOI] [PubMed] [Google Scholar]
- 110.Weinstock M, Razin M, Schorer-Apelbaum D, Men D, McCarty R. Gender differences in sympathoadrenal activity in rats at rest and in response to footshock stress. Int J Dev Neurosci. 1998;16:289–95. doi: 10.1016/s0736-5748(98)00021-5. [DOI] [PubMed] [Google Scholar]
- 111.Critchlow V, Liebelt RA, Bar-Sela M, Mountcastle W, Lipscomb HS. Sex difference in resting pituitary-adrenal function in the rat. Am J Physiol. 1963;205:807–15. doi: 10.1152/ajplegacy.1963.205.5.807. [DOI] [PubMed] [Google Scholar]
- 112.Handa RJ, Burgess LH, Kerr JE, O’Keefe JA. Gonadal steroid hormone receptors and sex differences in the hypothalamo-pituitary-adrenal axis. Horm Behav. 1994;28:464–76. doi: 10.1006/hbeh.1994.1044. [DOI] [PubMed] [Google Scholar]
- 113.Rivier C. Gender, sex steroids, corticotropin-releasing factor, nitric oxide, and the HPA response to stress. Pharmacol Biochem Behav. 1999;64:739–51. doi: 10.1016/s0091-3057(99)00148-3. [DOI] [PubMed] [Google Scholar]
- 114.Iwasaki-Sekino A, Mano-Otagiri A, Ohata H, Yamauchi N, Shibasaki T. Gender differences in corticotropin and corticosterone secretion and corticotropin-releasing factor mRNA expression in the paraventricular nucleus of the hypothalamus and the central nucleus of the amygdala in response to footshock stress or psychological stress in rats. Psychoneuroendocrinology. 2009;34:226–37. doi: 10.1016/j.psyneuen.2008.09.003. [DOI] [PubMed] [Google Scholar]
- 115.Heinsbroek RP, Van Haaren F, Feenstra MG, Endert E, Van de Poll NE. Sex- and time-dependent changes in neurochemical and hormonal variables induced by predictable and unpredictable footshock. Physiol Behav. 1991;49:1251–6. doi: 10.1016/0031-9384(91)90359-v. [DOI] [PubMed] [Google Scholar]
- 116.Seale JV, Wood SA, Atkinson HC, Harbuz MS, Lightman SL. Gonadal steroid replacement reverses gonadectomy-induced changes in the corticosterone pulse profile and stress-induced hypothalamic-pituitary-adrenal axis activity of male and female rats. J Neuroendocrinol. 2004;16:989–98. doi: 10.1111/j.1365-2826.2004.01258.x. [DOI] [PubMed] [Google Scholar]
- 117.Weathington JM, Arnold AR, Cooke BM. Juvenile social subjugation induces a sex-specific pattern of anxiety and depression-like behaviors in adult rats. Hormones and behavior. 2012;61:91–9. doi: 10.1016/j.yhbeh.2011.10.008. [DOI] [PubMed] [Google Scholar]
- 118.Carey MP, Deterd CH, de Koning J, Helmerhorst F, de Kloet ER. The influence of ovarian steroids on hypothalamic-pituitary-adrenal regulation in the female rat. The Journal of endocrinology. 1995;144:311–21. doi: 10.1677/joe.0.1440311. [DOI] [PubMed] [Google Scholar]
- 119.Viau V, Meaney MJ. Variations in the hypothalamic-pituitary-adrenal response to stress during the estrous cycle in the rat. Endocrinology. 1991;129:2503–11. doi: 10.1210/endo-129-5-2503. [DOI] [PubMed] [Google Scholar]
- 120.Atkinson HC, Waddell BJ. Circadian variation in basal plasma corticosterone and adrenocorticotropin in the rat: sexual dimorphism and changes across the estrous cycle. Endocrinology. 1997;138:3842–8. doi: 10.1210/endo.138.9.5395. [DOI] [PubMed] [Google Scholar]
- 121.Figueiredo HF, Ulrich-Lai YM, Choi DC, Herman JP. Estrogen potentiates adrenocortical responses to stress in female rats. Am J Physiol Endocrinol Metab. 2007;292:E1173–82. doi: 10.1152/ajpendo.00102.2006. [DOI] [PubMed] [Google Scholar]
- 122.Duncko R, Kiss A, Skultetyova I, Rusnak M, Jezova D. Corticotropin-releasing hormone mRNA levels in response to chronic mild stress rise in male but not in female rats while tyrosine hydroxylase mRNA levels decrease in both sexes. Psychoneuroendocrinology. 2001;26:77–89. doi: 10.1016/s0306-4530(00)00040-8. [DOI] [PubMed] [Google Scholar]
- 123.Sterrenburg L, Gaszner B, Boerrigter J, Santbergen L, Bramini M, Roubos EW, Peeters BW, Kozicz T. Sex-dependent and differential responses to acute restraint stress of corticotropin-releasing factor-producing neurons in the rat paraventricular nucleus, central amygdala, and bed nucleus of the stria terminalis. Journal of neuroscience research. 2012;90:179–92. doi: 10.1002/jnr.22737. [DOI] [PubMed] [Google Scholar]
- 124.Bohler HC, Jr, Zoeller RT, King JC, Rubin BS, Weber R, Merriam GR. Corticotropin releasing hormone mRNA is elevated on the afternoon of proestrus in the parvocellular paraventricular nuclei of the female rat. Brain Res Mol Brain Res. 1990;8:259–62. doi: 10.1016/0169-328x(90)90025-9. [DOI] [PubMed] [Google Scholar]
- 125.Lund TD, Munson DJ, Haldy ME, Handa RJ. Androgen inhibits, while oestrogen enhances, restraint-induced activation of neuropeptide neurones in the paraventricular nucleus of the hypothalamus. J Neuroendocrinol. 2004;16:272–8. doi: 10.1111/j.0953-8194.2004.01167.x. [DOI] [PubMed] [Google Scholar]
- 126.Bingaman EW, Magnuson DJ, Gray TS, Handa RJ. Androgen inhibits the increases in hypothalamic corticotropin-releasing hormone (CRH) and CRH-immunoreactivity following gonadectomy. Neuroendocrinology. 1994;59:228–34. doi: 10.1159/000126663. [DOI] [PubMed] [Google Scholar]
- 127.Lunga P, Herbert J. 17Beta-oestradiol modulates glucocorticoid, neural and behavioural adaptations to repeated restraint stress in female rats. J Neuroendocrinol. 2004;16:776–85. doi: 10.1111/j.1365-2826.2004.01234.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Green PG, Dahlqvist SR, Isenberg WM, Strausbaugh HJ, Miao FJ, Levine JD. Sex steroid regulation of the inflammatory response: sympathoadrenal dependence in the female rat. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1999;19:4082–9. doi: 10.1523/JNEUROSCI.19-10-04082.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Cutler GB, Jr, Barnes KM, Sauer MA, Loriaux DL. Estrogen receptor in rat adrenal gland. Endocrinology. 1978;102:252–7. doi: 10.1210/endo-102-1-252. [DOI] [PubMed] [Google Scholar]
- 130.Bentvelsen FM, McPhaul MJ, Wilson CM, Wilson JD, George FW. Regulation of immunoreactive androgen receptor in the adrenal gland of the adult rat. Endocrinology. 1996;137:2659–63. doi: 10.1210/endo.137.7.8770883. [DOI] [PubMed] [Google Scholar]
- 131.Shughrue PJ, Lane MV, Scrimo PJ, Merchenthaler I. Comparative distribution of estrogen receptor-α (ER-α) and β (ER-β) mRNA in the rat pituitary, gonad, and reproductive tract. Steroids. 1998;63:498–504. doi: 10.1016/s0039-128x(98)00054-3. [DOI] [PubMed] [Google Scholar]
- 132.Sar M, Lubahn DB, French FS, Wilson EM. Immunohistochemical localization of the androgen receptor in rat and human tissues. Endocrinology. 1990;127:3180–6. doi: 10.1210/endo-127-6-3180. [DOI] [PubMed] [Google Scholar]
- 133.Maejima Y, Aoyama M, Ookawara S, Hirao A, Sugita S. Distribution of the androgen receptor in the diencephalon and the pituitary gland in goats: Co-localisation with corticotrophin releasing hormone, arginine vasopressin and corticotrophs. Veterinary journal. 2009;181:193–9. doi: 10.1016/j.tvjl.2008.02.021. [DOI] [PubMed] [Google Scholar]
- 134.Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogen receptor-alpha and -beta mRNA in the rat central nervous system. The Journal of comparative neurology. 1997;388:507–25. doi: 10.1002/(sici)1096-9861(19971201)388:4<507::aid-cne1>3.0.co;2-6. [DOI] [PubMed] [Google Scholar]
- 135.Simerly RB, Chang C, Muramatsu M, Swanson LW. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: an in situ hybridization study. The Journal of comparative neurology. 1990;294:76–95. doi: 10.1002/cne.902940107. [DOI] [PubMed] [Google Scholar]
- 136.Vamvakopoulos NC, Chrousos GP. Evidence of direct estrogenic regulation of human corticotropin-releasing hormone gene expression. Potential implications for the sexual dimophism of the stress response and immune/inflammatory reaction. J Clin Invest. 1993;92:1896–902. doi: 10.1172/JCI116782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Shapiro RA, Xu C, Dorsa DM. Differential transcriptional regulation of rat vasopressin gene expression by estrogen receptor alpha and beta. Endocrinology. 2000;141:4056–64. doi: 10.1210/endo.141.11.7796. [DOI] [PubMed] [Google Scholar]
- 138.Bao AM, Fischer DF, Wu YH, Hol EM, Balesar R, Unmehopa UA, Zhou JN, Swaab DF. A direct androgenic involvement in the expression of human corticotropin-releasing hormone. Mol Psychiatry. 2006;11:567–76. doi: 10.1038/sj.mp.4001800. [DOI] [PubMed] [Google Scholar]
- 139.Burgess LH, Handa RJ. Chronic estrogen-induced alterations in adrenocorticotropin and corticosterone secretion, and glucocorticoid receptor-mediated functions in female rats. Endocrinology. 1992;131:1261–9. doi: 10.1210/endo.131.3.1324155. [DOI] [PubMed] [Google Scholar]
- 140.Weiser MJ, Handa RJ. Estrogen impairs glucocorticoid dependent negative feedback on the hypothalamic-pituitary-adrenal axis via estrogen receptor alpha within the hypothalamus. Neuroscience. 2009;159:883–95. doi: 10.1016/j.neuroscience.2008.12.058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Herman JP, McKlveen JM, Solomon MB, Carvalho-Netto E, Myers B. Neural regulation of the stress response: glucocorticoid feedback mechanisms. Brazilian journal of medical and biological research = Revista brasileira de pesquisas medicas e biologicas/Sociedade Brasileira de Biofisica … [et al.] 2012;45:292–8. doi: 10.1590/S0100-879X2012007500041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Turner BB. Sex difference in glucocorticoid binding in rat pituitary is estrogen dependent. Life sciences. 1990;46:1399–406. doi: 10.1016/0024-3205(90)90340-w. [DOI] [PubMed] [Google Scholar]
- 143.Turner BB, Weaver DA. Sexual dimorphism of glucocorticoid binding in rat brain. Brain research. 1985;343:16–23. doi: 10.1016/0006-8993(85)91153-9. [DOI] [PubMed] [Google Scholar]
- 144.Burgess LH, Handa RJ. Estrogen-induced alterations in the regulation of mineralocorticoid and glucocorticoid receptor messenger RNA expression in the female rat anterior pituitary gland and brain. Molecular and cellular neurosciences. 1993;4:191–8. doi: 10.1006/mcne.1993.1023. [DOI] [PubMed] [Google Scholar]
- 145.Bourke CH, Raees MQ, Malviya S, Bradburn CA, Binder EB, Neigh GN. Glucocorticoid sensitizers Bag1 and Ppid are regulated by adolescent stress in a sex-dependent manner. Psychoneuroendocrinology. 2013;38:84–93. doi: 10.1016/j.psyneuen.2012.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Steiniger B, Kniess T, Bergmann R, Pietzsch J, Wuest FR. Radiolabeled glucocorticoids as molecular probes for imaging brain glucocorticoid receptors by means of positron emission tomography (PET) Mini Rev Med Chem. 2008;8:728–39. doi: 10.2174/138955708784567403. [DOI] [PubMed] [Google Scholar]
- 147.Tooby J, Cosmides L. The Past Explains the Present Emotional Adaptations and the Structure of Ancestral Environments. Ethology and Sociobiology. 1990;11:375–424. [Google Scholar]
- 148.Moons WG, Eisenberger NI, Taylor SE. Anger and fear responses to stress have different biological profiles. Brain, behavior, and immunity. 2010;24:215–9. doi: 10.1016/j.bbi.2009.08.009. [DOI] [PubMed] [Google Scholar]
- 149.Cole PM, Michel MK, Teti LO. The development of emotion regulation and dysregulation: a clinical perspective. Monogr Soc Res Child Dev. 1994;59:73–100. [PubMed] [Google Scholar]
- 150.Critchley H. Emotion and its disorders. Br Med Bull. 2003;65:35–47. doi: 10.1093/bmb/65.1.35. [DOI] [PubMed] [Google Scholar]
- 151.Grossman M, Wood W. Sex differences in intensity of emotional experience: a social role interpretation. J Pers Soc Psychol. 1993;65:1010–22. doi: 10.1037//0022-3514.65.5.1010. [DOI] [PubMed] [Google Scholar]
- 152.Bradley MM, Codispoti M, Sabatinelli D, Lang PJ. Emotion and motivation II: sex differences in picture processing. Emotion. 2001;1:300–19. [PubMed] [Google Scholar]
- 153.Sharp C, van Goozen S, Goodyer I. Children’s subjective emotional reactivity to affective pictures: gender differences and their antisocial correlates in an unselected sample of 7–11-year-olds. J Child Psychol Psychiatry. 2006;47:143–50. doi: 10.1111/j.1469-7610.2005.01464.x. [DOI] [PubMed] [Google Scholar]
- 154.Fujita F, Diener E, Sandvik E. Gender differences in negative affect and well-being: the case for emotional intensity. J Pers Soc Psychol. 1991;61:427–34. doi: 10.1037//0022-3514.61.3.427. [DOI] [PubMed] [Google Scholar]
- 155.Costa PT, Jr, Zonderman AB, McCrae RR, Cornoni-Huntley J, Locke BZ, Barbano HE. Longitudinal analyses of psychological well-being in a national sample: stability of mean levels. J Gerontol. 1987;42:50–5. doi: 10.1093/geronj/42.1.50. [DOI] [PubMed] [Google Scholar]
- 156.Kelly MM, Tyrka AR, Price LH, Carpenter LL. Sex differences in the use of coping strategies: predictors of anxiety and depressive symptoms. Depress Anxiety. 2008;25:839–46. doi: 10.1002/da.20341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Thomsen DK, Mehlsen MY, Viidik A, Sommerlund B, Zachariae R. Age and gender differences in negative affect—Is there a role for emotion regulation? Personality and Individual Differences. 2005;38:1935–1946. [Google Scholar]
- 158.Leach LS, Christensen H, Mackinnon AJ, Windsor TD, Butterworth P. Gender differences in depression and anxiety across the adult lifespan: the role of psychosocial mediators. Soc Psychiatry Psychiatr Epidemiol. 2008;43:983–98. doi: 10.1007/s00127-008-0388-z. [DOI] [PubMed] [Google Scholar]
- 159.Kraft JM, Werner JS. Aging and the saturation of colors. 1. Colorimetric purity discrimination. J Opt Soc Am A Opt Image Sci Vis. 1999;16:223–30. doi: 10.1364/josaa.16.000223. [DOI] [PubMed] [Google Scholar]
- 160.Phan KL, Wager TD, Taylor SF, Liberzon I. Functional neuroimaging studies of human emotions. CNS Spectr. 2004;9:258–66. doi: 10.1017/s1092852900009196. [DOI] [PubMed] [Google Scholar]
- 161.Sabatinelli D, Fortune EE, Li Q, Siddiqui A, Krafft C, Oliver WT, Beck S, Jeffries J. Emotional perception: meta-analyses of face and natural scene processing. Neuroimage. 2011;54:2524–33. doi: 10.1016/j.neuroimage.2010.10.011. [DOI] [PubMed] [Google Scholar]
- 162.Goosens KA. Hippocampal regulation of aversive memories. Curr Opin Neurobiol. 2011;21:460–6. doi: 10.1016/j.conb.2011.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Domes G, Schulze L, Bottger M, Grossmann A, Hauenstein K, Wirtz PH, Heinrichs M, Herpertz SC. The neural correlates of sex differences in emotional reactivity and emotion regulation. Human brain mapping. 2010;31:758–69. doi: 10.1002/hbm.20903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Lebron-Milad K, Abbs B, Milad MR, Linnman C, Rougemount-Bucking A, Zeidan MA, Holt DJ, Goldstein JM. Sex differences in the neurobiology of fear conditioning and extinction: a preliminary fMRI study of shared sex differences with stress-arousal circuitry. Biol Mood Anxiety Disord. 2012;2:7. doi: 10.1186/2045-5380-2-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Stevens JS, Hamann S. Sex differences in brain activation to emotional stimuli: a meta-analysis of neuroimaging studies. Neuropsychologia. 2012;50:1578–93. doi: 10.1016/j.neuropsychologia.2012.03.011. [DOI] [PubMed] [Google Scholar]
- 166.Bellace M, Williams JM, Mohamed FB, Faro SH. An FMRI study of the activation of the hippocampus by emotional memory. Int J Neurosci. 2013;123:121–7. doi: 10.3109/00207454.2012.742894. [DOI] [PubMed] [Google Scholar]
- 167.Hofer A, Siedentopf CM, Ischebeck A, Rettenbacher MA, Verius M, Felber S, Wolfgang Fleischhacker W. Sex differences in brain activation patterns during processing of positively and negatively valenced emotional words. Psychol Med. 2007;37:109–19. doi: 10.1017/S0033291706008919. [DOI] [PubMed] [Google Scholar]
- 168.Cahill L. Sex-related influences on the neurobiology of emotionally influenced memory. Annals of the New York Academy of Sciences. 2003;985:163–73. doi: 10.1111/j.1749-6632.2003.tb07080.x. [DOI] [PubMed] [Google Scholar]
- 169.Cahill L, Haier RJ, White NS, Fallon J, Kilpatrick L, Lawrence C, Potkin SG, Alkire MT. Sex-related difference in amygdala activity during emotionally influenced memory storage. Neurobiology of learning and memory. 2001;75:1–9. doi: 10.1006/nlme.2000.3999. [DOI] [PubMed] [Google Scholar]
- 170.Cahill L, Uncapher M, Kilpatrick L, Alkire MT, Turner J. Sex-related hemispheric lateralization of amygdala function in emotionally influenced memory: an FMRI investigation. Learning & memory. 2004;11:261–6. doi: 10.1101/lm.70504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Canli T, Desmond JE, Zhao Z, Gabrieli JD. Sex differences in the neural basis of emotional memories. Proceedings of the National Academy of Sciences of the United States of America. 2002;99:10789–94. doi: 10.1073/pnas.162356599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.MacQueen GM, Campbell S, McEwen BS, Macdonald K, Amano S, Joffe RT, Nahmias C, Young LT. Course of illness, hippocampal function, and hippocampal volume in major depression. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:1387–92. doi: 10.1073/pnas.0337481100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Sheline YI. 3D MRI studies of neuroanatomic changes in unipolar major depression: the role of stress and medical comorbidity. Biol Psychiatry. 2000;48:791–800. doi: 10.1016/s0006-3223(00)00994-x. [DOI] [PubMed] [Google Scholar]
- 174.Sheline YI, Wang PW, Gado MH, Csernansky JG, Vannier MW. Hippocampal atrophy in recurrent major depression. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:3908–13. doi: 10.1073/pnas.93.9.3908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.McKinnon MC, Yucel K, Nazarov A, MacQueen GM. A meta-analysis examining clinical predictors of hippocampal volume in patients with major depressive disorder. Journal of psychiatry & neuroscience: JPN. 2009;34:41–54. [PMC free article] [PubMed] [Google Scholar]
- 176.Vakili K, Pillay SS, Lafer B, Fava M, Renshaw PF, Bonello-Cintron CM, Yurgelun-Todd DA. Hippocampal volume in primary unipolar major depression: a magnetic resonance imaging study. Biol Psychiatry. 2000;47:1087–90. doi: 10.1016/s0006-3223(99)00296-6. [DOI] [PubMed] [Google Scholar]
- 177.Kong L, Chen K, Womer F, Jiang W, Luo X, Driesen N, Liu J, Blumberg H, Tang Y, Xu K, Wang F. Sex differences of gray matter morphology in cortico-limbic-striatal neural system in major depressive disorder. Journal of psychiatric research. 2013;47:733–9. doi: 10.1016/j.jpsychires.2013.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Kitayama N, Vaccarino V, Kutner M, Weiss P, Bremner JD. Magnetic resonance imaging (MRI) measurement of hippocampal volume in posttraumatic stress disorder: A meta-analysis. Journal of Affective Disorders. 2005;88:79–86. doi: 10.1016/j.jad.2005.05.014. [DOI] [PubMed] [Google Scholar]
- 179.Karl A, Schaefer M, Malta LS, Dörfel D, Rohleder N, Werner A. A meta-analysis of structural brain abnormalities in PTSD. Neuroscience & Biobehavioral Reviews. 2006;30:1004–1031. doi: 10.1016/j.neubiorev.2006.03.004. [DOI] [PubMed] [Google Scholar]
- 180.Gilbertson MW, Shenton ME, Ciszewski A, Kasai K, Lasko NB, Orr SP, Pitman RK. Smaller hippocampal volume predicts pathologic vulnerability to psychological trauma. Nature neuroscience. 2002;5:1242–7. doi: 10.1038/nn958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Kasai K, Yamasue H, Gilbertson MW, Shenton ME, Rauch SL, Pitman RK. Evidence for acquired pregenual anterior cingulate gray matter loss from a twin study of combat-related posttraumatic stress disorder. Biol Psychiatry. 2008;63:550–6. doi: 10.1016/j.biopsych.2007.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Christoffel DJ, Golden SA, Russo SJ. Structural and synaptic plasticity in stress-related disorders. Rev Neurosci. 2011;22:535–49. doi: 10.1515/RNS.2011.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Leuner B, Shors TJ. Stress, anxiety, and dendritic spines: What are the connections? Neuroscience. 2012 doi: 10.1016/j.neuroscience.2012.04.021. [DOI] [PubMed] [Google Scholar]
- 184.Rochefort NL, Konnerth A. Dendritic spines: from structure to in vivo function. EMBO Rep. 2012;13:699–708. doi: 10.1038/embor.2012.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Kasai H, Hayama T, Ishikawa M, Watanabe S, Yagishita S, Noguchi J. Learning rules and persistence of dendritic spines. The European journal of neuroscience. 2010;32:241–9. doi: 10.1111/j.1460-9568.2010.07344.x. [DOI] [PubMed] [Google Scholar]
- 186.Xu T, Yu X, Perlik AJ, Tobin WF, Zweig JA, Tennant K, Jones T, Zuo Y. Rapid formation and selective stabilization of synapses for enduring motor memories. Nature. 2009;462:915–9. doi: 10.1038/nature08389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Lang C, Barco A, Zablow L, Kandel ER, Siegelbaum SA, Zakharenko SS. Transient expansion of synaptically connected dendritic spines upon induction of hippocampal long-term potentiation. Proceedings of the National Academy of Sciences of the United States of America. 2004;101:16665–70. doi: 10.1073/pnas.0407581101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Wong RO, Ghosh A. Activity-dependent regulation of dendritic growth and patterning. Nature reviews. Neuroscience. 2002;3:803–12. doi: 10.1038/nrn941. [DOI] [PubMed] [Google Scholar]
- 189.Yang G, Pan F, Gan WB. Stably maintained dendritic spines are associated with lifelong memories. Nature. 2009;462:920–4. doi: 10.1038/nature08577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Vyas A, Pillai AG, Chattarji S. Recovery after chronic stress fails to reverse amygdaloid neuronal hypertrophy and enhanced anxiety-like behavior. Neuroscience. 2004;128:667–73. doi: 10.1016/j.neuroscience.2004.07.013. [DOI] [PubMed] [Google Scholar]
- 191.McEwen BS. Stress, sex, and neural adaptation to a changing environment: mechanisms of neuronal remodeling. Annals of the New York Academy of Sciences. 2010;1204(Suppl):E38–59. doi: 10.1111/j.1749-6632.2010.05568.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Shors TJ, Chua C, Falduto J. Sex differences and opposite effects of stress on dendritic spine density in the male versus female hippocampus. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2001;21:6292–7. doi: 10.1523/JNEUROSCI.21-16-06292.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Garrett JE, Wellman CL. Chronic stress effects on dendritic morphology in medial prefrontal cortex: sex differences and estrogen dependence. Neuroscience. 2009;162:195–207. doi: 10.1016/j.neuroscience.2009.04.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Woolley CS, Gould E, Frankfurt M, McEwen BS. Naturally occurring fluctuation in dendritic spine density on adult hippocampal pyramidal neurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1990;10:4035–9. doi: 10.1523/JNEUROSCI.10-12-04035.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Gould E, Woolley CS, Frankfurt M, McEwen BS. Gonadal steroids regulate dendritic spine density in hippocampal pyramidal cells in adulthood. The Journal of neuroscience: the official journal of the Society for Neuroscience. 1990;10:1286–91. doi: 10.1523/JNEUROSCI.10-04-01286.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Rubinow MJ, Drogos LL, Juraska JM. Age-related dendritic hypertrophy and sexual dimorphism in rat basolateral amygdala. Neurobiology of aging. 2009;30:137–46. doi: 10.1016/j.neurobiolaging.2007.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Cooke BM, Woolley CS. Sexually dimorphic synaptic organization of the medial amygdala. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:10759–67. doi: 10.1523/JNEUROSCI.2919-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Leuner B, Shors TJ. New spines, new memories. Mol Neurobiol. 2004;29:117–30. doi: 10.1385/MN:29:2:117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199.Vyas A, Mitra R, Shankaranarayana Rao BS, Chattarji S. Chronic stress induces contrasting patterns of dendritic remodeling in hippocampal and amygdaloid neurons. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2002;22:6810–8. doi: 10.1523/JNEUROSCI.22-15-06810.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Farrell MR, Sengelaub DR, Wellman CL. Sex differences and chronic stress effects on the neural circuitry underlying fear conditioning and extinction. Physiology & behavior. 2013 doi: 10.1016/j.physbeh.2013.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Kim MJ, Loucks RA, Palmer AL, Brown AC, Solomon KM, Marchante AN, Whalen PJ. The structural and functional connectivity of the amygdala: from normal emotion to pathological anxiety. Behav Brain Res. 2011;223:403–10. doi: 10.1016/j.bbr.2011.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Sotres-Bayon F, Quirk GJ. Prefrontal control of fear: more than just extinction. Curr Opin Neurobiol. 2010;20:231–5. doi: 10.1016/j.conb.2010.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Shansky RM, Hamo C, Hof PR, McEwen BS, Morrison JH. Stress-induced dendritic remodeling in the prefrontal cortex is circuit specific. Cereb Cortex. 2009;19:2479–84. doi: 10.1093/cercor/bhp003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Shansky RM, Hamo C, Hof PR, Lou W, McEwen BS, Morrison JH. Estrogen promotes stress sensitivity in a prefrontal cortex-amygdala pathway. Cereb Cortex. 2010;20:2560–7. doi: 10.1093/cercor/bhq003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.McEwen BS. Plasticity of the hippocampus: adaptation to chronic stress and allostatic load. Annals of the New York Academy of Sciences. 2001;933:265–77. doi: 10.1111/j.1749-6632.2001.tb05830.x. [DOI] [PubMed] [Google Scholar]
- 206.Galea LA, McEwen BS, Tanapat P, Deak T, Spencer RL, Dhabhar FS. Sex differences in dendritic atrophy of CA3 pyramidal neurons in response to chronic restraint stress. Neuroscience. 1997;81:689–97. doi: 10.1016/s0306-4522(97)00233-9. [DOI] [PubMed] [Google Scholar]
- 207.McLaughlin KJ, Gomez JL, Baran SE, Conrad CD. The effects of chronic stress on hippocampal morphology and function: an evaluation of chronic restraint paradigms. Brain Res. 2007;1161:56–64. doi: 10.1016/j.brainres.2007.05.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Luine VN, Spencer RL, McEwen BS. Effects of chronic corticosterone ingestion on spatial memory performance and hippocampal serotonergic function. Brain Res. 1993;616:65–70. doi: 10.1016/0006-8993(93)90193-q. [DOI] [PubMed] [Google Scholar]
- 209.Luine V, Villegas M, Martinez C, McEwen BS. Repeated stress causes reversible impairments of spatial memory performance. Brain Res. 1994;639:167–70. doi: 10.1016/0006-8993(94)91778-7. [DOI] [PubMed] [Google Scholar]
- 210.Luine V, Villegas M, Martinez C, McEwen BS. Stress-dependent impairments of spatial memory. Role of 5-HT. Annals of the New York Academy of Sciences. 1994;746:403–4. doi: 10.1111/j.1749-6632.1994.tb39268.x. [DOI] [PubMed] [Google Scholar]
- 211.Song L, Che W, Min-Wei W, Murakami Y, Matsumoto K. Impairment of the spatial learning and memory induced by learned helplessness and chronic mild stress. Pharmacology, biochemistry, and behavior. 2006;83:186–93. doi: 10.1016/j.pbb.2006.01.004. [DOI] [PubMed] [Google Scholar]
- 212.McEwen BS, Conrad CD, Kuroda Y, Frankfurt M, Magarinos AM, McKittrick C. Prevention of stress-induced morphological and cognitive consequences. Eur Neuropsychopharmacol. 1997;7(Suppl 3):S323–8. doi: 10.1016/s0924-977x(97)00064-3. [DOI] [PubMed] [Google Scholar]
- 213.Watanabe Y, Gould E, Daniels DC, Cameron H, McEwen BS. Tianeptine attenuates stress-induced morphological changes in the hippocampus. European journal of pharmacology. 1992;222:157–62. doi: 10.1016/0014-2999(92)90830-w. [DOI] [PubMed] [Google Scholar]
- 214.Conrad CD, Galea LA, Kuroda Y, McEwen BS. Chronic stress impairs rat spatial memory on the Y maze, and this effect is blocked by tianeptine pretreatment. Behavioral neuroscience. 1996;110:1321–34. doi: 10.1037//0735-7044.110.6.1321. [DOI] [PubMed] [Google Scholar]
- 215.McLaughlin KJ, Baran SE, Conrad CD. Chronic stress- and sex-specific neuromorphological and functional changes in limbic structures. Mol Neurobiol. 2009;40:166–82. doi: 10.1007/s12035-009-8079-7. [DOI] [PubMed] [Google Scholar]
- 216.McLaughlin KJ, Wilson JO, Harman J, Wright RL, Wieczorek L, Gomez J, Korol DL, Conrad CD. Chronic 17beta-estradiol or cholesterol prevents stress-induced hippocampal CA3 dendritic retraction in ovariectomized female rats: possible correspondence between CA1 spine properties and spatial acquisition. Hippocampus. 2010;20:768–86. doi: 10.1002/hipo.20678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Bowman RE, Zrull MC, Luine VN. Chronic restraint stress enhances radial arm maze performance in female rats. Brain Res. 2001;904:279–89. doi: 10.1016/s0006-8993(01)02474-x. [DOI] [PubMed] [Google Scholar]
- 218.Kitraki E, Kremmyda O, Youlatos D, Alexis MN, Kittas C. Gender-dependent alterations in corticosteroid receptor status and spatial performance following 21 days of restraint stress. Neuroscience. 2004;125:47–55. doi: 10.1016/j.neuroscience.2003.12.024. [DOI] [PubMed] [Google Scholar]
- 219.Wood GE, Shors TJ. Stress facilitates classical conditioning in males, but impairs classical conditioning in females through activational effects of ovarian hormones. Proceedings of the National Academy of Sciences of the United States of America. 1998;95:4066–71. doi: 10.1073/pnas.95.7.4066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Bangasser DA, Shors TJ. The hippocampus is necessary for enhancements and impairments of learning following stress. Nat Neurosci. 2007;10:1401–3. doi: 10.1038/nn1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221.Servatius RJ, Shors TJ. Early acquisition, but not retention, of the classically conditioned eyeblink response is N-methyl-D-aspartate (NMDA) receptor dependent. Behavioral neuroscience. 1996;110:1040–8. doi: 10.1037//0735-7044.110.5.1040. [DOI] [PubMed] [Google Scholar]
- 222.Shors TJ, Falduto J, Leuner B. The opposite effects of stress on dendritic spines in male vs. female rats are NMDA receptor-dependent. The European journal of neuroscience. 2004;19:145–50. doi: 10.1046/j.1460-9568.2003.03065.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Dalla C, Whetstone AS, Hodes GE, Shors TJ. Stressful experience has opposite effects on dendritic spines in the hippocampus of cycling versus masculinized females. Neurosci Lett. 2009;449:52–6. doi: 10.1016/j.neulet.2008.10.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Shors TJ, Miesegaes G. Testosterone in utero and at birth dictates how stressful experience will affect learning in adulthood. Proc Natl Acad Sci U S A. 2002;99:13955–60. doi: 10.1073/pnas.202199999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Yildirim M, Janssen WG, Tabori NE, Adams MM, Yuen GS, Akama KT, McEwen BS, Milner TA, Morrison JH. Estrogen and aging affect synaptic distribution of phosphorylated LIM kinase (pLIMK) in CA1 region of female rat hippocampus. Neuroscience. 2008;152:360–70. doi: 10.1016/j.neuroscience.2008.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Yildirim M, Janssen WG, Lou WY, Akama KT, McEwen BS, Milner TA, Morrison JH. Effects of estrogen and aging on the synaptic distribution of phosphorylated Akt-immunoreactivity in the CA1 region of the female rat hippocampus. Brain Res. 2011;1379:98–108. doi: 10.1016/j.brainres.2010.07.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Yuen GS, McEwen BS, Akama KT. LIM kinase mediates estrogen action on the actin depolymerization factor Cofilin. Brain Res. 2011;1379:44–52. doi: 10.1016/j.brainres.2010.07.067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Liu RJ, Aghajanian GK. Stress blunts serotonin- and hypocretin-evoked EPSCs in prefrontal cortex: role of corticosterone-mediated apical dendritic atrophy. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:359–64. doi: 10.1073/pnas.0706679105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Magarinos AM, McEwen BS. Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: comparison of stressors. Neuroscience. 1995;69:83–8. doi: 10.1016/0306-4522(95)00256-i. [DOI] [PubMed] [Google Scholar]
- 230.Goldwater DS, Pavlides C, Hunter RG, Bloss EB, Hof PR, McEwen BS, Morrison JH. Structural and functional alterations to rat medial prefrontal cortex following chronic restraint stress and recovery. Neuroscience. 2009;164:798–808. doi: 10.1016/j.neuroscience.2009.08.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231.Martin KP, Wellman CL. NMDA receptor blockade alters stress-induced dendritic remodeling in medial prefrontal cortex. Cereb Cortex. 2011;21:2366–73. doi: 10.1093/cercor/bhr021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Christian KM, Miracle AD, Wellman CL, Nakazawa K. Chronic stress-induced hippocampal dendritic retraction requires CA3 NMDA receptors. Neuroscience. 2011;174:26–36. doi: 10.1016/j.neuroscience.2010.11.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Magarinos AM, Deslandes A, McEwen BS. Effects of antidepressants and benzodiazepine treatments on the dendritic structure of CA3 pyramidal neurons after chronic stress. European journal of pharmacology. 1999;371:113–22. doi: 10.1016/s0014-2999(99)00163-6. [DOI] [PubMed] [Google Scholar]
- 234.American Psychiatric Association., and American Psychiatric Association. DSM-5 Task Force. Diagnostic and statistical manual of mental disorders: DSM-5. American Psychiatric Association; Washington, D.C: 2013. [Google Scholar]
- 235.Coker AL, Weston R, Creson DL, Justice B, Blakeney P. PTSD symptoms among men and women survivors of intimate partner violence: the role of risk and protective factors. Violence Vict. 2005;20:625–43. [PubMed] [Google Scholar]
- 236.Breslau N, Chilcoat HD, Kessler RC, Peterson EL, Lucia VC. Vulnerability to assaultive violence: further specification of the sex difference in post-traumatic stress disorder. Psychol Med. 1999;29:813–21. doi: 10.1017/s0033291799008612. [DOI] [PubMed] [Google Scholar]
- 237.Hall M, Buysse DJ, Nowell PD, Nofzinger EA, Houck P, Reynolds CF, 3rd, Kupfer DJ. Symptoms of stress and depression as correlates of sleep in primary insomnia. Psychosomatic medicine. 2000;62:227–30. doi: 10.1097/00006842-200003000-00014. [DOI] [PubMed] [Google Scholar]
- 238.Perlis ML, Giles DE, Buysse DJ, Tu X, Kupfer DJ. Self-reported sleep disturbance as a prodromal symptom in recurrent depression. J Affect Disord. 1997;42:209–12. doi: 10.1016/s0165-0327(96)01411-5. [DOI] [PubMed] [Google Scholar]
- 239.Plante DT, Landsness EC, Peterson MJ, Goldstein MR, Riedner BA, Wanger T, Guokas JJ, Tononi G, Benca RM. Sex-related differences in sleep slow wave activity in major depressive disorder: a high-density EEG investigation. BMC Psychiatry. 2012;12:146. doi: 10.1186/1471-244X-12-146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 240.Plante DT, Goldstein MR, Landsness EC, Peterson MJ, Riedner BA, Ferrarelli F, Wanger T, Guokas JJ, Tononi G, Benca RM. Topographic and sex-related differences in sleep spindles in major depressive disorder: a high-density EEG investigation. J Affect Disord. 2013;146:120–5. doi: 10.1016/j.jad.2012.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 241.Kobayashi I, Mellman TA. Gender differences in sleep during the aftermath of trauma and the development of posttraumatic stress disorder. Behav Sleep Med. 2012;10:180–90. doi: 10.1080/15402002.2011.654296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Richards A, Metzler TJ, Ruoff LM, Inslicht SS, Rao M, Talbot LS, Neylan TC. Sex differences in objective measures of sleep in post-traumatic stress disorder and healthy control subjects. J Sleep Res. 2013 doi: 10.1111/jsr.12064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 243.Nolen-Hoeksema S, Larson J, Grayson C. Explaining the gender difference in depressive symptoms. J Pers Soc Psychol. 1999;77:1061–72. doi: 10.1037//0022-3514.77.5.1061. [DOI] [PubMed] [Google Scholar]
- 244.Mezo PG, Baker RM. The moderating effects of stress and rumination on depressive symptoms in women and men. Stress Health. 2012;28:333–9. doi: 10.1002/smi.2417. [DOI] [PubMed] [Google Scholar]
- 245.Thomsen DK, Yung Mehlsen M, Christensen S, Zachariae R. Rumination—relationship with negative mood and sleep quality. Personality and Individual Differences. 2003;34:1293–1301. [Google Scholar]
- 246.Pedersen WC, Denson TF, Goss RJ, Vasquez EA, Kelley NJ, Miller N. The impact of rumination on aggressive thoughts, feelings, arousal, and behaviour. Br J Soc Psychol. 2011;50:281–301. doi: 10.1348/014466610X515696. [DOI] [PubMed] [Google Scholar]
- 247.Andersen SB, Moore RA, Venables L, Corr PJ. Electrophysiological correlates of anxious rumination. International Journal of Psychophysiology. 2009;71:156–169. doi: 10.1016/j.ijpsycho.2008.09.004. [DOI] [PubMed] [Google Scholar]
- 248.Nolan S, Roberts J, Gotlib I. Neuroticism and Ruminative Response Style as Predictors of Change in Depressive Symptomatology. Cognitive Therapy and Research. 1998;22:445–455. [Google Scholar]
- 249.Nolen-Hoeksema S, Wisco BE, Lyubomirsky S. Rethinking Rumination. Perspectives on Psychological Science. 2008;3:400–424. doi: 10.1111/j.1745-6924.2008.00088.x. [DOI] [PubMed] [Google Scholar]
- 250.O’Donnell T, Hegadoren KM, Coupland NC. Noradrenergic mechanisms in the pathophysiology of post-traumatic stress disorder. Neuropsychobiology. 2004;50:273–83. doi: 10.1159/000080952. [DOI] [PubMed] [Google Scholar]
- 251.Southwick SM, Bremner JD, Rasmusson A, Morgan CA, 3rd, Arnsten A, Charney DS. Role of norepinephrine in the pathophysiology and treatment of posttraumatic stress disorder. Biol Psychiatry. 1999;46:1192–204. doi: 10.1016/s0006-3223(99)00219-x. [DOI] [PubMed] [Google Scholar]
- 252.Gold PW, Wong ML, Chrousos GP, Licinio J. Stress system abnormalities in melancholic and atypical depression: molecular, pathophysiological, and therapeutic implications. Mol Psychiatry. 1996;1:257–64. [PubMed] [Google Scholar]
- 253.Gold PW, Chrousos GP. The endocrinology of melancholic and atypical depression: relation to neurocircuitry and somatic consequences. Proc Assoc Am Physicians. 1999;111:22–34. doi: 10.1046/j.1525-1381.1999.09423.x. [DOI] [PubMed] [Google Scholar]
- 254.Koob GF. Corticotropin-releasing factor, norepinephrine, and stress. Biol Psychiatry. 1999;46:1167–80. doi: 10.1016/s0006-3223(99)00164-x. [DOI] [PubMed] [Google Scholar]
- 255.Swanson LW, Hartman BK. The central adrenergic system. An immunofluorescence study of the location of cell bodies and their efferent connections in the rat utilizing dopamine-beta-hydroxylase as a marker. J Comp Neurol. 1975;163:467–505. doi: 10.1002/cne.901630406. [DOI] [PubMed] [Google Scholar]
- 256.Foote SL, Bloom FE, Aston-Jones G. Nucleus locus ceruleus: new evidence of anatomical and physiological specificity. Physiol Rev. 1983;63:844–914. doi: 10.1152/physrev.1983.63.3.844. [DOI] [PubMed] [Google Scholar]
- 257.Berridge CW, Foote SL. Effects of locus coeruleus activation on electroencephalographic activity in the neocortex and hippocampus. J Neurosci. 1991;11:3135–3145. doi: 10.1523/JNEUROSCI.11-10-03135.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258.Berridge CW, Page ME, Valentino RJ, Foote SL. Effects of locus coeruleus inactivation on electroencephalographic activity in neocortex and hippocampus. Neuroscience. 1993;55:381–383. doi: 10.1016/0306-4522(93)90507-c. [DOI] [PubMed] [Google Scholar]
- 259.Abercrombie ED, Keller RW, Zigmond MJ. Characterization of hippocampal norepinephrine release as measured by microdialysis perfusion: pharmacological and behavioral studies. Neuroscience. 1988;27:897–904. doi: 10.1016/0306-4522(88)90192-3. [DOI] [PubMed] [Google Scholar]
- 260.Berridge CW, Waterhouse BD. The locus coeruleus-noradrenergic system: modulation of behavioral state and state-dependent cognitive processes. Brain Res Brain Res Rev. 2003;42:33–84. doi: 10.1016/s0165-0173(03)00143-7. [DOI] [PubMed] [Google Scholar]
- 261.Gary MGS, PhD, Aston-Jones S, Doran GSPD, Aston-Jones Monica Gonzalez, Doran S. Brain Norepinephrine. Cambridge University Press; 2007. Role of the locus coeruleus-norepinephrine system in arousal and circadian regulation of the sleep–wake cycle. [Google Scholar]
- 262.Geracioti TD, Jr, Baker DG, Ekhator NN, West SA, Hill KK, Bruce AB, Schmidt D, Rounds-Kugler B, Yehuda R, Keck PE, Jr, Kasckow JW. CSF norepinephrine concentrations in posttraumatic stress disorder. Am J Psychiatry. 2001;158:1227–30. doi: 10.1176/appi.ajp.158.8.1227. [DOI] [PubMed] [Google Scholar]
- 263.Wong ML, Kling MA, Munson PJ, Listwak S, Licinio J, Prolo P, Karp B, McCutcheon IE, Geracioti TD, Jr, DeBellis MD, Rice KC, Goldstein DS, Veldhuis JD, Chrousos GP, Oldfield EH, McCann SM, Gold PW. Pronounced and sustained central hypernoradrenergic function in major depression with melancholic features: relation to hypercortisolism and corticotropin-releasing hormone. Proc Natl Acad Sci U S A. 2000;97:325–30. doi: 10.1073/pnas.97.1.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 264.Garcia-Falgueras A, Pinos H, Fernandez R, Collado P, Pasaro E, Segovia S, Guillamon A. Sexual dimorphism in hybrids rats. Brain Res. 2006;1123:42–50. doi: 10.1016/j.brainres.2006.09.053. [DOI] [PubMed] [Google Scholar]
- 265.Garcia-Falgueras A, Pinos H, Collado P, Pasaro E, Fernandez R, Segovia S, Guillamon A. The expression of brain sexual dimorphism in artificial selection of rat strains. Brain Res. 2005;1052:130–8. doi: 10.1016/j.brainres.2005.05.066. [DOI] [PubMed] [Google Scholar]
- 266.Guillamon A, de Blas MR, Segovia S. Effects of sex steroids on the development of the locus coeruleus in the rat. Brain Res. 1988;468:306–10. doi: 10.1016/0165-3806(88)90143-5. [DOI] [PubMed] [Google Scholar]
- 267.Bangasser DA, Reyes BA, Piel D, Garachh V, Zhang XY, Plona ZM, Van Bockstaele EJ, Beck SG, Valentino RJ. Increased vulnerability of the brain norepinephrine system of females to corticotropin-releasing factor overexpression. Mol Psychiatry. 2013;18:166–73. doi: 10.1038/mp.2012.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 268.Bangasser DA, Zhang X, Garachh V, Hanhauser E, Valentino RJ. Sexual dimorphism in locus coeruleus dendritic morphology: a structural basis for sex differences in emotional arousal. Physiol Behav. 2011;103:342–51. doi: 10.1016/j.physbeh.2011.02.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Van Bockstaele EJ, Bajic D, Proudfit H, Valentino RJ. Topographic architecture of stress-related pathways targeting the noradrenergic locus coeruleus. Physiol and Behav. 2001;73:273–283. doi: 10.1016/s0031-9384(01)00448-6. [DOI] [PubMed] [Google Scholar]
- 270.Aston-Jones G, Ennis M, Pieribone VA, Nickell WT, Shipley MT. The brain nucleus locus coeruleus: restricted afferent control of a broad efferent network. Science. 1986;234:734–737. doi: 10.1126/science.3775363. [DOI] [PubMed] [Google Scholar]
- 271.Valentino RJ, Chen S, Zhu Y, Aston-Jones G. Evidence for divergent projections of corticotropin-releasing hormone neurons of Barrington’s nucleus to the locus coeruleus and spinal cord. Brain Res. 1996;732:1–15. doi: 10.1016/0006-8993(96)00482-9. [DOI] [PubMed] [Google Scholar]
- 272.Van Bockstaele EJ, Colago EE, Valentino RJ. Amygdaloid corticotropin-releasing factor targets locus coeruleus dendrites: substrate for the co-ordination of emotional and cognitive limbs of the stress response. J Neuroendocrinol. 1998;10:743–57. doi: 10.1046/j.1365-2826.1998.00254.x. [DOI] [PubMed] [Google Scholar]
- 273.Van Bockstaele EJ, Peoples J, Valentino RJ A.E. Bennett Research Award. Anatomic basis for differential regulation of the rostrolateral peri-locus coeruleus region by limbic afferents. Biol Psychiatry. 1999;46:1352–63. doi: 10.1016/s0006-3223(99)00213-9. [DOI] [PubMed] [Google Scholar]
- 274.Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving rats anticipates fluctuations in the sleep-waking cycle. J Neurosci. 1981;1:876–886. doi: 10.1523/JNEUROSCI.01-08-00876.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 275.Aston-Jones G, Cohen JD. An integrative theory of locus coeruleus-norepinephrine function: adaptive gain and optimal performance. Annu Rev Neurosci. 2005;28:403–50. doi: 10.1146/annurev.neuro.28.061604.135709. [DOI] [PubMed] [Google Scholar]
- 276.Clayton EC, Rajkowski J, Cohen JD, Aston-Jones G. Phasic activation of monkey locus ceruleus neurons by simple decisions in a forced-choice task. J Neurosci. 2004;24:9914–20. doi: 10.1523/JNEUROSCI.2446-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277.Foote SL, Aston-Jones G, Bloom FE. Impulse activity of locus coeruleus neurons in awake rats and monkeys is a function of sensory stimulation and arousal. Proc Natl Acad Sci USA. 1980;77:3033–3037. doi: 10.1073/pnas.77.5.3033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 278.Curtis AL, Bethea T, Valentino RJ. Sexually dimorphic responses of the brain norepinephrine system to stress and corticotropin-releasing factor. Neuropsychopharmacology. 2006;31:544–54. doi: 10.1038/sj.npp.1300875. [DOI] [PubMed] [Google Scholar]
- 279.Curtis AL, Drolet G, Valentino RJ. Hemodynamic stress activates locus coeruleus neurons of unanesthetized rats. Brain Res Bull. 1993;31:737–44. doi: 10.1016/0361-9230(93)90150-a. [DOI] [PubMed] [Google Scholar]
- 280.Valentino RJ, Foote SL. Corticotropin-releasing factor disrupts sensory responses of brain noradrenergic neurons. Neuroendocrinology. 1987;45:28–36. doi: 10.1159/000124700. [DOI] [PubMed] [Google Scholar]
- 281.Valentino RJ, Foote SL. Corticotropin-releasing hormone increases tonic but not sensory-evoked activity of noradrenergic locus coeruleus neurons in unanesthetized rats. J Neurosci. 1988;8:1016–25. doi: 10.1523/JNEUROSCI.08-03-01016.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 282.Page ME, Berridge CW, Foote SL, Valentino RJ. Corticotropin-releasing factor in the locus coeruleus mediates EEG activation associated with hypotensive stress. Neurosci Lett. 1993;164:81–84. doi: 10.1016/0304-3940(93)90862-f. [DOI] [PubMed] [Google Scholar]
- 283.Melia KR, Duman RS. Involvement of corticotropin-releasing factor in chronic stress regulation of the brain noradrenergic system. Proc Natl Acad Sci. 1991;88:8382–8386. doi: 10.1073/pnas.88.19.8382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 284.Smagin GN, Zhou J, Harris RB, Ryan DH. CRF receptor antagonist attenuates immobilization stress-induced norepinephrine release in the prefrontal cortex in rats. Brain Res Bull. 1997;42:431–4. doi: 10.1016/s0361-9230(96)00368-1. [DOI] [PubMed] [Google Scholar]
- 285.Valentino RJ, Wehby RG. Corticotropin-releasing factor: evidence for a neurotransmitter role in the locus ceruleus during hemodynamic stress. Neuroendocrinology. 1988;48:674–7. doi: 10.1159/000125081. [DOI] [PubMed] [Google Scholar]
- 286.Valentino RJ, Page ME, Curtis AL. Activation of noradrenergic locus coeruleus neurons by hemodynamic stress is due to local release of corticotropin-releasing factor. Brain Res. 1991;555:25–34. doi: 10.1016/0006-8993(91)90855-p. [DOI] [PubMed] [Google Scholar]
- 287.Curtis AL, Grigoriadis DE, Page ME, Rivier J, Valentino RJ. Pharmacological comparison of two corticotropin-releasing factor antagonists: in vivo and in vitro studies. J Pharmacol Exp Ther. 1994;268:359–65. [PubMed] [Google Scholar]
- 288.Snyder K, Wang WW, Han R, McFadden K, Valentino RJ. Corticotropin-Releasing Factor in the Norepinephrine Nucleus, Locus Coeruleus, Facilitates Behavioral Flexibility. Neuropsychopharmacology. 2011 doi: 10.1038/npp.2011.218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289.Curtis AL, Pavcovich LA, Grigoriadis DE, Valentino RJ. Previous stress alters corticotropin-releasing factor neurotransmission in the locus coeruleus. Neuroscience. 1995;65:541–50. doi: 10.1016/0306-4522(94)00496-r. [DOI] [PubMed] [Google Scholar]
- 290.Curtis AL, Pavcovich LA, Valentino RJ. Long-term regulation of locus ceruleus sensitivity to corticotropin-releasing factor by swim stress. J Pharmacol Exp Ther. 1999;289:1211–9. [PubMed] [Google Scholar]
- 291.Jedema HP, Grace AA. Corticotropin-releasing hormone directly activates noradrenergic neurons of the locus ceruleus recorded in vitro. J Neurosci. 2004;24:9703–13. doi: 10.1523/JNEUROSCI.2830-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 292.Schulz C, Lehnert H. Activation of noradrenergic neurons in the locus coeruleus by corticotropin-releasing factor, a microdialysis study. Neuroendocrinology. 1996;63:454–458. doi: 10.1159/000127071. [DOI] [PubMed] [Google Scholar]
- 293.Hillhouse EW, Grammatopoulos DK. The molecular mechanisms underlying the regulation of the biological activity of corticotropin-releasing hormone receptors: implications for physiology and pathophysiology. Endocr Rev. 2006;27:260–86. doi: 10.1210/er.2005-0034. [DOI] [PubMed] [Google Scholar]
- 294.Chen FM, Bilezikjian LM, Perrin MH, Rivier J, Vale W. Corticotropin releasing factor receptor-mediated stimulation of adenylate cyclase activity in the rat brain. Brain Res. 1986;381:49–57. doi: 10.1016/0006-8993(86)90688-8. [DOI] [PubMed] [Google Scholar]
- 295.Grammatopoulos DK, Randeva HS, Levine MA, Kanellopoulou KA, Hillhouse EW. Rat cerebral cortex corticotropin-releasing hormone receptors: evidence for receptor coupling to multiple G-proteins. J Neurochem. 2001;76:509–19. doi: 10.1046/j.1471-4159.2001.00067.x. [DOI] [PubMed] [Google Scholar]
- 296.Bangasser DA, Curtis A, Reyes BA, Bethea TT, Parastatidis I, Ischiropoulos H, Van Bockstaele EJ, Valentino RJ. Sex differences in corticotropin-releasing factor receptor signaling and trafficking: potential role in female vulnerability to stress-related psychopathology. Mol Psychiatry. 2010;15:877, 896–904. doi: 10.1038/mp.2010.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 297.Oakley RH, Olivares-Reyes JA, Hudson CC, Flores-Vega F, Dautzenberg FM, Hauger RL. Carboxyl-terminal and intracellular loop sites for CRF1 receptor phosphorylation and beta-arrestin-2 recruitment: a mechanism regulating stress and anxiety responses. Am J Physiol Regul Integr Comp Physiol. 2007;293:R209–22. doi: 10.1152/ajpregu.00099.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298.Teli T, Markovic D, Levine MA, Hillhouse EW, Grammatopoulos DK. Regulation of corticotropin-releasing hormone receptor type 1alpha signaling: structural determinants for G protein-coupled receptor kinase-mediated phosphorylation and agonist-mediated desensitization. Mol Endocrinol. 2005;19:474–90. doi: 10.1210/me.2004-0275. [DOI] [PubMed] [Google Scholar]
- 299.Holmes KD, Babwah AV, Dale LB, Poulter MO, Ferguson SS. Differential regulation of corticotropin releasing factor 1alpha receptor endocytosis and trafficking by beta-arrestins and Rab GTPases. J Neurochem. 2006;96:934–49. doi: 10.1111/j.1471-4159.2005.03603.x. [DOI] [PubMed] [Google Scholar]
- 300.Reyes BA, Fox K, Valentino RJ, Van Bockstaele EJ. Agonist-induced internalization of corticotropin-releasing factor receptors in noradrenergic neurons of the rat locus coeruleus. Eur J Neurosci. 2006;23:2991–8. doi: 10.1111/j.1460-9568.2006.04820.x. [DOI] [PubMed] [Google Scholar]
- 301.Reyes BA, Valentino RJ, Van Bockstaele EJ. Stress-induced intracellular trafficking of corticotropin-releasing factor receptors in rat locus coeruleus neurons. Endocrinology. 2008;149:122–30. doi: 10.1210/en.2007-0705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 302.Krupnick JG, Benovic JL. The role of receptor kinases and arrestins in G protein-coupled receptor regulation. Annu Rev Pharmacol Toxicol. 1998;38:289–319. doi: 10.1146/annurev.pharmtox.38.1.289. [DOI] [PubMed] [Google Scholar]
- 303.Premont RT. Once and future signaling: G protein-coupled receptor kinase control of neuronal sensitivity. Neuromolecular Med. 2005;7:129–47. doi: 10.1385/NMM:7:1-2:129. [DOI] [PubMed] [Google Scholar]
- 304.Lefkowitz RJ. G protein-coupled receptors. III. New roles for receptor kinases and beta-arrestins in receptor signaling and desensitization. J Biol Chem. 1998;273:18677–80. doi: 10.1074/jbc.273.30.18677. [DOI] [PubMed] [Google Scholar]
- 305.Lefkowitz RJ, Shenoy SK. Transduction of receptor signals by beta-arrestins. Science. 2005;308:512–7. doi: 10.1126/science.1109237. [DOI] [PubMed] [Google Scholar]
- 306.Violin JD, Lefkowitz RJ. Beta-arrestin-biased ligands at seven-transmembrane receptors. Trends Pharmacol Sci. 2007;28:416–22. doi: 10.1016/j.tips.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 307.DeWire SM, Ahn S, Lefkowitz RJ, Shenoy SK. Beta-arrestins and cell signaling. Annu Rev Physiol. 2007;69:483–510. doi: 10.1146/annurev.physiol.69.022405.154749. [DOI] [PubMed] [Google Scholar]
- 308.Shukla AK, Xiao K, Lefkowitz RJ. Emerging paradigms of beta-arrestin-dependent seven transmembrane receptor signaling. Trends Biochem Sci. 2011;36:457–69. doi: 10.1016/j.tibs.2011.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309.Valentino RJ, Van Bockstaele E, Bangasser D. Sex-specific cell signaling: the corticotropin-releasing factor receptor model. Trends Pharmacol Sci. 2013;34:437–44. doi: 10.1016/j.tips.2013.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 310.Valentino RJ, Bangasser D, Van Bockstaele EJ. Sex-biased stress signaling: the corticotropin-releasing factor receptor as a model. Molecular pharmacology. 2013;83:737–45. doi: 10.1124/mol.112.083550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311.Bangasser DA, Valentino RJ. Sex differences in molecular and cellular substrates of stress. Cell Mol Neurobiol. 2012;32:709–23. doi: 10.1007/s10571-012-9824-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312.Arnold AP. The organizational-activational hypothesis as the foundation for a unified theory of sexual differentiation of all mammalian tissues. Hormones and behavior. 2009;55:570–8. doi: 10.1016/j.yhbeh.2009.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 313.Juraska JM, Sisk CL, DonCarlos LL. Sexual differentiation of the adolescent rodent brain: hormonal influences and developmental mechanisms. Hormones and behavior. 2013;64:203–10. doi: 10.1016/j.yhbeh.2013.05.010. [DOI] [PubMed] [Google Scholar]
- 314.Schulz KM, Molenda-Figueira HA, Sisk CL. Back to the future: The organizational-activational hypothesis adapted to puberty and adolescence. Horm Behav. 2009;55:597–604. doi: 10.1016/j.yhbeh.2009.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 315.Phoenix CH, Goy RW, Gerall AA, Young WC. Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology. 1959;65:369–82. doi: 10.1210/endo-65-3-369. [DOI] [PubMed] [Google Scholar]
- 316.Schulz KM, Richardson HN, Zehr JL, Osetek AJ, Menard TA, Sisk CL. Gonadal hormones masculinize and defeminize reproductive behaviors during puberty in the male Syrian hamster. Horm Behav. 2004;45:242–9. doi: 10.1016/j.yhbeh.2003.12.007. [DOI] [PubMed] [Google Scholar]
- 317.Angold A, Worthman CW. Puberty onset of gender differences in rates of depression: a developmental, epidemiologic and neuroendocrine perspective. J Affect Disord. 1993;29:145–58. doi: 10.1016/0165-0327(93)90029-j. [DOI] [PubMed] [Google Scholar]
- 318.Douma SL, Husband C, O’Donnell ME, Barwin BN, Woodend AK. Estrogen-related mood disorders: reproductive life cycle factors. ANS Adv Nurs Sci. 2005;28:364–75. doi: 10.1097/00012272-200510000-00008. [DOI] [PubMed] [Google Scholar]
- 319.Schmidt PJ, Rubinow DR. Sex hormones and mood in the perimenopause. Annals of the New York Academy of Sciences. 2009;1179:70–85. doi: 10.1111/j.1749-6632.2009.04982.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320.Hendrick V, Altshuler LL, Suri R. Hormonal Changes in the Postpartum and Implications for Postpartum Depression. Psychosomatics. 1998;39:93–101. doi: 10.1016/S0033-3182(98)71355-6. [DOI] [PubMed] [Google Scholar]
- 321.Solomon MB, Herman JP. Sex differences in psychopathology: of gonads, adrenals and mental illness. Physiology & behavior. 2009;97:250–8. doi: 10.1016/j.physbeh.2009.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 322.Rubinow DR, Roca CA, Schmidt PJ. Chapter 24 - Estrogens and depression in women. In: Rogerio AL, MDMDA, Lobo Rogerio A, editors. Treatment of the Postmenopausal Woman. 3. Academic Press; St. Louis: 2007. pp. 307–320. [Google Scholar]
- 323.Steiner M, Dunn E, Born L. Hormones and mood: from menarche to menopause and beyond. Journal of Affective Disorders. 2003;74:67–83. doi: 10.1016/s0165-0327(02)00432-9. [DOI] [PubMed] [Google Scholar]
- 324.Schmidt PJ, Nieman L, Danaceau MA, Tobin MB, Roca CA, Murphy JH, Rubinow DR. Estrogen replacement in perimenopause-related depression: a preliminary report. Am J Obstet Gynecol. 2000;183:414–20. doi: 10.1067/mob.2000.106004. [DOI] [PubMed] [Google Scholar]
- 325.Soares CN, Almeida OP, Joffe H, Cohen LS. Efficacy of estradiol for the treatment of depressive disorders in perimenopausal women: a double-blind, randomized, placebo-controlled trial. Archives of general psychiatry. 2001;58:529–34. doi: 10.1001/archpsyc.58.6.529. [DOI] [PubMed] [Google Scholar]
- 326.Glover EM, Jovanovic T, Mercer KB, Kerley K, Bradley B, Ressler KJ, Norrholm SD. Estrogen levels are associated with extinction deficits in women with posttraumatic stress disorder. Biol Psychiatry. 2012;72:19–24. doi: 10.1016/j.biopsych.2012.02.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327.Bangasser DA, Shors TJ. The bed nucleus of the stria terminalis modulates learning after stress in masculinized but not cycling females. J Neurosci. 2008;28:6383–7. doi: 10.1523/JNEUROSCI.0831-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 328.Kopsida E, Lynn PM, Humby T, Wilkinson LS, Davies W. Dissociable effects of sry and sex chromosome complement on activity, feeding and anxiety-related behaviours in mice. PLoS One. 2013;8:e73699. doi: 10.1371/journal.pone.0073699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 329.Rush AJ, Trivedi MH, Wisniewski SR, Nierenberg AA, Stewart JW, Warden D, Niederehe G, Thase ME, Lavori PW, Lebowitz BD, McGrath PJ, Rosenbaum JF, Sackeim HA, Kupfer DJ, Luther J, Fava M. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: a STAR*D report. Am J Psychiatry. 2006;163:1905–17. doi: 10.1176/ajp.2006.163.11.1905. [DOI] [PubMed] [Google Scholar]
- 330.Foa EB, Keane TM, Friedman MJ, Cohen JA. Effective treatments for PTSD: practice guidelines from the International Society for Traumatic Stress Studies. Guilford Press; 2008. [Google Scholar]
- 331.Drake MT, Violin JD, Whalen EJ, Wisler JW, Shenoy SK, Lefkowitz RJ. beta-arrestin-biased agonism at the beta2-adrenergic receptor. J Biol Chem. 2008;283:5669–76. doi: 10.1074/jbc.M708118200. [DOI] [PubMed] [Google Scholar]
- 332.Ahn S, Shenoy SK, Wei H, Lefkowitz RJ. Differential kinetic and spatial patterns of beta-arrestin and G protein-mediated ERK activation by the angiotensin II receptor. J Biol Chem. 2004;279:35518–25. doi: 10.1074/jbc.M405878200. [DOI] [PubMed] [Google Scholar]
- 333.Whalen EJ, Rajagopal S, Lefkowitz RJ. Therapeutic potential of beta-arrestin- and G protein-biased agonists. Trends Mol Med. 2011;17:126–39. doi: 10.1016/j.molmed.2010.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 334.Wei H, Ahn S, Shenoy SK, Karnik SS, Hunyady L, Luttrell LM, Lefkowitz RJ. Independent beta-arrestin 2 and G protein-mediated pathways for angiotensin II activation of extracellular signal-regulated kinases 1 and 2. Proc Natl Acad Sci U S A. 2003;100:10782–7. doi: 10.1073/pnas.1834556100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 335.Beery AK, Zucker I. Sex bias in neuroscience and biomedical research. Neurosci Biobehav Rev. 2011;35:565–72. doi: 10.1016/j.neubiorev.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 336.NC Survey. 2005 http://www.hcp.med.harvard.edu/ncs/ftpdir/NCS-R_Lifetime_Prevalence_Estimates.pdf.