

Abstract
Background
Chronic social defeat stress (CSDS) produces persistent behavioral adaptations in mice. In many behavioral assays, it can be difficult to determine if these adaptations reflect core signs of depression. We designed studies to characterize the effects of CSDS on sensitivity to reward, since anhedonia (reduced sensitivity to reward) is a defining characteristic of depressive disorders in humans. We also examined the effects of striatal ΔFosB overexpression or the N-methyl-D-aspartate antagonist ketamine, both of which promote resilience, on CSDS-induced alterations in reward function and social interaction.
Methods
We used intracranial self-stimulation (ICSS) to quantify CSDS-induced changes in reward function. Mice were implanted with lateral hypothalamic (LH) electrodes and ICSS thresholds were measured following each of 10 daily CSDS sessions, and during a 5-day recovery period. We also examined if acute administration of ketamine (2.5–20 mg/kg, intraperitoneal) reverses CSDS-induced effects on reward or, in separate mice, social interaction.
Results
CSDS increased ICSS thresholds, indicating decreases in the rewarding impact of LH stimulation (anhedonia). This effect was attenuated in mice overexpressing ΔFosB in striatum, consistent with pro-resilient actions of this transcription factor. High but not low doses of ketamine administered after completion of the CSDS regimen attenuated social avoidance in defeated mice, although this effect was transient. Ketamine did not block CSDS-induced anhedonia in the ICSS test.
Conclusions
Our findings demonstrate that CSDS triggers persistent anhedonia, and confirm that ΔFosB overexpression produces stress resilience. They also indicate that acute ketamine fails to attenuate CSDS-induced anhedonia despite reducing other depression-related behavioral abnormalities.
Keywords: defeat, stress, anhedonia, intracranial self-stimulation (ICSS), social interaction, ketamine, antidepressant
INTRODUCTION
Chronic stress is implicated in the etiology and pathophysiology of anxiety and depressive disorders (1–3). Although these disorders are increasingly prevalent (4) and tend to be persistent and resistant to current treatments (5,6), the mechanisms by which stress triggers them remain poorly understood (7). Validating depression models is crucial to better understand the consequences of stress, elucidate the neurobiology of affective disorders, and develop novel anti-stress and antidepressant treatments.
Animal models of depression rely on their ability to mimic or produce core symptoms of the disorder in humans, including social avoidance and anhedonia (reduced sensitivity to reward) (8,9). Chronic social defeat stress (CSDS) is an increasingly utilized model that exploits the ethological relevance of territorial aggression (10,11) and produces these core symptoms as assessed in tests quantifying social interaction and preference for sucrose and other natural rewards (12–14). Furthermore, CSDS effects are reversed by chronic but not acute administration of fluoxetine or imipramine (12,15,16), standard antidepressant medications widely used to treat depressive disorders in humans. In contrast, standard anxiolytic medications are ineffective (12). Thus CSDS is considered to have construct, face, and predicative validity (10). However, it has been proposed that CSDS-triggered behaviors have their basis in anxiety (17,18), and what is often interpreted as anhedonia in sucrose preference tests may actually reflect anxiety-enhanced neophobia.
The primary goal of the present studies was to examine the ability of CSDS to produce anhedonia, a core feature of depressive but not anxiety disorders (8). We used intracranial self-stimulation (ICSS), an operant paradigm in which mice self-administer rewarding electrical brain stimulation, to directly assess the effects of CSDS on reward sensitivity (19,20). ICSS behavior is attenuated in rodents under conditions that cause depressive-like states in humans, including drug withdrawal (21–25), unpredictable and chronic mild stress (26,27), and administration of kappa-opioid receptor agonists (28,29). Specifically, these treatments increase the threshold frequency at which the stimulation supports responding, an indicator of anhedonia (19). Furthermore, the ICSS paradigm allows for the study of manipulation-induced alterations in reward sensitivity over time, and is impervious to anxiety- and satiety-related factors that confound other paradigms used to assess reward system function (e.g. sucrose preference, sex, drugs of abuse) (19).
In parallel, we examined the ability of ketamine, an NMDA receptor antagonist (30), to mitigate the effects of CSDS on social avoidance and ICSS thresholds. While standard antidepressant treatments have delayed therapeutic efficacy (often several weeks), recent studies demonstrate that a single dose of ketamine can produce rapid (though transient) antidepressant responses in depressed patients (31–37)—including patients that are treatment-resistant (32,34,38,39)—and antidepressant-like effects in numerous models of depression (40–51). To determine if the therapeutic-like actions of ketamine are accompanied by amnestic (learning- and memory-disrupting) effects often associated with NMDA antagonists (52,53) or anxiolytic effects (43) we examined performance in the passive-avoidance and elevated plus maze (EPM) tests. As a way of evaluating whether it is possible to mitigate the effects of CSDS on ICSS, we included studies using ΔFosB-overexpressing mice, which are less sensitive (resilient) to CSDS (54).
MATERIALS AND METHODS
Animals and Drugs
Male C57BL/6J mice (6–8 weeks) were purchased from Jackson Laboratories (Bar Harbor, ME), and male CD1 mice (retired breeders) were purchased from Charles River Laboratories (Wilmington, MA). Inducible bitransgenic male mice that overexpress ΔFosB were generated from crosses of NSE-tTA (line A) and TetOP-ΔFosB (line A11) mice, and fully backcrossed to a C57BL/6J background, using a tetracycline-regulated gene expression system (55). ΔFosB mice were raised on water containing doxycycline (DOX) (100 μg/ml) (Sigma, St. Louis, Missouri), which represses transgene expression. Experiments were conducted ~8 weeks after DOX discontinuation, when transgene expression of ΔFosB is maximal (ΔFosB–ON group) (55). Half of the mice remained on DOX for the duration of the experiment to serve as controls (ΔFosB-Control group). Mice had free access to food and water and were maintained on a 12-h light/dark cycle. All procedures were conducted in accordance with National Institutes of Health and McLean Hospital policies. Ketamine was obtained from Sigma-Aldrich (St. Louis, MO), dissolved in 0.9% saline (Vehicle), and administered intraperitoneally (IP) at 10 ml/kg. Control mice received identical treatments as defeated mice.
Behavioral Manipulations and Tests
CSDS was performed as described previously (10,12). CD1 mice (residents) were screened for consistent aggressive behavior (attack latencies <30 sec for 3 consecutive screening tests). On each of 10 consecutive days, the intruder (defeated) mouse was placed in the homecage of a resident mouse and subjected to 10 min of social defeat stress. Following the defeat session, the mice were separated in the cage with a perforated Plexiglas divider, which allowed for continuous protected sensory exposure. Defeated mice were exposed to a new resident and cage each day. Control mice were handled daily and housed in identical cage set-ups as the defeated mice, but opposite a conspecific mouse. Separate cohorts were used for ICSS and social interaction (SI) experiments.
ICSS was performed as previously described (19,28). Briefly, mice (25–30 g) were implanted with monopolar electrodes aimed at the lateral hypothalamus (LH). Mice were trained with a descending series (or “pass”) of 15 stimulation frequency trials (0.05 log10 unit steps), 4 per day, at the minimum effective current. CSDS and control groups had equivalent minimum currents (~75 μA). ICSS thresholds (Theta-0) were calculated using a least-squares line of best-fit analysis (19,56). After stable baseline thresholds were established (+/− 15% for 5 consecutive days; BL1-5), mice were subjected to CSDS for 10 days (D1-10). Mice were initially separated into two groups to test whether the effects of CSDS on ICSS thresholds depend on the interval of time between the defeat session and ICSS testing: mice in the long interval (LInt) group were tested in ICSS ~16 hrs after defeat, whereas mice in the short interval (ShInt) group were tested in ICSS ~6 hrs after defeat (Figure 1A). Following CSDS, mice were returned to their home cages and tested post-defeat in ICSS for 5 days (P1-5). For ketamine experiments, mice received either vehicle or ketamine (20 mg/kg) 1 hr after the final defeat session.
Figure 1.

Chronic social defeat stress (CSDS) increases reward thresholds in the intracranial self-stimulation (ICSS) test. (A) Experimental design; LInt=long interval (16 hr) between a defeat bout and ICSS testing, ShInt=short interval (6 hr). (B) CSDS increases reward thresholds in the LInt (n=10) and ShInt (n=6) groups compared to controls by the second day of defeat. Controls in the LInt group (n=6) and ShInt group (n=3) were not significantly different and were pooled. Data are expressed as mean (± SEM) % change from the average baseline threshold. Gray background represents days in which mice were defeated (D1-10). (C) Average thresholds for baseline (BL1-5), defeat (D1-10), and post-defeat (P1-5) for each group expressed as % change from average baseline threshold (BL1-5). CSDS increases reward thresholds in the LInt and ShInt groups on D1-10 and P1-5 compared to within-group baseline threshold (BL1-5) and compared to controls (D) Rate-frequency functions for individual control and defeated representative mice demonstrate that CSDS causes parallel rightward shifts. *p<0.05, **p<0.01, ***p<0.001 for within-group comparisons to baseline (BL1-5); ^p<0.05, ^^p<0.01, ^^^p<0.001 for between-group comparisons to controls; #p<0.05 for comparisons between ShInt and LInt groups; ns=not significant.
To assess ketamine effects on SI, mice received vehicle, a low (2.5 mg/kg), or a high (20 mg/kg) dose of ketamine 24 hrs before the first day of CSDS (Day 0) or 1 hr after the final defeat session (Day 10). Mice were habituated to the interaction arena in red light for 15 min on days 8-10 of CSDS. Twenty-four hr after the final defeat session (Day 11), social approach behavior in the presence of an unfamiliar CD-1 enclosed in a wire cage was assessed as previously described (12, 57), with minor modifications. SI scores were defined as the amount of time the mouse spent near an enclosure containing a CD-1 (social target) over a 2.5 min period compared to when the target enclosure was empty. Because control mice spend more time interacting with a social target present, an SI score of 1 (equal time near social target versus empty enclosure) was used as a cut-off: SI scores >1 were considered “stress-resilient” and scores <1 were considered “stress-susceptible” (13). The segregation of defeated mice into susceptible and resilient subpopulations is supported by extensive behavioral, neurobiological, and electrophysiological analyses (13,54).
Passive-avoidance conditioning was conducted in a Gemini Avoidance System apparatus (San Diego Instruments, San Diego CA) as previously described (50) with minor modifications. During training, mice were given 1 min acclimation to the light compartment before access to the dark compartment. Once a cross was made to the dark compartment, mice were conditioned with two consecutive 2 sec (inescapable) footshocks (0.2 mA), followed by a 1 min time-out. Mice were given either vehicle or ketamine (20 mg/kg) 1 hr after conditioning. Step-through latencies were measured 24 hrs later. To assess the effects of our ketamine on anxiety-like behavior, a separate cohort of mice were given either vehicle or ketamine (20 mg/kg) 24 hrs prior to EPM testing. Mice were placed in the center of an elevated plus maze (each arm 33 cm long and 5 cm wide, with 2 opposite arms closed by 16.5 cm high walls, maze elevated 81 cm from floor) in red light and allowed to explore for 5 min.
Statistical Analysis
Two- and three-way repeated measures ANOVA was performed for CSDS, ICSS, and passive-avoidance data. Significant ANOVAs were further analyzed with Bonferroni post hoc tests. Effects of ketamine on SI were analyzed with preplanned contrasts (Bonferroni tests) between control and defeated mice within each treatment group, based on a specific a priori hypothesis that ketamine treatment would mitigate depressive-like behavior in defeated mice. Effects on EPM behavior were analyzed using Student’s t-test. The SI and EPM tests were videotaped and scored by raters blinded to treatment conditions.
RESULTS
The effects of social defeat on ICSS thresholds were evaluated after each episode of defeat, enabling us to track changes in responsiveness across the entire CSDS regimen (Figure 1A, B). LInt and ShInt data are presented together to facilitate a side-by-side comparison of interval duration effects on ICSS thresholds (Figure 1B, C). The effects of CSDS on ICSS thresholds depended on group [F(2,22)=13.53, p<0.001] and day [F(15,330)=2.98, p<0.001], with a marginal group X day interaction (p=0.054). CSDS significantly increased mean ICSS thresholds (expressed as % baseline) in both LInt and ShInt defeated mice compared to controls by the second day of defeat (LInt: p<0.05, ShInt: p<0.001). ShInt mice had higher thresholds compared to LInt mice only on P1 (p<0.05) (Figure 1B). A small proportion of mice were resilient to the effects of CSDS on anhedonia in ICSS (not shown), consistent with findings in other tests (13). Since there were no overall differences in ICSS thresholds between control mice in the LInt and ShInt groups (data not shown), these data were consolidated. When data are expressed as single means for BL1-5, D1-10, and P1-5 for each group, the effects of CSDS on ICSS thresholds depended on defeat [F(2,22)=9.68, p<0.01], day [F(2,44)=21.57, p<0.001], and a defeat X day interaction [F(4,44)=5.09, p<0.01] (Figure 1C). Within group comparisons revealed that ICSS thresholds were increased in both LInt and ShInt defeated mice on D1-10 and P1-5 compared to BL1-5 (p<0.001). Between group comparisons revealed that ICSS thresholds were significantly increased in LInt and ShInt defeated mice on D1-10 (p<0.001), and in defeated ShInt mice on P1-5 (p<0.01), compared to controls (Figure 1C). Raw data from individual representative control and defeated (LInt) mice illustrate how CSDS can cause a rightward shift in ICSS rate-frequency functions on D1-10 compared to BL1-5 (Figure 1D).
Because mice that inducibly overexpress ΔFosB in striatal regions are resilient to CSDS (54) and stress-like conditions (58), we hypothesized that ΔFosB-ON mice, but not ΔFosB-Control mice, would be resilient to the effects of CSDS on ICSS thresholds. When data are expressed as single means for BL1-5, D1-10, and P1-5 for each group, the effects of ΔFosB overexpression on CSDS-mediated changes in ICSS thresholds depended on DOX treatment [F(1,4)=13.25, p<0.05], day [F(2,8)=23.89, p<0.001], and a DOX treatment X day interaction [F(2,8)=16.40, p<0.01] (Figure 2). CSDS increased ICSS thresholds in ΔFosB-Control mice on D1-10 and P1-5 compared to BL1-5 (p<0.001) and on D1-10 and P1-5 compared to ΔFosB-ON mice (p<0.05), whereas ΔFosB-ON mice were resilient to the anhedonic effects of CSDS. ΔFosB overexpression had no effect on minimum currents, confirming that increased ΔFosB itself does not affect sensitivity to LH stimulation (58).
Figure 2.
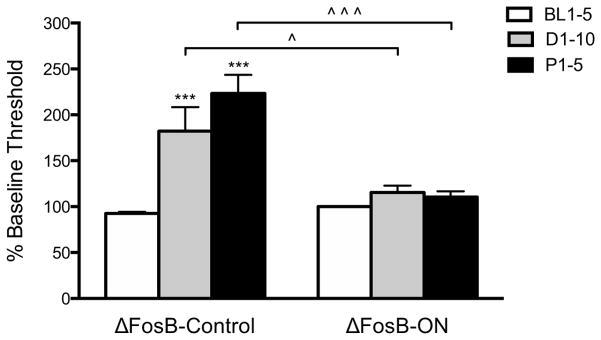
Inducible ΔFosB overexpression in striatum blocks the anhedonic effects of CSDS in the ICSS test. ΔFosB-Control mice (n=3), but not ΔFosB-ON mice (n=3) tested 8 weeks after DOX discontinuation, show increases in ICSS thresholds on D1-10 and P1-5 compared to BL1-5. ***p<0.001 for within-group comparisons to baseline (BL1-5); ^p<0.05, ^^^p<0.001 for between-group comparisons.
CSDS produced social avoidance in defeated mice treated with vehicle or a low dose of ketamine (2.5 mg/kg) (p’s<0.05) but not in defeated mice treated with a high dose of ketamine (20 mg/kg) (Figure 3A); there was a main effect of group [F(1,60)=15.75, p<0.001], but no main effect of dose or dose X group interaction. A plot of individual SI scores for defeated mice show that a small number of mice treated with vehicle (n=3/12) or a low dose of ketamine (n=2/10) exhibited resilience, whereas a larger proportion of mice treated with a high dose of ketamine (n=8/11) exhibited resilience (Figure 3A, inset). There were no significant differences in either distance traveled or velocity during the SI test (Figure 3B,C). Mice were retested 1 week later to determine if the antidepressant-like effects of the high dose of ketamine, as measured in the SI test, are persistent. Both defeated mice that received vehicle and defeated mice that received ketamine (20 kg/kg) exhibited social avoidance when retested 1 week later (vehicle: p<0.001, ketamine: p<0.05) (Figure 3D); there was a main effect of group [F(1,36)=21.10, p<0.0001], but no main effect of dose or dose X group interaction. A single administration of ketamine (20 mg/kg) 24 hrs prior to the first day of CSDS did not attenuate social avoidance. CSDS produced social avoidance in defeated mice treated previously with vehicle or a high dose of ketamine (20 mg/kg) (p’s<0.05) (Figure 3E); there was a main effect of group [F(1,21)=1.57, p<0.001], but no main effect of dose or dose X group interaction.
Figure 3.

Ketamine attenuates CSDS-induced social avoidance. (A) A high (20 mg/kg) but not low (2.5 mg/kg) dose of ketamine given 1 hr after the final defeat bout (Day 10) attenuates social avoidance in defeated mice. Inset: Individual SI scores in defeated mice from each dose group. (B–C) There were no effects of group (control, defeat) or treatment (vehicle, 20 mg/kg ketamine) on distance traveled (B) or velocity (C) during the SI test. (D) Mice in the vehicle and ketamine (20 mg/kg) groups exhibit comparable degrees of social avoidance compared to controls when retested 1 week later (Day 18). (E) Ketamine (20 mg/kg) administered 24 hrs before the first day of defeat does not attenuate social avoidance measured at the end of the CSDS procedure. *p<0.05 compared to controls within each dose, n=6–12 mice/group.
To determine if ketamine-effects on CSDS-induced social avoidance could be due to amnestic effects of this dosage, we compared passive avoidance memory retention in a separate cohort of vehicle and ketamine-treated mice. All mice exhibited increased step-through latencies on the memory retention test day [F(1,28)=22.82, p<0.0001] (Figure 4A). There was no main effect of treatment on retention of fear memory [F(1,28)=0.14, n.s.] or day X treatment interaction [F(1,28)=0.24, n.s.]. In the EPM, there were no differences between vehicle and ketamine-treated mice in open arm time (t(20)=0.61, n.s.) (Figure 4B) or number of open arm entries (t(20)=0.34, n.s.) (not shown).
Figure 4.
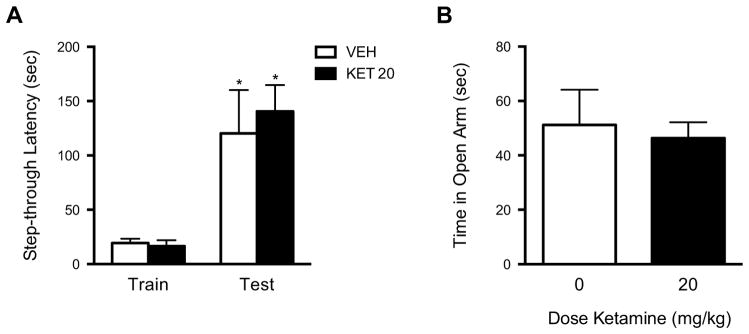
(A) Ketamine, at the effective dose (20 mg/kg) used in the SI test, does not interfere with fear memory retention in the passive avoidance test. All mice, regardless of treatment with vehicle or ketamine 1 hr after the training day, avoid entering the dark compartment on the testing day. (B) Ketamine treatment (20 mg/kg) 24 hrs prior to testing does not affect behavior in the elevated plus maze (EPM). *p<0.05 for within-group comparisons, n=8–11 mice/group.
Mice received ketamine or vehicle after the final defeat session, and continued ICSS testing post-treatment (D10, P1-5). ICSS thresholds obtained on pre-treatment days (D1-9) were compiled for each group. Thresholds on D10 and P1-5 were analyzed to assess the temporal effects of ketamine. The same dose of ketamine (20 mg/kg) that attenuated social avoidance in defeated mice in the SI test did not attenuate anhedonia in the ICSS test (Figure 5); there was a main effect of group [F(1,45)=48.65, p<0.0001], but no significant group X treatment X day interaction. ICSS electrode placements were indistinguishable from those depicted previously (28).
Figure 5.
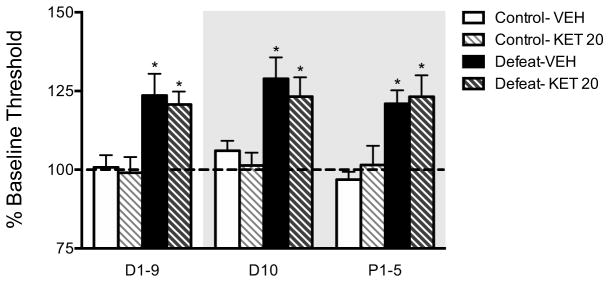
Ketamine (20 mg/kg) administered 1 hr after the final defeat bout does not block the anhedonic effects of CSDS in the ICSS test on D10 or P1-5. Gray background represents the thresholds obtained post-treatment. Mice were defeated for 10 days (LInt schedule) and matched into saline and ketamine groups. Ketamine had no effect in control mice. *p<0.05 for between-treatment group comparisons to controls, n=4–6 mice/group.
DISCUSSION
CSDS produces anhedonia in the ICSS paradigm in mice. Specifically, we show that that CSDS decreases the rewarding impact of LH stimulation, as measured by elevations in ICSS thresholds (19), with effects persisting up to 5 days post-CSDS. These results are broadly consistent with the results of previous studies in rats (59) and hamsters (60), which utilized other methodologies to quantify brain stimulation reward strength. As expected on the basis of previous work (54), which demonstrated that mice that overexpress ΔFosB are resilient to the development of social avoidance and sucrose preference deficits, we found that mice were also resilient to the anhedonia-inducing effects of CSDS in the ICSS test. In addition, we examined the antidepressant-like properties of ketamine in the CSDS paradigm. Acute ketamine treatment in defeated mice attenuated social avoidance in the SI test. This acute effect is remarkable considering similar effects are observed only after chronic treatment with fluoxetine or imipramine (12,16). However, acute ketamine had no effect on anhedonia in the ICSS test. Our findings complement previous work showing that CSDS decreases preference for natural rewards (12,13,15), and indicate that ICSS is a sensitive, reliable, and quantifiable method for detecting anhedonic effects of CSDS over time in mice. The results also demonstrate that ketamine has rapid antidepressant-like effects on some—but not all—CSDS-induced behavioral abnormalities.
In the CSDS paradigm, susceptible mice show deficits in sucrose preference that are often interpreted as anhedonia. However, numerous factors complicate the use of sucrose tests as a measure of anhedonia. First, they lack reliability: CSDS has been reported to decrease (14–18), increase (61,62) or have no effect (63,64) on sucrose preference, with similar discrepancies reported using the chronic mild stress paradigm (65,66). Second, sucrose tests may be confounded by the novelty of the sucrose solution, since chronic stressors can cause neophobia (67,68). Finally, their translational relevance is unclear, since there are no differences in preference for sweet solutions between individuals with major depressive disorder and healthy controls (69,70), and depression and chronic stress can result in both weight loss or gain (8,71–73). These factors suggest that sucrose preference and consumption tests alone may not represent a face-valid approach for assessing anhedonia. Although our ICSS studies do not address whether these sucrose-related tests reflect reward function, they confirm that CSDS indeed produces anhedonia.
One advantage of the ICSS test is that it enables day-to-day measurements of reward function, enabling precise analysis of the amount of CSDS required to produce anhedonia. Repeated tests are not feasible in SI or sucrose preference tests, since experience can affect the results. Here we demonstrate that CSDS begins to significantly elevate ICSS thresholds by the second defeat session. Thresholds remained elevated throughout the CSDS period and for up to one week following its termination, demonstrating persistent anhedonia. These threshold elevations are consistent with the effects of other pro-depressive treatments including chronic unpredictable stress (26,27), drug withdrawal (22–25), and kappa-opioid receptor agonists (29,58). Although thresholds in control mice remained stable during D1-10, the nominal elevations during P1-5 may be due to an idiosyncracy of the experimental design. Specifically, control mice were housed across a divider from a conspecific during D1-10, but (like all subjects) were socially isolated during P1-5. It is established that social isolation of adult rodents produces anhedonia-related signs (74).
Importantly, we demonstrate that it is possible to mitigate CSDS-induced anhedonia in the ICSS test. Mice that overexpress ΔFosB in D1-type medium spiny neurons of striatum (55) are less sensitive to CSDS-induced increases in ICSS thresholds. This is consistent with previous reports that they are also less sensitive to CSDS-induced social avoidance (54) and to the threshold increasing effects of U50488, a kappa-opioid receptor agonist known to produce dysphoria (58). The ability of ΔFosB overexpression to block the anhedonic effects of both CSDS and KOR agonists suggests potential overlap in molecular mechanisms, as proposed previously (75,76). One possible mechanism by which ΔFosB may mediate stress-resilience is via induction of GluR2 in the nucleus accumbens (NAc), which dampens glutamatergic tone (54). Although ΔFosB-Control mice show larger increases in ICSS thresholds compared to those seen in C57BL/6J mice, this may be due to different background of the founder strains; indeed ΔFosB-Control mice are also more sensitive to CSDS-induced social avoidance (54).
It is possible that the acute painful effects of CSDS contribute to elevations in ICSS thresholds, considering that acute pain can increase ICSS thresholds (77). It seems unlikely that the observed ICSS threshold increases are due to pain alone, however, for several reasons. First, exposure to stressful stimuli including CSDS often produces stress-induced analgesia (75,78). Second, ICSS thresholds remained elevated after the termination of defeat, indicating that the effects of CSDS are prolonged and independent of acute pain. Third, mice that “witness” social defeat sessions exhibit depressive-like behaviors despite the lack of physical contact (79). Finally, CSDS failed to produce anhedonia in mice that overexpress ΔFosB in striatum, where it mediates stress resilience (54). The present results suggest that CSDS produces adaptations within brain reward pathways (12,13,15,54) that result in the anhedonic phenotype.
Although standard antidepressants can reverse the behavioral effects of CSDS, chronic treatment is required (12,16). Because acute ketamine has rapid antidepressant effects in humans (31–39), we assessed whether acute ketamine treatment reverses the effects of CSDS in mice. We found that a single dose of ketamine (20 mg/kg, IP) causes rapid (~24 hours) antidepressant-like effects in the SI test following exposure to CSDS. This is consistent with numerous studies describing the rapid effects (within hours) of subanesthetic doses of ketamine in humans (31–39), even in patients with treatment-resistant depression (32,34,38,39). Ketamine also has rapid antidepressant-like effects in other animal models including the forced swim test (FST) and tail suspension test (TST) (40,41,42,43,44,48,50), learned helplessness paradigm (45,50,51), and chronic mild stress paradigm (46,47,49). Neither CSDS nor acute ketamine affected locomotor activity, an important factor to rule out non-specific effects on gross motor output that can complicate data interpretation. Considering that ketamine produces amnestic effects in rodents (52,53), we assessed the possibility that ketamine interferes with stress-related learning and memory. Consistent with previous work (50), we found that 20 mg/kg ketamine does not interfere with performance in the passive avoidance test, suggesting that the antidepressant-like effects of the drug in our studies are not due to memory impairment. We further show that the acute ketamine treatment that reduces social avoidance does not affect behavior in the EPM, although previous reports are inconclusive (42,43). Together, our findings suggest that the effects of ketamine in the SI test can be differentiated from effects on learning and memory or anxiety.
Interestingly, a single injection of 20 mg/kg ketamine 24 hr prior to day one of CSDS had no effect on social avoidance, suggesting an inability to prevent the development of repeated stress-induced adaptations, at least with this single dose. Additionally, the antidepressant-like effects of ketamine in the SI test were not persistent: mice re-tested 1 week later did not show continued attenuation of CSDS-induced social avoidance. This finding is broadly consistent with clinical studies reporting that patients who respond to acute ketamine often experience relapse within several days (31,39,80) and with preclinical reports of acute but not persistent antidepressant-like effects of ketamine in the FST and TST in mice (41,51,81), although there are also reports of persistent effects (45,49,50,82). These differences may be due to the intensity or type of stress (e.g. social stress versus FST or TST) or strain differences (41). Repeated ketamine treatment may be necessary to induce reliably persistent effects. It was recently reported that repeated ketamine treatment in rats was necessary to produce a long-lasting resilient phenotype in the chronic unpredictable stress paradigm (81). Furthermore, there is evidence that chronic ketamine administration induces antidepressant-like effects at doses that are not acutely effective (83). There are reports that repeated infusions in patients with treatment-resistant depression may result in a more sustained antidepressant response compared to a single infusion (84–86). However, the widespread use of ketamine as a therapeutic may be limited by abuse liability and other side effects (87–89). Characterizing the time-course of ketamine effects in animal models may help optimize clinical treatment regimens.
Despite its effects on social avoidance, acute ketamine failed to block the anhedonic effects of CSDS in the ICSS test. This was unexpected considering prior reports that ketamine reverses stress-related decreases in sucrose preference (45,48), and increases dopamine efflux in the NAc (90), an effect often associated with elevated mood (91). To the best of our knowledge there are no reports on the ability of standard antidepressants to reverse the effects of CSDS on ICSS. We cannot exclude the possibility that ketamine treatment might reverse CSDS-induced anhedonia in the ICSS test using other treatment regimens (e.g., different doses, repeated administration) or mouse strains, although such studies are beyond the scope of this report considering the number of possible permutations in experimental design. However, animal studies involving repeated ketamine administration may be of limited value in understanding why the drug is clinically effective after acute administration in people (31).
CSDS also induces robust anxiety-like behaviors as measured in exploratory assays such as the elevated plus maze or open field (12,13). However, an important feature of the CSDS paradigm is that it can distinguish these anxiety-like measures from more depressive-like signs such as social avoidance or sucrose preference deficits. Specifically, social avoidance and sucrose preference deficits are treated effectively with standard antidepressants but not anxiolytics (12). Moreover, mice that display a resilient phenotype (i.e., those that lack social avoidance and sucrose preference deficits) display equivalent levels of anxiety-like behavior (15), as seen in mice that overexpress ΔFosB (54). Similarly, we show that acute ketamine (20 mg/kg) affected one dimension of depressive behavior (social avoidance), but not others (anhedonia, anxiety, learning and memory), suggesting that these behaviors represent distinct domains regulated by separate, yet overlapping circuits in the brain (92). The molecular mechanisms by which ketamine produces these antidepressant-like effects are not well understood, but may involve alterations in glutamate function or protein synthesis within brain regions including hippocampus and frontal cortex (45,46,47,50).
Acknowledgments
This research was funded by grants from the National Institutes of Health (MH063266 to WC; R01MH51399 and P50MH096890 to EJN; MH090264 to SJR). RJD was supported by a Training Grant (T32MH020017) from the National Institute of Health.
Footnotes
FINANCIAL DISCLOSURES
The authors report no biomedical financial interests or potential conflicts of interest.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Finlay-Jones R, Brown GW. Types of stressful life event and the onset of anxiety and depressive disorders. Psychol Med. 1981;11:803–15. doi: 10.1017/s0033291700041301. [DOI] [PubMed] [Google Scholar]
- 2.Keller MC, Neale MC, Kendler KS. Association of different adverse life events with distinct patterns of depressive symptoms. Am J Psychiatry. 2007;164:1521–1529. doi: 10.1176/appi.ajp.2007.06091564. [DOI] [PubMed] [Google Scholar]
- 3.Kendler KS, Karkowski LM, Prescott CA. Causal relationship between stressful life events and the onset of major depression. Am J Psychiatry. 1999;156:837–48. doi: 10.1176/ajp.156.6.837. [DOI] [PubMed] [Google Scholar]
- 4.Kessler RC, Berglund P, Demler O, Jim R, Merikangas KR, Walters EE. Lifetime Prevalence and Age-of-Onset Distributions of DSM-IV Disorders in the National Comorbidity Survey Replication. Arch Gen Psychiatry. 2005;62:593–602. doi: 10.1001/archpsyc.62.6.593. [DOI] [PubMed] [Google Scholar]
- 5.Keller MB, Boland RJ. Implications of failing to achieve successful long-term maintenance treatment of recurrent unipolar major depression. Biol Psychiatry. 1998;44:348–360. doi: 10.1016/s0006-3223(98)00110-3. [DOI] [PubMed] [Google Scholar]
- 6.Nemeroff CB. Prevalence and management of treatment-resistant depression. J Clin Psychiatry. 2007;68:17–25. [PubMed] [Google Scholar]
- 7.Krishnan V, Nestler EJ. The molecular neurobiology of depression. Nature. 2008;455:894–902. doi: 10.1038/nature07455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 5. Washington, DC: American Psychiatric Association; 2000. [Google Scholar]
- 9.Nestler EJ, Hyman SE. Animal models of neuropsychiatric disorders. Nat Neurosci. 2010;13:1161–1169. doi: 10.1038/nn.2647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Golden SA, Covington HE, III, Berton O, Russo SJ. A standardized protocol for repeated social defeat stress in mice. Nat Protoc. 2011;6:1183–1191. doi: 10.1038/nprot.2011.361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Huhman KL. Social conflict models: can they inform us about human psychopathology? Horm Behav. 2006;50:640–646. doi: 10.1016/j.yhbeh.2006.06.022. [DOI] [PubMed] [Google Scholar]
- 12.Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science. 2006;311:864–868. doi: 10.1126/science.1120972. [DOI] [PubMed] [Google Scholar]
- 13.Krishnan V, Han MH, Graham DL, Berton O, Renthal W, Russo SJ, et al. Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions. Cell. 2007;131:391–404. doi: 10.1016/j.cell.2007.09.018. [DOI] [PubMed] [Google Scholar]
- 14.Kudryavtseva NN, Bakshtanovskaya IV, Koryakina LA. Social model of depression in mice of C57BL/6J strain. Pharmacol Biochem Behav. 1991;38:315–320. doi: 10.1016/0091-3057(91)90284-9. [DOI] [PubMed] [Google Scholar]
- 15.Cao JL, Covington HE, III, Friedman AK, Wilkinson MB, Walsh JJ, Cooper DC, et al. Mesolimbic dopamine neurons in the brain reward circuit mediate susceptibility to social defeat and antidepressant action. J Neurosci. 2010;30:16453–16458. doi: 10.1523/JNEUROSCI.3177-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Covington HE, III, Maze I, LaPlant QC, Vialou VF, Yoshinori ON, Berton O, et al. Antidepressant actions of histone deacetylase inhibitors. J Neurosci. 2009;29:11451–11461. doi: 10.1523/JNEUROSCI.1758-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chaouloff F. Social stress models in depression research: what do they tell us? Cell Tissue Res. 2013:1–12. doi: 10.1007/s00441-013-1606-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kalueff AV, Avgustinovich DF, Kudryavtseva NN, Murphy DL. BDNF in anxiety and depression. Science. 2006;312:1598–1599. doi: 10.1126/science.312.5780.1598. [DOI] [PubMed] [Google Scholar]
- 19.Carlezon WA, Jr, Chartoff EH. Intracranial self-stimulation (ICSS) in rodents to study the neurobiology of motivation. Nat Protoc. 2007;2:2987–2995. doi: 10.1038/nprot.2007.441. [DOI] [PubMed] [Google Scholar]
- 20.Der-Avakian A, Markou A. The neurobiology of anhedonia and other reward-related deficits. Trends Neurosci. 2012;35:68–77. doi: 10.1016/j.tins.2011.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barr AM, Markou A, Phillips AG. A ‘crash’ course on psychostimulant withdrawal as a model of depression. Trends Pharmacol Sci. 2003;23:475–482. doi: 10.1016/s0165-6147(02)02086-2. [DOI] [PubMed] [Google Scholar]
- 22.Epping-Jordan MP, Watkins SS, Koob GF, Markou A. Dramatic decreases in brain reward function during nicotine withdrawal. Nature. 1998;393:76–79. doi: 10.1038/30001. [DOI] [PubMed] [Google Scholar]
- 23.Johnson PM, Hollander JA, Kenny PJ. Decreased brain reward function during nicotine withdrawal in C57BL6 mice: Evidence from intracranial self-stimulation (ICSS) studies. Pharmacol Biochem Behav. 2008;90:409–415. doi: 10.1016/j.pbb.2008.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Liu J, Schulteis G. Brain reward deficits accompany naloxone-precipitated withdrawal from acute opioid dependence. Pharmacol Biochem Behav. 2004;79:101–108. doi: 10.1016/j.pbb.2004.06.006. [DOI] [PubMed] [Google Scholar]
- 25.Markou A, Hauger RL, Koob GF. Desmethylimipramine attenuates cocaine withdrawal in rats. Psychopharmacology (Berl) 1992;109:305–314. doi: 10.1007/BF02245878. [DOI] [PubMed] [Google Scholar]
- 26.Lin D, Bruijnzeel AW, Schmidt P, Markou A. Exposure to chronic mild stress alters thresholds for lateral hypothalamic stimulation reward and subsequent responsiveness to amphetamine. Neuroscience. 2002;114:925–933. doi: 10.1016/s0306-4522(02)00366-4. [DOI] [PubMed] [Google Scholar]
- 27.Moreau JL, Jenck F, Martin JR, Mortas P, Haefely WE. Antidepressant treatment prevents chronic unpredictable mild stress-induced anhedonia as assessed by ventral tegmentum self-stimulation behavior in rats. Eur J Neuropsychopharmacol. 1992;2:43–49. doi: 10.1016/0924-977x(92)90035-7. [DOI] [PubMed] [Google Scholar]
- 28.DiNieri JA, Nemeth CL, Parsegian A, Carle T, Gurevich VV, Gurevich E, et al. Altered sensitivity to rewarding and aversive drugs in mice with inducible disruption of cAMP response element-binding protein function within the nucleus accumbens. J Neurosci. 2006;29:1855–1859. doi: 10.1523/JNEUROSCI.5104-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Todtenkopf MS, Marcus JF, Portoghese PS, Carlezon WA., Jr Effects of kappa-opioid ligands on intracranial self-stimulation in rats. Psychopharmacology (Berl) 2004;172:463–470. doi: 10.1007/s00213-003-1680-y. [DOI] [PubMed] [Google Scholar]
- 30.Martin D, Lodge D. Ketamine acts as a non-competitive N-methyl-d-aspartate antagonist on frog spinal cord in vitro. Neuropharmacology. 1985;24:999–1003. doi: 10.1016/0028-3908(85)90128-5. [DOI] [PubMed] [Google Scholar]
- 31.Berman RM, Cappiello A, Anand A, Oren DA, Heninger GR, Charney DS, et al. Antidepressant effects of ketamine in depressed patients. Biol Psychiatry. 2000;47:351–354. doi: 10.1016/s0006-3223(99)00230-9. [DOI] [PubMed] [Google Scholar]
- 32.Carlson PJ, Diazgranados N, Nugent AC, Ibrahim L, Luckenbaugh DA, Brutsche N, et al. Neural correlates of rapid antidepressant response to ketamine in treatment-resistant unipolar depression: A preliminary positron emission tomography study. Biol Psychiatry. 2013;73:1213–1221. doi: 10.1016/j.biopsych.2013.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Correll GE, Futter GE. Two case studies of patients with major depressive disorder given low-dose (subanesthetic) ketamine infusions. Pain Med. 2006;7:92–95. doi: 10.1111/j.1526-4637.2006.00101.x. [DOI] [PubMed] [Google Scholar]
- 34.Diazgranados N, Ibrahim L, Brutsche NE, Newberg A, Kronstein P, Khalife S, et al. A randomized add-on trial of an N-methyl-D-aspartate antagonist in treatment-resistant bipolar depression. Arch Gen Psychiatry. 2010;67:793–802. doi: 10.1001/archgenpsychiatry.2010.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Larkin GL, Beautrais A preliminary naturalistic study of low-dose ketamine for depression and suicide ideation in the emergency department. Int J Neuropsychopharmacol. 2011;14:1127–1131. doi: 10.1017/S1461145711000629. [DOI] [PubMed] [Google Scholar]
- 36.Phelps LE, Brutsche N, Moral JR, Luckenbaugh DA, Manji HK, Zarate CA., Jr Family history of alcohol dependence and initial antidepressant response to an N-methyl-D-aspartate antagonist. Biol Psychiatry. 2009;65:181–184. doi: 10.1016/j.biopsych.2008.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Salvadore G, Cornwell BR, Sambataro F, Latov D, Colon-Rosario V, Carver F, et al. Anterior cingulated desynchronization and functional connectivity with the amygdala during a working memory task predict rapid antidepressant response to ketamine. Neuropsychopharmacology. 2010;35:1415–1422. doi: 10.1038/npp.2010.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Price RB, Nock MK, Charney DS, Mathew SJ. Effects of intravenous ketamine on explicit and implicit measures of suicidality in treatment-resistant depression. Biol Psychiatry. 2009;66:522–526. doi: 10.1016/j.biopsych.2009.04.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zarate CA, Jr, Singh JB, Carlson PJ, Brutsche NE, Ameli R, Luckenbaugh DA, et al. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch Gen Psychiatry. 2006;63:856–864. doi: 10.1001/archpsyc.63.8.856. [DOI] [PubMed] [Google Scholar]
- 40.Autry AE, Adachi M, Nosyreva E, Na ES, Los MF, Cheng PF, et al. NMDA receptor blockade at rest triggers rapid behavioral antidepressant responses. Nature. 2011;475:91–95. doi: 10.1038/nature10130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bechtholt-Gompf AJ, Smith KL, John CS, Kang HH, Carlezon WA, Jr, Cohen BM, et al. CD-1 and Balb/cJ mice do not show enduring antidepressant-like effects of ketamine in tests of acute antidepressant efficacy. Psychopharmacology. 2011;215:689–695. doi: 10.1007/s00213-011-2169-8. [DOI] [PubMed] [Google Scholar]
- 42.Da Silva FC, do Carmo de Oliveira Cito M, da Silva MI, Moura BA, de Aquino Neto MR, Feitosa ML, et al. Behavioral alterations and pro-oxidant effect of a single ketamine administration to mice. Brain Res Bull. 2010;83:9–15. doi: 10.1016/j.brainresbull.2010.05.011. [DOI] [PubMed] [Google Scholar]
- 43.Engin E, Treit D, Dickson CT. Anxiolytic- and antidepressant-like properties of ketamine in behavioral and neurophysiological animal models. Neuroscience. 2009;161:359–369. doi: 10.1016/j.neuroscience.2009.03.038. [DOI] [PubMed] [Google Scholar]
- 44.Garcia LS, Comim CM, Valvassori SS, Réus GZ, Barbosa LM, Andreazza AC, et al. Acute administration of ketamine induces antidepressant-like effects in the forced swimming test and increases BDNF levels in the rat hippocampus. Prog Neuropsychopharmacol Biol Psych. 2008;32:140–144. doi: 10.1016/j.pnpbp.2007.07.027. [DOI] [PubMed] [Google Scholar]
- 45.Koike H, Iijima M, Chaki S. Involvement of AMPA receptor in both the rapid and sustained antidepressant-like effects of ketamine in animal models of depression. Behav Brain Res. 2011;224:107–111. doi: 10.1016/j.bbr.2011.05.035. [DOI] [PubMed] [Google Scholar]
- 46.Li N, Liu RJ, Dwyer JM, Banasr M, Lee B, Son H, et al. Glutamate N-methyl-D-aspartate receptor antagonists rapidly reverse behavioral and synaptic deficits caused by chronic stress exposure. Biol Psychiatry. 2011;69:754–761. doi: 10.1016/j.biopsych.2010.12.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Li N, Lee B, Lui RJ, Banasr M, Dwyer JM, Iwata M, et al. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science. 2010;329:959–964. doi: 10.1126/science.1190287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lindholm JSO, Autio H, Vesa L, Antila H, Lindemann L, Hoener MC, et al. The antidepressant-like effects of glutamatergic drugs ketamine and AMPA receptor potentiators LY 451646 are preserved in bdnf (+/−) heterozygous null mice. Neuropharmacology. 2012;62:391–397. doi: 10.1016/j.neuropharm.2011.08.015. [DOI] [PubMed] [Google Scholar]
- 49.Ma XC, Dang YH, Jia M, Ma R, Wang F, Wu J, et al. Long-lasting antidepressant action of ketamine, but not glycogen synthase kinase-3 inhibitor SB216763, in the chronic mild stress model of mice. PLoS One. 2013;8:e56053. doi: 10.1371/journal.pone.0056053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Maeng S, Zarate CA, Jr, Du J, Schloesser RJ, McCammon J, Chen G, et al. Cellular mechanisms underlying the antidepressant effects of ketamine: Role of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptors. Biol Psychiatry. 2008;63:349–352. doi: 10.1016/j.biopsych.2007.05.028. [DOI] [PubMed] [Google Scholar]
- 51.Popik P, Kos T, Sowa-Kucma M, Nowak G. Lack of persistent effects of ketamine in rodent models of depression. Psychopharmacology (Berl) 2008;198:421–430. doi: 10.1007/s00213-008-1158-z. [DOI] [PubMed] [Google Scholar]
- 52.Boultadakis A, Pitsikas N. Anesthetic ketamine impairs rats’ recall of previous information: the nitric oxide synthase inhibitor N-nitro-L-arginine methylester antagonizes this ketamine-induced recognition memory deficit. Anesthesiology. 2011;114:1345–1353. doi: 10.1097/ALN.0b013e318219524e. [DOI] [PubMed] [Google Scholar]
- 53.Wang JH, Fu Y, Wilson FAW, Ma YY. Ketamine affects memory consolidation: differential effects in T-maze and passive avoidance paradigms in mice. Neuroscience. 2006;140:993–1002. doi: 10.1016/j.neuroscience.2006.02.062. [DOI] [PubMed] [Google Scholar]
- 54.Vialou V, Robison AJ, LaPlant QC, Covington HE, Dietz DM, Ohnishi YN, et al. ΔFosB in brain reward circuits mediates resilience to stress and antidepressant responses. Nat Neurosci. 2010;13:745–752. doi: 10.1038/nn.2551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kelz MB, Chen J, Carlezon WA, Whisler K, Gilden L, Beckmann AM, et al. Expression of the transcription factor ΔFosB in the brain controls sensitivity to cocaine. Nature. 1999;401:272–276. doi: 10.1038/45790. [DOI] [PubMed] [Google Scholar]
- 56.Miliaressis E, Rompre PP, Labiolette P, Philippe L, Coulombe D. The curve-shift paradigm in self-stimulation methodology. Physiol Behav. 1986;37:85–91. doi: 10.1016/0031-9384(86)90388-4. [DOI] [PubMed] [Google Scholar]
- 57.Bruchas MR, Schindler AG, Shankar H, Messinger DI, Miyatake M, Land BB, et al. Selective p383 MAPK deletion in serotonergic neurons produces stress resilience in models of depression and addiction. Neuron. 2011;71:498–511. doi: 10.1016/j.neuron.2011.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Muschamp JW, Nemeth CL, Robison AJ, Nestler EJ, Carlezon WA., Jr ΔFosB enhances the rewarding effects of cocaine while reducing the pro-depressive effects of the kappa-opioid receptor agonist U50488. Biol Psychiatry. 2012;71:44–50. doi: 10.1016/j.biopsych.2011.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Der-Avakian A, Markou A. Neonatal maternal separation exacerbates the reward-enhancing effect of acute amphetamine administration and the anhedonic effect of repeated social defeat in adult rats. Neuroscience. 2010;170:1189–1198. doi: 10.1016/j.neuroscience.2010.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kureta Y, Watanabe S. Influence of social dominance on self-stimulation behavior in male golden hamsters. Physiol Behav. 1996;59:621–624. doi: 10.1016/0031-9384(95)02123-x. [DOI] [PubMed] [Google Scholar]
- 61.Dubreucq S, Kambire S, Conforzi M, Metna-Laurent M, Cannich A, Soria-Gomez E, et al. Cannabinoid type 1 receptors located on single-minded 1-expressing neurons control emotional behaviours. Neuroscience. 2012;204:230–244. doi: 10.1016/j.neuroscience.2011.08.049. [DOI] [PubMed] [Google Scholar]
- 62.Dubreucq S, Matias I, Cardinal P, Häring M, Lutz B, Marsicano G, et al. Genetic dissection of the role of cannabinoid type-1 receptors in the emotional consequences of repeated social stress in mice. Neuropsychopharmacology. 2012;37:1885–1900. doi: 10.1038/npp.2012.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Croft AP, Brooks SP, Cole J, Little HJ. Social defeat increases alcohol preference of C57BL/10 strain mice; effect prevented by a CCKB antagonist. Psychopharmacology. 2005;183:163–170. doi: 10.1007/s00213-005-0165-6. [DOI] [PubMed] [Google Scholar]
- 64.Hollis F, Wang H, Dietz D, Gunjan A, Kabbaj M. The effects of repeated social defeat on long-term depressive-like behaviour and short-term histone modifications in the hippocampus in male Sprague-Dawley rats. Psychopharmacology. 2010;211:69–77. doi: 10.1007/s00213-010-1869-9. [DOI] [PubMed] [Google Scholar]
- 65.Moreau JL. Reliable monitoring of hedonic deficits in the chronic mild stress model of depression. Psychopharmacology. 1997;134:357–358. doi: 10.1007/s002130050467. [DOI] [PubMed] [Google Scholar]
- 66.Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology. 2005;52:90–110. doi: 10.1159/000087097. [DOI] [PubMed] [Google Scholar]
- 67.Bondar NP, Kovalenko IL, Avgustinovich DF, Smagin DA, Kudryavtseva NN. Anhedonia in the shadow of chronic social defeat stress, or when the experimental context matters. Open Behav Sci J. 2009;3:17–27. [Google Scholar]
- 68.File SE. Factors controlling measures of anxiety and responses to novelty in the mouse. Behav Brain Res. 2013;125:151–157. doi: 10.1016/s0166-4328(01)00292-3. [DOI] [PubMed] [Google Scholar]
- 69.Berlin I, Givry-Steiner L, Lecrubier Y, Puech AJ. Measures of anhedonia and hedonic responses to sucrose in depressive and schizophrenic patients in comparison with healthy subjects. Eur Psychiatry. 1998;13:303–309. doi: 10.1016/S0924-9338(98)80048-5. [DOI] [PubMed] [Google Scholar]
- 70.Dichter GS, Smoski MJ, Kampov Polevoy AB, Gallop R, Garbutt JC. Unipolar depression does not moderate responses to the Sweet Taste Test. Depress Anxiety. 2010;27:859–863. doi: 10.1002/da.20690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Dallman MF, Pecoraro N, Akana SF, La Fleur SE, Gomez F, Houshyar H, et al. Chronic stress and obesity: a new view of “comfort food”. Proc Natl Acad Sci USA. 2003;100:11696–11701. doi: 10.1073/pnas.1934666100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Dallman MF, Pecoraro NC, La Fleur SE. Chronic stress and comfort foods: self-medication and abdominal obesity. Brain Behav Immun. 2005;19:275–280. doi: 10.1016/j.bbi.2004.11.004. [DOI] [PubMed] [Google Scholar]
- 73.Heiskanena TH, Koivumaa-Honkanena HT, Niskanenb LK, Lehtoa SM, Honkalampie KM. Depression and major weight gain: A 6-year prospective follow-up of outpatients. Compr Psychiatry. 2013 doi: 10.1016/j.comppsych.2013.02.001. [DOI] [PubMed] [Google Scholar]
- 74.Wallace DL, Han MH, Graham DL, Green TA, Vialou V, Iñiguez SD, et al. CREB regulation of nucleus accumbens excitability mediates social isolation–induced behavioral deficits. Nat Neurosci. 2009;12:200–209. doi: 10.1038/nn.2257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.McLaughlin JP, Li S, Valdez J, Chavkin TA, Chavkin C. Social defeat stress-induced behavioral responses are mediated by the endogenous kappa opioid system. Neuropsychopharmacology. 2005;31:1241–1248. doi: 10.1038/sj.npp.1300872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Van’t Veer A, Carlezon WA., Jr Role of kappa-opioid receptors in stress and anxiety-related behavior. Psychopharmacology. 2013;229:435–452. doi: 10.1007/s00213-013-3195-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Pereira Do Carmo G, Stevenson GW, Carlezon WA, Negus SS. Effects of pain-and analgesia-related manipulations on intracranial self-stimulation in rats: further studies on pain-depressed behavior. Pain. 2009;144:170–177. doi: 10.1016/j.pain.2009.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Butler RK, Finn DP. Stress-induced analgesia. Prog Neurobiol. 2009;88:184–202. doi: 10.1016/j.pneurobio.2009.04.003. [DOI] [PubMed] [Google Scholar]
- 79.Warren BL, Vialou VF, Iñiguez SD, Alcantara LF, Wright KN, Feng J, et al. Neurobiological sequelae of witnessing stressful events in adult mice. Biol Psychiatry. 2013;73:7–14. doi: 10.1016/j.biopsych.2012.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ibrahim L, Diazgranados N, Franco-Chaves J, Brutsche N, Henter ID, Kronstein P, et al. Course of improvement in depressive symptoms to a single intravenous infusion of ketamine vs add-on riluzole: Results from a 4-week, double-blind, placebo-controlled study. Neuropsychopharmacology. 2012;37:1526–1533. doi: 10.1038/npp.2011.338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Parise EM, Alcantara LF, Warren Bl, Wright KN, Hadad R, Sial OK, et al. Repeated ketamine exposure induces an enduring resilient phenotype in adolescent and adult rats. Biol Psychiatry. 2013;74:750–759. doi: 10.1016/j.biopsych.2013.04.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Yilmaz A, Schulz D, Aksoy A, Canbeyli R. Prolonged effect of an anesthetic dose of ketamine on behavioral despair. Pharmacol Biochem Behav. 2002;71:341–344. doi: 10.1016/s0091-3057(01)00693-1. [DOI] [PubMed] [Google Scholar]
- 83.Garcia LS, Comim CM, Valvassori SS, Réus GZ, Andreazza AC, Stertz L, et al. Chronic administration of ketamine elicits antidepressant-like effects in rats without affecting hippocampal brain-derived neurotrophic factor protein levels. Basic Clin Pharmacol Toxicol. 2008;103:502–506. doi: 10.1111/j.1742-7843.2008.00210.x. [DOI] [PubMed] [Google Scholar]
- 84.aan het Rot M, Collins KA, Murrough JW, Perez AM, Reich DL, Charney DS, et al. Safety and efficacy of repeated-dose intravenous ketamine for treatment-resistant depression. Biol Psychiatry. 2010;67:139–145. doi: 10.1016/j.biopsych.2009.08.038. [DOI] [PubMed] [Google Scholar]
- 85.Murrough JW, Perez AM, Pillemer S, Stern J, Parides MK, aan het Rot M, et al. Rapid and longer-term antidepressant effects of repeated ketamine infusions in treatment-resistant major depression. Biol Psychiatry. 2012;74:250–6. doi: 10.1016/j.biopsych.2012.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Rasmussen KG, Lineberry TW, Galardy CW, Kung S, Lapid MI, Palmer BA, et al. Serial infusions of low-dose ketamine for major depression. J Psychopharmacol. 2013;27:444–450. doi: 10.1177/0269881113478283. [DOI] [PubMed] [Google Scholar]
- 87.de la Peña IJ, Lee HL, de la Peña I, Shin CY, Sohn AR, et al. Pre-exposure to related substances induced place preference and self-administration of the NMDA receptor antagonist-benzodiazepine combination, zoletil. Behav Pharmacol. 2013;24:20–28. doi: 10.1097/FBP.0b013e32835cf442. [DOI] [PubMed] [Google Scholar]
- 88.Herberg LJ, Rose IC. The effect of MK-801 and other antagonists of NMDA-type glutamate receptors on brain-stimulation reward. Psychopharmacology. 1989;99:87–90. doi: 10.1007/BF00634458. [DOI] [PubMed] [Google Scholar]
- 89.Morgan CJA, Mofeez A, Brandner B, Bromley L, Curran HV. Ketamine impairs response inhibition and is positively reinforcing in healthy volunteers: a dose-response study. Psychopharmacology. 2004;172:298–308. doi: 10.1007/s00213-003-1656-y. [DOI] [PubMed] [Google Scholar]
- 90.Hancock PJ, Stamford JA. Stereospecific effects of ketamine on dopamine efflux and uptake in the rat nucleus accumbens. Br J Anaesth. 1999;82:603–608. doi: 10.1093/bja/82.4.603. [DOI] [PubMed] [Google Scholar]
- 91.Wise RA. Dopamine and reward: the anhedonia hypothesis 30 years on. Neurotox Res. 2008;14:169–183. doi: 10.1007/BF03033808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Morris SE, Cuthbert BN. Research Domain Criteria: cognitive systems, neural circuits, and dimensions of behavior. Dialogues Clin Neurosci. 2012;14:29–37. doi: 10.31887/DCNS.2012.14.1/smorris. [DOI] [PMC free article] [PubMed] [Google Scholar]