

Abstract
A bioenergetical analysis of swimming at intensities near competitive distances is inexistent. It was aimed to compare the transient VO2 kinetics responses and metabolic contributions whilst swimming at different velocities around VO2max. 12 trained male swimmers performed (i) an incremental protocol to determine the velocity at VO2max (vVO2max) and (ii) three square wave exercises from rest to 95, 100, and 105% of vVO2max. VO2 was directly measured using a telemetric portable gas analyser and its kinetics analysed through a double-exponential model. Metabolic contributions were assessed through the sum of three energy components. No differences were observed in the fast component response (τ 1—15, 18, and 16 s, A 1—36, 34, and 37 mL · kg−1 · min−1, and Gain—32, 29, and 30 mL · min−1 at 95, 100, and 105% of the vVO2max, resp.) but A2 was higher in 95 and 100% compared to 105% intensity (480.76 ± 247.01, 452.18 ± 217.04, and 147.04 ± 60.40 mL · min−1, resp.). The aerobic energy contribution increased with the time sustained (83 ± 5, 74 ± 6, and 59 ± 7% for 95, 100, and 105%, resp.). The adjustment of the cardiovascular and/or pulmonary systems that determine O2 delivery and diffusion to the exercising muscles did not change with changing intensity, with the exception of VO2 slow component kinetics metabolic profiles.
1. Introduction
In the 1920s a sustained period of research in human exercise physiology emerged, with swimming (along with cycling and running) as one of the primary areas of research in sport sciences. In this sport, the maximal oxygen uptake (VO2max) reveals itself as an important physiological parameter, expressing the swimmers' maximal metabolic aerobic performance [1, 2] that is one of the primary areas of interest in training and performance diagnosis [3]. In fact, determining the VO2max is the main aim of some studies, but the capacity to sustain this intensity in time is a recent topic of research. A big temporal gap exists between the pioneer study where VO2max was measured in swimming [4] and the first studies where time to exhaustion at the velocity corresponding to VO2max (Tlim-vVO2max) was assessed [5–7]. Though being conducted in nonspecific swimming pool conditions (swimming flume), these studies evidenced that Tlim-vVO2max depended inversely on the vVO2max but was not related with VO2max. To date, few studies have been conducted in conventional competition conditions [1, 8–13], and none has proposed how VO2 kinetics might impact concerning distinct vVO2max swimming intensities.
The quantification of the dynamic characteristics of VO2 kinetics has gained popularity in exercise physiology as a mean to unveil the mechanisms underlying the control O2 muscular consumption in humans during exercise [14]. Traditionally, the dynamic VO2 response to exercise has been studied at three intensity domains: moderate—below the anaerobic threshold, heavy—above the anaerobic threshold, and below the critical power and severe—above the critical power until the VO2max boundary [15, 16]. In the severe intensity domain, neither VO2 nor blood lactate concentrations ([La−]) can be stabilized, rising inexorably until fatigue ensues, with VO2 achieving its maximum value [17]. Here, the VO2 slow component is more pronounced compared to during heavy exercise, with its magnitude dependent on the duration and type of exercise [18]. More recently, the extreme exercise domain has been proposed for power outputs that lead to exhaustion before VO2max is attained [19], with the kinetics of VO2 characterized by the development of an evident fast component with insufficient time for the appearance of a discernible VO2 slow component [20]. Although the characteristics of the VO2 kinetics in the moderate and heavy intensity domains are well described (particularly in cycle and running ergometer exercise), few studies have investigated it during swimming around the VO2max intensity [7, 8, 11]. Moreover, these studies have only conducted a simplistic VO2 kinetics characterization by presenting the VO2 slow component as the unique kinetics parameter.
Given the current level of interest in exercise tolerance and VO2 kinetics, it is surprising that no study has examined both Tlim-vVO2max and VO2 kinetics response at intensities close to VO2max. Trying to overcome this absence of data and aiming to disseminate knowledge to other forms of human locomotion, the purpose of this study is to compare the transient VO2 kinetics responses whilst swimming until exhaustion at different velocities around the VO2max intensity. In addition, the different metabolic contributions of each exhaustion exercise will also be analyzed. It was hypothesized that 5% of variability in swimming velocity will not promote significant changes in the VO2 fast component kinetics response phase but will promote distinct VO2 kinetics in the slow component phase.
2. Material and Methods
2.1. Subjects
Twelve well trained male swimmers (mean ± SD; age: 18.2 ± 4.1 yr, height: 179.4 ± 6.5 cm, body mass: 70.5 ± 5.8 kg, and mean performance for long course 200 m freestyle of 86.5 ± 3.7% of the 2013 world record for this event), participants in national level competitions, volunteered to participate. All the swimmers, specialized in middle distance freestyle events, trained at least eight times per week and competed in National Championships for at least 4 years. Their maturational index (according to Tanner scale) corresponded to the 4 or 5 stages. Swimmers were familiar with the testing procedures as they were involved in previous physiological evaluations. All subjects avoided strenuous exercise in the 24 h before each testing session and were well hydrated and abstained from food, caffeine, and alcohol 3 h before testing. The protocols were conducted at the same time of the day for each subject and were separated by, at least, 24 h. All participants (or parent/guardians when subjects were under 18 yrs) provided informed written consent before data collection.
2.2. Experimental Design
Subjects visited the swimming pool facilities on four different occasions over a two-week period. In the first session, VO2max and vVO2max were determined through an intermittent incremental protocol until exhaustion. In the subsequent visits, subjects completed three square wave exercises from rest to distinct percentages of vVO2max intensity until exhaustion to assess its time sustained. Encouragement was given to motivate the swimmers to perform their best effort in the incremental protocol and to perform as long as possible during the square wave exercises.
2.3. Incremental Protocol
Briefly, each subject performed an individualized intermittent incremental protocol for front crawl vVO2max assessment, with increments of 0.05 m · s−1 and 30 s intervals between each 200 m stage until exhaustion [9, 21]. According to these studies, the initial velocity was established according to the individual level of fitness and was set at the swimmer's individual performance on the 400 m front crawl minus seven increments of velocity. The velocity was controlled at each stage by a visual pacer with flashing lights in the bottom of the pool (TAR.1.1, GBK-electronics, Aveiro, Portugal). VO2max was considered to be reached according to primary and secondary criteria [22] and all ventilatory parameters mean values were measured over the last 60 s of the exercise. If a plateau less than 2.1 mL · min−1 · kg−1 could not be observed, vVO2max was calculated as previously described [23].
2.4. Time to Exhaustion Tests
A total of three experimental exhaustive conditions were conducted in randomized order: 95, 100, and 105% of vVO2max. Each square wave exercise consisted of three distinct phases: 10 min warm-up at 50% of the VO2max, a short rest period (300 s), and the maintenance of the specific swimming intensity until exhaustion [8, 11]. The square wave exercises ended when the swimmers could no longer maintain the required velocity dictated by the visual feedback. Ventilatory parameters were calculated as the average values measured over the last 60 s of each exhaustive exercise.
2.5. Experimental Measurements
Respiratory and pulmonary gas-exchange variables were directly measured using a telemetric portable gas analyzer (K4b2, Cosmed, Rome, Italy), suspended over the water (at a 2 m height) in a steel cable following the swimmer along the pool and minimizing disturbances of the normal swimming movements. This apparatus was connected to the swimmer by a low hydrodynamic resistance respiratory snorkel and valve system (Aquatrainer, Cosmed, Rome, Italy [24]), which presents inspiratory and expiratory tubes of 86 cm length counting a volume of 847 mL from the mouthpiece to the turbine. It contains a dead space at the valves assembly, of 11.3 mL. In-water starts and open turns, without underwater gliding, were used. Heart rate (HR) was monitored and registered continuously by a Polar Vantage NV (Polar electro Oy, Kempele, Finland) that telemetrically emitted the data to the K4b2 portable unit. The gas analysers were calibrated before each test with gases of known concentration (16% O2 and 5% CO2) and the turbine volume transducer was calibrated by using a 3 L syringe. Capillary blood samples (25 μL) for [La−] were collected from the earlobe during the 30 s intervals (incremental protocol) and immediately at the end of exercise during the 1st, 3rd, 5th, and 7th min of the recovery period, until maximal values were reached ([La−]max), in both incremental and square wave exercises (Lactate Pro, Arkay, Inc., Kyoto, Japan).
2.6. Data Analysis
Firstly, occasional VO2 breath values were omitted from the analysis by including only those in-between VO2 mean ± 4 standard deviations. After verification of the data, individual breath-by-breath VO2 responses were smoothed by using a 3-breath moving average and time average of 5 s [21]. For VO2 kinetics analysis, the first 20 s of data after the onset of exercise (cardiodynamic phase) were not considered for model analysis. For this, a double-exponential (1) equation was used where a nonlinear least squares method was implemented in the MatLab Software to fit the VO2 data with the model:
| (1) |
where VO2(t) represents the relative VO2 at the time t, A 0 is the VO2 at rest (mL · kg−1 · min−1) and A 1 and A 2 (mL · kg−1 · min−1), TD1 and TD2 (s), and τ 1 and τ 2 (s) are the amplitudes, the corresponding time delays, and time constants of the fast and slow VO2 components, respectively. The oxygen deficit (DefO2) was calculated as the multiplication of A 1 and τ 1. In turn, A 1 was used to determine the gain (A 1/velocity) of the primary fast component. The relative contribution of the slow component to the overall increase in VO2 at the end exercise was calculated as [A 2/(A 1 + A 2)]. For the primary fast component to be accurately described by an exponential model, the duration of the VO2 primary rise before the onset of the VO2 slow component must be sufficiently long (≥4 times τ 1), in all subjects at all swimming intensities.
The maximal metabolic expenditure (E max) amounted during the square wave exercises was assumed to be the sum of the three components: aerobic (Aer), anaerobic lactic (Analac), and anaerobic alactic (Anaalac) energies [25–27]. The Aer contribution was calculated from the time integral of the net VO2 versus time relationship in the appropriate time ranges (mL O2) and then expressed in kJ assuming an energy equivalent of 20.9 kJ · lO2 −1 [28]. The Analac was estimated through the energy derived from lactic acid production (2):
| (2) |
where [La]bnet is the net accumulation of lactate after exercise, b is the energy equivalent for lactate accumulation in blood (2.7 mL O2 mM−1 · kg−1 [29]), and M (kg) is the mass of the swimmers (mL O2). Then, it was expressed in kJ assuming an energy equivalent of 20.9 kJ · lO2 −1 [28].
The Anaalac was assessed from the maximal PCr splitting in the contracting muscle:
| (3) |
where Anaalac is the anaerobic alactic contribution, t is the exercise time, τ is time constant of the PCr splitting at the onset of exhausting exercise (23.4 s [30]), M is the body mass, and PCr is the phosphocreatine concentration at rest. This latter was estimated assuming that, in transition from rest to exhaustion, its concentration decreases by 18.55 m-mole · kg−1 muscle wet weight (in a maximally working muscle mass equal to 30% of the overall body mass). Anaalac was, thus, expressed in kJ by assuming an energy equivalent of 0.468 kJ · mole−1 and a P/O2 ratio of 6.25 [31].
2.7. Statistical Analysis
Individual, mean, and standard deviations (SD) values were used for descriptive analysis for all studied variables and measures of skewness, kurtosis, and the Shapiro-Wilk test allowed to assess the normality and homogeneity of the data. The differences in ventilatory, metabolic, and VO2 kinetics parameters and time sustained between the square wave exercises at 95, 100, and 105% of vVO2max were tested for statistical significance using ANOVA for repeated measures. When a significant F value was achieved, the Bonferroni post hoc procedures were conducted to locate the pairwise differences between the averages. Simple linear regression and Pearson's correlation coefficients were also used. All statistical procedures were conducted with SPSS 10.05 and the significance level was set at 5%.
3. Results
Time to exhaustion decreased with increasing swimming velocity: 344.09 ± 63.64, 194.17 ± 47.79 and 122.64 ± 20.06 s at 1.34 ± 0.05, 1.39 ± 0.06 and 1.46 ± 0.07 m · s−1 (corresponding to 95, 100 and 105% of the vVO2max, resp.). Table 1 lists the mean ± SD data regarding the ventilatory and metabolic parameters assessed during the incremental and the square wave bouts performed at different percentages of vVO2max.
Table 1.
Mean ± SD values for VO2 max, HRmax, R, Ve, and [La−]max obtained at the end of the incremental protocol and the time to exhaustion tests (n = 12).
| Incremental protocol | 95% of vVO2 max | 100% of vVO2 max | 105% of vVO2 max | |
|---|---|---|---|---|
| VO2 max (L·min−1) | 4.21 ± 0.61 | 4.36 ± 0.55 | 4.41 ± 0.74 | 4.24 ± 0.64 |
| VO2 max (mL·kg−1 ·min−1) | 60.75 ± 5.17 | 61.34 ± 5.58 | 60.05 ± 6.10 | 59.78 ± 6.45 |
| HRmax (beats·min−1) | 180.6 ± 6.96 | 176.75 ± 9.63 | 176.91 ± 8.57 | 177.00 ± 8.90 |
| R | 0.93 ± 0.05 | 0.95 ± 0.06 | 1.01 ± 0.09 | 0.97 ± 0.06 |
| Ve (L·min−1) | 112.04 ± 23.38 | 116.54 ± 21.94 | 115.75 ± 34.25 | 119.27 ± 17.17 |
| [La−]max (mmol·L−1) | 7.18 ± 2.52 | 8.04 ± 1.70 | 8.86 ± 1.63 | 8.45 ± 1.91 |
VO2 max = maximal oxygen uptake; HRmax = maximal heart rate; R = respiratory quotient; Ve = ventilation; [La−]max = maximal blood lactate concentrations.
There were no differences between the determined physiologic parameters obtained in the incremental protocol and the square wave exercises and between the different time to exhaustion tests. Table 2 shows the VO2 kinetics parameters obtained at 95, 100, and 105% of vVO2max during the square wave exercises.
Table 2.
Mean ± SD values for the VO2 kinetics responses during at the time to exhaustion tests (n = 12).
| VO2 kinetics parameters | 95% of vVO2 max | 100% of vVO2 max | 105% of vVO2 max |
|---|---|---|---|
| A 0 (mL·min−1) | 1214.71 ± 351.79 | 1358.53 ± 368.71 | 1389.55 ± 257.32 |
| A 1 (mL·min−1) | 2568.31 ± 384.22 | 2402.64 ± 327.82 | 2628.24 ± 410.31 |
| TD1 (s) | 11.28 ± 3.98 | 8.60 ± 2.49 | 8.05 ± 3.49 |
| τ 1 (s) | 14.82 ± 4.01 | 18.06 ± 3.07 | 16.37 ± 3.81 |
| 95% confidence intervals (s) | (12.3–17.4) | (16.1–20.1) | (13.9–18.8) |
| DefO2 (L) | 0.60 ± 0.12 | 0.77 ± 0.24 | 0.74 ± 0.19 |
| Gain (mL·m−1) | 32.07 ± 4.54 | 29.39 ± 4.72 | 29.84 ± 4.43 |
| A 2 (mL·min−1) | 480.76 ± 247.01b | 452.18 ± 217.04b | 147.07 ± 60.40 |
| TD2 (s) | 106.29 ± 28.67a,b | 59.99 ± 12.50 | 69.07 ± 5.70 |
| τ 2 (s) | 120.23 ± 31.77b | 121.12 ± 31.71b | 61.46 ± 27.29 |
| % A 2 | 15.81 ± 7.87b | 16.36 ± 6.04b | 5.32 ± 1.99 |
A 0 = VO2 just before the beginning of exercise; A 1, TD1, τ 1, DefO2, and Gain = fast component amplitude, time delay, time constant, O2 deficit and gain, respectively; A 2, TD2, τ 2, and % A 2 = slow component amplitude, time delay, time constant, and relative contribution of slow component in relation to the end exercise VO2 of that bout, respectively. Differences between intensities are identified by a and b (100 and 105% of vVO2 max) (P ≤ 0.05).
At all intensities, the best data fit was obtained when the model incorporated a slow component (R 2 = 0.94, 0.92, and 0.90 for 95, 100, and 105% of vVO2max intensity) as opposed to a single exponential model, once a significant decrease in the sum of squared residuals occurred (the criterion measure used for each model). In fact, the F-test values evidenced a high heterogeneity between both models variances, also confirmed by the differences between their mean values. No differences were found between different time to exhaustion bouts regarding the fast component phase, but A 2 was higher at 95 and 100% compared to the 105% vVO2max intensity and with physiological meaning (≥200 mL · min−1) only at former intensities. In addition, the relative contribution of A 2 to total VO2 kinetics response was similar in-between 95 and 100% and higher than 105% vVO2max. Moreover, the TD2 was higher at 95% compared to 100 and 105% of vVO2max and τ 2 was higher in 95 and 100% compared to 105% of vVO2max. An individual VO2 kinetics response and the mean and SD values for the swimming velocity and the metabolic contributions at 95, 100, and 105% of vVO2max are shown in Figures 1(a) and 1(b).
Figure 1.
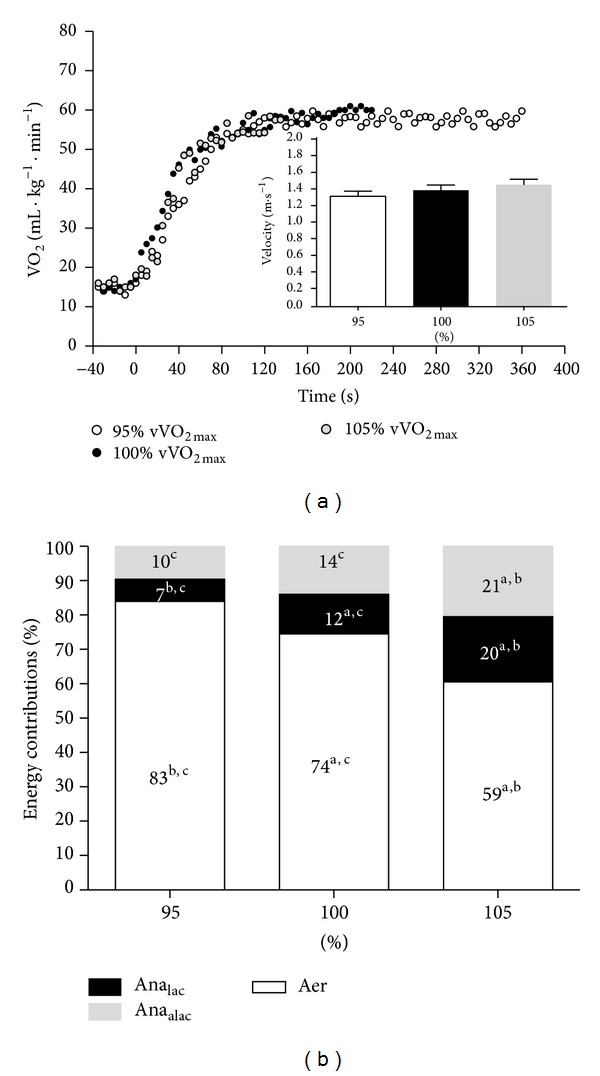
(a) VO2 kinetics individual response 5 s time average at 95, 100, and 105% of vVO2max. The insets represent the mean velocity values of each time to exhaustion bout; (b) mean aerobic, anaerobic lactic, and anaerobic alactic perceptual contributions obtained during the square wave exercises at 95, 100, and 105% of vVO2max (differences between intensities are identified by a, b, and c for 95, 100, and 105% of vVO2max, resp.; P ≤ 0.05).
Each square wave started with a sudden and exponential VO2 increase, independently of the swimming intensity. Differences between the Ana, Analac, and Anaalac contributions were found between time to exhaustion intensities, with exception to the Anaalac that was similar between 95 and 100% of vVO2max.
In Figure 2 it is possible to observe the relationships between swimming performance indicators and VO2 kinetics parameters obtained at the different time to exhaustion bouts, with significant correlation values found between (i) HRmax and τ 1 at 95% of vVO2max (a); (ii) vVO2max and time sustained at 100% of vVO2max (b); and (iii) vVO2max and time sustained at 105% of vVO2max, and, vVO2max and A 1 (c).
Figure 2.
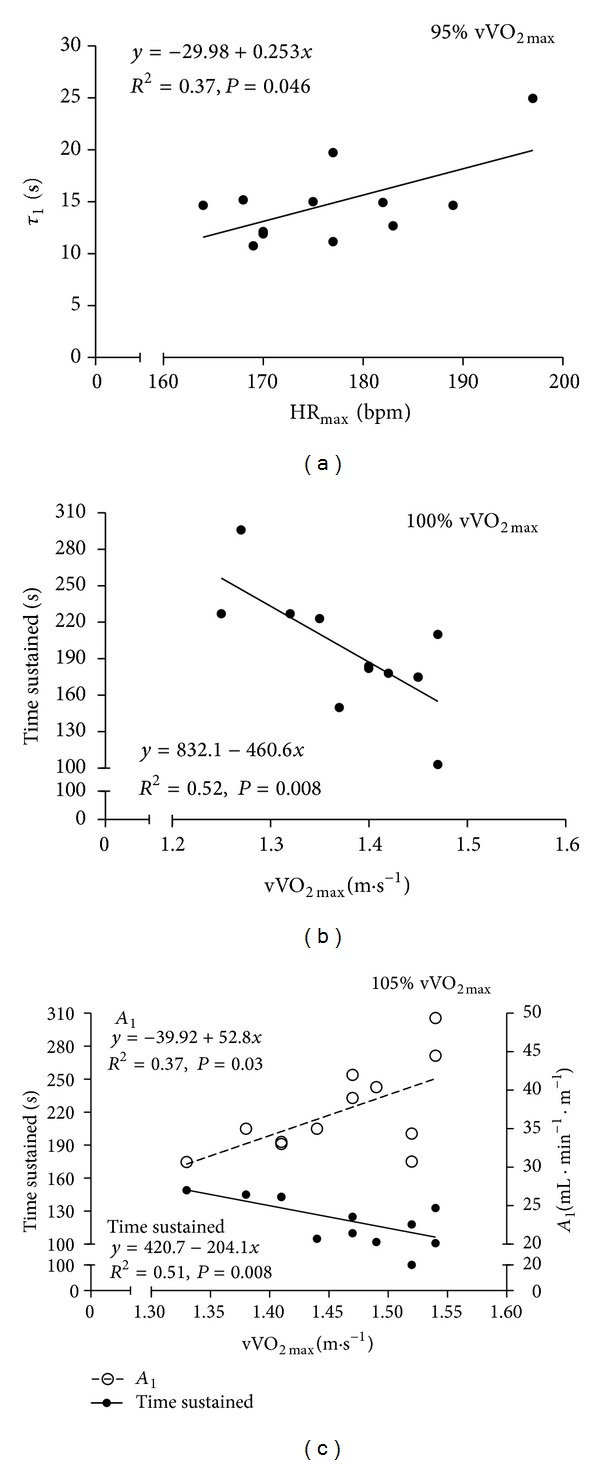
Relationships between maximum peak heart rate (HRmax) and fast component time constant (τ 1) at 95% of vVO2max intensity (a), velocity and time sustained at 100% of vVO2max intensity (b), and velocity and time sustained (filled circles) and velocity and fast component amplitude (unfilled circles) at 105% of vVO2max intensity (c). The regression equations, determination coefficients, and significance level values are also identified.
4. Discussion
Studies investigating VO2 kinetics when performing to exhaustion have been conducted mainly in cycle ergometry and treadmill exercise, presenting a pretty simplistic approach when comparing different exercise intensities. The current study is the first attempt to examine and compare the VO2 kinetics during swimming to exhaustion at different velocities around the VO2max intensity. The exercise duration decreased when intensity increased, similarly to what was proposed for other cyclic sports. In addition, no differences were found in the VO2 fast component related parameters (τ 1, A 1, and Gain) between 95, 100, and 105% of vVO2max, supporting our hypothesis that a 5% change in swimming velocity surrounding the VO2max intensity would not be sufficient to promote changes in the primary phase of VO2 kinetics response. However, A 2 was higher at 95 and 100% compared to 105% of vVO2max corroborating the hypothesis that different swimming intensities near vVO2max would promote distinct VO2 slow component kinetic profiles. In addition, E max was different between the studied intensities.
VO2max is the most commonly measured parameter in applied physiological sciences. The mean values obtained at the end of the incremental protocol are in accordance with those presented for middle distance swimmers [5, 6], but lower than those described for elite swimmers [8–11], runners [32, 33], cyclists [34, 35], and rowers [36, 37], probably explained by the use of a larger muscle mass in these sports. Also, the observed [La−]max mean values are lower compared to other exercise modes, which can explain the lower R mean values found. This fact suggests that a lower metabolic acidosis occurs in swimming compared to other sports or that swimmers are less sensitive to it [5]. Furthermore, no differences were observed in the ventilatory and metabolic parameters between the incremental protocol and the square wave exercises, in agreement with the literature for 100% vVO2max swimming exercise [1, 5, 11]. In-between intensities comparison did not evidence changes in ventilatory and metabolic mean values, conversely to the differences found between 100 and 105% of vVO2max in running [32].
Specifically in swimming, only Demarie et al. [7] analysed a time to exhaustion at intensities different from 100% of vVO2max, showing that swimmers were able to sustain ~375 s at 96% of vVO2max, studying agreement with our current data (~344 s). The values reported in the current study for 100% vVO2max are also similar to those presented for highly trained swimmers performing at the same intensity [9–11] but are lower than others obtained in nonreal competition conditions [5, 6]. In addition, Alberty et al. [12, 13] conducted studies performed at 95 and 100% of the velocity of the 400 m front crawl (not measuring ventilatory parameters), observing a longer time to exhaustion compared to ours (~670 and 238 s). Collectively, these studies seem to evidence that time sustained at intensities around vVO2max depends also on the conditions in which they occurred. In fact, it has been reported that swimming flume might influence the VO2max, vVO2max, and the Tlim-vVO2max assessment, as well as the swimming technique, and, therefore, could explain the differences found [8].
Concerning the VO2 kinetics, the observed τ 1 mean value is lower than values obtained for to 100 and 400 m front crawl all-out efforts [38, 39] but higher compared to the 200 m front crawl all-out efforts [2, 40]. Conversely, the value found is in accordance with previous reports for 7 min swimming at heavy [41, 42] and severe intensities [42] and for 400 m performed at 100% vVO2max [43]. The VO2 kinetics response was also characterized by a similar τ 1 in-between the time to exhaustion tests, in line with previous cycling and treadmill ergometer studies who have showed that it remains constant as exercise intensity increases from moderate to heavy and to severe intensity domains, despite the increasing acidosis [44–47]. The current study also corroborates the absence of differences in τ 1 between intensities around VO2max cycling exercise (90, 100, and 120%, [47]). It has been suggested that the characteristics of VO2 kinetics provide insights into the physiological mechanisms responsible for the control of, and the limitations to, VO2 kinetics following the onset of exercise [48]. Thus, similar τ 1 values observed seem to suggest that an O2 delivery and diffusion are not influenced by a 5% external arousal in swimming at vVO2max intensity. In fact, at 95% of vVO2max, a positive correlation was observed between HRmax and τ 1, evidencing that if a limiting factor exists, it may be related to peripheral factors (from convective O2 transport, to its diffusion and utilization in the muscles) and not to central ones (O2 delivery and transportation to the working muscles).
Complementarily, the A 1 mean value obtained in this study is in accordance with previous reports for swimming at heavy intensity [41] but lower compared to higher intensities [2, 40, 42]. The similar A 1 values across conditions do not corroborate the fact that an increase in amplitude is linearly related to the increase in exercise intensity [45–47]. However, when comparing intensities around VO2max in cycling exercise [47], differences have only been observed between 90% and 110% of vVO2max. These results suggest that the 5% velocity changes on our experimental set were not sufficient to induce modifications in A 1, as previously noted for τ 1. Well linked to the VO2 first component amplitude is the fast component Gain that evidenced a tendency to decrease with increasing intensity, in line with the decrease reported as the exercise intensity approaches the individuals' VO2max [47]. Comparison in the VO2 gain between intensities around VO2max (90, 100, and 110% of VO2max) have been reported only for cycling exercise [47], being this study the first attempt to assess the VO2 gain during swimming exercise surrounding the VO2max intensity. The lack of differences seems to indicate that the 5% changes in swimming velocity were insufficient to induce an adjustment of O2 delivery and diffusion to the exercising muscles.
The magnitude of the VO2 slow component is considered to have physiological meaning only when it is ≥200 mL · min−1 [49] (although this value is still a matter of debate), occurring at 95 and 100% of vVO2max. At these intensities (the severe intensity domain), the attainment of a VO2 steady state is delayed due to the emergence of a supplementary slowly developing component of the VO2 response [18], corroborating the lack of differences found in A 2 between 95 and 100% of vVO2max intensities. The A 2 values found in the current study for 95 and 100% of vVO2max intensities are in accordance with those previously reported at 96% of vVO2max [7] but are higher than those presented for 100% of vVO2max [8, 11] and for the heavy intensity exercise domain [41, 42]. This lack of agreement could be explained by the method that was used to assess the VO2 slow component-fixed interval method that seems to result in lower values compared to the mathematical modeling method [39] and is considerd a simple rough estimate of this parameter [48].
At intensities higher than VO2max (the extreme intensity domain), the exercise duration is so short (≤2 min) that a VO2 slow component is not readily observed [16], confirmed at 105% of vVO2max. The relative contribution of the VO2 slow component to the overall increase in VO2 at the end exercise was higher than those presented for heavy domain [41, 42], but being A 1 and A 2 dependent variables, this was an expectable outcome. Although some explanations to better understand the VO2 slow component phenomenon have been proposed [3], its origin in swimming is even more uncertain than in running and cycling [7]. The current study seems to indicate that possibly the type and pattern of recruitment of the available motor units were clearly modified. The lack of direct relationships between time sustained and VO2 slow component, as found previously [8, 17, 50], suggests that near the vVO2max intensity the slow component is not linked with the time sustained.
Regarding E max values, the Aer contribution was higher at 95% compared to 100 and 105% of vVO2max and the anaerobic (lactic and alactic) contribution evidenced the opposite trend. These relative values (percentage of the total energy spent) show similar absolute values for the Analac and Anaalac contributions (~21, 26, and 28 kJ and 29.3, 29.2, and 29.1 kJ for 95, 100, and 105% of vVO2max, resp.) but different ones for the Aer contribution (265, 169, and 184 kJ at 95, 100, and 105% of vVO2max, resp.). This fact can be explained by the time sustained at each intensity once the assessment of the Aer contribution was through the time integral from the VO2 to time curve. To date, this metabolic comparison has never been conducted at different intensities around vVO2max. In fact, the literature regarding all energetic contributions in all sports is very scarce and, particularly in swimming, has been applied remotely [25–27]. Thus, caution must be taken when comparisons between the present results with others studies are made once the method by which the energy release was determined can have a significant influence on the calculated relative contribution of the energy systems during periods of maximal exercise [51]. At 95% of vVO2max, the Aer contribution (~83%) was similar to the percentage proposed previously for 333 s exercise duration [26], but the Analac (~7%) and Anaalac (~10%) contributions were lower than those presented for 112 s [25] and 200 m all-out effort [27]. At 100 and 105% of vVO2max, the Aer contributions (~74 and 59%) were similar to previous reports [25–27] but some inconsistencies were found in the remaining fractions of Analac (~12 and 20%) and Anaalac (~14 and 21%) contributions [25, 27]. The observed inverse relationship between time sustained and vVO2max at 100 and 105% of vVO2max (Figure 2) is in accordance with previous reports for similar swimming intensities [1, 5, 6, 8, 9] and could indicate a significative strenuous effort, with a more pronounced recruitment of anaerobic energy pathways (Figure 1). This fact could lead to earlier fatigue stages and, consequently, to shorter time sustained efforts. The inverse trend (greater dependency of the aerobic energy pathway) could explain the lack of an inverse relationship between time to exhaustion and vVO2peak at 95% of vVO2max.
Further studies to compare the transient VO2 kinetics responses and metabolic contributions whilst swimming at different velocities around maximal intensities are supported by the data from this study. However, the fact that only one transition from rest to 95, 100, and 105% of vVO2max intensity was done could lead to a low signal-to-noise-ratio. Consequently, this factor could have influenced the swimmers' performance and subsequent VO2 kinetics and, for that, should be construed as a possible limitation of the present study.
5. Conclusions
This study was the first attempt to examine and compare the VO2 kinetics and the metabolic profile during time to exhaustion exercise at different velocities around the vVO2max intensity. The 5% velocity variability across conditions was not sufficient to promote changes in the kinetics of the VO2 fast component (τ 1, A 1, and Gain) but resulted in differences in the kinetics of the VO2 slow component (A 2) and the corresponding metabolic profiles. Though being well documented in cycling and running exercise, VO2 kinetics has received considerably less research attention in swimming, even though it is providing a noninvasive into oxidative metabolism at the muscle level. Since athletes typically train at intensities surrounding the VO2max, understanding how subtle variations in intensity surrounding the VO2max impacts on oxidative metabolism and performance might have important implications for optimising high-intensity interval training.
Acknowledgments
The authors acknowledge the Portuguese Science and Technology Foundation (FCT): PTDC/DES/101224/2008 Grant. Ana Sousa acknowledges the Portuguese Science and Technology Foundation (FCT) for the PhD Grant (SFRH/BD/72610/2010).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Renoux J-C. Evaluating the time limit at maximum aerobic speed in elite swimmers. Training implications. Archives of Physiology and Biochemistry. 2001;109(5):424–429. doi: 10.1076/apab.109.5.424.8036. [DOI] [PubMed] [Google Scholar]
- 2.Sousa AC, Figueiredo P, Oliveira NL, et al. VO2 kinetics in 200-m race-pace front crawl swimming. International Journal of Sports Medicine. 2011;32(10):765–770. doi: 10.1055/s-0031-1279772. [DOI] [PubMed] [Google Scholar]
- 3.Fernandes RJ, Vilas-Boas JP. Time to exhaustion at the VO2max velocity in swimming: a review. Journal of Human Kinetics. 2012;32(1):121–134. doi: 10.2478/v10078-012-0029-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Liljestrand G, Lindhard J. Über das minutenvolumen des herzens beim schwimmen. Skandinavisches Archiv für Physiologie. 1920;39(1):64–77. [Google Scholar]
- 5.Billat V, Faina M, Sardella F, et al. A comparison of time to exhaustion at VO2 max in élite cyclists, kayak paddlers, swimmers and runners. Ergonomics. 1996;39(2):267–277. doi: 10.1080/00140139608964457. [DOI] [PubMed] [Google Scholar]
- 6.Faina M, Billat V, Squadrone R, de Angelis M, Koralsztein JP, dal Monte A. Anaerobic contribution to the time to exhaustion at the minimal exercise intensity at which maximal oxygen uptake occurs in elite cyclists, kayakists and swimmers. European Journal of Applied Physiology and Occupational Physiology. 1997;76(1):13–20. doi: 10.1007/s004210050207. [DOI] [PubMed] [Google Scholar]
- 7.Demarie S, Sardella F, Billat V, Magini W, Faina M. The VO2 slow component in swimming. European Journal of Applied Physiology. 2001;84(1-2):95–99. doi: 10.1007/s004210000348. [DOI] [PubMed] [Google Scholar]
- 8.Fernandes R, Cardoso C, Soares S, Ascenção A, Colaço P, Vilas-Boas J. Time limit and VO2 slow part at intensities corresponding to VO2max in swimmers. International Journal of Sports Medicine. 2003;24(8):576–581. doi: 10.1055/s-2003-43274. [DOI] [PubMed] [Google Scholar]
- 9.Fernandes RJ, Billat VL, Cruz AC, Colaço PJ, Cardoso CS, Vilas-Boas JP. Does net energy cost of swimming affect time to exhaustion at the individual’s maximal oxygen consumption velocity? Journal of Sports Medicine and Physical Fitness. 2006;46(3):373–380. [PubMed] [Google Scholar]
- 10.Fernandes RJ, Marinho DA, Barbosa TM, Vilas-Boas JP. Is time limit at the minimum swimming velocity of VO2 max influenced by stroking parameters? Perceptual and Motor Skills. 2006;103(1):67–75. doi: 10.2466/pms.103.1.67-75. [DOI] [PubMed] [Google Scholar]
- 11.Fernandes RJ, Keskinen KL, Colaço P, et al. Time limit at VO2max velocity in elite crawl swimmers. International Journal of Sports Medicine. 2008;29(2):145–150. doi: 10.1055/s-2007-965113. [DOI] [PubMed] [Google Scholar]
- 12.Alberty M, Potdevin F, Dekerle J, Pelayo P, Gorce P, Sidney M. Changes in swimming technique during time to exhaustion at freely chosen and controlled stroke rates. Journal of Sports Sciences. 2008;26(11):1191–1200. doi: 10.1080/02640410801974984. [DOI] [PubMed] [Google Scholar]
- 13.Alberty M, Sidney M, Pelayo P, Toussaint HM. Stroking characteristics during time to exhaustion tests. Medicine & Science in Sports & Exercise. 2009;41(3):637–644. doi: 10.1249/MSS.0b013e31818acfba. [DOI] [PubMed] [Google Scholar]
- 14.Adami A, Pogliaghi S, de Roia G, Capelli C. Oxygen uptake, cardiac output and muscle deoxygenation at the onset of moderate and supramaximal exercise in humans. European Journal of Applied Physiology. 2011;111(7):1517–1527. doi: 10.1007/s00421-010-1786-y. [DOI] [PubMed] [Google Scholar]
- 15.Xu F, Rhodes EC. Oxygen uptake kinetics during exercise. Sports Medicine. 1999;27(5):313–327. doi: 10.2165/00007256-199927050-00003. [DOI] [PubMed] [Google Scholar]
- 16.Jones AM, Burnley M. Oxygen uptake kinetics: an underappreciated determinant of exercise performance. International Journal of Sports Physiology and Performance. 2009;4(4):524–532. doi: 10.1123/ijspp.4.4.524. [DOI] [PubMed] [Google Scholar]
- 17.Gaesser GA, Poole DC. The slow component of oxygen uptake kinetics in humans. Exercise and Sport Sciences Reviews. 1996;24:35–70. [PubMed] [Google Scholar]
- 18.Jones AM, Grassi B, Christensen PM, Krustrup P, Bangsbo J, Poole DC. Slow component of VO2 kinetics: mechanistic bases and practical applications. Medicine & Science in Sports & Exercise. 2011;43(11):2046–2062. doi: 10.1249/MSS.0b013e31821fcfc1. [DOI] [PubMed] [Google Scholar]
- 19.Hill DW, Poole DC, Smith JC. The relationship between power and the time to achieve VO2max. Medicine & Science in Sports & Exercise. 2002;34(4):709–714. doi: 10.1097/00005768-200204000-00023. [DOI] [PubMed] [Google Scholar]
- 20.Burnley M, Jones AM. Oxygen uptake kinetics as a determinant of sports performance. European Journal of Sport Science. 2007;7(2):63–79. [Google Scholar]
- 21.Fernandes RJ, de Jesus K, Baldari C, et al. Different VO2max time-averaging intervals in swimming. International Journal of Sports Medicine. 2012;33(12):1010–1015. doi: 10.1055/s-0032-1316362. [DOI] [PubMed] [Google Scholar]
- 22.Howley ET, Bassett DR, Jr., Welch HG. Criteria for maximal oxygen uptake: review and commentary. Medicine & Science in Sports & Exercise. 1995;27(9):1292–1301. [PubMed] [Google Scholar]
- 23.Kuipers H, Verstappen FTJ, Keizer HA, Geurten P, van Kranenburg G. Variability of aerobic performance in the laboratory and its physiologic correlates. International Journal of Sports Medicine. 1985;6(4):197–201. doi: 10.1055/s-2008-1025839. [DOI] [PubMed] [Google Scholar]
- 24.Baldari C, Fernandes RJ, Meucci M, Ribeiro J, Vilas-Boas JP, Guidetti L. Is the new AquaTrainer snorkel valid for VO2 assessment in swimming? International Journal of Sports Medicine. 2013;34(4):336–344. doi: 10.1055/s-0032-1321804. [DOI] [PubMed] [Google Scholar]
- 25.Capelli C, Pendergast DR, Termin B. Energetics of swimming at maximal speeds in humans. European Journal of Applied Physiology and Occupational Physiology. 1998;78(5):385–393. doi: 10.1007/s004210050435. [DOI] [PubMed] [Google Scholar]
- 26.Zamparo P, Capelli C, Cautero M, di Nino A. Energy cost of front-crawl swimming at supra-maximal speeds and underwater torque in young swimmers. European Journal of Applied Physiology. 2000;83(6):487–491. doi: 10.1007/s004210000318. [DOI] [PubMed] [Google Scholar]
- 27.Figueiredo P, Zamparo P, Sousa A, Vilas-Boas JP, Fernandes RJ. An energy balance of the 200 m front crawl race. European Journal of Applied Physiology. 2011;111(5):767–777. doi: 10.1007/s00421-010-1696-z. [DOI] [PubMed] [Google Scholar]
- 28.Zamparo P, Capelli C, Pendergast D. Energetics of swimming: a historical perspective. European Journal of Applied Physiology. 2011;111(3):367–378. doi: 10.1007/s00421-010-1433-7. [DOI] [PubMed] [Google Scholar]
- 29.di Prampero P, Pendergast D, Wilson D, Rennie D. Blood lactic acid concentrations in high velocity swimming. In: Eriksson B, Furberg B, editors. Swimming Medicine IV. Baltimore, Md, USA: University Park Press; 1978. pp. 249–261. [Google Scholar]
- 30.Binzoni T, Ferretti G, Schenker K, Cerretelli P. Phosphocreatine hydrolysis by 31P-NMR at the onset of constant-load exercise in humans. Journal of Applied Physiology. 1992;73(4):1644–1649. doi: 10.1152/jappl.1992.73.4.1644. [DOI] [PubMed] [Google Scholar]
- 31.Sousa A, Figueiredo P, Zamparo P, Vilas-Boas JP, Fernandes RJ. Anaerobic alactic energy assessment in middle distance swimming. European Journal of Applied Physiology. 2013;113(8):2153–2158. doi: 10.1007/s00421-013-2646-3. [DOI] [PubMed] [Google Scholar]
- 32.Billat V, Renoux JC, Pinoteau J, Petit B, Koralsztein JP. Times to exhaustion at 90, 100 and 105% of velocity at VO2 max (maximal aerobic speed) and critical speed in elite long-distance runners. Archives of Physiology and Biochemistry. 1995;103(2):129–135. doi: 10.3109/13813459508996126. [DOI] [PubMed] [Google Scholar]
- 33.Renoux JC, Petit B, Billat V, Koralsztein JP. Oxygen deficit is related to the exercise time to exhaustion at maximal aerobic speed in middle distance runners. Archives of Physiology and Biochemistry. 1999;107(4):280–285. doi: 10.1076/13813455199908107041qft280. [DOI] [PubMed] [Google Scholar]
- 34.Chavarren J, Calbet JAL. Cycling efficiency and pedalling frequency in road cyclists. European Journal of Applied Physiology and Occupational Physiology. 1999;80(6):555–563. doi: 10.1007/s004210050634. [DOI] [PubMed] [Google Scholar]
- 35.Coyle EF, Sidossis LS, Horowitz JF, Beltz JD. Cycling efficiency is related to the percentage of Type I muscle fibers. Medicine & Science in Sports & Exercise. 1992;24(7):782–788. [PubMed] [Google Scholar]
- 36.Jensen RL. The prediction of power and efficiency during near-maximal rowing. European Journal of Applied Physiology and Occupational Physiology. 1996;73(1-2):98–104. doi: 10.1007/BF00262816. [DOI] [PubMed] [Google Scholar]
- 37.Sousa A, Ribeiro J, Sousa M, Vilas-Boas JP, Fernandes RJ. Influence of prior exercise on VO2 kinetics subsequent exhaustive rowing performance. PLoS ONE. 2014;9(1) doi: 10.1371/journal.pone.0084208.e84208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Rodríguez F, Keskinen K, Malvela M, Keskinen O. Oxygen uptake kinetics during free swimming: a pilot study. In: Chatard JE, editor. IX Biomechanics and Medicine in Swimming. Publications de l’Université de Saint-Étienne; 2003. pp. 379–384. [Google Scholar]
- 39.Reis J, Santos E, Oliveira D, Gonçalves L, Carneiro A, Fernandes R. Oxygen uptake slow component at submaximal swimming. Gazzetta Medica Italiana: Archivio per le Scienze Mediche. 2013;172:603–610. [Google Scholar]
- 40.Sousa A, Figueiredo P, Keskinen KL, et al. VO2 off transient kinetics in extreme intensity swimming. Journal of Sports Science and Medicine. 2011;10(3):546–552. [PMC free article] [PubMed] [Google Scholar]
- 41.Reis JF, Alves FB, Bruno PM, Vleck V, Millet GP. Effects of aerobic fitness on oxygen uptake kinetics in heavy intensity swimming. European Journal of Applied Physiology. 2012;112(5):1689–1697. doi: 10.1007/s00421-011-2126-6. [DOI] [PubMed] [Google Scholar]
- 42.Filho DMP, Alves FB, Reis JF, Greco CC, Denadai BS. VO2 kinetics during heavy and severe exercise in swimming. International Journal of Sports Medicine. 2012;33(9):744–748. doi: 10.1055/s-0031-1299753. [DOI] [PubMed] [Google Scholar]
- 43.Bentley DJ, Roels B, Hellard P, Fauquet C, Libicz S, Millet GP. Physiological responses during submaximal interval swimming training: effects of interval duration. Journal of Science and Medicine in Sport. 2005;8(4):392–402. doi: 10.1016/s1440-2440(05)80054-4. [DOI] [PubMed] [Google Scholar]
- 44.Barstow TJ. Characterization of VO2 kinetics during heavy exercise. Medicine & Science in Sports & Exercise. 1994;26(11):1327–1334. [PubMed] [Google Scholar]
- 45.Barstow TJ, Jones AM, Nguyen PH, Casaburi R. Influence of muscle fiber type and pedal frequency on oxygen uptake kinetics of heavy exercise. Journal of Applied Physiology. 1996;81(4):1642–1650. doi: 10.1152/jappl.1996.81.4.1642. [DOI] [PubMed] [Google Scholar]
- 46.Pringle JSM, Doust JH, Carter H, Tolfrey K, Campbell IT, Jones AM. Oxygen uptake kinetics during moderate, heavy and severe intensity “submaximal” exercise in humans: the influence of muscle fibre type and capillarisation. European Journal of Applied Physiology. 2003;89(3-4):289–300. doi: 10.1007/s00421-003-0799-1. [DOI] [PubMed] [Google Scholar]
- 47.Scheuermann BW, Barstow TJ. O2 uptake kinetics during exercise at peak O2 uptake. Journal of Applied Physiology. 2003;95(5):2014–2022. doi: 10.1152/japplphysiol.00590.2002. [DOI] [PubMed] [Google Scholar]
- 48.Jones AM, Poole DC. Oxygen Uptake Kinetics in Sport, Exercise and Medicine. London, UK: Routledge; 2005. [Google Scholar]
- 49.Billat VL, Morton RH, Blondel N, et al. Oxygen kinetics and modelling of time to exhaustion whilst running at various velocities at maximal oxygen uptake. European Journal of Applied Physiology. 2000;82(3):178–187. doi: 10.1007/s004210050670. [DOI] [PubMed] [Google Scholar]
- 50.Whipp BJ. The slow component of O2 uptake kinetics during heavy exercise. Medicine & Science in Sports & Exercise. 1994;26(11):1319–1326. [PubMed] [Google Scholar]
- 51.Gastin PB. Energy system interaction and relative contribution during maximal exercise. Sports Medicine. 2001;31(10):725–741. doi: 10.2165/00007256-200131100-00003. [DOI] [PubMed] [Google Scholar]