

Abstract
AIM: To study the effects of hepatic ischemia/reperfusion (I/R) injury on store-operated calcium channel (SOC) currents (ISOC) in freshly isolated rat Kupffer cells, and the effects of Ca2+ channel blockers, 2-aminoethoxydiphenyl borate (2-APB), SK&F96365, econazole and miconazole, on ISOC in isolated rat Kupffer cells after hepatic I/R injury.
METHODS: The model of rat hepatic I/R injury was established. Whole-cell patch-clamp techniques were performed to investigate the effects of 2-APB, SK&F96365, econazole and miconazole on ISOC in isolated rat Kupffer cells after hepatic I /R injury.
RESULTS: I/R injury significantly increased ISOC from -80.4 ± 25.2pA to -159.5 ± 34.5pA (bP < 0.01, n = 30). 2-APB (20, 40, 60, 80, 100 μmol/L), SK&F96365 (5, 10, 20, 40, 50 μmol/L), econazole (0.1, 0.3, 1, 3, 10 μmol/L) and miconazole (0.1, 0.3, 1, 3, 10 μmol/L) inhibited ISOC in a concentration-dependent manner with IC50 of 37.41 μmol/L (n = 8), 5.89 μmol/L (n = 11), 0.21 μmol/L (n = 13), and 0.28 μmol/L (n = 10). The peak value of ISOC in the I-V relationship was decreased by the blockers in different concentrations, but the reverse potential of ISOC was not transformed.
CONCLUSION: SOC is the main channel for the influx of Ca2+ during hepatic I/R injuries. Calcium channel blockers, 2-APB, SK&F96365, econazole and miconazole, have obviously protective effects on I/R injury, probably by inhibiting Isoc in Kupffer cells and preventing the activation of Kupffer cells.
Keywords: Kupffer cell, Ischemia/reperfusion, Store-operated calcium channel currents, 2-aminoethoxydiphenyl borate, SK&F96365, Econazole, Miconazole
INTRODUCTION
In clinical practice of hepatobiliary surgery, various factors, such as shock, inflammation, hepatic trauma, operation of liver and biliary tract (in case of necessities of interrupting hepatic portal vein), liver transplantation, are mutually related to common pathophysiological procedures, named hepatic ischemia/reperfusion (I/R) injury. Kupffer cells play an important role in hepatic I/R injury, hepatic I/R injury can be relieved when Kupffer cells are inactivated, and store-operated Ca2+ channels (SOC) are present both in almost all non-excitable cells and in some excitable cells[1-4], but there is no report about SOC in Kupffer cells. 2-APB, SK&F96365, econazole and miconazole have been used as the blockers of SOC in many cells. In the present study, we investigated the effects of hepatic I/R injury on store-operated calcium channel currents (Isoc) in freshly isolated rat Kupffer cells, and the effects of calcium channel blockers, 2-APB, SK&F96365, econazole and miconazole on Isoc in isolated rat Kupffer cells after hepatic I/R injury.
MATERIALS AND METHODS
Materials
Male Sprague Dawley rats (Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences), weighing 200-300 g, were used in all experiments. Thapsigargin was obtained from Alexis Company. 2-aminoethoxydiphenyl borate (2-APB), SK&F96365 were obtained from Merck KcaA (Darmstadt, Germany). Collangenase IV, HEPES, EGTA, trypan blue, econazole, miconazole, CsCl, CsOH and the other chemicals were from Sigma (USA). The pipette solution contained 120.0 mmol/L CsCl, 1.0 mmol/L MgCl2, 10.0 mmol/L EGTA, 10.0 mmol/L HEPES, 2 μmol/L thasigargin, pH 7.3 (adjusted with CsOH). The bath solution contained 145 mmol/L NaCl, 2.8 mmol/L KCl, 10.0 mmol/L CaCl2, 1.0 mmol/L CSCl, 2.0 mmol/L MgCl2, 10.0 mmol/L glucose, 10.0 mmol/L HEPES, pH 7.3 (adjusted with NaOH). Ca2+-Mg2+-free Krebs-ringer-HEPES buffer solution contained 5 mmol/L KCl, 1 mmol/L KH2PO4, 115 mmol/L NaCl, 25 mmol/L HEPES, 0.5 mmol/L EGTA, pH 7.3 (adjusted with NaOH).
Hepatic I/R injury model
The model of rat partial hepatic I/R injury was established according to the procedures reported by Colletti et al[5] with minor modifications. Briefly, adult animals were anesthetized with pentobarbital sodium (50 mg/kg) and heparinized (1 U/g) via an intraperitoneal injection. A midline laparotomy was performed, then an atraumatic clip was used to interrupt the arterial and portal venous blood supply to the cephalad three lobes of liver. After 20 min for hepatic ischemia, the clip was removed followed by 40 min for hepatic reperfusion. Sham-operated control animals were treated in an identical fashion with the omission of vascular occlusion.
Cell preparation
Rat kupffer cells were enzymatically isolated from SD rats. Briefly, after the model was achieved, the portal vein and inferior vena cava were cannulated. The liver was initially perfused at a flow rate of 20-30 mL/min with a constant-flow system with modified oxygenated Ca2+-Mg2+-free Krebs-ringer-HEPES buffer solution (37°C) for about 5 min, followed by Krebs-ringer-HEPES buffer solution (37°C) containing collagenase IV for 10 min. After the perfusions, cephalad three lobes of the liver were excised and minced in Ca2+-Mg2+-free Krebs-ringer-HEPES solution (37°C) for 20 min. The cells were filtered through a 200 μm nylon mesh, and washed by centrifugation at 700 g for 7 min. Then we reserved sediment and poured about 20-30 mL PBS into the sediment, washed it by centrifugation at 50 g for 4 min, reserved supernatant and washed it by centrifugation at 700 g for 5 min, plated the sediment on glass cover-slips and incubated it at 37°C in DMEM for 1-2 h. Then the cells were washed 3-5 times with the bath solution. The cells attached to the bottom of glass cover-slips were kupffer cells. The spherical, smooth cells were used for the whole-cell patch-clamp studies. All experiments were performed at room temperature (20-24°C).
Electrophysiological recordings
An automatic micropipette puller (Model P-97, Sutter Instruments, Novato, CA) was used to pull the electrodes. The resistance of the capillary glass electrodes (GG217, Would Precision, USA) used was 4-8 MΩ when filled with internal solution. An Axopatch 200B amplifier (Axon Instruments, USA) was used to record whole-cell currents with the filter set at 3 kHz, digitized at 5 kHz. The protocol of a series of depolarizing pulses from a holding potential of 0 mV to different membrane potentials (-120 mV to +80 mV) with a 20 mV increment was used for voltage clamp. Data analysis was performed using software (pCLAMP9.0, Axon Instruments, USA). The data were stored in computer for subsequent analysis. The blockers were added to the place around the cells with a rapid solution changer to investigate their effects on Isoc.
Statistical analysis
All values were expressed as mean ± SD. Appropriate t-test was used for the statistical analysis. aP < 0.05 and bP < 0.01 were considered statistically significant and apparently significant respectively.
RESULTS
Viability of isolated rat Kupffer cells
The isolated Kupffer cells showed polymorphism with typical polygon-like and star-like shapes. The purity and adhesion rate were 85% and 39.4%, respectively. The viability was over 80%. The isolated rat Kupffer cells were suitable for whole-cell patch clamp technique.
Isoc in isolated rat Kupffer cells
After the whole-cell configuration was established, cell membrane potential was clamped at 0 mV, and different test potentials from -120 to +80 mV with a 20 mV increment at a frequency of 0.2Hz were applied. Following break-in, the currents in ischemia/reperfusion group (-159.5 ± 34.5 pA, n = 30) were greater than those in sham-operated control group (-80.4 ± 25.2 pA, n = 30) at the test potential of -100 mV (bP < 0.01, Figure 1). But the I-V curve relation and reversal potential were not transformed (Figure 2).
Figure 1.
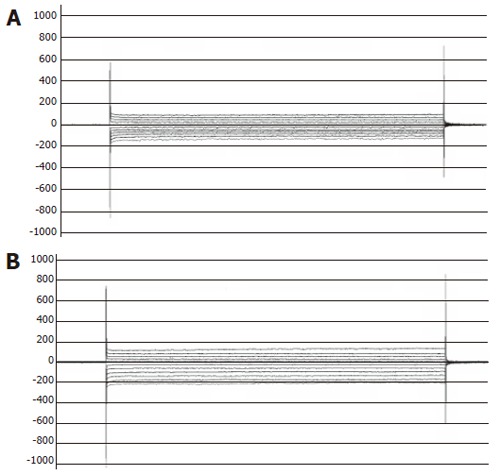
Whole-cell currents measured at the test potential from -120 mV to + 80 mV in sham-operated control (A) and in rats with ischemia/reperfusion injuries (B).
Figure 2.
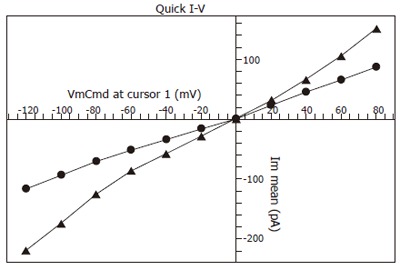
I-V curve of whole-cell currents measured at -100 mV test potential. bP < 0.01 vs sham-operated control (●) (▲ I/R).
Effects of 2-APB on Isoc
In order to observe the effects of 2-APB on Isoc, 2-APB with different concentrations (20, 40, 60, 80, 100 μmol/L) was respectively added to periphery of Kupffer cells after the whole cell configuration was established. According to the whole-cell currents at the test potential of -100mV (Table 1), Isoc was blocked by 2-APB in a concentration-dependent fashion with the IC50 of 37.41 μmol/L. But the I-V curve relation and reversal potential were not transformed (Figure 3A).
Table 1.
Effects of 2-APB on Isoc at the test potential of -100 mV (mean ± SD, n = 8)
| Group (μmol/L) | Peak current (pA) | t | Inhibitive rate (%) |
| Control | -227.8 ± 68.5 | ||
| 2-APB (20) | -139.6 ± 52.9b | 7.6 | 39.0 ± 10.3 |
| 2-APB (40) | -109.6 ± 36.4b | 6.2 | 50.8 ± 12.2 |
| 2-APB (60) | -88.1 ± 37.6b | 6.7 | 61.0 ± 15.1 |
| 2-APB (80) | -72.4 ± 37.1b | 6.8 | 67.7 ± 15.1 |
| 2-APB (100) | -57.8 ± 33.4b | 6.9 | 73.7 ± 14.3 |
P < 0.01 vs control.
Figure 3.
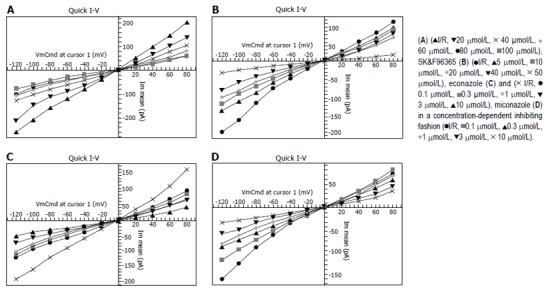
I-V curve of whole-cell currents at -100 mV test potential affected by 2-APB
Effects of SK&F96365 on Isoc
In order to observe the effects of SK&F96365 on Isoc, SK&F96365 with different concentrations (5, 10, 20, 40, 50 μmol/L) was respectively added to periphery of Kupffer cells after the whole-cell configuration was established. According to the whole-cell currents at the test potential of -100mV (Table 2), Isoc was blocked by SK&F96365 in a concentration-dependent fashion with the IC50 of 5.89 μmol/L. But the I-V curve relation and reversal potential were not transformed (Figure 3B).
Table 2.
Effects of SK&F96365 on Isoc at the test potential of -100 mV (mean ± SD, n = 11)
| Group (μmol/L) | Peak current (pA) | t | Inhibitive rate (%) |
| Control | -161.7 ± 58.7 | ||
| SK&F96365 (5) | -82.6 ± 29.5b | 5.3 | 46.1 ± 17.3 |
| SK&F96365 (10) | -57.0 ± 24.9b | 6.1 | 61.2 ± 20.5 |
| SK&F96365 (20) | -47.1 ± 20.5b | 6.5 | 67.2 ± 19.5 |
| SK&F96365 (40) | -35.6 ± 15.4b | 7.1 | 74.8 ± 15.2 |
| SK&F96365 (50) | -25.3 ± 11.9b | 7.7 | 82.1 ± 11.0 |
P < 0.01 vs control.
Effects of econazole on Isoc
In order to observe the effects of econazole on Isoc, econazole with different concentrations (0.1, 0.3, 1, 3, 10 μmol/L) was respectively added to periphery of Kupffer cells after the whole-cell configuration was established. According to the whole-cell currents at the test potential of -100 mV (Table 3), Isoc was blocked by econazole in a concentration-dependent fashion with the IC50 of 0.21 μmol/L. But the I-V curve relation and reversal potential were not transformed (Figure 3C).
Table 3.
Effects of econazole on Isoc at the test potential of -100 mV (mean ± SD, n = 13)
| Group (μmol/L) | Peak current (pA) | t | Inhibitive rate (%) |
| Control | -147.2 ± 35.7 | ||
| econazole (0.1) | -83.5 ± 22.8b | 6.7 | 42.0 ± 15.1 |
| econazole (0.3) | -64.7 ± 18.9b | 8.7 | 54.6 ± 14.7 |
| econazole (1) | -56.6 ± 19.9b | 9.8 | 60.7 ± 13.7 |
| econazole (3) | -47.9 ± 18.2b | 10.3 | 66.5 ± 13.5 |
| econazole (10) | -39.9 ± 15.9b | 11.1 | 71.9 ± 12.6 |
P < 0.01 vs control.
Effects of miconazole on Isoc
In order to observe the effects of miconazole on Isoc, miconazole with different concentrations (0.1, 0.3, 1, 3, 10 μmol/L) was respectively added to periphery of Kupffer cells after the whole-cell configuration was established. According to the whole-cell currents at the test potential of -100 mV (Table 4), Isoc was blocked by miconazole in a concentration-dependent fashion with the IC50 of 0.28 μmol/L. But the I-V curve relation and reversal potential were not transformed (Figure 3D).
Table 4.
Effects of miconazole on Isoc at the test potential of -100 mV (mean ± SD, n = 10)
| Group (μmol/L) | Peak current (pA) | t | Inhibitive rate (%) |
| Control | -133.2 ± 32.0 | ||
| miconazole (0.1) | -86.4 ± 31.7b | 7.1 | 35.6 ± 14.9 |
| miconazole (0.3) | -63.8 ± 23.4b | 10.7 | 52.3 ± 10.0 |
| miconazole (1) | -45.9 ± 13.1b | 9.2 | 64.5 ± 10.6 |
| miconazole (3) | -36.8 ± 11.4b | 9.9 | 71.3 ± 10.2 |
| miconazole (10) | -29.7 ± 10.5b | 9.9 | 76.5 ± 9.4 |
P < 0.01 vs control.
DISCUSSION
In this study, a rat hepatic I/R injury model was established. Kupffer cells were isolated and Isoc in Kupffer cells were detected.
Isoc can be activated by two ways, one is active way induced by IP3[6-8], and the other way is EGTA induced by thasigargin[9]. The latter was chosen in our study.
Hepatic I/R injury can cause calcium overload in liver cells[10-12]. Kupffer cells are considered to play a major role in hepatic I/R injury. The hepatic partial I/R process leads to activation of kupffer cells in ischemic and nonischemic areas of the liver, superoxide generation and proinflammatory cytokine production in Kupffer cells[13]. An excess of reactive oxygen species (ROS) is generated by Kupffer cells activated during hepatic I/R injuries. All these events cause pathophysiological changes initiating a cascade of hepatocellular injury, necrosis, apoptosis, and subsequent inflammation[14-16]. Inhibiting activation of Kupffer cells can relieve hepatic I/R injuries. Calcium overload is one of the important reasons for activation of Kupffer cells. In our study, I/R injuries could significantly increase Isoc of the Kupffer cells, suggesting that SOC has a close relation with hepatic I/R injuries and is the main channel for the influx of Ca2+ during calcium overload. Broad et al[17] showed that phospolipase C and polyphosphoinositides can activate capacitative calcium entry (CCE). Recent findings indicate that receptor-mediated activation of phospholipase C in intact cells activates TRPC3 diacylglycerol production, independently of G proteins, protein kinase C, or inositol 1, 4, 5-trisphosphate[18]. The last source of ion channel is the transient receptor potential (TPR) channel family, which forms non-selective cations[19] all these indicate that hepatic I/R injuries activate phospholipase C which induces activation of store-operated calcium channels or TRPC3 channels, finally leading to calcium overload of Kupffer cells which could exacerbate hepatic I/R injuries.
Sequentially, Isoc of Kupffer cells after hepatic I/R injuries could be blocked by 2-APB, SK&F96365, econazole and miconazole in a concentration-dependent fashion.
2-APB has the restraint effect on Isoc in many kinds of cells[20,21] and is a blocker of SOC and TRP channels[22-24]. Our data have shown that Isoc of Kupffer cells after hepatic I/R injuries is blocked by 2-APB in a concentration-dependent fashion from 20 μmol/L to 100 μmol/L. 2-APB inhibits SOC by IP3 way[25,26]. Recent studies indicate that 2-APB may act as a direct blocker rather than as an IP3 receptor antagonist while Icrac activity is rapidly blocked by extracellular 2-APB, but not by intracellular 2-APB[27-30]. Furthermore, Broad et al[17] showed 2-APB abolishes CCE induced by thapsigargin even in DT40 cells deficient for all isoforms of IP3 receptor, consistent with a direct action of 2-APB on the SOC themselves. Our data also suggest that 2-APB may act as a direct blocker for SOC. Therefore, 2-APB protects Kupffer cells against being activated after hepatic I/R injuries by blocking Isoc.
SK&F96365 could block Isoc in many cells such as HL-60 cells, thyroid gland FRTL-5 cells, thrombocytes as well as voltage-dependent Ca2+ channels (VDCCs) in GH3 pituitary cells and smooth muscle cells[31], suggesting that SK&F96365 can be used as a blocker of SOC in non-excited cells without VDCCs. VDCCs have been proved to be Kupffer cells, but Ca2+ current induced by EGTA and thapsigargin could not be blocked by verapamil (a kind of blocker of VDCCs). In our study, Isoc of Kupffer cells after hepatic I/R injuries was blocked by SK&F96365 in a concentration-dependent fashion. The mechanism is not completely clear. Since SK&F96365 could block two different channels, SOC and VDCCs, it may act on the SOC directly.
Econazole and miconazole decrease intracellular calcium levels after activation of SOC channels[32] and block Isoc in Jurkat T-cells, HL-60 leukocytes, HEL cells, etc. Our data demonstrate that Isoc of Kupffer cells after hepatic I/R injuries is blocked by econazole and miconazole in a concentration-dependent fashion.
In conclusion, hepatic I/R injuries can activate Kupffer cells, probably by increasing Isoc in Kupffer cells and activated Kupffer cells exacerbate hepatic I/R injuries. 2-APB, SK&F96365, econazole and miconazole can inhibit Isoc of Kupffer cells after hepatic I/R injuries in a concentration-dependent fashion. They have obvious protective effects on I/R injury, probably by inhibiting Isoc of Kupffer cells and preventing activation of Kupffer cells.
Footnotes
Supported by the National Natural Science Foundation of China, No.30270532; Trans-Century Training Programme Foundation for the Talents by the Ministry of Education of China, No. 2002-48; Shuguang Program Project of Shanghai Educational Committee, No.02SG20
S- Editor Pan BR L- Editor Wang XL E- Editor Bi L
References
- 1.Vazquez G, Wedel BJ, Aziz O, Trebak M, Putney JW Jr. The mammalian TRPC cation channels. Biochim Biophys Acta. 2004;1742:21–36. doi: 10.1016/j.bbamcr.2004.08.015. [DOI] [PubMed] [Google Scholar]
- 2.Penner R, Fleig A. Store-operated calcium entry: a tough nut to CRAC. Sci STKE. 2004;2004:pe38. doi: 10.1126/stke.2432004pe38. [DOI] [PubMed] [Google Scholar]
- 3.Putney JW Jr. Store-operated calcium channels: how do we measure them, and why do we care. Sci STKE. 2004;2004:pe37. doi: 10.1126/stke.2432004pe37. [DOI] [PubMed] [Google Scholar]
- 4.Gregory RB, Hughes R, Riley AM, Potter BV, Wilcox RA, Barritt GJ. Inositol trisphosphate analogues selective for types I and II inositol trisphosphate receptors exert differential effects on vasopressin-stimulated Ca2+ inflow and Ca2+ release from intracellular stores in rat hepatocytes. Biochem J. 2004;381:519–526. doi: 10.1042/BJ20040637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Colletti LM, Remick DG, Burtch GD, Kunkel SL, Strieter RM, Campbell DA Jr. Role of tumor necrosis factor-alpha in the pathophysiologic alterations after hepatic ischemia/reperfusion injury in the rat. J Clin Invest. 1990;85:1936–1943. doi: 10.1172/JCI114656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gregory RB, Sykiotis D, Barritt GJ. Evidence that store-operated Ca2+ channels are more effective than intracellular messenger-activated non-selective cation channels in refilling rat hepatocyte intracellular Ca2+ stores. Cell Calcium. 2003;34:241–251. doi: 10.1016/s0143-4160(03)00106-4. [DOI] [PubMed] [Google Scholar]
- 7.Gregory RB, Barritt GJ. Evidence that Ca2+-release-activated Ca2+ channels in rat hepatocytes are required for the maintenance of hormone-induced Ca2+ oscillations. Biochem J. 2003;370:695–702. doi: 10.1042/BJ20021671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Wissing F, Nerou EP, Taylor CW. A novel Ca2+-induced Ca2+ release mechanism mediated by neither inositol trisphosphate nor ryanodine receptors. Biochem J. 2002;361:605–611. doi: 10.1042/0264-6021:3610605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rychkov G, Brereton HM, Harland ML, Barritt GJ. Plasma membrane Ca2+ release-activated Ca2+ channels with a high selectivity for Ca2+ identified by patch-clamp recording in rat liver cells. Hepatology. 2001;33:938–947. doi: 10.1053/jhep.2001.23051. [DOI] [PubMed] [Google Scholar]
- 10.Silomon M, Pizanis A, Rose S. Oxyradical-mediated hepatocellular Ca2+ alterations during hemorrhagic shock and resuscitation. Shock. 1999;11:193–198. doi: 10.1097/00024382-199903000-00007. [DOI] [PubMed] [Google Scholar]
- 11.Sung YJ, Sung Z, Ho CL, Lin MT, Wang JS, Yang SC, Chen YJ, Lin CH. Intercellular calcium waves mediate preferential cell growth toward the wound edge in polarized hepatic cells. Exp Cell Res. 2003;287:209–218. doi: 10.1016/s0014-4827(03)00160-5. [DOI] [PubMed] [Google Scholar]
- 12.Ben Abdennebi H, Steghens JP, Hadj-Aïssa A, Barbieux A, Ramella-Virieux S, Gharib C, Boillot O. A preservation solution with polyethylene glycol and calcium: a possible multiorgan liquid. Transpl Int. 2002;15:348–354. doi: 10.1007/s00147-002-0427-8. [DOI] [PubMed] [Google Scholar]
- 13.Nakamitsu A, Hiyama E, Imamura Y, Matsuura Y, Yokoyama T. Kupffer cell function in ischemic and nonischemic livers after hepatic partial ischemia/reperfusion. Surg Today. 2001;31:140–148. doi: 10.1007/s005950170198. [DOI] [PubMed] [Google Scholar]
- 14.Jambrina E, Alonso R, Alcalde M, del Carmen Rodríguez M, Serrano A, Martínez-A C, García-Sancho J, Izquierdo M. Calcium influx through receptor-operated channel induces mitochondria-triggered paraptotic cell death. J Biol Chem. 2003;278:14134–14145. doi: 10.1074/jbc.M211388200. [DOI] [PubMed] [Google Scholar]
- 15.Lemasters JJ, Qian T, He L, Kim JS, Elmore SP, Cascio WE, Brenner DA. Role of mitochondrial inner membrane permeabilization in necrotic cell death, apoptosis, and autophagy. Antioxid Redox Signal. 2002;4:769–781. doi: 10.1089/152308602760598918. [DOI] [PubMed] [Google Scholar]
- 16.Nieminen AL. Apoptosis and necrosis in health and disease: role of mitochondria. Int Rev Cytol. 2003;224:29–55. doi: 10.1016/s0074-7696(05)24002-0. [DOI] [PubMed] [Google Scholar]
- 17.Broad LM, Braun FJ, Lievremont JP, Bird GS, Kurosaki T, Putney JW Jr. Role of the phospholipase C-inositol 1,4,5-trisphosphate pathway in calcium release-activated calcium current and capacitative calcium entry. J Biol Chem. 2001;276:15945–15952. doi: 10.1074/jbc.M011571200. [DOI] [PubMed] [Google Scholar]
- 18.Trebak M, St J Bird G, McKay RR, Birnbaumer L, Putney JW Jr. Signaling mechanism for receptor-activated canonical transient receptor potential 3 (TRPC3) channels. J Biol Chem. 2003;278:16244–16252. doi: 10.1074/jbc.M300544200. [DOI] [PubMed] [Google Scholar]
- 19.Clapham DE, Runnels LW, Strübing C. The TRP ion channel family. Nat Rev Neurosci. 2001;2:387–396. doi: 10.1038/35077544. [DOI] [PubMed] [Google Scholar]
- 20.Wilson SM, Mason HS, Ng LC, Montague S, Johnston L, Nicholson N, Mansfield S, Hume JR. Role of basal extracellular Ca2+ entry during 5-HT-induced vasoconstriction of canine pulmonary arteries. Br J Pharmacol. 2005;144:252–264. doi: 10.1038/sj.bjp.0706077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tamareille S, Mignen O, Capiod T, Rücker-Martin C, Feuvray D. High glucose-induced apoptosis through store-operated calcium entry and calcineurin in human umbilical vein endothelial cells. Cell Calcium. 2006;39:47–55. doi: 10.1016/j.ceca.2005.09.008. [DOI] [PubMed] [Google Scholar]
- 22.Iwasaki H, Mori Y, Hara Y, Uchida K, Zhou H, Mikoshiba K. 2-Aminoethoxydiphenyl borate (2-APB) inhibits capacitative calcium entry independently of the function of inositol 1,4,5-trisphosphate receptors. Receptors Channels. 2001;7:429–439. [PubMed] [Google Scholar]
- 23.Bootman MD, Collins TJ, Mackenzie L, Roderick HL, Berridge MJ, Peppiatt CM. 2-aminoethoxydiphenyl borate (2-APB) is a reliable blocker of store-operated Ca2+ entry but an inconsistent inhibitor of InsP3-induced Ca2+ release. FASEB J. 2002;16:1145–1150. doi: 10.1096/fj.02-0037rev. [DOI] [PubMed] [Google Scholar]
- 24.Braun FJ, Aziz O, Putney JW Jr. 2-aminoethoxydiphenyl borane activates a novel calcium-permeable cation channel. Mol Pharmacol. 2003;63:1304–1311. doi: 10.1124/mol.63.6.1304. [DOI] [PubMed] [Google Scholar]
- 25.Maruyama T, Kanaji T, Nakade S, Kanno T, Mikoshiba K. 2APB, 2-aminoethoxydiphenyl borate, a membrane-penetrable modulator of Ins(1,4,5)P3-induced Ca2+ release. J Biochem. 1997;122:498–505. doi: 10.1093/oxfordjournals.jbchem.a021780. [DOI] [PubMed] [Google Scholar]
- 26.Wang JP, Tseng CS, Sun SP, Chen YS, Tsai CR, Hsu MF. Capsaicin stimulates the non-store-operated Ca2+ entry but inhibits the store-operated Ca2+ entry in neutrophils. Toxicol Appl Pharmacol. 2005;209:134–144. doi: 10.1016/j.taap.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 27.Ma HT, Patterson RL, van Rossum DB, Birnbaumer L, Mikoshiba K, Gill DL. Requirement of the inositol trisphosphate receptor for activation of store-operated Ca2+ channels. Science. 2000;287:1647–1651. doi: 10.1126/science.287.5458.1647. [DOI] [PubMed] [Google Scholar]
- 28.Braun FJ, Broad LM, Armstrong DL, Putney JW Jr. Stable activation of single Ca2+ release-activated Ca2+ channels in divalent cation-free solutions. J Biol Chem. 2001;276:1063–1070. doi: 10.1074/jbc.M008348200. [DOI] [PubMed] [Google Scholar]
- 29.Kukkonen JP, Lund PE, Akerman KE. 2-aminoethoxydiphenyl borate reveals heterogeneity in receptor-activated Ca(2+) discharge and store-operated Ca(2+) influx. Cell Calcium. 2001;30:117–129. doi: 10.1054/ceca.2001.0219. [DOI] [PubMed] [Google Scholar]
- 30.Lievremont JP, Bird GS, Putney JW Jr. Mechanism of inhibition of TRPC cation channels by 2-aminoethoxydiphenylborane. Mol Pharmacol. 2005;68:758–762. doi: 10.1124/mol.105.012856. [DOI] [PubMed] [Google Scholar]
- 31.Merritt JE, Armstrong WP, Benham CD, Hallam TJ, Jacob R, Jaxa-Chamiec A, Leigh BK, McCarthy SA, Moores KE, Rink TJ. SK&F 96365, a novel inhibitor of receptor-mediated calcium entry. Biochem J. 1990;271:515–522. doi: 10.1042/bj2710515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Harper JL, Shin Y, Daly JW. Loperamide: a positive modulator for store-operated calcium channels. Proc Natl Acad Sci USA. 1997;94:14912–14917. doi: 10.1073/pnas.94.26.14912. [DOI] [PMC free article] [PubMed] [Google Scholar]