

Abstract
AIM: To investigate reactivated Epstein-Barr virus (EBV) infection as a cause for chronic hepatitis.
METHODS: Patients with occasionally established elevated serum aminotransferases were studied. HIV, HBV and HCV-infections were excluded as well as any other immunosuppressive factors, metabolic or toxic disorders. EBV viral capsid antigen (VCA) IgG and IgM, EA-R and EA-D IgG and Epstein-Barr nuclear antigen (EBNA) were measured using IFA kits. Immunophenotyping of whole blood was performed by multicolor flow cytometry. CD8+ T cell responses to EBV and PHA were determined according to the intracellular expression of IFN-γ.
RESULTS: The mean alanine aminotransferase (ALT) and gamma glutamyl transpeptidase (GGTP) values exceeded twice the upper normal limit, AST/ALT ratio < 1. Serology tests showed reactivated EBV infection in all patients. Absolute number and percentages of T, B and NK cells were within the reference ranges. Fine subset analysis, in comparison to EBV+ healthy carriers, revealed a significant decrease of naive T cells (P < 0.001), accompanied by increased percentage of CD45RA- (P < 0.0001), and terminally differentiated CD28-CD27-CD8+ T cells (P < 0.01). Moderately elevated numbers of CD38 molecules on CD8+ T cells (P < 0.05) proposed a low viral burden. A significantly increased percentage of CD8+ T cells expressing IFN-γ in response to EBV and PHA stimulation was registered in patients, as compared to controls (P < 0.05). Liver biopsy specimens from 5 patients revealed nonspecific features of low-grade hepatitis.
CONCLUSION: Chronic hepatitis might be a manifestation of chronic EBV infection in the lack of detectable immune deficiency; the expansion of CD28-CD27- and increase of functional EBV-specific CD8+ T cells being the only surrogate markers of viral activity.
Keywords: Chronic hepatitis, Epstein-Barr, Epstein-Barr virus-specific CD8+ T cell
INTRODUCTION
Epstein-Barr virus (EBV) infects more than 90% of humans by the time of adulthood and persists as a life-long latency, suggesting sophisticated mechanisms for effective immune evasion. Primary EBV infection is usually asymptomatic or results in infectious mononucleosis, which resolves spontaneously after the emergence of EBV-specific immunity. Similar to other herpes viruses, EBV persists in the infected immunocompetent organism, without causing apparent disease. CD8+ T cell-mediated responses control the virus both during primary infection and the following carrier state[1,2].
So far, studies on the interaction between the immune system and EBV have focused mostly on congenital or acquired immune deficiency states, which can lead to viral reactivation. Rare cases of fulminant chronic EBV infection have been described and termed severe chronic active EBV infection (SCAEBV or CAEBV)[3,4]. Criteria of CAEBV include severe infection lasting more than 6 mo, histologic evidence of major organ involvement, abnormal EBV antibody titers, and increased quantities of EBV in affected tissues in apparently immunocompetent hosts[5]. However, congenital or acquired defects of cell-mediated immune mechanisms in CAEBV patients are very probable, associated with signalling defects, reduced T-cell or NK cell activity, or reduced perforin levels[6].
While hepatitis is a common feature of primary EBV infection[7,8], the role of EBV in chronic liver disease is less obvious. Bertolini et al showed typical histopathological lesions of chronic hepatitis without involvement of other organs after inoculation of normal human bone marrow-derived B cell, carrying an endogenous EBV line in vivo in nu/nu mice[9]. Single cases of persistent hepatitis related to EBV infection were reported in immunocompromised persons[10]. Liver injury may be one of the manifestations of CAEBV[11-13]. However, only limited knowledge exists about EBV-related hepatitis in immunocompetent patients. We analyzed the clinical and laboratory findings in 15 patients with chronic hepatitis and no data of immune deficiency. Based on the serological profile of reactivated EBV infection and the absence of other major etiological agents, we propose reactivated EBV infection as a cause for the chronic liver disease. Furthermore, we demonstrate that the only significant changes at the level of peripheral blood lymphocytes were the prevalence of CD45RA- T cells, combined with expansion of CD8+ T cells lacking CD27 and CD28 expression, and an increased percentage of lytic EBV epitope-specific CTL.
MATERIALS AND METHODS
Patients
Fifteen Caucasian HLA-A2+ patients (11 males, 4 females, mean age 40.6 years (range 19-55) with chronic hepatitis, referred to the Clinic of Gastroenterology, Medical Institute, Ministry of Interior, Sofia between Sep 2004 and Mar 2005, were included in this study. Chronic liver injury was detected during routine prophylactic examination and was defined as an elevation of serum aminotransferases of more than 6 mo duration. All patients were HIV-1, HBV and HCV-negative, had normal serum protein levels, and had no evidence of metabolic, or alcohol-related disorders, or drug toxicity (Table 1). Furthermore, they had no history of prior immune deficiency or of any other recent, recurring or chronic infection, or immunosuppressive factors that might explain the observed pathological changes.
Table 1.
Exclusion criteria
| Alcohol consumption > 40 mL per day |
| Diabetes or impaired glucose tolerance |
| (including hyperinsulinaemia, OGTT and fasting blood glucose) |
| Under- or overweight (BMI < 18 or > 26 kg/m2) |
| Dyslipidaemia |
| Hyper- or hypothyroidism |
| Gluten enteropathy |
| Autoimmune hepatitis or PBC/PSH |
| Viral hepatitis B or C |
| Hereditary hemochromatosis or Iron overload |
| Wilson disease |
| Anamnesis for hepatotoxic drugs use (including herbal and over-the-counters in the last 12 mo) |
| Hereditary muscular disorders |
A control group included 15 HLA-A2+ EBV+ sex- and age-matched healthy volunteers with normal aminotransferase levels. All samples were obtained after informed consent.
Serological and routine laboratory tests
EBV viral capsid antigen (VCA) IgG and IgM, EA-R and EA-D IgG and Epstein-Barr nuclear antigen (EBNA) were measured, using immunofluorescent assay (IFA FDA Atlanta kits). HIV-1, HBsAg, anti-HBc and anti-HCV were tested with ELISA method. Serum aspartate aminotransferase (AST), alanine aminotransferase (ALT), gamma-glutamyl transpeptidase (gamma-GT, GGTP), creatinine, bilirubin, glucose levels, serum proteins and coagulation were measured according to standard laboratory procedures.
Immunophenotypic studies
Whole blood was obtained in heparinized vacutainer tubes. Lymphocyte subset absolute counts were determined by a lysis/no wash procedure with TruCOUNT tubes and CD3/CD4/CD45/CD8 MultiTest; T cell subsets were evaluated by multicolor immunophenotyping and a standard lysis/wash technique (Table 2). The quantitative expression of CD38 on CD8+ T cells (CD38 ABC) was assessed by the QUANTIBRITE PE fluorescence quantitation kit, according to the manufacturer’s instructions and analyzed with QUANTIBRITE software. All reagents were products of BD Biosciences (B-D, San Jose, CA). At least 5000 lymphocytes were collected per sample and analysed using FACSCalibur flow cytometer and CellQuest software (B-D).
Table 2.
T cell subsets analyzed in this study
| mAb combination | Subset | Phenotype |
| CCR7/CD45RA/ CD8/CD4 | Naïve CD8 | CD8+CD45RA+CCR7+ |
| Antigen-primed/memory CD8 | CD8+CD45RA- | |
| Effector CD8 | CD8+CD45RA+CCR7- | |
| Naïve CD4 | CD4+CD45RA+CCR7+ | |
| Antigen-primed/memory CD4 | CD4+CD45RA- | |
| CD27/CD28/CD8 | Early CD8 | |
| Intermediate CD8 | CD8+CD27+CD28+ | |
| Terminally differentiated CD8 | CD8+CD27+CD28- |
Intracellular cytokine analysis
The HLA-A2 restricted epitope GLCTLVAML from the EBV lytic cycle protein BMLF1 was synthesized according to the standard Fmoc protocol[14]. EBV peptide (10 mg/L) or PHA (Sigma) were used for overnight stimulation of 250 μL whole blood (at 37°C and humidified 5% CO2 atmosphere). Two hours later 12.5 mg/L Brefeldin A (BD) was added. Cells were further washed in cold phosphate buffered saline (PBS), incubated with EDTA for 10 min, washed again and processed according to the protocol for surface and intracellular staining. For intracellular staining, 100 μL of stimulated blood was lysed with 1 mL FACSTM Lysing solution (BD) followed by permeabilization with 0.5 mL 1 × FACS permeabilizing solution (BD) for 10 min. After washing with PBS containing 0.5% BSA, a simultaneous staining for IFN-γ/CD69/CD8/CD4 was performed for 30 min at room temperature in the dark. For analysis of intracellular staining at least 25 000 CD8high cells were collected. Detection of at least 0.05% IFN-γ+CD69+CD8high cells, after subtraction of non-stimulated control values was considered as significant response.
Liver biopsy and imaging
All patients underwent conventional abdominal ultrasono-graphy. Liver biopsy specimens were obtained from 5 patients.
Statistical analysis
Man-Witney non-parametric test was used to evaluate differences between patient and control groups. P values less than 0.05 were considered statistically significant.
RESULTS
Clinical findings
The clinical characteristics of each patient are summarized in Table 3. The estimated mean disease duration before the prophylactic examination was 10.8 (min 6, max 36) mo. At the time of diagnosis of the chronic liver disease none of the 15 patients had an infectious mononucleosis-like illness or any other significant complaints except for one, who reported periods of sub-febrile temperature. None of the patients reported previous opportunistic infections or other significant chronic diseases, or congenital immunodeficiency, or immunosuppressive treatment. The average score of Karnofski performance status was 95%[15].
Table 3.
Characteristics of the patients with chronic hepatitis related to Epstein-Barr virus infection
| Pt No. | Age (yr) | Sex | Duration of illness (mo) | AST | ALT | ALP | GGT | Serology EBV (IFA) | ||||
| VCA IgG | VCA IgM | EA-R IgG | EA-D IgG | EBNA | ||||||||
| 1 | 40 | m | 24 | 52 | 90 | 300 | 200 | 320 | 10 | 80 | 80 | 0 |
| 2 | 60 | m | 6 | 50 | 187 | 280 | 70 | 160 | 0 | 160 | 160 | 40 |
| 3 | 27 | m | 6 | 31 | 107 | 200 | 20 | 640 | 0 | 160 | 160 | 40 |
| 4 | 40 | m | 6 | 30 | 60 | 300 | 40 | 160 | 0 | 160 | 160 | 40 |
| 5 | 57 | f | 24 | 66 | 84 | 280 | 110 | 320 | 0 | 160 | 160 | 40 |
| 6 | 55 | f | 6 | 66 | 85 | 850 | 171 | 160 | 0 | 80 | 80 | 40 |
| 7 | 28 | m | 6 | 40 | 96 | 200 | 60 | 160 | 10 | 80 | 80 | 20 |
| 8 | 38 | m | 12 | 60 | 60 | 150 | 130 | 640 | 0 | 80 | 80 | 20 |
| 9 | 30 | m | 6 | 50 | 125 | 250 | 180 | 640 | 0 | 80 | 80 | 20 |
| 10 | 25 | m | 6 | 20 | 60 | 225 | 90 | 160 | 0 | 80 | 80 | 20 |
| 11 | 35 | m | 6 | 40 | 80 | 136 | 180 | 160 | 0 | 80 | 80 | 40 |
| 12 | 31 | m | 6 | 50 | 80 | 310 | 80 | 320 | 0 | 160 | 160 | 40 |
| 13 | 43 | m | 10 | 60 | 100 | 300 | 100 | 320 | 10 | 160 | 160 | 0 |
| 14 | 50 | f | 6 | 100 | 200 | 130 | 200 | 640 | 0 | 160 | 160 | 40 |
| 15 | 50 | f | 6 | 80 | 120 | 400 | 200 | 320 | 0 | 160 | 160 | 40 |
Laboratory tests showed normal blood counts. Mean values (95% CI) are as follows: white blood cell count, 6.61 × 109/L (5.945 to 7.281); red blood cells 5.08 × 106/L (4.7893 to 5.3707); platelets, 243 × 109/L (207.31 to 277.89). The mean range of serum ALT at the time of diagnosis was 103 UI/mL (UNL = 40) (SD 40.9, SE 10.57), AST/ALT ratio < 1, gamma-GT 122 UI/mL (UNL = 48) (SD 62.41, SE 16.12). Normal serum protein and albumin levels were established in all patients, and no hyperbilirubinaemia or coagulation disturbances were found. Fasting glucose levels, as well as the glucose load test were within the reference ranges. A serum insulin level in 12 patients was below upper limit, while 3 patients were not tested. The abdominal ultrasonography series revealed normal liver and spleen image.
EBV-serology at the time of diagnosis
The EBV-specific antibody titers of each patient are shown in Table 2. Serology tests indicated a reactivated EBV infection in all 15 patients. The average EBV VCA IgG titer was 320 UA/mL (160-640), EBNA average titer was 20 UA/mL (10-40), and EA-R and EA-D were elevated (80-160). EBV VCA IgM was not detected in any patient. All controls were EBV VCA IgM-negative.
Lymphocyte subset analysis
The percentage and absolute counts of total lymphocytes, T, B and NK cells, CD4+ T cells as well as the CD4/CD8 ratio were within the established reference ranges for healthy age-matched controls. Using the mAb combinations CD45RA/CCR7/CD8/CD4 and CD27/CD8/CD28, naïve, central and effector memory, as well as terminally differentiated effector T cells (CTL) were defined (Table 2). The fine subset analysis of T cells revealed a significant decrease of naïve (CD45RA+CCR7+) CD4+ (P < 0.05) and CD8+ (P < 0.01) T cells accompanied by an increased percentage of memory (CD45RA-) CD4+ (P < 0.01) and CD8+ T cells (P < 0.001), as well as of the terminally differentiated (CD28-CD27-) CTL subset (P < 0.01) (Figure 1A-E). A moderately increased number of CD38 molecules on CD8+ T cells was established (mean, 2150; min, 1090; max, 4413) as compared to controls (mean, 1522; min, 510; max, 2210; P < 0.05), most probably corresponding to a low level viral replication.
Figure 1.
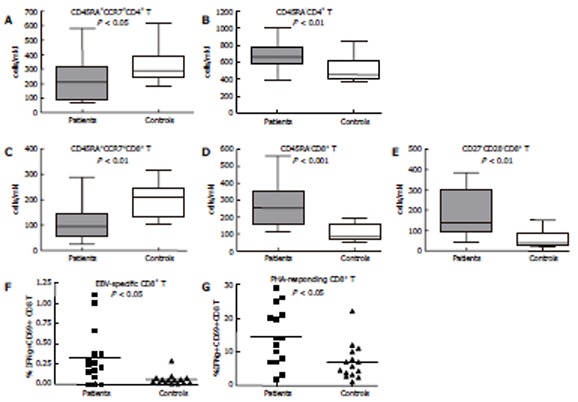
Significant differences detected between patients and control group by immunophenotyping. Patients were characterized by a decreased absolute number of naive CCCR7+CD45RA+CD4+ (A) and CD8+ (C) T cells, increased absolute number of memory CD45RA-CD4+ (B) and CD8+ (D)T cells, increased absolute number of effector CD28-CD27-CD8+ T cells (E), as well as by increased percentage of IFNγ+CD69+CD8+ T cells after stimulation with EBV peptide GLCTMVL (F) and with PHA (G). Box-and-whiskers indicate the 25-75 percentile and min-max values for each group. Mean values are designated on all plots by a horizontal line. Man-Witney. P values are indicated for each comparison.
Circulating CD8+ T cells, expressing IFN-γ in response to the lytic BMLF1 epitope were registered in all patients, and their percentage was significantly increased (15/15; mean, 0.39%), as compared to EBV-seropositive donors (6/15; mean, 0.06%; P < 0.05). The percentage of PHA-responding CD8+ T cells was also significantly increased in the patients group in comparison to controls (14.5% vs 7.8%, P < 0.05) (Figure 1F and G).
Histopathology
Liver biopsy specimens revealed non-specific histo-pathologic features of low-grade hepatitis. On routine HE slides mild portal and periportal mononuclear infiltrates were seen, composed of small lymphocytes lacking significant cytologic atypia. According to the METAVIR system all patients were staged as A2. No fibrosis was found in any patient except in one which was graded F1 (Figure 2).
Figure 2.
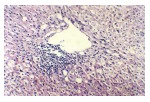
Liver biopsy, showing lymphocyte infiltrate within the lobule and some hepatocyte injury, with mild steatosis displayed in some cells.
DISCUSSION
EBV infects the majority of the human population and persists in the host organism for life. CD8+ T lymphocytes specific for lytic and latent EBV antigens control the virus both during primary infection and the long-term carrier state and almost no symptoms indicate these events[2,16]. Therefore, the development of cellular immune deficiency, related to HIV-infection, transplantation, prolonged immunosuppressive therapy, or even psychological/physical stress is often combined with complications due to EBV reactivation[17]. A broad array of clinical entities has, therefore, been related to EBV infection in immunocompromised host, including persistent and necrotizing hepatitis[18,19]. CAEBV, characterized by a specific pattern of serological and clinical findings, may also induce liver injury[11,12].
We reported the clinical and laboratory features of 15 Caucasian non-related patients with chronic hepatitis in the settings of reactivated EBV infection that had no apparent immune deficiency and did not meet the classical criteria for CAEBV infection. The serological evidence for EBV reactivation was the presence of EBV-NA and EA-R antibodies, combined with elevated but not extreme titers of IgG anti-VCA[20]. None of the patients had significant clinical manifestation of the infection, except for one. Although we did not determine plasma and tissue viral load, the moderately elevated CD38 expression on CD8+ T cells suggested rather a low-level of viral replication[21]. In fact, reactivated EBV infection cases have been reported with mild to moderate clinical presentation, and without extreme EBV-specific Ab titers or viral load[3]. Our data further suggest that the reactivation of EBV infection in the settings of a functional immune system may be more frequent than previously assessed.
Since the latency of EBV infection depends almost entirely on efficient cellular immune responses, phenotypic and functional changes in the lymphocyte compartment might be expected in case of reactivation. CAEBV is usually characterized by oligoclonal expansions of either T or NK cells[22,23]. Although we detected no significant changes in the main circulating lymphocyte populations, the fine subset analysis revealed a significant prevalence of antigen-primed (CD45RA-) over naïve (CD45RA+CCR7+) CD4+ and CD8+ T cells, combined with increased percentage of terminally differentiated (CD28-CD27-) CD8+ T cells. In general, the activation and differentiation state of the circulating T cell pool reflects any major antigen-driven process in the organism. T-cell activation induces a shift from CD45RA to CD45RO expression, while long-lasting chronic viral infections like HIV-1 or CMV are characterized with prevalence of terminally differentiated non-proliferating effectors in the CD8+ T cell compartment[24,25], similar to the natural age-related senescence of cellular immunity[26]. Moreover, particular viruses may be associated with specific phenotypic changes in latency, as the CMV-driven expansion of CD45RA+CD27-CD8+ T cells[27]. Logically, such a phenotypic shift must involve both virus-specific and non-specific by-stander activation and differentiation.
The EBV-specific memory pool in asymptomatic carriers is reported to be heterogeneous: CD8+ T cells specific for latent epitopes being mostly CD45RA-CD28+, while those, specific for lytic epitopes, CD45RA+/CD45RA-CD28-CD27+[28]. CD27 is a co-stimulatory molecule promoting the survival of activated CD8+ T cells in the absence of CD28[29], with a limited reexpression capacity. CD27 probably enhances EBV-specific responses, as most of the EBV-specific cytotoxic activity was reported in the CD27+ subpopulation[30]. In our patients group we established a significant expansion of CD8+ T cells lacking both CD28 and CD27. We may speculate that in case of long lasting or repeated low-level EBV reactivation, the CD27+ pool would be gradually replaced by CD27- CD8+ T cells, less efficient in controlling the persistent EBV infection.
EBV-specific cells detected by tetramer staining in latency account for 1%-3% of the circulating CD8+ T cell pool[31]. Again by tetramer staining, a similar detection rate of lytic epitope-specific CD8+ T cells has been reported in CAEBV and asymptomatic EBV carriers[16]. In our series, patients with EBV-related hepatitis had a significantly higher lytic epitope-specific CTL response, as compared to controls, combined with a higher response to non-specific stimulation, as estimated by IFN-γ expression. These results indicate that the identification of functional EBV-specific CD8+ T cells may better differentiate between asymptomatic carriers and clinically unapparent but active infection.
Several viruses have been implicated in the develop-ment of chronic liver disease, either as single agents or via cross-activation of T cells. A close association between liver lesions and a previous EBV infection has been demonstrated, as well as the presence of EBV-positive cells in affected livers in the settings of continuous low level EBV replication[32,33]. Sugawara et al[34-36] linked EBV to HCV-related hepatocellular cancerogenesis, supporting the EBNA-1 promoted HCV replication hypothesis. EBV was suggested to be involved in the hepatocellular carcinogenesis, although this role is still controversial[37]. Our patients had serological evidence for EBV reactivation and, on the other hand, immunophenotypic evidence for persisting low-level cellular immunity activation and chronic liver inflammation. As the mechanism of EBV-related liver damage remains unclear, indirect, cytokine-induced hepatocellular injury is being discussed[38,39]. Despite the lack of typical mononucleosis characteristics, liver lymphocyte infiltrates in our cases might be involved in the EBV-provoked immune response.
Although based mostly on exclusion criteria, our data suggest that chronic liver disease could be a manifestation of chronic EBV infection, with persistent low-level virus replication or in case of frequent reactivations. It remains to elucidate the underlying mechanisms of inefficient viral control in these immunocompetent patients and whether such patients may further revert to a fulminant course of the infection.
In conclusion, a reactivated EBV infection might be the possible reason for a chronic elevation of serum aminotransferases without a detectable immune deficiency. The prevalence of memory (CD45RA-) and terminally differentiated (CD28-CD27-) CD8+ T cells together with the increased frequency of functionally active EBV-specific CD8+ T cells in the circulating CD8+ T cell compartment might be the only surrogate markers of reactivated viral infection. The identification and follow-up of such patients may help the prevention of classic CAEBV or other severe manifestations of chronic EBV infection.
Footnotes
S- Editor Liu Y L- Editor Zhu LH E- Editor Liu WF
References
- 1.Rickinson AB, Kieff E. Epsterin-Barr virus. In: Knipe DM, Howley PM, editors. Field's Virology, 4th ed, Vol.2. Philadelphia: Lippincott Williams & Wilkins; 2001. pp. 2575–2627. [Google Scholar]
- 2.Cohen JI. Epstein-Barr virus infection. N Engl J Med. 2000;343:481–492. doi: 10.1056/NEJM200008173430707. [DOI] [PubMed] [Google Scholar]
- 3.Kimura H, Hoshino Y, Kanegane H, Tsuge I, Okamura T, Kawa K, Morishima T. Clinical and virologic characteristics of chronic active Epstein-Barr virus infection. Blood. 2001;98:280–286. doi: 10.1182/blood.v98.2.280. [DOI] [PubMed] [Google Scholar]
- 4.Kimura H, Morishima T, Kanegane H, Ohga S, Hoshino Y, Maeda A, Imai S, Okano M, Morio T, Yokota S, et al. Prognostic factors for chronic active Epstein-Barr virus infection. J Infect Dis. 2003;187:527–533. doi: 10.1086/367988. [DOI] [PubMed] [Google Scholar]
- 5.Straus SE. The chronic mononucleosis syndrome. J Infect Dis. 1988;157:405–412. doi: 10.1093/infdis/157.3.405. [DOI] [PubMed] [Google Scholar]
- 6.Katano H, Ali MA, Patera AC, Catalfamo M, Jaffe ES, Kimura H, Dale JK, Straus SE, Cohen JI. Chronic active Epstein-Barr virus infection associated with mutations in perforin that impair its maturation. Blood. 2004;103:1244–1252. doi: 10.1182/blood-2003-06-2171. [DOI] [PubMed] [Google Scholar]
- 7.Kimura H, Nagasaka T, Hoshino Y, Hayashi N, Tanaka N, Xu JL, Kuzushima K, Morishima T. Severe hepatitis caused by Epstein-Barr virus without infection of hepatocytes. Hum Pathol. 2001;32:757–762. doi: 10.1053/hupa.2001.25597. [DOI] [PubMed] [Google Scholar]
- 8.White NJ, Juel-Jensen BE. Infectious mononucleosis hepatitis. Semin Liver Dis. 1984;4:301–306. doi: 10.1055/s-2008-1040659. [DOI] [PubMed] [Google Scholar]
- 9.Bertolini L, Iacovacci S, Bosman C, Carloni G, Monaco V, Bangrazi C, Serafino A, Gualandi G, Prantera G, Fruscalzo A. Low cell dosage of lymphoblastoid human cell lines EBV(+) is associated to chronic hepatitis in a minority of inoculated Nu/Nu mice. J Med Virol. 2002;66:70–81. doi: 10.1002/jmv.2113. [DOI] [PubMed] [Google Scholar]
- 10.Catalina MD, Sullivan JL, Bak KR, Luzuriaga K. Differential evolution and stability of epitope-specific CD8(+) T cell responses in EBV infection. J Immunol. 2001;167:4450–4457. doi: 10.4049/jimmunol.167.8.4450. [DOI] [PubMed] [Google Scholar]
- 11.Yuge A, Kinoshita E, Moriuchi M, Ohno Y, Haga H, Moriuchi H. Persistent hepatitis associated with chronic active Epstein-Barr virus infection. Pediatr Infect Dis J. 2004;23:74–76. doi: 10.1097/01.inf.0000105182.51471.4b. [DOI] [PubMed] [Google Scholar]
- 12.Shibuya A, Tsuchihashi T, Watanabe M, Nakazawa T, Takeuchi A, Sakurai K, Mitomi H, Saigenji K. Severe chronic active Epstein-Barr virus infection associated with multiple necrotic lesions in the liver. Hepatol Res. 2003;25:447–454. doi: 10.1016/s1386-6346(02)00311-x. [DOI] [PubMed] [Google Scholar]
- 13.Ohshima K, Suzumiya J, Sugihara M, Nagafuchi S, Ohga S, Kikuchi M. Clinicopathological study of severe chronic active Epstein-Barr virus infection that developed in association with lymphoproliferative disorder and/or hemophagocytic syndrome. Pathol Int. 1998;48:934–943. doi: 10.1111/j.1440-1827.1998.tb03864.x. [DOI] [PubMed] [Google Scholar]
- 14.Fields GB, Noble RL. Solid phase peptide synthesis utilizing 9-fluorenylmethoxycarbonyl amino acids. Int J Pept Protein Res. 1990;35:161–214. doi: 10.1111/j.1399-3011.1990.tb00939.x. [DOI] [PubMed] [Google Scholar]
- 15.Karnofsky DA, Abelman WH, Craver LF, Burchenal JH. The use of nitrogen mustards in the palliative treatment of carcinoma. Cancer. 1948;1:634–656. [Google Scholar]
- 16.Sugaya N, Kimura H, Hara S, Hoshino Y, Kojima S, Morishima T, Tsurumi T, Kuzushima K. Quantitative analysis of Epstein-Barr virus (EBV)-specific CD8+ T cells in patients with chronic active EBV infection. J Infect Dis. 2004;190:985–988. doi: 10.1086/423285. [DOI] [PubMed] [Google Scholar]
- 17.Maurmann S, Fricke L, Wagner HJ, Schlenke P, Hennig H, Steinhoff J, Jabs WJ. Molecular parameters for precise diagnosis of asymptomatic Epstein-Barr virus reactivation in healthy carriers. J Clin Microbiol. 2003;41:5419–5428. doi: 10.1128/JCM.41.12.5419-5428.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Klein E. The complexity of the Epstein-Barr virus infection in humans. Pathol Oncol Res. 1998;4:3–7. doi: 10.1007/BF02904687. [DOI] [PubMed] [Google Scholar]
- 19.Nicolas JC, Maréchal V, Dehée A. [Epstein-Barr virus] Bull Acad Natl Med. 1997;181:981–996; discussion 996-997. [PubMed] [Google Scholar]
- 20.Sumaya CV. Endogenous reactivation of Epstein-Barr virus infections. J Infect Dis. 1977;135:374–379. doi: 10.1093/infdis/135.3.374. [DOI] [PubMed] [Google Scholar]
- 21.Zidovec Lepej S, Vince A, Dakovic Rode O, Remenar A, Jeren T. Increased numbers of CD38 molecules on bright CD8+ T lymphocytes in infectious mononucleosis caused by Epstein-Barr virus infection. Clin Exp Immunol. 2003;133:384–390. doi: 10.1046/j.1365-2249.2003.02219.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ishii M, Yamaguchi N, Ohshima S, Ishii T, Mori KL, Kimura H, Morishima T, Kawase I, Saeki Y. Possibility of preventive treatment for EBV-associated NK cell-lineage proliferative disorders. Intern Med. 2003;42:250–254. doi: 10.2169/internalmedicine.42.250. [DOI] [PubMed] [Google Scholar]
- 23.Toyabe S, Harada W, Uchiyama M. Biclonal expansion of T cells infected with monoclonal Epstein-Barr virus (EBV) in a patient with chronic, active EBV infection. Clin Exp Immunol. 2003;134:92–97. doi: 10.1046/j.1365-2249.2003.02270.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Gamadia LE, van Leeuwen EM, Remmerswaal EB, Yong SL, Surachno S, Wertheim-van Dillen PM, Ten Berge IJ, Van Lier RA. The size and phenotype of virus-specific T cell populations is determined by repetitive antigenic stimulation and environmental cytokines. J Immunol. 2004;172:6107–6114. doi: 10.4049/jimmunol.172.10.6107. [DOI] [PubMed] [Google Scholar]
- 25.Papagno L, Spina CA, Marchant A, Salio M, Rufer N, Little S, Dong T, Chesney G, Waters A, Easterbrook P, et al. Immune activation and CD8+ T-cell differentiation towards senescence in HIV-1 infection. PLoS Biol. 2004;2:E20. doi: 10.1371/journal.pbio.0020020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nociari MM, Telford W, Russo C. Postthymic development of CD28-CD8+ T cell subset: age-associated expansion and shift from memory to naive phenotype. J Immunol. 1999;162:3327–3335. [PubMed] [Google Scholar]
- 27.Kuijpers TW, Vossen MT, Gent MR, Davin JC, Roos MT, Wertheim-van Dillen PM, Weel JF, Baars PA, van Lier RA. Frequencies of circulating cytolytic, CD45RA+CD27-, CD8+ T lymphocytes depend on infection with CMV. J Immunol. 2003;170:4342–4348. doi: 10.4049/jimmunol.170.8.4342. [DOI] [PubMed] [Google Scholar]
- 28.Hislop AD, Annels NE, Gudgeon NH, Leese AM, Rickinson AB. Epitope-specific evolution of human CD8(+) T cell responses from primary to persistent phases of Epstein-Barr virus infection. J Exp Med. 2002;195:893–905. doi: 10.1084/jem.20011692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ochsenbein AF, Riddell SR, Brown M, Corey L, Baerlocher GM, Lansdorp PM, Greenberg PD. CD27 expression promotes long-term survival of functional effector-memory CD8+ cytotoxic T lymphocytes in HIV-infected patients. J Exp Med. 2004;200:1407–1417. doi: 10.1084/jem.20040717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hislop AD, Gudgeon NH, Callan MF, Fazou C, Hasegawa H, Salmon M, Rickinson AB. EBV-specific CD8+ T cell memory: relationships between epitope specificity, cell phenotype, and immediate effector function. J Immunol. 2001;167:2019–2029. doi: 10.4049/jimmunol.167.4.2019. [DOI] [PubMed] [Google Scholar]
- 31.Tan LC, Gudgeon N, Annels NE, Hansasuta P, O'Callaghan CA, Rowland-Jones S, McMichael AJ, Rickinson AB, Callan MF. A re-evaluation of the frequency of CD8+ T cells specific for EBV in healthy virus carriers. J Immunol. 1999;162:1827–1835. [PubMed] [Google Scholar]
- 32.Santolamazza M, Marinelli RM, Bacosi M, D'Innocenzo S, Miglioresi L, Patrizi F, Delle Monache M, Ricci GL. What kind of hepatitis. J Int Med Res. 2001;29:441–444. doi: 10.1177/147323000102900510. [DOI] [PubMed] [Google Scholar]
- 33.Stránský J, Benda R, Vodák M, Zikmundová L, Honzáková E. Causes of acute exacerbations of chronic hepatitis B. Sb Lek. 1995;96:75–83. [PubMed] [Google Scholar]
- 34.Sugawara Y, Mizugaki Y, Uchida T, Torii T, Imai S, Makuuchi M, Takada K. Detection of Epstein-Barr virus (EBV) in hepatocellular carcinoma tissue: a novel EBV latency characterized by the absence of EBV-encoded small RNA expression. Virology. 1999;256:196–202. doi: 10.1006/viro.1999.9619. [DOI] [PubMed] [Google Scholar]
- 35.Sugawara Y, Makuuchi M, Takada K. Detection of Epstein-Barr virus DNA in hepatocellular carcinoma tissues from hepatitis C-positive patients. Scand J Gastroenterol. 2000;35:981–984. doi: 10.1080/003655200750023075. [DOI] [PubMed] [Google Scholar]
- 36.Sugawara Y, Makuuchi M, Kato N, Shimotohno K, Takada K. Enhancement of hepatitis C virus replication by Epstein-Barr virus-encoded nuclear antigen 1. EMBO J. 1999;18:5755–5760. doi: 10.1093/emboj/18.20.5755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Li W, Wu BA, Zeng YM, Chen GC, Li XX, Chen JT, Guo YW, Li MH, Zeng Y. Epstein-Barr virus in hepatocellular carcinogenesis. World J Gastroenterol. 2004;10:3409–3413. doi: 10.3748/wjg.v10.i23.3409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Collin L, Moulin P, Jungers M, Geubel AP. Epstein-Barr virus (EBV)-induced liver failure in the absence of extensive liver-cell necrosis: a case for cytokine-induced liver dysfunction. J Hepatol. 2004;41:174–175. doi: 10.1016/j.jhep.2004.03.028. [DOI] [PubMed] [Google Scholar]
- 39.Drebber U, Kasper HU, Krupacz J, Haferkamp K, Kern MA, Steffen HM, Quasdorff M, Zur Hausen A, Odenthal M, Dienes HP. The role of Epstein-Barr virus in acute and chronic hepatitis. J Hepatol. 2006;44:879–885. doi: 10.1016/j.jhep.2006.02.006. [DOI] [PubMed] [Google Scholar]