

Abstract
The hepatic sinusoids are lined by a unique population of hepatic sinusoidal endothelial cells (HSEC), which is one of the first hepatic cell populations to come into contact with blood components. However, HSEC are not simply barrier cells that restrict the access of blood-borne compounds to the parenchyma. They are functionally specialised endothelial cells that have complex roles, including not only receptor-mediated clearance of endotoxin, bacteria and other compounds, but also the regulation of inflammation, leukocyte recruitment and host immune responses to pathogens. Thus understanding the differentiation and function of HSEC is critical for the elucidation of liver biology and pathophysiology. This article reviews methods for isolating and studying human hepatic endothelial cell populations using in vitro models. We also discuss the expression and functions of phenotypic markers, such as the presence of fenestrations and expression of VAP-1, Stabilin-1, L-SIGN, which can be used to identify sinusoidal endothelium and to permit discrimination from vascular and lymphatic endothelial cells.
Keywords: Human, Liver, Endothelium, Sinusoid, Phenotype
INTRODUCTION
The liver has a unique dual blood supply receiving blood from both the portal vein and the hepatic artery. Unlike other organs, which are supplied by arterial blood through arterioles, the liver receives venous blood at low pressures through the portal vein as well as arterial blood via the hepatic artery. The intrahepatic portal venous system consists of conducting and distributing systems that ensure blood is carried throughout the parenchyma and evenly delivered to individual hepatocytes via the sinusoidal network[1]. A uniform and strict branching pattern appears at the level of the terminal vessels and is maintained throughout the distributing system. The first order branches of the distributing system arise from the terminal branches of the conducting system and give rise to second order vessels of approximately 70 μm diameter that correspond to the terminal portal vein branches seen in portal tracts. Third order vessels arise from these and correspond to the septal or interlobular branches. These exhibit a classical sinusoidal appearance, lack a connective tissue sheath and have no basement membrane. The distributing system then branches into the hexagonal lobule before draining into the hepatic vein. The hepatic artery supplies 25% of hepatic blood flow and provides blood to both the parenchyma and the portal tracts. Axial hepatic arterial branches run parallel to the conducting portal veins and terminate in inlet venules, terminal portal veins and the sinusoids, thereby supplying blood to the parenchyma. The axial arteries also give rise to peribiliary branches that supply the accompanying bile ducts and portal interstitium. These arteries form the peribiliary plexus consisting of efferent and afferent capillaries that wrap around the surface of bile ducts. Small channels from the peribiliary plexus drain into the sinusoids or portal vein branches. Thus, complex anastomoses exist between the axial arteries and peribiliary arteries[2].
ISOLATION AND CULTURE OF HEPATIC SINUSOIDAL ENDOTHELIAL CELLS
Isolated cultures of hepatic sinusoidal endothelial cells present a valuable tool for the study of liver physiology and pathophysiology. Most published studies used cells isolated from rodent livers and have defined the role of fenestrations and specific receptors in uptake and processing of circulating factors including pathogens (reviewed in[3]). Most investigators use a combination of enzymatic digestion and density gradient centrifugation to isolate HSEC from liver tissue although there is considerable variation in the protocols used by different groups and for cells from different species. HSEC from rodents are commonly isolated by enzymatic digestion, either by perfusion of an intact organ with an enzymatic cocktail or by mechanical disruption followed by enzymatic digestion[4-7]. The cell suspension generated by such methods is fractionated using differential centrifugation techniques, including counterflow elutriation or density gradient centrifugation. Endothelial cells are then grown on matrix coated flasks in selective growth media containing growth factors.
Similar methods can be used to isolate HSEC from human liver samples[8-10] and between 103-106 cells have been isolated from whole livers unsuitable for transplantation[9]. However, the limited access to intact human liver means that most groups use diseased tissue removed at transplantation or surgical resection or biopsy specimens that generate low numbers of cells per isolation. Nevertheless, successful isolation is aided by the large numbers of HSEC relative to other vascular endothelial cells present in the liver. Some researchers further reduce the potential of contaminating vascular endothelium by excising visible large vascular structures from the liver tissue prior to enzymatic digestion. Although several different isolation procedures have been described these are not all equally effective in our hands. Our experience, and that of other groups, suggests that counterflow elutriation alone is not useful for selection of human HSEC from a mixed non-parenchymal cell preparation as most of the cells have similar centrifugal densities[6,11]. This led us to routinely include a step of immunomagnetic depletion to remove common contaminating cell populations, such as biliary epithelial cells, followed by positive selection of endothelial cells using antibody against CD31 (see later)[10]. Similar methods using magnetic beads coated with Evonymus europaeus agglutinin have been used to isolate HSEC from the liver of other primates[12] and there are now commercial antibody-based magnetic kits for isolation of rodent HSEC. Despite these refinements there is considerable variability in the yield and viability of cells obtained from diseased human liver tissue. Thus, although there are compelling reasons to study human HSEC in vitro low cell yields mean that it is difficult to use primary cells without passage.
PROBLEMS WITH EXISTING PHENOTYPIC MARKERS AND USE OF HSEC IN MONO-CULTURE
As scientific technology advances it is possible to carry out increasingly complex genetic, proteomic and functional analyses on cells grown in culture. However such techniques require highly purified populations of cells of defined phenotype. Some of the methods previously used to confirm HSEC purity and phenotype, such as AcLDL uptake[8] and binding of Ulex lectin[8], are not specific. For example, other hepatic cell populations, including dendritic cells, take up AcLDL and Ulex lectin binds to fucosylated receptors on both Kupffer cells and HSEC[13]. Similarly, endothelial cells share many cell surface receptors with leukocytes, including CD31, CD4[14], CD11b, and CD11c[11], which may contaminate endothelial cell preparations in culture. Care also needs to be taken when using antibody staining to define HSEC phenotype because HSEC express high levels of FcγR[15] allowing them to bind antibodies non-specifically. In light of such problems, it has been suggested that the presence of open fenestrations arranged in sieve plates is the only true marker of hepatic sinusoidal endothelial cells[16]. These pores are indeed classic features of liver sinusoidal endothelial cells in vivo but present problems when used to identify cells in vitro (see below).
All of these problems are compounded by the fact that HSEC are most commonly cultured as a monolayer of cells on matrix-coated tissue culture plates in vitro. This perturbs the normal morphology of the cells and they become flattened and rapidly lose fenestrations. Part of this effect may be the loss of paracrine signals from other cells of the sinusoid that maintain the phenotype and differentiation of HSEC in vivo. For example, crosstalk between hepatic sinusoidal endothelial cells and closely juxtaposed hepatocytes is essential for the maintenance of sinusoidal endothelial cell growth and differentiation. This is demonstrated by studies where implantation of foetal liver fragments into quail chorioallantoid membrane resulted in the acquisition of a sinusoidal phenotype by the chorioallantoid microvessels[17] and also in vitro studies where co-culture of HSEC with other liver cells resulted in a more stable endothelial phenotype and function[18]. Thus, markers used to determine phenotypes of HSEC must take into account alterations in phenotype as a consequence of culturing cells in isolation in the absence of local paracrine signals.
FENESTRATIONS
Endothelial cells throughout the adult organism are derived from common early embryological precursors and have broadly similar functions and histological appearance. However, there is important organ and tissue-specific heterogeneity that results in phenotypic and functional variations (reviewed in[19]). For example, high endothelial venules in lymph nodes are lined by morphologically and phenotypically distinct endothelial cells that have the unique ability to promote the recruitment of naïve lymphocytes whereas lymphatic endothelium express several receptors that allow uptake of macromolecules found in lymph[20]. Sinusoidal endothelial cells are found in the spleen and bone marrow, as well as in the liver, and in all these sites they have a minimal basement membrane and lack classical tight junctions. Hepatic sinusoidal endothelium differs from sinusoidal endothelium in these other beds by its discontinuous nature, being interspersed with kupffer cells and by the presence of open fenestrations arranged in sieve plates[16].
The vascular architecture in the human liver develops by 17-25 wk of gestation, and the sinusoids acquire their distinctive fenestrated phenotype by wk 20 (reviewed in[21]). The fenestrations act as a ‘dynamic filter’[3] allowing macromolecules in blood controlled access to parenchymal cells[22]. Evidence from animal studies suggests that fenestrations constitute up to 40% of the cell and that the size, distribution and clustering of the pores in sieve plates varies with the zonal distribution of the endothelium[23] and across the endothelial surface. Although normal hepatic sinusoidal cells in most mammals are characterised by the presence of fenestrations (reviewed in[3]), caution must be exercised when translating these observations to human cells. Studies of fenestrations in human liver samples are rare, and suggest that the number and size of fenestrations differs from that observed in other mammals[3]. Furthermore, the number of fenestrations per endothelial cell decreases in disease[24,25], following viral infection[22] or with ageing[26]. During cirrhosis and chronic hepatitis, HSEC develop a more vascular morphology and produce a basement membrane in a process known as ‘capillarisation’ (reviewed in[27]). This is associated with increased expression of CD31 and VCAM-1 and loss of fenestrations[27]. These changes may impede the transfer of materials to and from the parenchyma and contribute towards regional hepatocyte hypoxia. Fenestrations are not unique to hepatic EC but are found in endothelium in endocrine glands, kidney, gastrointestinal tract, choroid plexus, lymphatic organs such as the spleen and are sometimes observed in tumour vasculature. Many studies have implicated VEGF as an essential factor for regulation of fenestrations in these organs (reviewed in[28]).
Thus considerable variations in the number, size and localisation of fenestrations are seen among species and also in health and disease. The situation becomes more complex when cells are removed from the hepatic microenvironment and cultured in vitro. The fenestrations documented in freshly isolated rat HSEC begin to disappear within 48 h of cell culture[29] and are almost gone within a week[30]. We have made similar observations with human cells from normal livers and also find very few fenestrated cells when HSEC are isolated from cirrhotic livers (Lalor and Adams unpublished observations). However, the number of fenestrations on rat HSEC can be maintained in vitro by the addition of VEGF and by culturing cells on extracellular matrix constituents, such as collagen, that are secreted by endothelial cells[29,31]. Both human and rodent HSEC need growth factors and attachment to appropriate extracellular matrix molecules to survive and will rapidly undergo apoptosis in the absence of these. Thus in order to maintain cell survival, cultured HSEC must be grown in the presence of VEGF which induces and maintains expression of fenestrations as well as promoting HSEC proliferation. VEGF is also a growth factor for vascular endothelial cells which induces the production of matrix molecules essential for survival and proliferation[32]. At higher concentrations, VEGF can induce the formation of fenestrations in vascular endothelial cells[28,32,33] and, although these pores are not organised into sieve plates, they can be very difficult to distinguish from the fenestrations that characterise HSEC. This inducibility of fenestrations in vascular endothelial cells together with the impracticality of using electron microscopy for routine phenotyping means that the presence of fenestrations alone cannot be used to define HSEC in most experimental situations.
NORMAL ‘ENDOTHELIAL’ CELL PHENO-TYPE AND FUNCTIONS
HSEC form a single cell barrier between the hepatocytes and the bloodstream and are strategically situated to interact with leukocytes and other blood constituents. The cells produce a minimal basement membrane[34] , which is mostly composed of type IV collagen in normal liver[35], and have a high endocytotic capacity[3]. They express some markers that are common to all endothelial cells and these provide a useful means to positively identify a cell of ‘endothelial’ lineage (Table 1).
Table 1.
The expression of classical markers of endothelial phenotype by human sinusoidal endothelial cells
| Marker | Extra-hepatic endothelial expression | Sinusoidal endothelial cells in vivo | Sinusoidal endothelial cells in vitro | Problems with use in phenotyping? |
| CD31 | Vascular and lymphatic endothelial cells | Yes, but at low levels | Yes | Widely expressed on all EC thus not specific for HSEC |
| vWF | Vascular endothelial cells | Controversial, Yes | Yes | Widely expressed on all EC thus not specific for HSEC |
| Ulex lectin binding | Endothelial cells | Yes | Yes | Widely expressed on all EC thus not specific for HSEC |
| Uptake of AcLDL | Endothelial cells | Yes | Yes | Also taken up by macrophages in liver and other EC |
| CD34 | Vascular and lymphatic endothelial cells | No | Absent or low | May be upregulated during capillarisation or with passage in vitro |
| E-Selectin | Vascular endothelial cells | Low or absent under normal conditions | Low, can be upregulated by cytokines | Widely expressed on activated vascular EC, not specific for HSEC |
| Pal-E antigen | Vascular endothelial cells | No | No? | May be upregulated during capillarisation |
| CD105/ endoglin | May be upregulated during capillarisation | Yes | Yes | Widely expressed on all EC, also by stellate cells and fibroblasts in liver |
While many of the above markers are indeed expressed on hepatic sinusoidal endothelial cells and provide a means of confirming “sinusoidal endothelial identity”, none is specific to sinusoidal endothelial cells.
CD31
CD31 or PECAM-1 is an abundantly expressed membr-ane glycoprotein member of the immunoglobulin superfamily (reviewed in[36]). It is constitutively expressed on endothelial cells and some haematopoetic cells and has functions in cell adhesion and signalling. Expression of CD31 is used widely as a marker of ‘continuous’ or classical vascular endothelium, but expression of CD31 by hepatic sinusoidal endothelial cells remains controversial. In rodent studies, CD31 has been localised to normal sinusoidal endothelial cells using a variety of techniques[4,37] and has been shown to be downregulated following CCl4 or TNFα mediated liver injury[37]. However, other studies suggest that the protein is absent under normal conditions[34] and indeed a lack of CD31 has been used to characterise rodent HSEC[7].
The situation is similarly complex in human HSEC. During human embryonic development CD31 is absent from HSEC until wk 25. In the adult, CD31 has been reported as present on HSEC by FACS and immunohistochemistry on sections from cirrhotic liver but is minimally present on normal liver[8] or alternately present on normal liver and enhanced on cirrhotic liver[38]. Our own data show that CD31 is expressed by both normal and diseased liver HSEC (Figures 1 and 2). In general, it seems that HSEC express lower levels of CD31 than vascular EC ([16] and our observations). However, one needs to interpret immunohistochemical analysis with caution because CD31 is present on kupffer cells in sinusoids. Studies on isolated HSEC using both PCR and antibody-based assessment of characteristic endothelial markers, such as CD31 and vWF (see below) in parallel, show varying results[39]. Some studies suggest that subcellular localisation of CD31 can be used as a marker of HSEC phenotype to indicate whether cells have dedifferentiated in culture[16]. Thus cells with cytoplasmic CD31 are ‘normal’ whereas dedifferentiated/capillarised EC demonstrate increased membranous expression.
Figure 1.
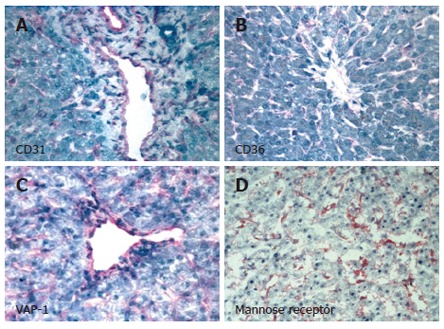
Human hepatic sinusoidal endothelial cells in situ express the classical endothelial phenotypic markers CD31 (A) and CD36 (B) as well as more recently identified markers such as VAP-1 (C) and Mannose receptor (D). Images represent immunohistochemical staining of human liver sections using specific primary antibodies in an indirect immunoperoxidase protocol with haematoxylin counterstain. Positive staining of sinusoidal endothelial cells is indicated by pink pigmentation. CD31, CD36 and VAP-1 are observed on both portal vessel endothelial cells and sinusoidal endothelium, whilst the mannose receptor is localised on sinusoidal endothelial cells and Kupffer cells.
Figure 2.
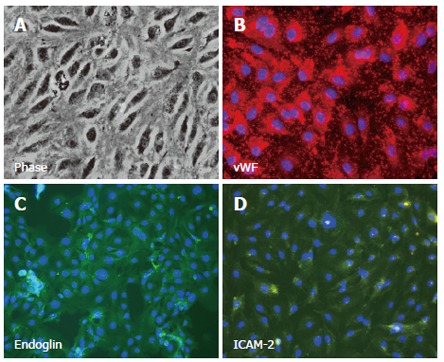
Isolated, cultured human hepatic sinusoidal endothelial cells exhibit classical morphology under phase contrast microscopy (A), and stain positively with antibody directed against endothelial phenotypic markers. Images represent immunofluorescent staining of cultured cells using specific primary antibodies in an indirect fluorescent protocol with DAPI nuclear counterstain (blue). Positive staining for vWF (B) is visualised using a Texas Red-labelled secondary antibody, whilst expression of endoglin (CD105, C) and ICAM-2 (D) are visualised with a FITC-conjugated secondary antibody (green).
Von Willebrand Factor (vWF)
vWF is a multimeric glycoprotein that binds and stabilises the coagulation factor FVIII as well as supports the adhesion of platelets to subendothelial structures during vascular damage. It is expressed by both platelets and endothelial cells and is often used as a marker to identify endothelium. In most vascular endothelial cells, von Willebrand Factor is stored in cytoplasmic vesicles called Weibel Palade bodies. Expression of vWF varies between different vascular beds in vivo[40] and particularly low levels are observed in the liver, most of which is detected in vascular rather than sinusoidal endothelial cells. The low levels of vWF detected in HSEC are consistent with the reported lack of Weibel Palade bodies. However, definitive evidence supporting the presence or absence of these structures is also lacking. Studies in mice suggest that HSEC contain Weibel-Palade bodies[41] and produce VWF at the mRNA and protein level[4,11,41]. Other groups working with rat HSEC report that vWF is not expressed in normal rat cells[42] and these findings are supported by porcine and rat studies showing absence of Weibel-Palade bodies in HSEC[9,43]. In human cells, vWF expression has been reported in both normal[8,44] and diseased samples[45] and we and others[8] have demonstrated expression on passaged, cultured HSEC in vitro (Figure 2).
E-Selectin
E-Selectin is a member of the selectin family of adhesion molecules that supports leukocyte binding. Expression of E-Selectin is restricted to cells of endothelial lineage[46] and is induced by inflammation in vivo and exposure of endothelial cells to proinflammatory cytokines and LPS in vitro. The ability to express E-selectin can thus be used to define endothelial cells in culture. However, expression of E-selectin is restricted to vascular endothelial cells in the normal liver[47] , although it may be upregulated on sinusoidal endothelium in disease and during metastatic processes[48] and animal studies demonstrate a minimal role for E-selectin in leukocyte recruitment to liver tissue[49]. However, we have reported expression of functional E-Selectin on cultured cytokine-stimulated human HSEC suggesting that HSEC can express E-Selectin under restricted circumstances in vivo and in vitro[18]. Recent studies demonstrating that E-Selectin expression by HUVEC in response to TNFα is reduced by pretreatment with HGF suggest that paracrine factors from adjacent hepatocytes may suppress E-selectin in the sinusoids in vivo[50].
Binding of lectins and Acetylated LDL uptake
The ability to bind Ulex lectin and take up acetylated LDL is often used to define HSEC. Ulex lectin, from the gorse family of plants, binds alpha-L fucose containing receptors and is commonly used as a histological marker for endothelial cells, although in some tissues it also binds epithelial structures[51]. In the liver, different lectins bind differentially within the vasculature. Concanavalin A binds with equal affinity for all segments of the microvasculature whereas wheat germ agglutinins show preferential binding to the sinusoidal vasculature as a consequence of differences in distribution of glycosylated ligands throughout the acinus. In most studies Ulex lectins do not bind preferentially to sinusoidal endothelium as a consequence of the relative paucity of alpha-L fucose motifs. The ability of wheat germ lectins to bind sinusoidal endothelium has led to their use in selectively purifying sinusoidal EC and their preference for periportal HSEC has even led to the suggestion that they can be used to differentially purify periportal verses perivenous HSEC[5]. Staining with Ulex lectin is increased in disease and is particularly pronounced during capillarisation of the sinusoids[52]. Thus although binding of Ulex lectin is indicative of an endothelial phenotype, it is not restricted to ‘sinusoidal’ endothelial cells[52] and is not a good marker of HSEC.
The liver is the major site for the scavenger receptor-mediated clearance of lipoproteins from the circulation. Acetylated LDL is mainly cleared by hepatic endothelial cells[53] by binding to scavenger receptors including scavenger receptor class A (SR-AI/II). However, these receptors are expressed by both HSEC and macrophages[54] and the ability to take up acetylated LDL is common to many extrahepatic endothelial cell populations, again reducing the specificity of acLDL uptake as a characteristic property of HSEC.
CD34
CD34 is a type 1 transmembrane sialomucin expressed by haematopoetic stem cells, capillary and lymphatic endothelial cells. CD34 is absent from most sinusoidal endothelial cells in normal liver but expression increases[8,27,55] during capillarisation in chronic inflamma-tory disease and in the sinusoidal-type vasculature within hepatocellular carcinomas[56,57]. CD34 expression has also been shown to increase in other tissues including the rheumatoid joint and at sites of neolymphoid development during chronic inflammation. We have described CD34 positive lymphatic-like vessels in portal associated lymphatic tissue in chronic inflammatory liver diseases including PSC[58] and hepatitis C (Heydtmann 2006 J Immunol in press). However, CD34 is not expressed on most HSEC in non-inflamed tissue in vivo and is absent from low passage human HSEC in vitro.
Pal-E Antigen
The antigen recognised by the Pal-E antibody is a widely used marker of vascular endothelial cells[59-61] The identity of the protein has been proposed as either a secreted form of vimentin produced by endothelial cells[61] or a protein designated PV-1/FELS (plasmalemmal vesicle-1/fenestrated endothelial-linked structural protein)[60]. PV-1 is particularly interesting in the context of HSEC. As its name implies, PV-1/FELS is expressed by fenestrated endothelial cells in the kidney and pancreas[62] but it is absent from fenestrated hepatic sinusoidal endothelial cells[60,62] and is restricted to vascular endothelium and neovessels in areas of capilliarisation in chronic liver disease and hepatocellular carcinoma[52,58].
Vascular Endothelial-Cadherin (VE-Cadherin)
Cadherins are a family of adhesion molecules that demonstrate cation- dependent homophilic and heterophilic binding. Endothelial cells express at least three cadherins: N-, P-, and VE-cadherin. VE-cadherin is localised to the inter-endothelial cell junction where it is an essential part of the adherens junctions that maintains endothelial permeability, monolayer integrity, morphogenesis and angiogenic reponses. Most studies suggest that sinusoidal endothelium in normal liver lacks VE-cadherin or expresses it at low levels, although it can be detected in chronic inflammation[38,63]. It seems likely that the relative lack of VE-cadherin on HSEC is a consequence of the absence of classical adherens junctions between HSEC and this is consistent with a lack of other junctional proteins, including vascular endothelial junctional adhesion molecule (VE-JAM/JAM-2), which is a member of the immunoglobulin superfamily structurally similar to JAM-1, which is absent from foetal and adult liver[64].
CD105/endoglin
CD105 (endoglin) is a hypoxia-inducible protein that is widely expressed on endothelial cells and is upregulated during angiogenesis. It is a receptor for transforming growth factor (TGF) -β1 and -β3 and modulates TGF-β signalling by interacting with TGF-β receptorsIand/or II (for review see[65]). CD105 has been used as a marker of angiogenesis, particularly in tumour tissue[66], and because it is a transmembrane molecule, it has been used in antibody-mediated positive selection strategies for endothelial cell isolation. However, expression of the molecule is not restricted to endothelial cells[65], and in the liver, expression has been reported in both stellate cells and myofibroblasts[67]. We have demonstrated that CD105 is expressed on hepatic sinusoidal endothelial cells (Figure 2) but again emphasise that this is not a tissue- or cell lineage-specific marker and urge caution when using it as a phenotypic identifier.
NEWER MARKERS OF ENDOTHELIAL PHENOTYPE RELATE TO THE FUNCTIONS OF HSEC WITHIN THE LIVER MICRO-ENVIRONMENT
As well as exhibiting features characteristic of all endo-thelial cells, hepatic sinusoidal endothelial cells fulfil many specific features within the liver environment. These include providing a barrier to minimise access of blood-borne material into the parenchyma and specific protein/antigen uptake and presentation. The embryonic origins of these cells and their similarity with lymphatic endothelial cells (see later) mean that they express several markers that are not present on vascular endothelium and these can be used to distinguish them from other endothelial cells. Thus, it is possible to use protein markers related to these specific origins and functions to confirm the phenotype of HSEC in vitro.
Scavenger functions/lipid uptake functions of EC
The exposure of sinusoidal endothelial cells to blood originating from both the systemic circulation and the gut means that HSEC are strategically situated to remove and recycle blood-borne proteins and lipids. In combination with Kupffer cells, HSEC constitute the most powerful scavenger system in the body[68]. The uptake of solutes is facilitated by the presence of fenestrae, the lack of a classical basement membrane and the expression of multiple scavenger receptors that allow them to bind and take up specific classes of molecules. These properties facilitate bidirectional transfer of materials to the parenchyma. Many of the scavenger receptor proteins can be used to determine the phenotype of HSEC.
The link family of proteins has recently been described as scavenger receptors responsible for clearance of a variety of proteins, including advanced glycation end products, modified LDL and bacteria[69]. Two members of this protein family, Stabilin-1 and -2, are constitutively expressed by hepatic sinusoidal endothelial cells[69,70]. Stabilin-2 is the major lymph node and liver hyaluronan and glycosaminoglycan scavenger receptor whilst Stabilin-1 (also called Feel-1 or CLEVER-1[71-73] is a more promiscuous scavenger receptor. In common with many other scavenger-type receptors, these proteins are present on sinusoidal endothelial cells in spleen and lymph node as well as the liver. Most of the scavenger functions assigned to this molecule relate to endocytosis of hyaluronic acid, acetylated LDL and glycation end products, but there is also evidence to support roles for Stabilin-1 in leukocyte adhesion and tumour metastasis[69,74]. Another member of the link protein family is LYVE-1, an endothelial hyaluronan receptor predominantly restricted to lymphatic endothelial cells (reviewed in[75]). Putative functions for LYVE-1 include the uptake of hyaluronic acid and regulation of leukocyte adhesion or migration within the lymphatic circulation. Interestingly, hepatic sinusoidal endothelial cells also express LYVE-1 constitutively[76] (Figure 3) with evidence of a zonal distribution, the highest levels being detected in acinar zone 2. This hyaluronan receptor is present on both normal and diseased human HSEC, although lower levels are observed in cirrhosis and the protein is absent in HCC[76]. Expression is also seen on portal-associated lymphatics in chronic liver disease[58,76].
Figure 3.
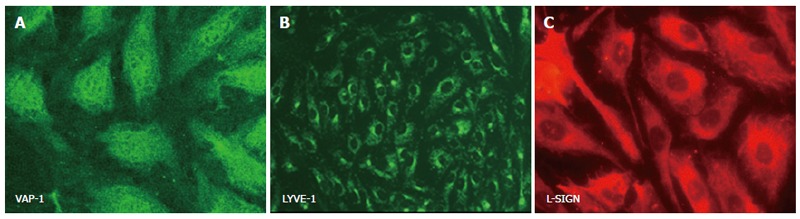
Cultured human hepatic sinusoidal endothelial cells stain positively with antibody directed against ‘non-classical endothelial’ phenotypic markers. Images represent immunofluorescent staining of cultured cells using specific primary antibodies in an indirect fluorescent protocol. Expression of VAP-1 (A) and LYVE-1 (B) are visualised with a FITC-conjugated secondary antibody (green), whilst positive staining for L-SIGN (C) is visualised using a Texas Red-labelled secondary antibody.
The liver is the major site for synthesis and metabolism of cholesterol and scavenger receptors of class A (SR-A) on both KC and HSEC[77] are responsible for the uptake of oxidised/acetylated LDL, which is subsequently passed on to hepatocytes. Another HDL/LDL receptor, CD36[78], also known as GPIV, is expressed at high levels on platelets, monocyte/macrophages and vascular endothelial cells. In the liver, CD36 is strongly expressed on sinusoidal endothelial cells[79] where it fulfills multiple functions including acting as a scavenger receptor for oxidised lipid[80] and as an adhesion receptor for red blood cells infected with malarial parasite[81]. There are two alternatively spliced members of the scavenger receptor B family (SR-BI and -BII). Scavenger receptor-B1 is expressed by HSEC and is responsible for the uptake of HDL cholesterol esters to liver parenchymal cells and also acts as a coreceptor for HCV infection[82].
The calcium-dependent C-type lectins, Dendritic Cell-specific ICAM-3 grabbing nonintegrin (DC-SIGN) or CD209 and the related molecule Liver/lymph node-specific intercellular adhesion molecule-3-grabbing nonintegrin (L-SIGN) or CD209L (Figure 3), are constitutively expressed on hepatic endothelial cells[83]. DC-SIGN is expressed at high levels on myeloid dendritic cells in tissues where it interacts with ICAM-3 on T cells as part of the immunological synapse as well as being an attachment factor for HCV, ebola virus, CMV, HIV and other lentiviruses (reviewed in[84,85]). We have recently reported that DC-SIGN is present on hepatic sinusoidal endothelial cells and that expression is increased in response to treatment with cytokines including IL-4[86]. DC-SIGN on endothelial cells acts as an attachment factor for HCV but does not mediate HCV entry directly but rather enhances infection of hepatocytes in trans. DC-SIGN is absent from most other vascular beds, although it has been reported on a brain microvascular cell line[87].
Liver/lymph node-specific ICAM-3-grabbing integrin (L-SIGN), otherwise known as DC-SIGN-related (DC SIGNR) (CD209L), shares 77% amino acid homology with DC-SIGN. Like DC-SIGN it can also bind ICAM-3, HIV and HCV. L-SIGN is strongly and constitutively expressed on sinusoidal endothelial cells in the liver and on endothelium in lymph nodes but not on DCs or on endothelium in other tissues. L-SIGN is thus an excellent marker of liver endothelium[44,88]. A related molecule, Liver and Lymph node Sinusoidal Endothelial C-type lectin (LSECtin) is also expressed on sinusoidal endothelial cells and has recently been demonstrated to mediate attachment of filovirus and coronavirus particles[89]. Thus sinusoidal endothelial cells[90] can bind a wide variety of pathogens after which they pass hepatotropic viruses on to adherent hepatocytes in trans[91] thereby concentrating viral pathogens within the liver.
Antigen presentation by HSEC
As well as being equipped with scavenger receptors that facilitate efficient uptake of viruses and potential antigens, HSEC also have the ability to phagocytose particles and to present antigen to lymphocytes(reviewed in [92-94]). There is evidence that such interactions are important for generating immunological tolerance to gut-derived antigens although recent work suggests that local antigen presentation cannot explain liver tolerance and that, on the contrary, the liver may be an excellent priming site for naive CD8+ T cells[95]. Antigen presentation is facilitated by the expression of MHC classIand II[96,97] molecules together with co-stimulatory molecules such as CD40 and more contentiously CD80 and CD86[98]. These receptors are upregulated on HSEC in fulminant liver failure[99] and may contribute to disease pathogenesis by allowing ongoing presentation of stimulatory antigen. HSEC also express the mannose receptor (Figure 1), a 175 kDa transmembrane glycosylated protein involved in uptake of Ag by both DCs and HSEC[97]. Competitive inhibition of this receptor by mannan reduces antigen-specific T cell activation by murine HSEC[97].
Similarities between lymphatic endothelial cells and sinusoidal endothelium
Both the liver and pancreas develop from buds of the embryonic endoderm[100], however, the vasculature components of the liver have distinct origins. The portal vessels are derived from viteine veins whereas the sinusoids develop from the capillary vessels of the septum transversum and acquire their distinctive fenestrated phenotype by wk 20 of gestation (reviewed in[21]). From this point onward, sinusoidal endothelial cells remain functionally and phenotypically distinct from the other vascular endothelial cells in the liver and express several receptors that are otherwise confined to lymphatic endothelial cells that are derived from buds from the cardinal vein. Hence, both lymphatic and sinusoidal endothelial cells have minimal basement membranes, loosely organised cell junctions and constitutively express LYVE-1 and VAP-1 (SSAO/AOC3) but lack CD34 (reviewed in[101]). VAP-1 is a type II transmembrane protein that can support leukocyte adhesion via interactions with sialic acid rich side chains. It is also an amine oxidase and enzyme activity is also involved in regulating leukocyte adhesion and transmigration[101]. VAP-1 is expressed on all vascular compartments within the liver (Figures 1 and 2) where it supports the adhesion and transmigration of leukocytes[10,102]. The only extrahepatic site where VAP-1 is constitutively expressed at high levels is endothelial cells in high endothelial venules within lymph nodes[103] where again it is proposed to have a role directing the adhesion of lymphocyte populations[104]. Similarly both lymphatic endothelium and HSEC express the Reeler gene product Reelin[105]. Reelin is a secreted glycoprotein with roles in embryonic development and organisation. Expression is restricted during embryogenesis but in the adult organism high levels are detected on lymphatic endothelial cells and within the sinusoids localised either to stellate cells[106] or HSEC[105]. This has led to the hypothesis that reelin may be involved in the regulation of lymphoangiogenesis or regulation of lymphatic endothelial phenotype and thus may have similar roles within the liver sinusoids.
Thus there are many similarities between HSEC and lymphatic endothelial cells and some antigens originally defined on lymphatic endothelium can also be used to differentiate between HSEC and vascular endothelial cells in the liver. It is possible to exclude contamination with lymphatic EC in HSEC cultures on the basis that LYVE-1 positive HSEC do not express PROX -1, a transcription factor found exclusively in lymphatic EC[107].
CONCLUSIONS
Cultures of endothelial cells are valuable tools to investigate mechanisms of liver physiology and pathophysiology in vitro. However, the study of endothelial cells in vitro is complicated by the marked heterogeneity of endothelial cells between and within different organs and the tendency for cells to lose tissue-specific markers when cultured in vitro. Although all endothelial cells share some characteristic features (as described in the first part of this review), there is a need for specific markers or combinations of markers that define distinct populations of endothelial cells. To date, the study of HSEC in vitro has been hampered by the lack of specific markers that can conclusively identify these cells and discriminate them from vascular or lymphatic endothelial cells. To some extent this remains the case, since many classical endothelial markers are widely expressed (Table 1) and there is considerable reported variability in detection of phenotypic markers between animal and human systems (eg CD31 and vWF). To date there is no known single molecule that is only expressed on hepatic sinusoidal and no other type of endothelia. However, the increasing knowledge of endothelial receptors is providing us with a larger and better defined set of phenotypic makers. In addition to their use in phenotyping or sorting/selecting specific endothelial cells for culture, receptors that show tissue-specific expression provide clues to specific functions of the cells being studied. Examples of this are the large number of scavenger receptors expressed by HSEC. Cultured HSEC do exhibit some useful identifying features, however. Very low passage cells retain fenestrations in vitro for a short time but these rapidly disappear within a passage or two in culture[8,16], as does expression of VAP-1[10]. Apart from these changes, however, the cells remain relatively phenotypically stable for 7-8 passages[8] and can be identified by expression of CD31, LYVE-1, DC-SIGNR(L-SIGN), Stabilin-1 and lack of CD34 and PROX-1[107]. These markers confirm endothelial identity, while excluding vascular and lymphatic endothelial contamination and in conjunction with markers to exclude cells of leukocyte origin can be used to confirm the sinusoidal nature of the cells.
Footnotes
S- Editor Wang J L- Editor Lutze M E- Editor Ma WH
References
- 1.Matsumoto T, Kawakami M. The unit-concept of hepatic parenchyma--a re-examination based on angioarchitectural studies. Acta Pathol Jpn. 1982;32 Suppl 2:285–314. [PubMed] [Google Scholar]
- 2.Takasaki S, Hano H. Three-dimensional observations of the human hepatic artery (Arterial system in the liver) J Hepatol. 2001;34:455–466. doi: 10.1016/s0168-8278(00)00058-1. [DOI] [PubMed] [Google Scholar]
- 3.Braet F, Wisse E. Structural and functional aspects of liver sinusoidal endothelial cell fenestrae: a review. Comp Hepatol. 2002;1:1. doi: 10.1186/1476-5926-1-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Do H, Healey JF, Waller EK, Lollar P. Expression of factor VIII by murine liver sinusoidal endothelial cells. J Biol Chem. 1999;274:19587–19592. doi: 10.1074/jbc.274.28.19587. [DOI] [PubMed] [Google Scholar]
- 5.Vidal-Vanaclocha F, Rocha M, Asumendi A, Barberá-Guillem E. Isolation and enrichment of two sublobular compartment-specific endothelial cell subpopulations from liver sinusoids. Hepatology. 1993;18:328–339. [PubMed] [Google Scholar]
- 6.Yamane A, Seetharam L, Yamaguchi S, Gotoh N, Takahashi T, Neufeld G, Shibuya M. A new communication system between hepatocytes and sinusoidal endothelial cells in liver through vascular endothelial growth factor and Flt tyrosine kinase receptor family (Flt-1 and KDR/Flk-1) Oncogene. 1994;9:2683–2690. [PubMed] [Google Scholar]
- 7.Knolle PA, Germann T, Treichel U, Uhrig A, Schmitt E, Hegenbarth S, Lohse AW, Gerken G. Endotoxin down-regulates T cell activation by antigen-presenting liver sinusoidal endothelial cells. J Immunol. 1999;162:1401–1407. [PubMed] [Google Scholar]
- 8.Daneker GW, Lund SA, Caughman SW, Swerlick RA, Fischer AH, Staley CA, Ades EW. Culture and characterization of sinusoidal endothelial cells isolated from human liver. In Vitro Cell Dev Biol Anim. 1998;34:370–377. doi: 10.1007/s11626-998-0018-9. [DOI] [PubMed] [Google Scholar]
- 9.Gerlach JC, Zeilinger K, Spatkowski G, Hentschel F, Schnoy N, Kolbeck S, Schindler RK, Neuhaus P. Large-scale isolation of sinusoidal endothelial cells from pig and human liver. J Surg Res. 2001;100:39–45. doi: 10.1006/jsre.2001.6224. [DOI] [PubMed] [Google Scholar]
- 10.Lalor PF, Edwards S, McNab G, Salmi M, Jalkanen S, Adams DH. Vascular adhesion protein-1 mediates adhesion and transmigration of lymphocytes on human hepatic endothelial cells. J Immunol. 2002;169:983–992. doi: 10.4049/jimmunol.169.2.983. [DOI] [PubMed] [Google Scholar]
- 11.Katz SC, Pillarisetty VG, Bleier JI, Shah AB, DeMatteo RP. Liver sinusoidal endothelial cells are insufficient to activate T cells. J Immunol. 2004;173:230–235. doi: 10.4049/jimmunol.173.1.230. [DOI] [PubMed] [Google Scholar]
- 12.Gomez DE, Hartzler JL, Corbitt RH, Nason AM, Thorgeirsson UP. Immunomagnetic separation as a final purification step of liver endothelial cells. In Vitro Cell Dev Biol Anim. 1993;29A:451–455. [PubMed] [Google Scholar]
- 13.Sawyer RT, Garner RE, Hudson JA. Effect of lectins on hepatic clearance and killing of Candida albicans by the isolated perfused mouse liver. Infect Immun. 1992;60:1041–1046. doi: 10.1128/iai.60.3.1041-1046.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Scoazec JY, Feldmann G. Both macrophages and endothelial cells of the human hepatic sinusoid express the CD4 molecule, a receptor for the human immunodeficiency virus. Hepatology. 1990;12:505–510. doi: 10.1002/hep.1840120310. [DOI] [PubMed] [Google Scholar]
- 15.Scoazec JY, Racine L, Couvelard A, Flejou JF, Feldmann G. Endothelial cell heterogeneity in the normal human liver acinus: in situ immunohistochemical demonstration. Liver. 1994;14:113–123. doi: 10.1111/j.1600-0676.1994.tb00059.x. [DOI] [PubMed] [Google Scholar]
- 16.DeLeve LD, Wang X, Hu L, McCuskey MK, McCuskey RS. Rat liver sinusoidal endothelial cell phenotype is maintained by paracrine and autocrine regulation. Am J Physiol Gastrointest Liver Physiol. 2004;287:G757–G763. doi: 10.1152/ajpgi.00017.2004. [DOI] [PubMed] [Google Scholar]
- 17.Módis L, Martinez-Hernandez A. Hepatocytes modulate the hepatic microvascular phenotype. Lab Invest. 1991;65:661–670. [PubMed] [Google Scholar]
- 18.Edwards S, Lalor PF, Nash GB, Rainger GE, Adams DH. Lymphocyte traffic through sinusoidal endothelial cells is regulated by hepatocytes. Hepatology. 2005;41:451–459. doi: 10.1002/hep.20585. [DOI] [PubMed] [Google Scholar]
- 19.Ghitescu L, Robert M. Diversity in unity: the biochemical composition of the endothelial cell surface varies between the vascular beds. Microsc Res Tech. 2002;57:381–389. doi: 10.1002/jemt.10091. [DOI] [PubMed] [Google Scholar]
- 20.Jackson DG. The lymphatics revisited: new perspectives from the hyaluronan receptor LYVE-1. Trends Cardiovasc Med. 2003;13:1–7. doi: 10.1016/s1050-1738(02)00189-5. [DOI] [PubMed] [Google Scholar]
- 21.Gouysse G, Couvelard A, Frachon S, Bouvier R, Nejjari M, Dauge MC, Feldmann G, Hénin D, Scoazec JY. Relationship between vascular development and vascular differentiation during liver organogenesis in humans. J Hepatol. 2002;37:730–740. doi: 10.1016/s0168-8278(02)00282-9. [DOI] [PubMed] [Google Scholar]
- 22.Steffan AM, Pereira CA, Bingen A, Valle M, Martin JP, Koehren F, Royer C, Gendrault JL, Kirn A. Mouse hepatitis virus type 3 infection provokes a decrease in the number of sinusoidal endothelial cell fenestrae both in vivo and in vitro. Hepatology. 1995;22:395–401. doi: 10.1016/0270-9139(95)90556-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Vidal-Vanaclocha F, Barberá-Guillem E. Fenestration patterns in endothelial cells of rat liver sinusoids. J Ultrastruct Res. 1985;90:115–123. doi: 10.1016/0889-1605(85)90102-8. [DOI] [PubMed] [Google Scholar]
- 24.Horn T, Christoffersen P, Henriksen JH. Alcoholic liver injury: defenestration in noncirrhotic livers--a scanning electron microscopic study. Hepatology. 1987;7:77–82. doi: 10.1002/hep.1840070117. [DOI] [PubMed] [Google Scholar]
- 25.Mak KM, Lieber CS. Alterations in endothelial fenestrations in liver sinusoids of baboons fed alcohol: a scanning electron microscopic study. Hepatology. 1984;4:386–391. doi: 10.1002/hep.1840040306. [DOI] [PubMed] [Google Scholar]
- 26.Le Couteur DG, Fraser R, Hilmer S, Rivory LP, McLean AJ. The hepatic sinusoid in aging and cirrhosis: effects on hepatic substrate disposition and drug clearance. Clin Pharmacokinet. 2005;44:187–200. doi: 10.2165/00003088-200544020-00004. [DOI] [PubMed] [Google Scholar]
- 27.Xu B, Broome U, Uzunel M, Nava S, Ge X, Kumagai-Braesch M, Hultenby K, Christensson B, Ericzon BG, Holgersson J, et al. Capillarization of hepatic sinusoid by liver endothelial cell-reactive autoantibodies in patients with cirrhosis and chronic hepatitis. Am J Pathol. 2003;163:1275–1289. doi: 10.1016/S0002-9440(10)63487-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chen J, Braet F, Brodsky S, Weinstein T, Romanov V, Noiri E, Goligorsky MS. VEGF-induced mobilization of caveolae and increase in permeability of endothelial cells. Am J Physiol Cell Physiol. 2002;282:C1053–C1063. doi: 10.1152/ajpcell.00292.2001. [DOI] [PubMed] [Google Scholar]
- 29.Funyu J, Mochida S, Inao M, Matsui A, Fujiwara K. VEGF can act as vascular permeability factor in the hepatic sinusoids through upregulation of porosity of endothelial cells. Biochem Biophys Res Commun. 2001;280:481–485. doi: 10.1006/bbrc.2000.4148. [DOI] [PubMed] [Google Scholar]
- 30.Krause P, Markus PM, Schwartz P, Unthan-Fechner K, Pestel S, Fandrey J, Probst I. Hepatocyte-supported serum-free culture of rat liver sinusoidal endothelial cells. J Hepatol. 2000;32:718–726. doi: 10.1016/s0168-8278(00)80239-1. [DOI] [PubMed] [Google Scholar]
- 31.Yokomori H, Oda M, Yoshimura K, Nagai T, Ogi M, Nomura M, Ishii H. Vascular endothelial growth factor increases fenestral permeability in hepatic sinusoidal endothelial cells. Liver Int. 2003;23:467–475. doi: 10.1111/j.1478-3231.2003.00880.x. [DOI] [PubMed] [Google Scholar]
- 32.Watanabe Y, Dvorak HF. Vascular permeability factor/vascular endothelial growth factor inhibits anchorage-disruption-induced apoptosis in microvessel endothelial cells by inducing scaffold formation. Exp Cell Res. 1997;233:340–349. doi: 10.1006/excr.1997.3583. [DOI] [PubMed] [Google Scholar]
- 33.Esser S, Wolburg K, Wolburg H, Breier G, Kurzchalia T, Risau W. Vascular endothelial growth factor induces endothelial fenestrations in vitro. J Cell Biol. 1998;140:947–959. doi: 10.1083/jcb.140.4.947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Scoazec JY, Feldmann G. The cell adhesion molecules of hepatic sinusoidal endothelial cells. J Hepatol. 1994;20:296–300. doi: 10.1016/s0168-8278(05)80072-8. [DOI] [PubMed] [Google Scholar]
- 35.Hahn E, Wick G, Pencev D, Timpl R. Distribution of basement membrane proteins in normal and fibrotic human liver: collagen type IV, laminin, and fibronectin. Gut. 1980;21:63–71. doi: 10.1136/gut.21.1.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Newton JP, Hunter AP, Simmons DL, Buckley CD, Harvey DJ. CD31 (PECAM-1) exists as a dimer and is heavily N-glycosylated. Biochem Biophys Res Commun. 1999;261:283–291. doi: 10.1006/bbrc.1999.1018. [DOI] [PubMed] [Google Scholar]
- 37.Neubauer K, Ritzel A, Saile B, Ramadori G. Decrease of platelet-endothelial cell adhesion molecule 1-gene-expression in inflammatory cells and in endothelial cells in the rat liver following CCl(4)-administration and in vitro after treatment with TNFalpha. Immunol Lett. 2000;74:153–164. doi: 10.1016/s0165-2478(00)00203-0. [DOI] [PubMed] [Google Scholar]
- 38.Medina J, Sanz-Cameno P, García-Buey L, Martín-Vílchez S, López-Cabrera M, Moreno-Otero R. Evidence of angiogenesis in primary biliary cirrhosis: an immunohistochemical descriptive study. J Hepatol. 2005;42:124–131. doi: 10.1016/j.jhep.2004.09.024. [DOI] [PubMed] [Google Scholar]
- 39.Unger RE, Krump-Konvalinkova V, Peters K, Kirkpatrick CJ. In vitro expression of the endothelial phenotype: comparative study of primary isolated cells and cell lines, including the novel cell line HPMEC-ST1.6R. Microvasc Res. 2002;64:384–397. doi: 10.1006/mvre.2002.2434. [DOI] [PubMed] [Google Scholar]
- 40.Yamamoto K, de Waard V, Fearns C, Loskutoff DJ. Tissue distribution and regulation of murine von Willebrand factor gene expression in vivo. Blood. 1998;92:2791–2801. [PubMed] [Google Scholar]
- 41.Harrison RL, Boudreau R. Human hepatic sinusoidal endothelial cells in culture produce von Willebrand factor and contain Weibel-Palade bodies. Liver. 1989;9:242–249. doi: 10.1111/j.1600-0676.1989.tb00406.x. [DOI] [PubMed] [Google Scholar]
- 42.Lenzi R, Alpini G, Liu MH, Rand JH, Tavoloni N. von Willebrand factor antigen is not an accurate marker of rat and guinea pig liver endothelial cells. Liver. 1990;10:372–379. doi: 10.1111/j.1600-0676.1990.tb00483.x. [DOI] [PubMed] [Google Scholar]
- 43.Wisse E. An ultrastructural characterization of the endothelial cell in the rat liver sinusoid under normal and various experimental conditions, as a contribution to the distinction between endothelial and Kupffer cells. J Ultrastruct Res. 1972;38:528–562. doi: 10.1016/0022-5320(72)90089-5. [DOI] [PubMed] [Google Scholar]
- 44.Pöhlmann S, Soilleux EJ, Baribaud F, Leslie GJ, Morris LS, Trowsdale J, Lee B, Coleman N, Doms RW. DC-SIGNR, a DC-SIGN homologue expressed in endothelial cells, binds to human and simian immunodeficiency viruses and activates infection in trans. Proc Natl Acad Sci USA. 2001;98:2670–2675. doi: 10.1073/pnas.051631398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Fukuda Y, Nagura H, Imoto M, Koyama Y. Immunohistochemical studies on structural changes of the hepatic lobules in chronic liver diseases. Am J Gastroenterol. 1986;81:1149–1155. [PubMed] [Google Scholar]
- 46.Tedder TF, Steeber DA, Chen A, Engel P. The selectins: vascular adhesion molecules. FASEB J. 1995;9:866–873. [PubMed] [Google Scholar]
- 47.Lalor PF, Shields P, Grant A, Adams DH. Recruitment of lymphocytes to the human liver. Immunol Cell Biol. 2002;80:52–64. doi: 10.1046/j.1440-1711.2002.01062.x. [DOI] [PubMed] [Google Scholar]
- 48.Khatib AM, Kontogiannea M, Fallavollita L, Jamison B, Meterissian S, Brodt P. Rapid induction of cytokine and E-selectin expression in the liver in response to metastatic tumor cells. Cancer Res. 1999;59:1356–1361. [PubMed] [Google Scholar]
- 49.Wong J, Johnston B, Lee SS, Bullard DC, Smith CW, Beaudet AL, Kubes P. A minimal role for selectins in the recruitment of leukocytes into the inflamed liver microvasculature. J Clin Invest. 1997;99:2782–2790. doi: 10.1172/JCI119468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Makondo K, Kimura K, Kitamura T, Yamaji D, Dong Jung B, Shibata H, Saito M. Hepatocyte growth factor/scatter factor suppresses TNF-alpha-induced E-selectin expression in human umbilical vein endothelial cells. Biochim Biophys Acta. 2004;1644:9–15. doi: 10.1016/j.bbamcr.2003.10.006. [DOI] [PubMed] [Google Scholar]
- 51.Holthöfer H, Virtanen I, Kariniemi AL, Hormia M, Linder E, Miettinen A. Ulex europaeus I lectin as a marker for vascular endothelium in human tissues. Lab Invest. 1982;47:60–66. [PubMed] [Google Scholar]
- 52.Hattori M, Fukuda Y, Imoto M, Koyama Y, Nakano I, Urano F. Histochemical properties of vascular and sinusoidal endothelial cells in liver diseases. Gastroenterol Jpn. 1991;26:336–343. doi: 10.1007/BF02781922. [DOI] [PubMed] [Google Scholar]
- 53.De Rijke YB, Biessen EA, Vogelezang CJ, van Berkel TJ. Binding characteristics of scavenger receptors on liver endothelial and Kupffer cells for modified low-density lipoproteins. Biochem J. 1994;304(Pt 1):69–73. doi: 10.1042/bj3040069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ling W, Lougheed M, Suzuki H, Buchan A, Kodama T, Steinbrecher UP. Oxidized or acetylated low density lipoproteins are rapidly cleared by the liver in mice with disruption of the scavenger receptor class A type I/II gene. J Clin Invest. 1997;100:244–252. doi: 10.1172/JCI119528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Couvelard A, Scoazec JY, Feldmann G. Expression of cell-cell and cell-matrix adhesion proteins by sinusoidal endothelial cells in the normal and cirrhotic human liver. Am J Pathol. 1993;143:738–752. [PMC free article] [PubMed] [Google Scholar]
- 56.Cui S, Hano H, Sakata A, Harada T, Liu T, Takai S, Ushigome S. Enhanced CD34 expression of sinusoid-like vascular endothelial cells in hepatocellular carcinoma. Pathol Int. 1996;46:751–756. doi: 10.1111/j.1440-1827.1996.tb03544.x. [DOI] [PubMed] [Google Scholar]
- 57.Frachon S, Gouysse G, Dumortier J, Couvelard A, Nejjari M, Mion F, Berger F, Paliard P, Boillot O, Scoazec JY. Endothelial cell marker expression in dysplastic lesions of the liver: an immunohistochemical study. J Hepatol. 2001;34:850–857. doi: 10.1016/s0168-8278(01)00049-6. [DOI] [PubMed] [Google Scholar]
- 58.Grant AJ, Goddard S, Ahmed-Choudhury J, Reynolds G, Jackson DG, Briskin M, Wu L, Hübscher SG, Adams DH. Hepatic expression of secondary lymphoid chemokine (CCL21) promotes the development of portal-associated lymphoid tissue in chronic inflammatory liver disease. Am J Pathol. 2002;160:1445–1455. doi: 10.1016/S0002-9440(10)62570-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Schlingemann RO, Dingjan GM, Emeis JJ, Blok J, Warnaar SO, Ruiter DJ. Monoclonal antibody PAL-E specific for endothelium. Lab Invest. 1985;52:71–76. [PubMed] [Google Scholar]
- 60.Niemelä H, Elima K, Henttinen T, Irjala H, Salmi M, Jalkanen S. Molecular identification of PAL-E, a widely used endothelial-cell marker. Blood. 2005;106:3405–3409. doi: 10.1182/blood-2005-01-0254. [DOI] [PubMed] [Google Scholar]
- 61.Xu B, deWaal RM, Mor-Vaknin N, Hibbard C, Markovitz DM, Kahn ML. The endothelial cell-specific antibody PAL-E identifies a secreted form of vimentin in the blood vasculature. Mol Cell Biol. 2004;24:9198–9206. doi: 10.1128/MCB.24.20.9198-9206.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Stan RV, Kubitza M, Palade GE. PV-1 is a component of the fenestral and stomatal diaphragms in fenestrated endothelia. Proc Natl Acad Sci USA. 1999;96:13203–13207. doi: 10.1073/pnas.96.23.13203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Medina J, Caveda L, Sanz-Cameno P, Arroyo AG, Martín-Vílchez S, Majano PL, García-Buey L, Sánchez-Madrid F, Moreno-Otero R. Hepatocyte growth factor activates endothelial proangiogenic mechanisms relevant in chronic hepatitis C-associated neoangiogenesis. J Hepatol. 2003;38:660–667. doi: 10.1016/s0168-8278(03)00053-9. [DOI] [PubMed] [Google Scholar]
- 64.Palmeri D, van Zante A, Huang CC, Hemmerich S, Rosen SD. Vascular endothelial junction-associated molecule, a novel member of the immunoglobulin superfamily, is localized to intercellular boundaries of endothelial cells. J Biol Chem. 2000;275:19139–19145. doi: 10.1074/jbc.M003189200. [DOI] [PubMed] [Google Scholar]
- 65.Duff SE, Li C, Garland JM, Kumar S. CD105 is important for angiogenesis: evidence and potential applications. FASEB J. 2003;17:984–992. doi: 10.1096/fj.02-0634rev. [DOI] [PubMed] [Google Scholar]
- 66.Saad RS, Liu YL, Nathan G, Celebrezze J, Medich D, Silverman JF. Endoglin (CD105) and vascular endothelial growth factor as prognostic markers in colorectal cancer. Mod Pathol. 2004;17:197–203. doi: 10.1038/modpathol.3800034. [DOI] [PubMed] [Google Scholar]
- 67.Meurer SK, Tihaa L, Lahme B, Gressner AM, Weiskirchen R. Identification of endoglin in rat hepatic stellate cells: new insights into transforming growth factor beta receptor signaling. J Biol Chem. 2005;280:3078–3087. doi: 10.1074/jbc.M405411200. [DOI] [PubMed] [Google Scholar]
- 68.Elvevold KH, Nedredal GI, Revhaug A, Smedsrød B. Scavenger properties of cultivated pig liver endothelial cells. Comp Hepatol. 2004;3:4. doi: 10.1186/1476-5926-3-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Prevo R, Banerji S, Ni J, Jackson DG. Rapid plasma membrane-endosomal trafficking of the lymph node sinus and high endothelial venule scavenger receptor/homing receptor stabilin-1 (FEEL-1/CLEVER-1) J Biol Chem. 2004;279:52580–52592. doi: 10.1074/jbc.M406897200. [DOI] [PubMed] [Google Scholar]
- 70.Hansen B, Longati P, Elvevold K, Nedredal GI, Schledzewski K, Olsen R, Falkowski M, Kzhyshkowska J, Carlsson F, Johansson S, et al. Stabilin-1 and stabilin-2 are both directed into the early endocytic pathway in hepatic sinusoidal endothelium via interactions with clathrin/AP-2, independent of ligand binding. Exp Cell Res. 2005;303:160–173. doi: 10.1016/j.yexcr.2004.09.017. [DOI] [PubMed] [Google Scholar]
- 71.Politz O, Gratchev A, McCourt PA, Schledzewski K, Guillot P, Johansson S, Svineng G, Franke P, Kannicht C, Kzhyshkowska J, et al. Stabilin-1 and -2 constitute a novel family of fasciclin-like hyaluronan receptor homologues. Biochem J. 2002;362:155–164. doi: 10.1042/0264-6021:3620155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Tamura Y, Adachi H, Osuga J, Ohashi K, Yahagi N, Sekiya M, Okazaki H, Tomita S, Iizuka Y, Shimano H, et al. FEEL-1 and FEEL-2 are endocytic receptors for advanced glycation end products. J Biol Chem. 2003;278:12613–12617. doi: 10.1074/jbc.M210211200. [DOI] [PubMed] [Google Scholar]
- 73.Irjala H, Elima K, Johansson EL, Merinen M, Kontula K, Alanen K, Grenman R, Salmi M, Jalkanen S. The same endothelial receptor controls lymphocyte traffic both in vascular and lymphatic vessels. Eur J Immunol. 2003;33:815–824. doi: 10.1002/eji.200323859. [DOI] [PubMed] [Google Scholar]
- 74.Salmi M, Koskinen K, Henttinen T, Elima K, Jalkanen S. CLEVER-1 mediates lymphocyte transmigration through vascular and lymphatic endothelium. Blood. 2004;104:3849–3857. doi: 10.1182/blood-2004-01-0222. [DOI] [PubMed] [Google Scholar]
- 75.Jackson DG. Biology of the lymphatic marker LYVE-1 and applications in research into lymphatic trafficking and lymphangiogenesis. APMIS. 2004;112:526–538. doi: 10.1111/j.1600-0463.2004.apm11207-0811.x. [DOI] [PubMed] [Google Scholar]
- 76.Mouta Carreira C, Nasser SM, di Tomaso E, Padera TP, Boucher Y, Tomarev SI, Jain RK. LYVE-1 is not restricted to the lymph vessels: expression in normal liver blood sinusoids and down-regulation in human liver cancer and cirrhosis. Cancer Res. 2001;61:8079–8084. [PubMed] [Google Scholar]
- 77.van Oosten M, van de Bilt E, van Berkel TJ, Kuiper J. New scavenger receptor-like receptors for the binding of lipopolysaccharide to liver endothelial and Kupffer cells. Infect Immun. 1998;66:5107–5112. doi: 10.1128/iai.66.11.5107-5112.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Malerød L, Juvet K, Gjøen T, Berg T. The expression of scavenger receptor class B, type I (SR-BI) and caveolin-1 in parenchymal and nonparenchymal liver cells. Cell Tissue Res. 2002;307:173–180. doi: 10.1007/s00441-001-0476-9. [DOI] [PubMed] [Google Scholar]
- 79.Scoazec JY, Feldmann G. In situ immunophenotyping study of endothelial cells of the human hepatic sinusoid: results and functional implications. Hepatology. 1991;14:789–797. doi: 10.1002/hep.1840140508. [DOI] [PubMed] [Google Scholar]
- 80.Ohgami N, Nagai R, Ikemoto M, Arai H, Kuniyasu A, Horiuchi S, Nakayama H. CD36, a member of class B scavenger receptor family, is a receptor for advanced glycation end products. Ann N Y Acad Sci. 2001;947:350–355. doi: 10.1111/j.1749-6632.2001.tb03961.x. [DOI] [PubMed] [Google Scholar]
- 81.Mota MM, Jarra W, Hirst E, Patnaik PK, Holder AA. Plasmodium chabaudi-infected erythrocytes adhere to CD36 and bind to microvascular endothelial cells in an organ-specific way. Infect Immun. 2000;68:4135–4144. doi: 10.1128/iai.68.7.4135-4144.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Bartosch B, Vitelli A, Granier C, Goujon C, Dubuisson J, Pascale S, Scarselli E, Cortese R, Nicosia A, Cosset FL. Cell entry of hepatitis C virus requires a set of co-receptors that include the CD81 tetraspanin and the SR-B1 scavenger receptor. J Biol Chem. 2003;278:41624–41630. doi: 10.1074/jbc.M305289200. [DOI] [PubMed] [Google Scholar]
- 83.Engering A, van Vliet SJ, Hebeda K, Jackson DG, Prevo R, Singh SK, Geijtenbeek TB, van Krieken H, van Kooyk Y. Dynamic populations of dendritic cell-specific ICAM-3 grabbing nonintegrin-positive immature dendritic cells and liver/lymph node-specific ICAM-3 grabbing nonintegrin-positive endothelial cells in the outer zones of the paracortex of human lymph nodes. Am J Pathol. 2004;164:1587–1595. doi: 10.1016/S0002-9440(10)63717-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.van Kooyk Y, Geijtenbeek TB. DC-SIGN: escape mechanism for pathogens. Nat Rev Immunol. 2003;3:697–709. doi: 10.1038/nri1182. [DOI] [PubMed] [Google Scholar]
- 85.Cocquerel L, Voisset C, Dubuisson J. Hepatitis C virus entry: potential receptors and their biological functions. J Gen Virol. 2006;87:1075–1084. doi: 10.1099/vir.0.81646-0. [DOI] [PubMed] [Google Scholar]
- 86.Bashirova AA, Geijtenbeek TB, van Duijnhoven GC, van Vliet SJ, Eilering JB, Martin MP, Wu L, Martin TD, Viebig N, Knolle PA, et al. A dendritic cell-specific intercellular adhesion molecule 3-grabbing nonintegrin (DC-SIGN)-related protein is highly expressed on human liver sinusoidal endothelial cells and promotes HIV-1 infection. J Exp Med. 2001;193:671–678. doi: 10.1084/jem.193.6.671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Mukhtar M, Harley S, Chen P, BouHamdan M, Patel C, Acheampong E, Pomerantz RJ. Primary isolated human brain microvascular endothelial cells express diverse HIV/SIV-associated chemokine coreceptors and DC-SIGN and L-SIGN. Virology. 2002;297:78–88. doi: 10.1006/viro.2002.1376. [DOI] [PubMed] [Google Scholar]
- 88.Cormier EG, Durso RJ, Tsamis F, Boussemart L, Manix C, Olson WC, Gardner JP, Dragic T. L-SIGN (CD209L) and DC-SIGN (CD209) mediate transinfection of liver cells by hepatitis C virus. Proc Natl Acad Sci USA. 2004;101:14067–14072. doi: 10.1073/pnas.0405695101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Gramberg T, Hofmann H, Möller P, Lalor PF, Marzi A, Geier M, Krumbiegel M, Winkler T, Kirchhoff F, Adams DH, et al. LSECtin interacts with filovirus glycoproteins and the spike protein of SARS coronavirus. Virology. 2005;340:224–236. doi: 10.1016/j.virol.2005.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Housset C, Lamas E, Courgnaud V, Boucher O, Girard PM, Marche C, Brechot C. Presence of HIV-1 in human parenchymal and non-parenchymal liver cells in vivo. J Hepatol. 1993;19:252–258. doi: 10.1016/s0168-8278(05)80579-3. [DOI] [PubMed] [Google Scholar]
- 91.Breiner KM, Schaller H, Knolle PA. Endothelial cell-mediated uptake of a hepatitis B virus: a new concept of liver targeting of hepatotropic microorganisms. Hepatology. 2001;34:803–808. doi: 10.1053/jhep.2001.27810. [DOI] [PubMed] [Google Scholar]
- 92.Knolle PA, Limmer A. Control of immune responses by savenger liver endothelial cells. Swiss Med Wkly. 2003;133:501–506. doi: 10.4414/smw.2003.10261. [DOI] [PubMed] [Google Scholar]
- 93.Onoe T, Ohdan H, Tokita D, Shishida M, Tanaka Y, Hara H, Zhou W, Ishiyama K, Mitsuta H, Ide K, et al. Liver sinusoidal endothelial cells tolerize T cells across MHC barriers in mice. J Immunol. 2005;175:139–146. doi: 10.4049/jimmunol.175.1.139. [DOI] [PubMed] [Google Scholar]
- 94.Tokita D, Ohdan H, Onoe T, Hara H, Tanaka Y, Asahara T. Liver sinusoidal endothelial cells contribute to alloreactive T-cell tolerance induced by portal venous injection of donor splenocytes. Transpl Int. 2005;18:237–245. doi: 10.1111/j.1432-2277.2004.00045.x. [DOI] [PubMed] [Google Scholar]
- 95.Klein I, Crispe IN. Complete differentiation of CD8+ T cells activated locally within the transplanted liver. J Exp Med. 2006;203:437–447. doi: 10.1084/jem.20051775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Lohse AW, Knolle PA, Bilo K, Uhrig A, Waldmann C, Ibe M, Schmitt E, Gerken G, Meyer Zum Büschenfelde KH. Antigen-presenting function and B7 expression of murine sinusoidal endothelial cells and Kupffer cells. Gastroenterology. 1996;110:1175–1181. doi: 10.1053/gast.1996.v110.pm8613007. [DOI] [PubMed] [Google Scholar]
- 97.Knolle PA, Uhrig A, Hegenbarth S, Löser E, Schmitt E, Gerken G, Lohse AW. IL-10 down-regulates T cell activation by antigen-presenting liver sinusoidal endothelial cells through decreased antigen uptake via the mannose receptor and lowered surface expression of accessory molecules. Clin Exp Immunol. 1998;114:427–433. doi: 10.1046/j.1365-2249.1998.00713.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ahmed-Choudhury J, Russell CL, Randhawa S, Young LS, Adams DH, Afford SC. Differential induction of nuclear factor-kappaB and activator protein-1 activity after CD40 ligation is associated with primary human hepatocyte apoptosis or intrahepatic endothelial cell proliferation. Mol Biol Cell. 2003;14:1334–1345. doi: 10.1091/mbc.E02-07-0378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Leifeld L, Trautwein C, Dumoulin FL, Manns MP, Sauerbruch T, Spengler U. Enhanced expression of CD80 (B7-1), CD86 (B7-2), and CD40 and their ligands CD28 and CD154 in fulminant hepatic failure. Am J Pathol. 1999;154:1711–1720. doi: 10.1016/S0002-9440(10)65427-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Lammert E, Cleaver O, Melton D. Role of endothelial cells in early pancreas and liver development. Mech Dev. 2003;120:59–64. doi: 10.1016/s0925-4773(02)00332-5. [DOI] [PubMed] [Google Scholar]
- 101.Salmi M, Jalkanen S. Cell-surface enzymes in control of leukocyte trafficking. Nat Rev Immunol. 2005;5:760–771. doi: 10.1038/nri1705. [DOI] [PubMed] [Google Scholar]
- 102.Bonder CS, Norman MU, Swain MG, Zbytnuik LD, Yamanouchi J, Santamaria P, Ajuebor M, Salmi M, Jalkanen S, Kubes P. Rules of recruitment for Th1 and Th2 lymphocytes in inflamed liver: a role for alpha-4 integrin and vascular adhesion protein-1. Immunity. 2005;23:153–163. doi: 10.1016/j.immuni.2005.06.007. [DOI] [PubMed] [Google Scholar]
- 103.Salmi M, Kalimo K, Jalkanen S. Induction and function of vascular adhesion protein-1 at sites of inflammation. J Exp Med. 1993;178:2255–2260. doi: 10.1084/jem.178.6.2255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Salmi M, Hellman J, Jalkanen S. The role of two distinct endothelial molecules, vascular adhesion protein-1 and peripheral lymph node addressin, in the binding of lymphocyte subsets to human lymph nodes. J Immunol. 1998;160:5629–5636. [PubMed] [Google Scholar]
- 105.Ikeda Y, Terashima T. Expression of reelin, the gene responsible for the reeler mutation, in embryonic development and adulthood in the mouse. Dev Dyn. 1997;210:157–172. doi: 10.1002/(SICI)1097-0177(199710)210:2<157::AID-AJA8>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 106.Samama B, Boehm N. Reelin immunoreactivity in lymphatics and liver during development and adult life. Anat Rec A Discov Mol Cell Evol Biol. 2005;285:595–599. doi: 10.1002/ar.a.20202. [DOI] [PubMed] [Google Scholar]
- 107.Hong YK, Detmar M. Prox1, master regulator of the lymphatic vasculature phenotype. Cell Tissue Res. 2003;314:85–92. doi: 10.1007/s00441-003-0747-8. [DOI] [PubMed] [Google Scholar]