

Abstract
During development,axon guidance receptors play a crucial role in regulating axons sensitivity to both attractive and repulsive cues. Indeed, activation of the guidance receptors is the first step of the signaling mechanisms allowing axon tips, the growth cones, to respond to the ligands. As such, the modulation of their availability at the cell surface is one of the mechanisms that participate in setting the growth cone sensitivity. We describe here a method to precisely visualize the spatio-temporal cell surface dynamics of an axon guidance receptor both in vitro and in vivo in the developing chick spinal cord. We took advantage of the pH-dependent fluorescence property of a green fluorescent protein (GFP) variant to specifically detect the fraction of the axon guidance receptor that is addressed to the plasma membrane. We first describe the in vitro validation of such pH-dependent constructs and we further detail their use in vivo, in the chick spinal chord, to assess the spatio-temporal dynamics of the axon guidance receptor of interest.
Keywords: Neuroscience, Issue 83, Neurons, Axons, Cell Differentiation, Embryonic Development, Life Sciences (General), Axon guidance receptor, trafficking, pHluorin, in ovo electroporation, commissural neurons, Plexin,
Introduction
During their navigation, axons integrate multiple environmental cues that guide them towards their target. These cues activate guidance receptors at the surface of axon terminals, the growth cones, which in turn initiate an appropriate signaling pathway. Thus, the temporal and spatial regulation of the cell surface distribution of the receptors is critical to set the sensitivity of the growth cone1. In this context, midline crossing by commissural axons is an excellent model to investigate the regulation of receptor cell surface levels. In the developing spinal cord, commissural axons are initially attracted towards the ventral floor plate where they cross the midline. After crossing, they lose their responsiveness to the floor plate attractants and gain response to floor plate repellents so that they can exit the floor plate and navigate towards their final destination in the contralateral side of the nervous system2,3. Regulation of receptor availability at the growth cone surface is one of the mechanisms underlying the switch of responsiveness to midline cues4,5. Thus, selective monitoring of the receptors present at the plasma membrane of growth cones is of prime importance. We describe here a method based on the pH-dependent fluorescence property of a green fluorescent protein (GFP) variant to specifically visualize the axon guidance receptors that are addressed to the plasma membrane in vitro and in vivo, in the developing chick spinal cord.
Rothman and colleagues engineered by point mutations pH-sensitive variants of GFP including the ecliptic pHluorin6. Ecliptic pHluorin has the property of being nonfluorescent when exposed to acidic pH (<6), while being fluorescent at neutral pH. This allows distinguishing nonfluorescent receptors localized in intracellular acidic compartments (i.e. endosomes, trafficking vesicles) from fluorescent receptors incorporated to the plasma membrane and thus exposed to the extracellular neutral pH7. We took advantage of this to monitor the plasma membrane localization of plexinA1, an axon guidance receptor mediating the growth cone response to the midline repellent semaphorin 3B5 (Figure 1A). We describe here the in vitro characterization of a pHluorin-plexinA1 construct, along with in ovo electroporation8-10 of this construct in the developing chick spinal cord followed by the microscopic analysis of cryosections which enable to follow the axon guidance receptor dynamics in vivo with both spatial and temporal resolutions.
Protocol
1. Cloning Strategy to Tag PlexinA1 Receptor with pHluorin
Choose an appropriate expression vector as a backbone (e.g. the mouse receptor plexinA1 expressing vector, a kind gift of Dr. Andreas Puschel11). Note: This plexinA1 vector was engineered to achieve efficient HA- or VSV-tagged receptor insertion in the plasma membrane.
Amplify by PCR the ecliptic pHluorin coding sequence using the adequate plasmid as a template (e.g. pHluorin-tagged GABA A receptor, a kind gift of Dr. Jacob2). If necessary, add a restriction site to the 5’ end of the primer in order to facilitate the cloning step in the backbone.
Insert the pHluorin sequence in frame between the signal peptide and the receptor coding sequence using restriction sites (e.g. NruI/Kpn2I restriction sites as described in Figure 1B). Note: Because the signal peptide that ensures the correct targeting of the receptors is cleaved, the pHluorin should be placed after it. This warrants the recognition of the signal peptide and prevents cleavage of the pHluorin from the receptor of interest.
Sequence the constructs obtained to ensure that no mutation was introduced by PCR.
2. Characterization of pHluorin-tagged Receptor In vitro in COS7 Cells
The ability of the fusion protein to reach the plasma membrane and its reversible loss of fluorescence as pH is lowered can be confirmed using the following procedure.
Day 1. Plate 1.5 x 105 COS7 cells in a glass-bottom 35 mm dish in 2 ml of complete Dulbecco’s Modified Eagle Medium (DMEM - 10% Fetal Bovine Serum - 1 mM sodium pyruvate - 25 U/ml penicillin/stretomycin - 2.5 µg/ml Amphotericin B - pH 7.4).
- Day 2. Transfect cells: Note: Cells should be 70-80% confluent.
- Prepare 200 µl NaCl 150 mM and add 3 µg DNA i.e. the vector encoding pHluorin-tagged receptor. Gently vortex and spin down briefly. Note: A map of the pHluorin-plexinA1 vector used in these experiments is shown in Figure 1B.
- Add 10 µl of transfection reagent (or the appropriate amount of the reagent used). Vortex immediately.
- Incubate 10 min at RT.
- Add 200 µl of the transfection reagent/DNA mix to the cells.
- Move the plate gently to achieve repartition of the mix and place the cells back to the 37 °C incubator.
Day 3. Remove transfection medium and replace it by 2 ml of fresh complete DMEM.
- Day 4. Perform live cell imaging of COS7 pHluorin-plexinA1 transfected cells:
- Prepare two aliquots of DMEM complete medium and adjust pH to 3.5 and to 9.5, respectively. Note: For one 35 mm plate, 1.5 ml of each solution is needed to perform the experiment.
- Remove cell medium and replace it by 1 ml of DMEM complete medium pH 7.4.
- Prepare a 5 ml syringe with an appropriate type of tubing in order to inject various components directly into the cell culture medium without opening the microscope chamber.
- Use a module that allows maintenance of a 37 °C, 5% CO2 humid working atmosphere. Note: An alternate approach to the use of a CO2 chamber is to use HEPES-buffered medium (usually in the range of 10-25 mM according to the cell type).
- Place cells in the chamber and adjust the tubing and the syringe. Note: The microscope should be equilibrated before starting to avoid mechanical drift during the recording.
- Open the imaging software and select the multidimensional acquisition program.
- Find transfected COS7 cells with the 40X objective and mark position in the software for each of them.
- Configure Z stack in order to have a 15 µm depth acquisition (the focus can change when adding media in the plate).
- Set up exposure for the GFP filter and the Phase.
- Configure acquisition timing. Note: For the whole experiment with 5 fields of interest, acquisition every 20 sec for 10 min should be sufficient.
- Start acquisition and take 5 control images in DMEM pH 7.4 medium.
- Pause acquisition, inject 1.25 ml of pH 3.5 complete DMEM to achieve a pH of 5.5 in the culture medium, mark event in the software and resume acquisition for 5 more time points. Note: Green fluorescence should progressively disappear.
- Pause acquisition, inject 1.2 ml of pH 9.5 complete DMEM to achieve a pH 7.4 in the culture medium, mark event in the software and resume acquisition for 5 more time points. Note: Green fluorescence should reappear at the plasma membrane.
- Analyze images. Figure 2 shows representative images obtained with such a protocol with the pHluorin-plexinA1 construct.
3. In ovo Electroporation of pHluorin-plexinA1 Construct
- Handling of eggs before electroporation:
- Store fertilized eggs in a fridge at 14 °C up to one week prior to incubation.
- Incubate eggs at 38.5 °C (101.3 °F) in an incubator with saturated humidity for 50-52 hr until the embryos reach stage HH1412. Note: The eggs must be placed horizontally during incubation so that the embryo is properly positioned for electroporation, floating on the top of the yolk. HH14 stage is suitable to obtain expression of the plasmids in differentiated neurons in the spinal cord and in the dorsal root ganglion with an appropriate survival rate.
- Electroporate embryos8-10:
- Prepare electroporation:
- Prepare endotoxin-free DNA plasmids with a concentration superior to 2 µg/µl, to be able to dilute it as the desired concentration.
- Pull enough glass capillaries to inject the different DNA solutions.
- Prepare sterile PBS (-Ca2+; -Mg2+) - 100 U/ml penicillin/streptomycin and equilibrate at 38.5 °C.
- Sterilize the hood, curved scissors and fine forceps.
- Control the electrode spacing. Note: A 4 mm space between the electrodes is generally used.
- Window the egg13 (Figure 3A):
- Use curved scissors to pierce the shell on the blunt side of the egg.
- Remove 2 ml of albumen using a 0.9 mm x 25 mm needle and a 5 ml syringe. Orient the needle vertically in order to avoid damaging the yolk sac.
- Cover the top of the egg with tape to maintain the integrity of the shell.
- Using curved scissors, pierce the shell in the middle of the tape to equalize the pressure when removing 2 ml of albumen from the egg. Then, cut a window large enough to visualize the embryo and be able to work on it.
- Add ~2 ml sterile warm PBS (-Ca2+; -Mg2+) - 100 U/ml penicillin/streptomycin to avoid dehydration of the embryo and make it more accessible to the manipulator.
- Inject DNA and electroporate the embryo
- Dilute plasmid in PBS (-Ca2+; -Mg2+) to a concentration between 0.5-2 µg/µl and add Fast green dye to reach a final 0.025% concentration. Load the DNA mix in a capillary. The use of an injector is recommended. Note: Check that the capillary resistance is neither too large (meaning there could be difficulties when injecting embryos) nor too small (meaning that the capillary could be too big and could damage the embryo). Also, concentration of nucleic acids higher than 2 µg/µl may cause unspecific effects and needs to be controlled.
- Puncture the yolk-sac and the neural tube at the caudal side with the loaded capillary. Enter the neural tube with a shallow angle and fill the lumen from the tail to the head with the DNA mix (Figure 3B). Note: Fast green allows controlling the accuracy of the injection.
- Quickly place the 4 mm platinum electrodes on either side of the neural tube at the level you wish to electroporate and apply 3 pulses at 31 V for 50 msec with 500 msec intervals (Figure 3C). Note: Avoid placing the electrodes on the heart or on big extra embryonic vessels to avoid damaging the developing embryo. Bubbles should form on the electrodes.
- With the needle, remove 2 ml of albumen to decrease the level in the shell.
- Seal the window and the blunt side hermetically with tape.
- Put the eggs back at 38.5 °C in the incubator until they reach the desired stage.
4. Embryos Embedding and Cryosectioning
48 hr after electroporation, carefully harvest electroporated embryos (HH24 stage). Cut the tape and half of the chorioallantoid membrane. To prevent embryos from sinking into the yolk, position a colander under the embryo and cut the second half of the chorioallantoid membrane.
Transfer the embryo to a dissection dish filled with ice cold PBS.
Check electroporation efficiency by looking for fluorescence in the neural tube with a fluorescence stereo microscope. Note : Coelectroporation of a control plasmid encoding the RFP may help to visualize the electroporated area.
Dissect embryos using a microscalpel in order to select the electroporated area of the spinal cord.
Transfer dissected embryos to a 24-well plate and fix in pH 7.4 4% Paraformaldehyde (PFA) - Phosphate Buffer Saline (PBS), O/N at 4 °C. Note: Fixation step is crucial to allow the stabilization of pHluorin in its “live” conformation and thus to be able to use pHluorin in fixed/permeabilized tissue. Although fixation slows drastically pH-dependent change of fluorescence, one must take into account that conformational/protonation changes can still occur after fixation. Thus, the following protocol (embedding, cryosections and observation) has to be performed within 3 days after the fixation step, with all buffers at pH 7. If fixation is not required, performing observation on live tissue sections is recommended.
Remove 4% PFA and wash embryos in pH 7.4 PBS.
Incubate embryos in PBS-15% sucrose and keep at 4 °C until the embryos sink.
Incubate fixed embryos in pH 7.4 7.5% gelatin- 15% sucrose for 45 min at 37 °C so embryos are completely embedded.
Place embedding molds on ice and add 400 µl of pH 7.4 7.5% gelatin- 15% sucrose to achieve a solid 2 mm base.
Aspirate the embedded embryo with a cut tip and place the embryo on the solid gelatin base.
Cover with pH 7.4 7.5% gelatin- 15% sucrose and position the embryo with forceps before the gelatin solidifies.
Once the gelatin is solid, prepare a -40 °C isopropanol bath (use dry ice or liquid nitrogen) and freeze the gelatin block for 5 min.
Keep the frozen blocks at -80 °C.
Place the frozen block at -20 °C for 1 hr.
Remove the mold and fix the block on a chuck with a polyethylene glycol medium.
Once the block is tightly fixed, place the chuck in the cryostat system. Note: Use coated slides to avoid tissue loss during staining.
Perform serial cryosections (20 µm cryosections are usually performed).
Let the cryosections dry for 15 min at RT. Note: cryosections should be protected from unnecessary light exposure in order to avoid bleaching of the GFP fluorescence.
5. Microscopic Analysis of Cryosections
Rehydrate cryosections in pH 7.4 PBS at RT for 10 min. Note: If needed, nuclei can be stained with Hoechst.
Use a 0.5 µg/ml Hoechst solution in PBS and incubate cryosections for 15 min.
Rinse the slides 3x with pH 7.4 PBS for 5 min.
Proceed to slide mounting. A pH 7.4 (or more basic) polyvinyl alcohol mounting solution that hardens O/N can be used: carefully position the coverslip to avoid the formation of air bubbles between the slide and the coverslip.
Let the mounting medium harden O/N at 4 °C in the dark.
Use an inverted confocal microscope to precisely visualize the pHluorin-plexinA1 fusion protein: perform z-stack at the optimal pinhole and optical resolution and use a 20X (NA 0.75) or 40X (NA 1.3) lenses. Note: pHluorin is detectable with the same parameters used to detect GFP (i.e. emission peak at 509 nm). Wavelength excitation and detection filter settings are optimally defined by the imaging software. Hoechst is detected between 425-460 nm (excitation is at 405 nm), GFP or pHluorin is detected between 485-545 nm (excitation is at 473 nm) and RFP is detected between 575-675 nm (excitation is at 559 nm).
Representative images of pHluorin-plexinA1 and eGFP expression in the chick embryo spinal cord are shown in Figure 4.
Representative Results
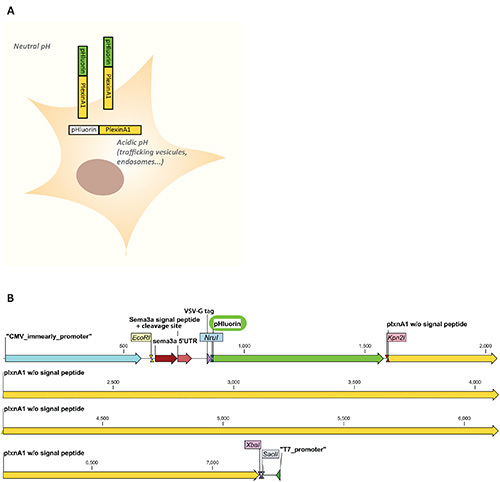 Figure 1. A. Scheme of the pHluorin-plexinA1 fluorescence properties in a cellular context. PHluorin is nonfluorescent in intracellular compartments where the pH is acidic (<6) such as in trafficking vesicules or in endosomes and is fluorescent when exposed to the extracellular medium where the pH is neutral. This allows visualizing only the cell surface pool of the pHluorin-plexinA1 receptor. B. Map of the pBK-CMV-pHluorin-plexinA1 cloning site. Mouse receptor plexinA1 expressing vector was used as a backbone. This vector was engineered to achieve efficient VSV-tagged receptor insertion at the plasma membrane. To do so, semaphorin 3a peptide signal and cleavage site were fused upstream of the VSV-tagged receptor ablated from its own signal peptide and cleavage site. By PCR, pHluorin sequence was inserted in frame between VSV-tag and PlxnA1 coding sequences using NruI/Kpn2I restriction sites. Single restriction sites are indicated for designing cloning strategy of other receptors. The scheme is adapted from bioinformatic software.
Figure 1. A. Scheme of the pHluorin-plexinA1 fluorescence properties in a cellular context. PHluorin is nonfluorescent in intracellular compartments where the pH is acidic (<6) such as in trafficking vesicules or in endosomes and is fluorescent when exposed to the extracellular medium where the pH is neutral. This allows visualizing only the cell surface pool of the pHluorin-plexinA1 receptor. B. Map of the pBK-CMV-pHluorin-plexinA1 cloning site. Mouse receptor plexinA1 expressing vector was used as a backbone. This vector was engineered to achieve efficient VSV-tagged receptor insertion at the plasma membrane. To do so, semaphorin 3a peptide signal and cleavage site were fused upstream of the VSV-tagged receptor ablated from its own signal peptide and cleavage site. By PCR, pHluorin sequence was inserted in frame between VSV-tag and PlxnA1 coding sequences using NruI/Kpn2I restriction sites. Single restriction sites are indicated for designing cloning strategy of other receptors. The scheme is adapted from bioinformatic software.
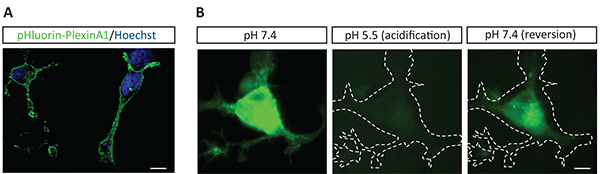 Figure 2. Characterization of the pHluorin-plexinA1 construct pH-dependent fluorescent properties in COS7 cells. A. COS7 cells transfected with the pHluorin-plexinA1 construct where observed with a confocal microscope allowing visualization of pHluorin-plexinA1 fluorescence at the plasma membrane. Scale bar: 20 µM. B. COS7 cells transfected with the pHluorin-plexinA1 construct where observed through live imaging in a pH 7.4 culture medium, after acidification of the culture medium up to pH 5.5 and after restoration of pH 7.4 in the culture medium. Cell outlines are indicated with a dashed line on GFP fluorescence acquisition. Scale bar: 10 µM. Click here to view larger image.
Figure 2. Characterization of the pHluorin-plexinA1 construct pH-dependent fluorescent properties in COS7 cells. A. COS7 cells transfected with the pHluorin-plexinA1 construct where observed with a confocal microscope allowing visualization of pHluorin-plexinA1 fluorescence at the plasma membrane. Scale bar: 20 µM. B. COS7 cells transfected with the pHluorin-plexinA1 construct where observed through live imaging in a pH 7.4 culture medium, after acidification of the culture medium up to pH 5.5 and after restoration of pH 7.4 in the culture medium. Cell outlines are indicated with a dashed line on GFP fluorescence acquisition. Scale bar: 10 µM. Click here to view larger image.
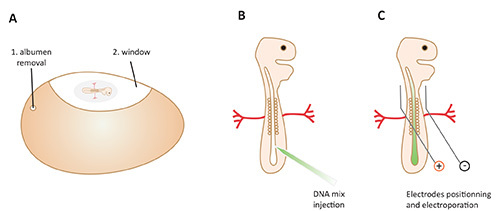 Figure 3. In ovo electroporation procedure. A. 1. Albumen removal from the largest side of the shell. 2. Windowing the egg shell to access the embryo. B. DNA/Fast Green mix injection in the neural tube. C. Placing electrodes on either sides of the neural tube, avoiding the heart and big vessels, and electroporation of the chick embryo.
Figure 3. In ovo electroporation procedure. A. 1. Albumen removal from the largest side of the shell. 2. Windowing the egg shell to access the embryo. B. DNA/Fast Green mix injection in the neural tube. C. Placing electrodes on either sides of the neural tube, avoiding the heart and big vessels, and electroporation of the chick embryo.
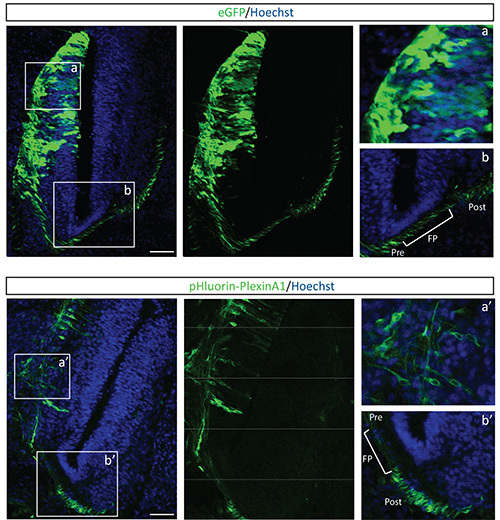 Figure 4. Microscopic analysis of chick neural tube cryosections after pHluorin-plexinA1 electroporation. A comparison between eGFP (upper panels) and pHluorin-plexinA1 (lower panels) expression pattern in the electroporated chick spinal cord is shown. Enlarged panels show the membranous fluorescence of pHluorin-plexinA1 (a’) as compared to the diffuse subcellular localization of eGFP (a). Regarding plexinA1, enlarged panel shows that this receptor is specifically enriched at the plasma membrane upon floor plate crossing (b’) which is not the case for eGFP that is equally expressed before and after floor plate crossing (b). Scale bars: 100 µM. FP: Floor Plate; Pre: Precrossing; Post: Postcrossing. Click here to view larger image.
Figure 4. Microscopic analysis of chick neural tube cryosections after pHluorin-plexinA1 electroporation. A comparison between eGFP (upper panels) and pHluorin-plexinA1 (lower panels) expression pattern in the electroporated chick spinal cord is shown. Enlarged panels show the membranous fluorescence of pHluorin-plexinA1 (a’) as compared to the diffuse subcellular localization of eGFP (a). Regarding plexinA1, enlarged panel shows that this receptor is specifically enriched at the plasma membrane upon floor plate crossing (b’) which is not the case for eGFP that is equally expressed before and after floor plate crossing (b). Scale bars: 100 µM. FP: Floor Plate; Pre: Precrossing; Post: Postcrossing. Click here to view larger image.
Discussion
This protocol provides a step-by-step procedure to follow the dynamics of an axon guidance receptor both in cell culture and in the developmental context of the chick embryo spinal cord.
To design a de novo pHluorin tagged protein, two points need to be considered regarding the cloning strategy. First, the pHluorin tag should be exposed to the lumen of the acidic endosomes, and consequently, to the extracellular compartment in order to visualize the plasma membrane receptor pool. Thus, the correct positioning of the pHluorin sequence regarding the receptor sequence is directly dependent on the type of receptor studied (i.e. whether the N-terminal or the C-terminal part of the receptor is exposed to the extracellular compartment). Second, as explained in the protocol, the position of the pHluorin sequence relative to the signal peptide should be considered to avoid subsequent cleavage between the pHluorin and the receptor of interest.
A limitation of the technique is linked to the trafficking of the receptors after their activation at the plasma membrane. Commonly, receptors are internalized and progress through the endocytic pathway composed of functionally and physically distinct compartments15. Due to ATP-driven proton pumps, endosomes maintain an acidic pH around 6 in early endosomes, around 5 in late endosomes, lower than 5 in lysosomes, but only around 6.4 in recycling endosomes or 7.0 in caveosomes allowing fluorescence in these two endosomic compartments. This issue can be resolved in vitro using total internal reflection fluorescence (TIRF) microscopy16. A second limitation inherent to this technique is that, due to its relatively big size, pHluorin tag could disrupt the receptor activity, depending on the tag insertion site.
Despite its advantages, pHluorin has not been widely used to study the dynamics and regulation of membrane proteins during axon navigation. Seminal works have used pHluorin to study the spatio-temporal dynamics of exocytosis during growth cone turning in vitro17. In the present protocol, we illustrate how pHluorin could be used to investigate the distribution of receptors at the plasma membrane of axons in vivo. Since pHluorins are genetically encoded, they are particularly appropriate to in vivo analysis. Although we describe its use in chick after in ovo electroporation, this approach can be used in other species. Indeed electroporation works efficiently in various animal models18,19. In addition, pHluorins have been successfully used in C. elegans, Drosophila, or mouse transgenic animals20-23.
Since pH-dependent change occurs in the millisecond range, the pHluorins are particularly adapted for live imaging6,24. PHluorin fusion has, for example, been used to monitor the in vivo dynamics of synaptobrevin exocytosis in olfactory sensory neurons from transgenic mice23. In vivo live imaging of pHluorin fusions holds considerable promise as confocal microscopes adapted to live imaging (fast imaging and low phototoxicity) become more efficient and accessible25. Live imaging in vivo could also be combined with fluorescence recovery after photobleaching (FRAP) so that exocytosis or diffusion could be assessed more directly26.
The use of other pH-sensitive variants and their combinations can further extend the range of applications. The dynamic distribution of the receptor in different organelles could be monitored by using ratiometric pHluorin that changes fluorescence according to the pH of the compartment6,27. Similarly, the addition of a pH-insensitive fluorescent protein to a membrane protein fused to an ecliptic pHluorin could provide important insights into its trafficking28. In addition, the recent cloning of pHtomato29 will enable monitoring of two receptors simultaneously. This could provide important insights into the formation of receptor complexes. Since pHluorin can also be used to tag guidance cues30, dual imaging of the ligand, and its receptor is also feasible.
In this protocol, the expression rate of the fusion protein is a critical point and particularly depends on the strength of the promoter, the stability of the fusion protein and the quantity of plasmid used to transfect the cells. Indeed, when overexpressed, fusion proteins may accumulate in intracellular vesicles and a fluorescent background may appear. Thus, several optimizations may be necessary to achieve a precise visualization of the pHluorin fused protein at the cell surface. Moreover, the use of pHluorin-fusion protein in fixed tissue requires particular care during and after fixation. Indeed, the stabilization of the conformations of pHluorin fusion proteins may only be temporary. Thus, pH must be kept above 7 to prevent a loss of the pHluorin signal. Moreover observations have to be carried out shortly after fixation. Optimal timing may need to be defined by users according to their particular pHluorin fusion protein.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We thank Homaira Nawabi, Frederic Moret and Isabelle Sanyas for their help. This work is supported by CNRS, Association Francaise contre les Myopathies (AFM), ANR YADDLE, Labex DevWeCan, Labex Cortex, ERC YODA to V.C.; C.D-B and A.J are supported by a La Ligue contre le cancer and Labex DevWeCan fellowships, respectively.
References
- Winckler B, Mellman I. Trafficking guidance receptors. Cold Spring Harb. Perspect. Biol. 2010;2 doi: 10.1101/cshperspect.a001826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacob TC, et al. J. Neurosci. 2005;25:10469–10478. doi: 10.1523/JNEUROSCI.2267-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nawabi H, Castellani V. Axonal commissures in the central nervous system: how to cross the midline. Cell Mol. Life Sci. 2011;68:2539–2553. doi: 10.1007/s00018-011-0691-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keleman K, Ribeiro C, Dickson BJ. Comm function in commissural axon guidance: cell-autonomous sorting of Robo in vivo. Nat. Neurosci. 2005;8:156–163. doi: 10.1038/nn1388. [DOI] [PubMed] [Google Scholar]
- Nawabi H, et al. A midline switch of receptor processing regulates commissural axon guidance in vertebrates. Genes Dev. 2010;24:396–410. doi: 10.1101/gad.542510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miesenbock G, De Angelis DA, Rothman JE. Visualizing secretion and synaptic transmission with pH-sensitive green fluorescent proteins. Nature. 1998;394:192–195. doi: 10.1038/28190. [DOI] [PubMed] [Google Scholar]
- Miesenbock G. Synapto-pHluorins: genetically encoded reporters of synaptic transmission. Cold Spring Harb. Protoc. 2012;2012:213–217. doi: 10.1101/pdb.ip067827. [DOI] [PubMed] [Google Scholar]
- Avraham O, Zisman S, Hadas Y, Vald L, Klar A. Deciphering axonal pathways of genetically defined groups of neurons in the chick neural tube utilizing in ovo electroporation. J. Vis. Exp. 2010. pp. 1792–17. [DOI] [PMC free article] [PubMed]
- Blank MC, Chizhikov V, Millen KJ. In ovo electroporations of HH stage 10 chicken embryos. J. Vis. Exp. 2007. [DOI] [PMC free article] [PubMed]
- Wilson NH, Stoeckli ET. In ovo electroporation of miRNA-based plasmids in the developing neural tube and assessment of phenotypes by DiI injection in open-book preparations. J. Vis. Exp. 2012. [DOI] [PMC free article] [PubMed]
- Rohm B, Ottemeyer A, Lohrum M, Puschel AW. Plexin/neuropilin complexes mediate repulsion by the axonal guidance signal semaphorin 3A. Mech. Dev. 2000;93:95–104. doi: 10.1016/s0925-4773(00)00269-0. [DOI] [PubMed] [Google Scholar]
- Hamburger V, Hamilton HL. A series of normal stages in the development of the chick embryo. 1951. Dev. Dyn. 1992;195:231–272. doi: 10.1002/aja.1001950404. [DOI] [PubMed] [Google Scholar]
- Korn MJ, Cramer KS. Windowing chicken eggs for developmental studies. J. Vis. Exp. 2007. [DOI] [PMC free article] [PubMed]
- Alberts P, et al. Cdc42 and actin control polarized expression of TI-VAMP vesicles to neuronal growth cones and their fusion with the plasma membrane. Mol. Biol. Cell. 2006;17:1194–1203. doi: 10.1091/mbc.E05-07-0643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perret E, Lakkaraju A, Deborde S, Schreiner R, Rodriguez-Boulan E. Evolving endosomes: how many varieties and why. Curr. Opin. Cell Biol. 2005;17:423–434. doi: 10.1016/j.ceb.2005.06.008. [DOI] [PubMed] [Google Scholar]
- Li Y, et al. Imaging pHluorin-tagged receptor insertion to the plasma membrane in primary cultured mouse neurons. J. Vis. Exp. 2012. [DOI] [PMC free article] [PubMed]
- Tojima T, Itofusa R, Kamiguchi H. Asymmetric clathrin-mediated endocytosis drives repulsive growth cone guidance. Neuron. 2010;66:370–377. doi: 10.1016/j.neuron.2010.04.007. [DOI] [PubMed] [Google Scholar]
- Matsui A, Yoshida AC, Kubota M, Ogawa M, Shimogori T. Mouse in utero electroporation: controlled spatiotemporal gene transfection. J. Vis. Exp. 2011. pp. 3024–30. [DOI] [PMC free article] [PubMed]
- Falk J, et al. Electroporation of cDNA/Morpholinos to targeted areas of embryonic CNS in Xenopus. BMC Dev. Biol. 2007;7(107) doi: 10.1186/1471-213X-7-107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holzhausen LC, Lewis AA, Cheong KK, Brockerhoff SE. Differential role for synaptojanin 1 in rod and cone photoreceptors. J. Comp. Neurol. 2009;517:633–644. doi: 10.1002/cne.22176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shang Y, Claridge-Chang A, Sjulson L, Pypaert M, Miesenbock G. Excitatory local circuits and their implications for olfactory processing in the fly antennal lobe. Cell. 2007;128:601–612. doi: 10.1016/j.cell.2006.12.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dittman JS, Kaplan JM. Factors regulating the abundance and localization of synaptobrevin in the plasma membrane. Proc. Natl. Acad. Sci. U.S.A. 2006;103:11399–11404. doi: 10.1073/pnas.0600784103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bozza T, McGann JP, Mombaerts P, Wachowiak M. In vivo imaging of neuronal activity by targeted expression of a genetically encoded probe in the mouse. Neuron. 2004;42:9–21. doi: 10.1016/s0896-6273(04)00144-8. [DOI] [PubMed] [Google Scholar]
- Sankaranarayanan S, Ryan TA. Real-time measurements of vesicle-SNARE recycling in synapses of the central nervous system. Nat. Cell. Biol. 2000;2:197–204. doi: 10.1038/35008615. [DOI] [PubMed] [Google Scholar]
- Stark DA, Kasemeier-Kulesa JC, Kulesa PM. Photoactivation cell labeling for cell tracing in avian development. CSH Protoc. 2008;2008 doi: 10.1101/pdb.prot4975. [DOI] [PubMed] [Google Scholar]
- Hildick KL, Gonzalez-Gonzalez IM, Jaskolski F, Henley JM. Lateral diffusion and exocytosis of membrane proteins in cultured neurons assessed using fluorescence recovery and fluorescence-loss photobleaching. J. Vis. Exp. 2012. [DOI] [PMC free article] [PubMed]
- Hanson GT, et al. Green fluorescent protein variants as ratiometric dual emission pH sensors. 1. Structural characterization and preliminary application. Biochemistry. 2002;41:15477–15488. doi: 10.1021/bi026609p. [DOI] [PubMed] [Google Scholar]
- Rose T, Schoenenberger P, Jezek K, Oertner TG. Developmental refinement of vesicle cycling at schaffer collateral synapses. Neuron. 2013;77:1109–1121. doi: 10.1016/j.neuron.2013.01.021. [DOI] [PubMed] [Google Scholar]
- Li Y, Tsien RW. pHTomato, a red, genetically encoded indicator that enables multiplex interrogation of synaptic activity. Nat. Neurosci. 2012;15:1047–1053. doi: 10.1038/nn.3126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Wit J, Toonen RF, Verhage M. Matrix-dependent local retention of secretory vesicle cargo in cortical neurons. J. Neurosci. 2009;29:23–37. doi: 10.1523/JNEUROSCI.3931-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]