

Abstract
Analysis of neuromuscular junction morphology can give important insight into the physiological status of a given motor neuron. Analysis of thin flat muscles can offer significant advantage over traditionally used thicker muscles, such as those from the hind limb (e.g. gastrocnemius). Thin muscles allow for comprehensive overview of the entire innervation pattern for a given muscle, which in turn permits identification of selectively vulnerable pools of motor neurons. These muscles also allow analysis of parameters such as motor unit size, axonal branching, and terminal/nodal sprouting. A common obstacle in using such muscles is gaining the technical expertise to dissect them. In this video, we detail the protocol for dissecting the transversus abdominis (TVA) muscle from young mice and performing immunofluorescence to visualize axons and neuromuscular junctions (NMJs). We demonstrate that this technique gives a complete overview of the innervation pattern of the TVA muscle and can be used to investigate NMJ pathology in a mouse model of the childhood motor neuron disease, spinal muscular atrophy.
Keywords: Neuroscience, Issue 83, Transversus Abdominis, neuromuscular junction, NMJ, dissection, mouse, immunofluorescence
Introduction
Neuromuscular junctions (NMJs) are the synaptic connection between a lower motor neuron and a skeletal muscle fiber. They are traditionally considered a tripartite synapse, made up of a neuron (presynaptic terminal), muscle fiber (post synaptic terminal), and terminal Schwann cell1. NMJs appear to be early and significant targets in pathology in a range of motor neuron diseases and mouse models2,3. Typical symptoms include denervation, where the motor endplate becomes devoid of a presynaptic innervation, swelling of the presynaptic terminal, and a reduction in the complexity of the NMJ morphology4-11. Compensatory responses can also be noted, which include terminal and nodal sprouting, where axonal processes extend from remaining synaptic terminals or internodes to reinnervated denervated endplates12,13. Due to the tight correlation between synaptic activity and NMJ morphology, a great deal of information can be gained about the functional status of motor neurons from analysis of NMJ morphology. As loss of NMJs frequently represents one of the first aspects of neuromuscular pathology4,10, quantification at the level of innervation can give important information about the progression of pathology and the potential effect of a therapeutic intervention. Furthermore, as NMJ loss represents a significant step in pathological progression, the development of therapeutics that can stabilize connections and encourage regeneration may yield significant benefit.
When analyzing NMJ morphology, muscle choice is of great importance. Some of the primary considerations might include muscle fiber type, body position, and comparative analysis to human conditions. In addition, where manipulations such as injection of substances or traumatic nerve injury are required, experimental accessibility is also important to consider. In general, it is preferable to analyze a range of muscles positioned throughout the body reflecting a range of motor unit subtypes. Often, however, muscle choice is influenced by ease of dissection. Consequently, NMJ analysis is often performed exclusively on large appendicular muscles such as gastrocnemius. To obtain good NMJ staining in such muscles, sectioning or mechanical disruption of muscle fibers is often required. As a result, the innervation pattern may become disrupted and a comprehensive and high quality analysis of innervation patterns, sprouting and denervation is often compromised. An alternative approach is to use thin flat muscles which do not require sectioning and can be stained and mounted intact, allowing a comprehensive overview of the entire innervation of the muscle. There are a number of muscles which can be used for such an analysis, including a group of cranial muscles, (encompassing levator auris longus, auricularis superior, and adductor auris longus)14, thoracic muscles (e.g. triangularis sterni)15, and abdominal (e.g. transversus abdominis (TVA)) muscles. The major hindrance in using such muscles is the technical expertise required to dissect them without damage.
In this video, we provide a protocol to dissect and perform immunofluorescent labeling of the TVA muscle from the mouse to allow a comprehensive analysis of innervation pattern and NMJ morphology. The TVA muscle is a predominantly slow twitch muscle comprising the deepest layer of abdominal musculature and is innervated by the lower intercostal nerves. Previous work has shown it to be consistently highly vulnerable to pathology in a number of mouse models of the childhood motor neuron disease spinal muscular atrophy (SMA) and in other mouse models of early onset motor neuron degeneration4,16. We therefore suggest that the TVA is a useful muscle for undertaking NMJ analysis in peripheral neuropathies.
Protocol
All procedures should be performed to the animal care standards set out by the institution.
1. Dissection of the Abdominal Musculature from the Mouse
Before starting, make up 4% paraformaldehyde (PFA). Caution: Always keep PFA within a fume hood and wear appropriate protective equipment.
Euthanize mouse by an approved method. Note: The mouse shown in the video was euthanized by overdose of CO2 and cervical dislocation. This mouse is a 4-week-old wild-type mouse on a CD1/C57Bl6 hybrid background. This mouse was bred in the animal facilities at the University of Ottawa from mice that were originally purchased.
Make an initial incision through the skin at the level of the hip and cut through the skin all the way around the mouse.
Peel off the skin, pulling upwards until you reach the level of the forelimb.
Cut through the abdominal musculature at the level of the hip and continue the incision until you reach the vertebral column.
At this point, start to cut upwards through the musculature and the ribs until you reach the upper limb.
Cut straight across until you reach the vertebral column on the other side.
Cut downwards until you reach the point at which you started.
Release the diaphragm from underneath (taking care not to damage the TVA muscle) and place it in phosphate buffered saline (PBS) in a SYLGARD-coated dissection dish with 0.2 mm dissection pins.
Pin out the rib cage and associated musculature in the dish, superficial side up, making sure the muscle is fully stretched out.
Pour off the 1x PBS and replace with 4% PFA.
Cover and leave on a rocking platform at room temperature for 15 min.
Wash 3x in 1x PBS at room temperature for 10 min.
* At this point the fixed muscles can be left in a fridge overnight before subsequent dissection.
2. Isolation of the TVA Muscle
To proceed with isolation of the TVA muscle, under a dissection microscope, begin by cutting through the external oblique muscle at the level of the last few ribs (See Figure 1 for an annotated guide).
Cut straight down beside the linea alba (taking care not to cut through the TVA below) until you reach the start of the internal oblique muscle.
Then cut across to release the overlying rectus abdominis and external oblique muscles from the upper part of the TVA below.
Remove the blood vessel and any overlying fat from the TVA muscle.
Release the muscle from the overlying last rib.
Cut around the margins of the muscle and remove to a 24-well plate containing PBS.
3. Immunofluorescent Labeling and Microscopy of the TVA Muscle
For all subsequent steps, remove liquid using a fine tipped pipette and leave the plate on a rocking platform. Unless otherwise stated, all subsequent steps are performed at room temperature.
Incubate in PBS containing 1:1,000 fluorescently tagged bungarotoxin for 30 min to label the NMJs. This can be done in a dark room or under foil.
Permeabilize muscle by adding 300 µl per well of 2% Triton X-100 in PBS and leave on a rocking platform for 30 min.
Incubate in blocking reagent (4% BSA, 1% Triton in PBS) for 30 min.
Incubate in blocking reagent containing primary antibodies (neurofilament 1:100; synaptic vesicle protein 2, 1:250) at 4 °C overnight.
Wash 3x in 1x PBS.
Incubate in PBS containing 1:250 secondary antibody for 2-4 hr.
Wash 3x in 1x PBS.
Mount muscles onto glass slides using sufficient fluorescent mounting media and coverslip.
Slides are best viewed using a double band pass filter on a standard epifluorescent microscope.
NMJs are best imaged using a z-series projection on a confocal microscope equipped with at least a 40X objective.
Representative Results
The above protocol directs the isolation and staining of the TVA muscle for NMJ analysis. This allows whole-mount analysis of muscle innervation patterns as well as high-resolution analysis of NMJ morphology (Figure 2). This technique can be successfully applied to reveal NMJ pathology in mouse models of motor neuron disease, such as SMA4,17(Figure 3). In mouse models of SMA there is significant intramuscular variability in pathology, however the TVA muscle is consistently highly affected. This is evidenced by denervation of motor endplates, accumulation of neurofilaments at the presynaptic terminal and terminal sprouting (Figure 3). The results presented here thus demonstrate that the technique described above can be a powerful method to provide a comprehensive overview of innervation and analysis of NMJ pathology in mouse models.
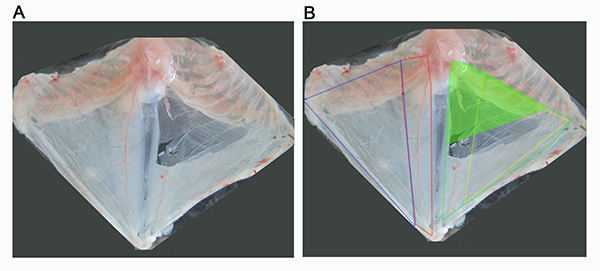 Figure 1. Overview of abdominal musculature of the mouse. (A) Image show the thoracic cage with attached abdominal musculature, which has been removed from a recently euthanized mouse and pinned out in a dissection dish superficial side up (A). (B) Dissection in A has been annotated to mark the approximate boundaries of the abdominal muscles. The superficial abdominal muscles can be seen on the left side of the dissection, and include external oblique (outlined in blue) and rectus abdominis (outlined in red). On the right hand side of the dissection, the superficial muscles have been removed allowed visualization of the transversus abdominis muscle (outlined in green) and the internal oblique muscle (outlined in yellow). The aim of this protocol is to direct the dissection of the superior part of the TVA muscle, shown here as a solid green triangle. Click here to view larger image.
Figure 1. Overview of abdominal musculature of the mouse. (A) Image show the thoracic cage with attached abdominal musculature, which has been removed from a recently euthanized mouse and pinned out in a dissection dish superficial side up (A). (B) Dissection in A has been annotated to mark the approximate boundaries of the abdominal muscles. The superficial abdominal muscles can be seen on the left side of the dissection, and include external oblique (outlined in blue) and rectus abdominis (outlined in red). On the right hand side of the dissection, the superficial muscles have been removed allowed visualization of the transversus abdominis muscle (outlined in green) and the internal oblique muscle (outlined in yellow). The aim of this protocol is to direct the dissection of the superior part of the TVA muscle, shown here as a solid green triangle. Click here to view larger image.
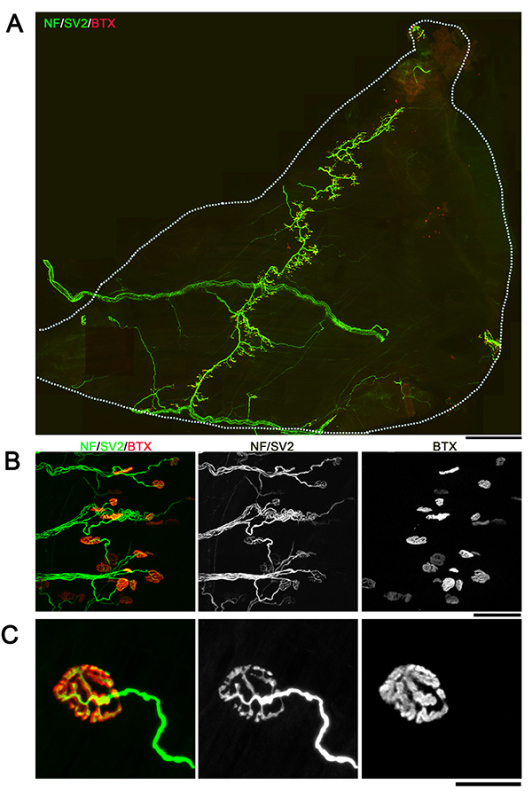 Figure 2. Whole-mount overview of neuromuscular junctions in TVA muscle. Images showing example NMJs from the TVA muscle visualized with immunofluorescent staining for neurofilament (NF; green), synaptic vesicle protein 2 (SV2; green) and bungarotoxin (BTX; red). Images are montaged fluorescent micrographs showing the whole muscle (A) or confocal images showing groups (B) or individual (C) NMJs. Scale bar = 800 µm (A), 70 µm (B), 25 µm (C). Click here to view larger image.
Figure 2. Whole-mount overview of neuromuscular junctions in TVA muscle. Images showing example NMJs from the TVA muscle visualized with immunofluorescent staining for neurofilament (NF; green), synaptic vesicle protein 2 (SV2; green) and bungarotoxin (BTX; red). Images are montaged fluorescent micrographs showing the whole muscle (A) or confocal images showing groups (B) or individual (C) NMJs. Scale bar = 800 µm (A), 70 µm (B), 25 µm (C). Click here to view larger image.
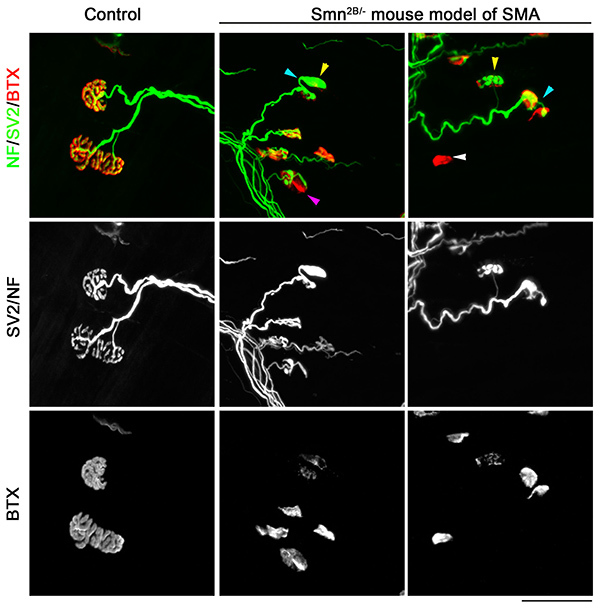 Figure 3. Neuromuscular junction pathology in TVA muscle from a mouse model of SMA. Confocal micrographs showing NMJs from the TVA muscle visualized with immunofluorescent staining for neurofilament (NF; green), synaptic vesicle protein 2 (SV2; green) and bungarotoxin (BTX; red) from either control (Smn2B/+) or SMA mouse model (Smn2B/-). Note that whilst normal NMJ morphology can be observed in control mice, in TVA muscles from Smn2B/- mice there is evidence of full denervation (white arrowhead), partial denervation (purple arrowhead) terminal sprouting (blue arrowhead) and presynaptic swelling (yellow arrowhead). The post-synaptic endplates are also less complex reflecting an apparently less mature phenotype. Scale bar = 50 µm. Click here to view larger image.
Figure 3. Neuromuscular junction pathology in TVA muscle from a mouse model of SMA. Confocal micrographs showing NMJs from the TVA muscle visualized with immunofluorescent staining for neurofilament (NF; green), synaptic vesicle protein 2 (SV2; green) and bungarotoxin (BTX; red) from either control (Smn2B/+) or SMA mouse model (Smn2B/-). Note that whilst normal NMJ morphology can be observed in control mice, in TVA muscles from Smn2B/- mice there is evidence of full denervation (white arrowhead), partial denervation (purple arrowhead) terminal sprouting (blue arrowhead) and presynaptic swelling (yellow arrowhead). The post-synaptic endplates are also less complex reflecting an apparently less mature phenotype. Scale bar = 50 µm. Click here to view larger image.
Discussion
In this video, we have detailed a protocol for the dissection of the TVA muscle from the mouse and for the whole-mount immunofluorescent labeling of NMJs within the muscle. We also present data showing this muscle can be used to analyze neuromuscular junction pathology in a mouse model of SMA.
Success in this technique is reliant on a number of factors. Some of the most common problems are outlined below. Firstly: poor immunohistochemical staining. There can be a number of reasons for this, one of the most common being use of different reagents to the ones listed in this protocol. A high quality electron microscopy grade PFA is very important to ensure good staining, as are the choice of antibodies listed in this protocol. In addition, in older animals (i.e. > 3 months), getting good quality staining can be more difficult. This is because of increased thickness of the fascia surrounding the muscle and the increase in fat accumulation between the external oblique and tranversus abdominis. It is important to strip off the fat, before proceeding to immunofluorescence. It may also be necessary to strip off some of the fascia covering the muscle, which can become thickened. It is sometimes difficult to strip the fascia and fat from the muscle without incurring some damage to the muscle fiber and a disruption to the innervation pattern. However if this technique is performed carefully, good quality staining can be acquired from mice up to at least 1 year of age. In younger mice (i.e. less than 3 months of age) it should not be necessary to perform any teasing or separation of muscle fibers. Secondly: difficultly in finding NMJs following dissection and staining. This is often because the dissection has not extended underneath the last rib. The majority of the NMJs are located just underneath the last rib and therefore care must be taken to ensure this part of the muscle is included in the dissection. Thirdly: adherence of the EO muscle to the TVA muscle. This is often a complaint when individuals attempt to extend the dissection below the level of the internal oblique (IO) muscle. The area of the TVA muscle where the IO is also present is more difficult to analyze as it can be difficult to distinguish which muscle is which. For this reason, we routinely just dissect the most superior part of the TVA muscle. At this level, there is no adherence between the EO and TVA muscles, and therefore this should not be a significant problem.
One significant hindrance to using the TVA muscle, compared to appendicular muscles, is accessibility for either surgical manipulations or injection of substances. These types of experiments can be crucial to investigate NMJ physiology in a given muscle. Although the TVA is certainly less easily accessible than more commonly used muscles such as tibialis anterior or gastrocnemius, previous work has shown it is possible to denervate the TVA by surgical injury of the intercostal nerves18. We have recently also used this muscle for local administration of substances under general anesthetic (unpublished data). Although these experiments can represent a moderate technical challenge, this work shows that they are feasible and thus extend the usefulness of this muscle for analysis of NMJs under both pathological and physiological manipulation.
The TVA muscle is one of a number of thin flat muscles situated throughout the body that can be used for whole-mount analysis of innervation patterns. Other muscles include a group of cranial muscles innervated by motor neurons originating from the facial nucleus of the brainstem, encompassing levator auris longus, auricularis superior, and adductor auris longus, the dissections for which have been described previously14,19. Furthermore, the musculature surrounding the TVA muscle, including EO, IO, and rectus abdominis, can also be labeled and used for NMJ analysis. For comprehensive analysis of NMJ pathology in a mouse model, it is important to consider a number of muscles situated throughout the body and not to restrict analysis to a single muscle. This is exemplified in mouse models of motor neuron diseases where there is significant heterogeneity in levels of NMJ pathology between different muscles20. Such intermuscular variability is an extremely valuable tool when investigating the mechanism of motor neuron vulnerability and therefore restricting analysis to a single muscle could significantly diminish the potential of the research.
Disclosures
The authors have nothing to disclose.
Acknowledgments
This work was supported by grants from the Canadian Institutes of Health Research (grant number MOP 38040) to R.K., Muscular Dystrophy Association (USA) to R.K., Families of SMA to R.K. and L.M.M, The SMA Trust to T.H.G., and The Muscular Dystrophy Campaign to T.H.G.. L.M.M is a recipient of a Multiple Sclerosis Society of Canada Postdoctoral Fellowship, and R.K. is a recipient of a University Health Research Chair from University of Ottawa.
References
- Sanes JR, Lichtman JW. Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci. 1999;22:389–442. doi: 10.1146/annurev.neuro.22.1.389. [DOI] [PubMed] [Google Scholar]
- Dupuis L, Loeffler JP. Neuromuscular junction destruction during amyotrophic lateral sclerosis: insights from transgenic models. Curr. Opin. Pharmacol. 2009;9:341–346. doi: 10.1016/j.coph.2009.03.007. [DOI] [PubMed] [Google Scholar]
- Murray LM, Talbot K, Gillingwater TH. Review: neuromuscular synaptic vulnerability in motor neurone disease: amyotrophic lateral sclerosis and spinal muscular atrophy. Neuropathol. Appl. Neurobiol. 2010;36:133–156. doi: 10.1111/j.1365-2990.2010.01061.x. [DOI] [PubMed] [Google Scholar]
- Murray LM, et al. Selective vulnerability of motor neurons and dissociation of pre- and post-synaptic pathology at the neuromuscular junction in mouse models of spinal muscular atrophy. Hum. Mol. Genet. 2008;17:949–962. doi: 10.1093/hmg/ddm367. [DOI] [PubMed] [Google Scholar]
- Kariya S, et al. Reduced SMN protein impairs maturation of the neuromuscular junctions in mouse models of spinal muscular atrophy. Hum. Mol. Genet. 2008;17:2552–2569. doi: 10.1093/hmg/ddn156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iguchi Y, et al. Loss of TDP-43 causes age-dependent progressive motor neuron degeneration. Brain. 2013;136:1371–1382. doi: 10.1093/brain/awt029. [DOI] [PubMed] [Google Scholar]
- Martinez-Hernandez R, et al. Synaptic defects in type I spinal muscular atrophy in human development. J. Pathol. 2013;229:49–61. doi: 10.1002/path.4080. [DOI] [PubMed] [Google Scholar]
- Fischer LR, Li Y, Asress SA, Jones DP, Glass JD. Absence of SOD1 leads to oxidative stress in peripheral nerve and causes a progressive distal motor axonopathy. Exp. Neurol. 2012;233:163–171. doi: 10.1016/j.expneurol.2011.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cifuentes-Diaz C, et al. Neurofilament accumulation at the motor endplate and lack of axonal sprouting in a spinal muscular atrophy mouse model. Hum. Mol. Genet. 2002;11:1439–1447. doi: 10.1093/hmg/11.12.1439. [DOI] [PubMed] [Google Scholar]
- Fischer LR, et al. Amyotrophic lateral sclerosis is a distal axonopathy: evidence in mice and man. Exp. Neurol. 2004;185:232–240. doi: 10.1016/j.expneurol.2003.10.004. [DOI] [PubMed] [Google Scholar]
- Diers A, Kaczinski M, Grohmann K, Hubner C, Stoltenburg-Didinger G. The ultrastructure of peripheral nerve, motor end-plate and skeletal muscle in patients suffering from spinal muscular atrophy with respiratory distress type 1 (SMARD1) Acta Neuropathol. 2005;110:289–297. doi: 10.1007/s00401-005-1056-y. [DOI] [PubMed] [Google Scholar]
- Ang ET, et al. Motor axonal sprouting and neuromuscular junction loss in an animal model of Charcot-Marie-Tooth disease. J. Neuropathol. Exp. Neurol. 2010;69:281–293. doi: 10.1097/NEN.0b013e3181d1e60f. [DOI] [PubMed] [Google Scholar]
- Simon CM, Jablonka S, Ruiz R, Tabares L, Sendtner M. Ciliary neurotrophic factor-induced sprouting preserves motor function in a mouse model of mild spinal muscular atrophy. Hum. Mol. Genet. 2010;19:973–986. doi: 10.1093/hmg/ddp562. [DOI] [PubMed] [Google Scholar]
- Murray LM, Gillingwater TH, Parson SH. Using mouse cranial muscles to investigate neuromuscular pathology in vivo. Neuromuscul. Disord. 2010;20:740–743. doi: 10.1016/j.nmd.2010.06.013. [DOI] [PubMed] [Google Scholar]
- Brill MS, Marinkovic P, Misgeld T. Sequential photo-bleaching to delineate single Schwann cells at the neuromuscular junction. J. Vis. Exp. 2013. [DOI] [PMC free article] [PubMed]
- Murray LM, Thomson D, Conklin A, Wishart TM, Gillingwater TH. Loss of translation elongation factor (eEF1A2) expression in vivo differentiates between Wallerian degeneration and dying-back neuronal pathology. J. Anat. 2008;213:633–645. doi: 10.1111/j.1469-7580.2008.01007.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowerman M, Murray LM, Beauvais A, Pinheiro B, Kothary R. A critical smn threshold in mice dictates onset of an intermediate spinal muscular atrophy phenotype associated with a distinct neuromuscular junction pathology. Neuromuscul. Disord. 2012;22:263–276. doi: 10.1016/j.nmd.2011.09.007. [DOI] [PubMed] [Google Scholar]
- Comley LH, et al. ApoE isoform-specific regulation of regeneration in the peripheral nervous system. Hum. Mol. Genet. 2011;20:2406–2421. doi: 10.1093/hmg/ddr147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright M, Kim A, Son Y. Subcutaneous administration of muscarinic antagonists and triple-immunostaining of the levator auris longus muscle in mice. J. Vis. Exp. 2011. [DOI] [PMC free article] [PubMed]
- Ling KK, Gibbs RM, Feng Z, Ko CP. Severe neuromuscular denervation of clinically relevant muscles in a mouse model of spinal muscular atrophy. Hum. Mol. Genet. 2012;21:185–195. doi: 10.1093/hmg/ddr453. [DOI] [PMC free article] [PubMed] [Google Scholar]