

Abstract
Craniofacial birth defects occur in 1 out of every 700 live births, but etiology is rarely known due to limited understanding of craniofacial development. To identify where signaling pathways and tissues act during patterning of the developing face, a 'face transplant' technique has been developed in embryos of the frog Xenopus laevis. A region of presumptive facial tissue (the "Extreme Anterior Domain" (EAD)) is removed from a donor embryo at tailbud stage, and transplanted to a host embryo of the same stage, from which the equivalent region has been removed. This can be used to generate a chimeric face where the host or donor tissue has a loss or gain of function in a gene, and/or includes a lineage label. After healing, the outcome of development is monitored, and indicates roles of the signaling pathway within the donor or surrounding host tissues. Xenopus is a valuable model for face development, as the facial region is large and readily accessible for micromanipulation. Many embryos can be assayed, over a short time period since development occurs rapidly. Findings in the frog are relevant to human development, since craniofacial processes appear conserved between Xenopus and mammals.
Keywords: Developmental Biology, Issue 85, craniofacial development, neural crest, Mouth, Nostril, transplantation, Xenopus
Introduction
To understand mechanisms underlying craniofacial birth defects1-2, important tissues and their signaling contributions during craniofacial development must be identified. In the frog Xenopus laevis, part of the face, including the mouth and nostrils form from the "Extreme Anterior Domain" (EAD), where ectoderm and endoderm are directly juxtaposed3-4. The EAD also acts as a signaling center to influence surrounding tissues, including the cranial neural crest, which forms the jaws and other facial regions5. To identify genes that contribute to EAD function, a 'face transplant' technique was developed, where tissue is transplanted from a donor into a host embryo, after removing the corresponding host region. Following the transplant, resulting facial development is assessed. Thus, the effects of loss of function (LOF) or gain of function (GOF) for a specific gene in the EAD are analyzed locally, where the rest of the head and body is composed of wild type tissue. The reciprocal transplant can be performed, where wild type tissue is transplanted into embryos with global LOF or GOF in specific genes. Transplantation has been frequently used in Xenopus and chick studies6. For example, Xenopus transplantation has addressed homogenetic neural induction, lens and neural competence, and neural crest migration7-10. Quail-chick chimeric grafting has analyzed development of the anterior neural plate, anterior neural ridge, neural crest, and cranial bones11-14. This is the first transplant technique for study of craniofacial development in Xenopus. This technique has demonstrated a novel role for the Wnt inhibitors Frzb1 and Crescent in regulating basement membrane formation in the presumptive mouth5. Xenopus laevis is an ideal model for study of craniofacial development as embryos are large, develop externally, and the face is readily visible, allowing micromanipulation and imaging of development. Mechanisms underlying facial development appear conserved, indicating that findings made in the frog provide insight into human development4,15-16.
Protocol
1. Preparing Reagents
10x MBS: Prepare 1 L of 10x Modified Barth's Saline (MBS) solution17. Refer to Table 1, Reagents, ingredients, and instructions. Use distilled water for all solutions. Mix in a beaker, using a stir bar, until full dissolution. All solutions should be made at room temperature.
1x MBS: Dilute 100 ml of 10x MBS solution in 900 ml of distilled water to make 1 L of 1x MBS. Add 0.7 ml of 1 M CaCl2 solution.
0.1x MBS: Dilute 1x MBS to prepare 1 L of 0.5x MBS solution and 2 L of 0.1x MBS solution. To 1 L of 0.1x MBS solution, add 1 ml of 10 mg/ml gentamycin solution. The 0.1x MBS solution with gentamycin will be used for long-term embryo culture.
Ficoll/MBS: Add 15 grams of Ficoll 400 to 500 ml of 0.5x MBS solution. Mix vigorously. Add a stir bar and mix until Ficoll is fully dissolved (several hours).
70% ethanol: Dilute 100% ethanol to 70% ethanol using distilled water.
2. Preparing Glass Operating Tools
- Needle preparation: Load capillary tubing into a needle puller.
- Pull the needles according to the settings shown in Table 2: Needle puller settings. The settings are for a Sutter Instrument Co. Model P-80/PC micropipette puller and capillary tubings, as described in the Table of specific reagents and equipment. However, these settings are specific to the Sutter needle puller, and will need to be adjusted for other machines. Settings can be determined using a ramp test, as specified by the machine's manufacturer.
- Pull 4-6 needles in preparation for the procedure. The needles should be broken such that the flexible, hair-like portion of the glass tip is fully removed, which is typically 2-3 mm long. The tip must be relatively rigid, but still narrow enough to be used as a cutting tool. See photo of an ideal needle in Figure 1A.
- Store the needles in a Petri dish with a strip of clay down the center. Press the shaft of each needle into the clay, to hold it in place and to keep the fragile sharp tip away from the bottom and sides.
For additional tools, obtain glass cover slips, a pair of long standard pattern forceps, a Bunsen burner, and 3-4 glass Pasteur pipettes (Size 5 ¾ in).
Pipette tool: To make a pipette tool, light the Bunsen burner and place the tip of a glass Pasteur pipette into the blue part of the flame while rotating it, such that the tip melts and the hole completely seals, forming a closed, rounded end. The rounded, sealed tip is later used to make depressions in the clay dish that holds the embryos during the operation. See Figure 1B.
Glass bridges: To make glass bridges, use long standard pattern forceps to carefully break off 3 mm by 3 mm chunks of cover slip glass. While holding the piece of cover slip with tweezers, place the glass in the flame until all four edges soften and curve downward, forming a tiny glass dome. The edges should no longer be sharp. See Figure 1C.
Store the glass cover slip bridges by inserting them, edges down, into modeling clay, lining the bottom of a Petri dish. Insert the bridges such that the top of the bridge remains above the clay surface. Do not push too hard such that the bridge breaks, and do not fully embed the bridge into the clay, because it can be difficult to remove. After sterilization with a flame or 70% ethanol, the bridges can be reused from experiment to experiment.
3. Preparing for the Embryo Operation
Line a small 60 mm plastic Petri dish with modeling clay. Between uses, the dish and clay surface is thoroughly washed with distilled water and then with 70% ethanol. NOTE: Use red, white or yellow, Van Aken Plastalina modeling clay, which can be bought at a local toy or art store. Black clay is not recommended because it releases a residue.
Fill the dish with 3% Ficoll 0.5x MBS solution. The higher salt concentration prevents tissue dissociation, and the polysaccharide Ficoll helps to thicken the solution, which assists in holding the face in position.
Use the flamed Pasteur pipette tool (see Figure 1) to make shallow, 2-3 mm depressions in the clay, about the depth of a stage 20 embryo body. Make 20-30 depressions, 1-2 mm apart, on each side of the dish, so there is a total of 40-60 depressions. Label one side LOF/GOF, and the other side wild type, by carving initials into the clay with forceps.
4. Preoperation Embryo Preparation
- Obtain and culture Xenopus laevis embryos using standard methods17. For a detailed description of frog husbandry, please see Sive et al.17
- Forty-eight hours prior to the experiment obtain eggs from female frogs and perform in vitro fertilization.
- Inject 0.5-1 ng of capped membrane GFP mRNA, plus any desired mRNA or anti-sense morpholino-modified antisense oligonucleotides ("morpholinos") at the one cell stage or into 2 cells at the 2 cell stage. The one cell stage lasts approximately 70-90 min. Inject a total volume of 1-3 nl. In place of RNA coding for a fluorescent protein (such as GFP or RFP RNA), one can inject FITC labeled morpholino or fluorescently labeled dextran. The fluorescence is important for determining whether the transplanted tissue heals and remains in the head.
- Capped mRNA can be prepared from a linearized plasmid using a mMessage mMachine SP6 or T7 kit. Morpholinos can be designed and ordered through Gene Tools LLC. The amount of morpholino required for a desired effect must be determined for each gene18.
- Store the injected embryos at 15 °C for 48 hr, until they reach stage 19-20. (For all subsequent steps, stage embryos according to the Normal table of Xenopus laevis by Nieuwkoop and Faber19.) NOTE: On the day of the surgery, both recipient and donor embryos must be within one stage of each other for the transplants to work optimally. However, embryos injected with morpholinos ("morphants") sometimes develop more slowly than wild type or control morphant embryos, making it necessary to coordinate experiments so both morphant and wild type embryos are at the same stage. Morphants may need to be maintained at a higher temperature for 12-24 hr prior to the procedure. To increase the likelihood that embryos can be found at matching stages, embryos can be maintained at several temperatures. Twenty-four hours prior to the experiment, one should divide the embryos into a several dishes, and place morphants at 18-20 °C and wild type embryos at 15-18 °C.
On the day of the transplantation experiment, remove the embryos from the15 °C incubator and stage them according to Nieuwkoop and Faber19. If they are younger than late neurula (stage 19), leave them at room temperature for 1-2 hr, until they reach stage 19.
Screen the injected embryos under a fluorescent microscope. Select embryos that show uniform, bright fluorescence for the experiment.
Remove clutter and possible contaminants near the operating area. Wipe down the operating surface, stereomicroscope and all tools with 70% ethanol.
- Once the embryos are at stage 19, remove the vitelline membrane using #5/45 Dumont forceps under a stereomicroscope.
- Remove the vitelline membrane of 20-30 of each of donor and host embryos.
- For the first several face transplant experiments, one should practice with a few embryos for each condition. The method is challenging and requires careful practice before it can be used on a larger scale, quickly and successfully. One can work up to doing 20 transplants/experiment.
Move host embryos into the operating dish using a plastic graduated transfer pipette with the tip cut off such that the opening is much wider than an embryo (at least a few millimeters). Be careful to avoid touching embryos to bubbles or the water surface, as embryos will explode in the surface tension.
Use #5/45 forceps to insert the embryos into the clay depressions, with the posterior of the embryo in the clay. Gently close the clay around the bases of the embryo using the forceps, leaving the top quarter of the embryo, the head, protruding from the depression.
Once all host embryos are secured in their depressions, move to the other side of the plate and begin to insert the donor embryos into their depressions. Repeat the process of securing the embryos in their wells.
5. Performing the Face Transplant Surgery
Cutting knife: As a cutting knife for removal of donor EAD tissue, use an appropriately broken glass capillary needle (see step 2.3 and Figure 1). NOTE: One can directly hold the capillary between fingers (this works well for people with small hands) or one can mount the needle in an insect pin holder. Electrosharpened tungsten needles17 loaded into a pin holder may be used instead of a capillary tube needle. Please see suggested pin holders and tungsten needles in the Table of Specific Reagents and Equipment.
- Under a stereomicroscope, insert the needle into the head of the embryo to the left of the cement gland. The needle should be inserted deeply, so that it passes from outside the embryo, through the head, and into the foregut. To transplant the entire EAD, cuts should extend from the outside of the embryo to the foregut. For ectoderm-only transplants, cuts should be shallower and extend only through the ectoderm.
- Flick the needle from the left to the right side of the head, across the entire width of the cement gland. The flicking is important as the motion gives a clean cut. Refer to Figure 2A for a summary of the technique and Figure 2B for a demonstration of the cuts. The cement gland and eyes are important landmarks for the cuts. The order of the cuts does not affect the outcome and can vary based on the user's preference or handedness.
Place the needle at the start of the previous cut, at the left border of the cement gland, and flick the needle upward until it reaches the bottom of the left eye. This will create a vertical cut from the left border of the cement gland to the bottom of the left eye.
Place the needle at the right border of the cement gland, and flick the needle upward until it reaches the bottom of the right eye. This will create a vertical cut from the right border of the cement gland to the bottom of the right eye.
To fully excise the tissue, flick the needle from the bottom of the left eye to the bottom of the right eye, creating a horizontal cut that will free the tissue. Cuts can extend from the outside of the embryo to the foregut, including both ectoderm and endoderm in the EAD explant. Alternately, shallow cuts can be used for ectoderm-only EAD transplants. Once the EAD tissue is removed, there should be a rectangular hole from the outside of the embryo into the foregut, stretching from the cement gland to just below the eyes (top to bottom). The hole should extend from the inside border of the left eye to the inside border of the right eye (side to side). See Figure 2Bb.
Gently push the excised tissue on the tip of the needle, and lift it through the buffer to the part of the dish containing host embryos. Do not expose the tissue to the surface air or it will become damaged.
Excise the same tissue from the host embryo, as for the donor. Discard the host EAD explant or save it to insert into the donor face, for reciprocal transplants.
Insert the donor explant into the resulting host hole using #5/45 forceps.
Once the donor tissue is correctly positioned and fully inserted, carefully place a glass bridge (see Figure 1 and Figure 2Bc) over the embryo's face to hold the transplant in place. The ends of the bridge should insert into the clay, holding it in place. The bridge should gently apply pressure to the transplanted tissue, such that the transplant lies flush with the host head, without it sticking out of the head or lying deep within the head. The head can be slightly flattened by the cover slip, but be careful not to damage the embryo with too much pressure. Transplants should be performed within 5 minutes. NOTE: An experienced investigator can perform around twenty transplants per experiment, over 2-3 hr. During this period the embryos will progress from stage 20-22. Completing the face transplants at stage 21 or 22 does not affect outcomes. Later transplants (at stages 22-26) can be done but are more difficult as the crest-free EAD midline region is narrowed as cranial neural crest moves into the face. Consistency across transplants is critical.
6. Face Transplant Post-operation Recovery
Healing typically takes 2-3 hr. Leave the embryos at room temperature undisturbed in their clay depressions with the glass bridges holding the donor tissue in place.
Once the transplants have healed, carefully remove the glass bridges, remove the clay from around the base of the embryos using forceps, and use a plastic graduated transfer pipette to gently vacuum the embryos out of their depressions.
Put the embryos into an appropriately labeled Petri dish, half filled with clean 0.1x MBS with gentamycin.
- Grow the embryos at 15 °C or 18 °C for several days until they reach feeding tadpole stage at stage 40, when facial phenotypes can be scored.
- The 0.1x MBS solution with gentamycin should be changed daily, and any dead embryos should be removed promptly to prevent contamination and death of other embryos.
- The mouth opens at stage 40. At stage 40-41, one must check that the transplanted tissue remains healed in place by viewing its fluorescence. Transplants occasionally fall out, so one must ensure that all scored embryos have the donor tissue in place.
Representative Results
Transplanted tissue should be fully inserted into the host head after transplantation as shown in Figure 3A, and have a glass bridge appropriately placed on the embryo's face, as shown in Figure 2Bc. The transplanted donor tissue must be correctly sized for the host opening, for the transplant to be successful. The EAD tissue should not protrude from the head, in any way, as seen in Figures 3B and 3C. Additionally, the face transplant should not be rotated relative to its position in the donor body, as shown in Figure 3D. After several hours, the transplanted tissue and surrounding face should heal, and by the next day, the embryo should appear as the example shown in Figures 4A and 4A'. One can observe, under fluorescence, that the transplant remains in place in Figure 4B. At stage 41, the transplanted control tissue will contribute to the mouth, and will remain florescent green, seen in Figure 4B'. Wild type EAD tissue transplanted into wild type hosts should give rise to normal faces, when compared with unperturbed wild type embryos. However, with LOF or GOF donor tissue, faces should heal and remain in the head, but these may or may not give rise to normal craniofacial structures, as shown in Figure 3 of Dickinson and Sive5.
| Reagent | Ingredients | Instructions |
| 1 M CaCl2 Solution | 111 g of CaCl2 per liter | Autoclave and store in 1 ml aliquots at -20 or 4 °C. |
| 10x Modified Barth's Saline (MBS) Solution | 880 mM NaCl, 51.4 g | Adjust volume up to 1 L with distilled water. Adjust final pH to 7.8 with NaOH and then autoclave. |
| 10 mM KCl, 745.5 mg | ||
| 10 mM MgSO4, 1.2 g | ||
| 50 mM HEPES (pH 7.8), 11.9 g | ||
| 25 mM NaHCO3, 2.1 g | ||
| 1x MBS Solution | Final Concentrations: | Prepare 1x MBS solution by mixing 100 ml of 10x MBS salts solution with 0.7 ml of 1M CaCl2 solution. Adjust volume up to 1 L with distilled water. Dilute this solution to make 0.5x MBS and 0.1x MBS. |
| 88 mM NaCl | ||
| 1 mM KCl | ||
| 0.7 mM CaCl2 | ||
| 1 mM MgSO4 | ||
| 5 mM HEPES (pH 7.8) | ||
| 2.5 mM NaHCO3 |
Table 1. Reagents, ingredients, and instructions.
| Heat | Pull | Vel. | Time |
| 800 | 70 | 40 | 50 |
Table 2. Needle Puller Settings*.
*Needle puller settings vary from machine to machine so each lab will probably need to optimize their own needle puller settings.
 Figure 1. Tools used.A) Shows an intact, unbroken needle and a broken needle after the flexible tip has been removed. B) Depicts two pipette tools with their ends fully sealed and rounded. C) Shows three sample glass bridges. Scale bars = 1,000 mm.
Figure 1. Tools used.A) Shows an intact, unbroken needle and a broken needle after the flexible tip has been removed. B) Depicts two pipette tools with their ends fully sealed and rounded. C) Shows three sample glass bridges. Scale bars = 1,000 mm.
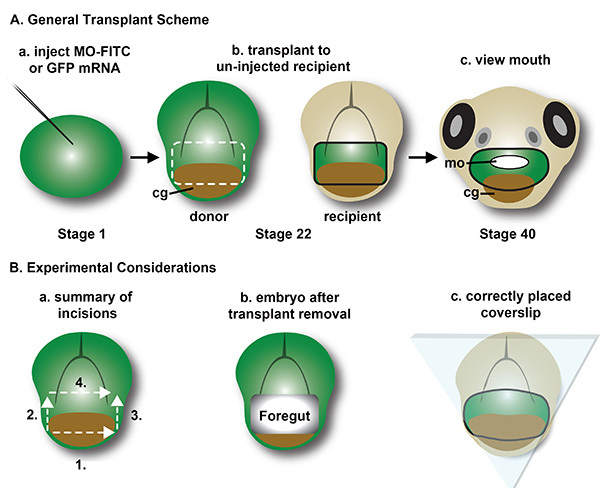 Figure 2. Summary of face transplant method.A) General Transplant Scheme: Schematic of the face transplant method. a. Embryos are injected with a fluorescent agent and an antisense oligonucleotide morpholino at the 1 cell stage. The fluorescent agent can either be GFP mRNA, FITC-tagged morpholino, or fluorescent dextran. b. Presumptive mouth is removed at stage 22 from a host wild type embryo and a donor morphant embryo. The transplanted tissue can also be enlarged to include the presumptive nose, which lies directly above the mouth region. The donor morphant tissue is transplanted to the wild type host, and then is secured in place with a glass bridge. Grey arch: hatching gland. c. Facial development is scored at stages 40-41. B) Experimental Considerations. a. Summary of incisions used to remove the face from a donor embryo. For surgical excision of the face, make incisions 1 through 4 in order. This is the preferred order of cuts, but the order of incisions can vary. b. The resulting donor is shown with the ectoderm and endoderm removed from the face, such that an opening exists from the outside of the embryo to the foregut. c. The diagram shows ideal placement of the glass bridge. The glass contacts both the transplant and host face, pressing the EAD tissue into the head to oppose extrusive forces from wound contraction during healing. Please click here to view a larger version of this figure.
Figure 2. Summary of face transplant method.A) General Transplant Scheme: Schematic of the face transplant method. a. Embryos are injected with a fluorescent agent and an antisense oligonucleotide morpholino at the 1 cell stage. The fluorescent agent can either be GFP mRNA, FITC-tagged morpholino, or fluorescent dextran. b. Presumptive mouth is removed at stage 22 from a host wild type embryo and a donor morphant embryo. The transplanted tissue can also be enlarged to include the presumptive nose, which lies directly above the mouth region. The donor morphant tissue is transplanted to the wild type host, and then is secured in place with a glass bridge. Grey arch: hatching gland. c. Facial development is scored at stages 40-41. B) Experimental Considerations. a. Summary of incisions used to remove the face from a donor embryo. For surgical excision of the face, make incisions 1 through 4 in order. This is the preferred order of cuts, but the order of incisions can vary. b. The resulting donor is shown with the ectoderm and endoderm removed from the face, such that an opening exists from the outside of the embryo to the foregut. c. The diagram shows ideal placement of the glass bridge. The glass contacts both the transplant and host face, pressing the EAD tissue into the head to oppose extrusive forces from wound contraction during healing. Please click here to view a larger version of this figure.
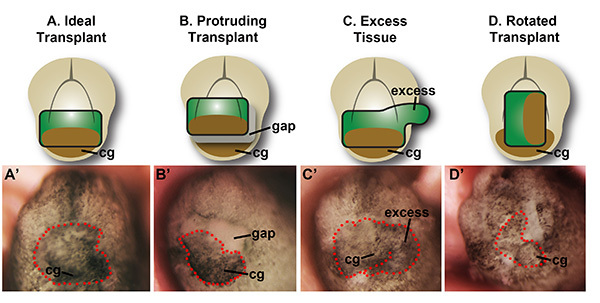 Figure 3. Schematic of embryos shortly after transplantation. Frontal views are shown. The transplanted tissue is outlined in red dots. A and A' depict an ideal outcome at stage 22. The tissue is fully inserted and correctly positioned in the head. B and B' show an incorrect transplant, with the tissue partially inserted into the head. C and C' show an incorrect transplant with most of the tissue inserted into the head, but the excess region is protruding from the healing site. This tissue will necrose, and inhibit healing of the surrounding, properly inserted regions. D and D' show an improper transplant, where the face has been rotated in the host, relative to its position in the donor. The tissue will heal into the head, but the face will not develop normally. Please click here to view a larger version of this figure.
Figure 3. Schematic of embryos shortly after transplantation. Frontal views are shown. The transplanted tissue is outlined in red dots. A and A' depict an ideal outcome at stage 22. The tissue is fully inserted and correctly positioned in the head. B and B' show an incorrect transplant, with the tissue partially inserted into the head. C and C' show an incorrect transplant with most of the tissue inserted into the head, but the excess region is protruding from the healing site. This tissue will necrose, and inhibit healing of the surrounding, properly inserted regions. D and D' show an improper transplant, where the face has been rotated in the host, relative to its position in the donor. The tissue will heal into the head, but the face will not develop normally. Please click here to view a larger version of this figure.
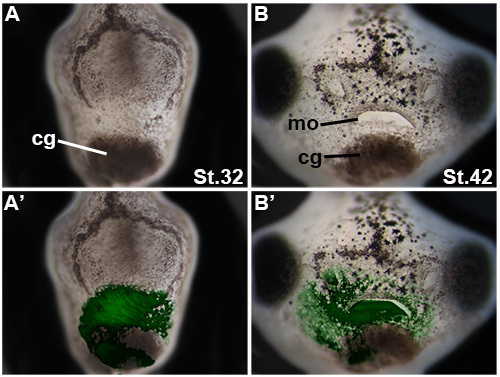 Figure 4. Older embryos with ideal outcomes. Frontal views are shown. cg: cement gland; mo: mouth. A) Displays an embryo one day after transplantation, showing a properly healed transplant at stage 32. A') Shows this same embryo with a fluorescent overlay, confirming that the transplanted tissue remains in place. B) Shows the same embryo at stage 42 which had a control morpholino, GFP+ transplant several days prior. The face has developed properly. B') Shows the same embryo with a fluorescent overlay, confirming that the transplanted tissue remains in the head and has contributed normally to facial structures. Please click here to view a larger version of this figure.
Figure 4. Older embryos with ideal outcomes. Frontal views are shown. cg: cement gland; mo: mouth. A) Displays an embryo one day after transplantation, showing a properly healed transplant at stage 32. A') Shows this same embryo with a fluorescent overlay, confirming that the transplanted tissue remains in place. B) Shows the same embryo at stage 42 which had a control morpholino, GFP+ transplant several days prior. The face has developed properly. B') Shows the same embryo with a fluorescent overlay, confirming that the transplanted tissue remains in the head and has contributed normally to facial structures. Please click here to view a larger version of this figure.
Discussion
Critical Steps and Limitations: The EAD face transplant procedure is time and work intensive. It requires practice, steady hands, and dexterity to perfect. The face transplant protocol relies on the researcher's ability to efficiently remove and transplant tissue. If one takes too long to insert the transplant into the host's face, the host face will begin to contract and heal. Forceps can be used to delicately expand the facial region. However, if significant wound contraction has occurred, the transplant will not heal as well and may need to be reduced in size to fit into the host's facial hole. The transplant's size and shape must roughly match the size and shape of the recipient's facial cavity, to allow successful insertion.
Face transplants are most successful when performed between stages 19-22, with embryos that are matched by stage. Older face transplants, between stages 22-26, are possible, but are more challenging and may disrupt neural crest migration into the sides of the face since the crest-free midline region becomes significantly narrower from stage 22-26.
Both the recipient and donor must be at similar stages for the transplant to work optimally. Ideally, they should be the same stage, and minimally have to be within a few hours of each other. Embryos injected with antisense-RNA morpholino oligonucleotides ("morphants") sometimes develop more slowly than wild type or control morphant embryos, requiring that morphants be grown at a higher temperature to match the wild type embryos' stages.
The donor face tissue should not be rotated in the host body relative to its original position in the donor embryo, otherwise the face will not develop normally. The cement gland does not need to be included in the face transplant for the procedure to work; the face develops normally without it. However, the cement gland is often included in the extirpated facial tissue because it serves as a distinct marker to indicate and position the bottom of the transplant. This marker will help avoid accidently rotating the tissue during the transfer of the face tissue to the recipient. If the transplant has been inserted incorrectly in the host face, the donor tissue can be removed and discarded. One can attempt to insert a new transplant into the same host, if the opening has not begun to constrict. However, if the host opening has noticeably shrunk, then discard the host and begin anew.
It is crucial to place the glass bridge directly on the transplanted face, so that the transplant is held in the host's head during healing. If the transplant is not held in place, the transplant can be extruded from host's face. Transplants that are not fully inserted into the face or that pop out during healing will undergo necrosis. Even in perfect transplants, small amounts of tissue death may occur around the edges of the transplant. Usually this does not cause adverse effects and a completely normal face can still be formed. In successful controls we have not observed any malformation after four weeks of development suggesting that the cartilages formed normally and that rejection of the tissue is rare.
Finally, if a morpholino perturbs a protein required for normal wound healing, this technique may not work, as the transplanted LOF tissue may not be incorporated into the host head.
Possible Modifications and Trouble Shooting: Modifications can be made to the procedure. With practice, the researcher can learn to transplant smaller regions, for example, half of the EAD. Shallow incisions allow ectodermal transplants, leaving the deeper endoderm undisturbed. Other regions of embryonic tissue can be transplanted, using similar approaches.
If the transplanted tissue dies after insertion, there are a couple of possible causes. Transplanted tissue that is not completely inserted into the host or appropriately held in place with a glass bridge, will die and impede proper healing. Make sure to fully insert the transplanted tissue and secure it in place with a glass bridge. If the transplant is derived from a morpholino or RNA injected donor, then the tissue may die due to toxicity from injected agents. Similarly, if the host embryos are morphants or RNA-injected and frequently die, then the amount of morpholino or RNA will need to be reduced. To resolve these issues, titrate the amount of injected RNA or morpholino and determine a safe amount for successful face transplants. Finally, increasing the salt concentration of the MBS solution above 0.1x can also assist healing.
Significance: The face transplant technique described allows analysis of the local requirements and activity of gene products during craniofacial development. This approach can clarify signaling between the developing noncrest, neural crest, and surrounding structures. It allows one to examine LOF or GOF (using any strategy) in all EAD-derived tissues, which is not possible with any single promoter driven construct. Though there is a long history of tissue transplantation in developmental biology this is the first application of transplantation to the study of craniofacial development in frogs and is crucial for mechanistic studies7. Thus the technique can help to unravel the complex mechanisms controlling patterning and formation of the vertebrate face and to clarify causes of craniofacial developmental defects.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We thank Radek Sindelka for his help, and Cas Bresilla for assisting with frog husbandry and embryo preparation. This work was funded by the NIH via the grant R01DE021109 to H.L.S. Laura Jacox was funded by the Herschel Smith Graduate Fellowship at Harvard University and an F30 individual fellowship grant F30DE022989-01 through the NIDCR.
References
- Gorlin RJ, Cohen M, Levin L. Syndromes of the head and neck. UK: Oxford University Press; 1990. [Google Scholar]
- Trainor P. Craniofacial birth defects: The role of neural crest cells in the etiology and pathogenesis of Treacher Collins syndrome and the potential for prevention. Am. J. Med. Gen. A. 2010;152:2984–2994. doi: 10.1002/ajmg.a.33454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dickinson AJ, Sive HL. Development of the primary mouth in Xenopus laevis. Dev. Bio. 2006;295:700–713. doi: 10.1016/j.ydbio.2006.03.054. [DOI] [PubMed] [Google Scholar]
- Dickinson AJ, Sive HL. Positioning the extreme anterior in Xenopus: cement gland, primary mouth and anterior pituitary. Sem. Cell Dev. Bio. 2007;18:525–533. doi: 10.1016/j.semcdb.2007.04.002. [DOI] [PubMed] [Google Scholar]
- Dickinson AJ, Sive HL. The Wnt antagonists Frzb-1 and Crescent locally regulate basement membrane dissolution in the developing primary mouth. Dev. 2009;136:1071–1081. doi: 10.1242/dev.032912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert SF. Developmental Biology. Sunderland, MA: Sinauer Associates, Inc; 2010. [Google Scholar]
- Borchers A, Epperlein HH, Wedlich D. An assay system to study migratory behavior of cranial neural crest cells in Xenopus. Dev. Genes Evol. 2000;210:217–222. doi: 10.1007/s004270050307. [DOI] [PubMed] [Google Scholar]
- Grunz H. Homoiogenetic neural inducing activity of the presumptive neural plate of Xenopus laevis. Dev. Growth Differ. 1990;32:583–589. doi: 10.1111/j.1440-169X.1990.00583.x. [DOI] [PubMed] [Google Scholar]
- Servetnick M, Grainger RM. Changes in neural and lens competence in Xenopus ectoderm: evidence for an autonomous developmental timer. Dev. Bio. 1991;112:177–188. doi: 10.1242/dev.112.1.177. [DOI] [PubMed] [Google Scholar]
- Servetnick M, Grainger RM. Homeogenetic neural induction in Xenopus. Dev. Bio. 1991;147:73–82. doi: 10.1016/s0012-1606(05)80008-9. [DOI] [PubMed] [Google Scholar]
- Couly G, Coltey P, Le Douarin N. The triple origin of skull in higher vertebrates: a study in quail-chick chimeras. Dev. 1993;117:409–429. doi: 10.1242/dev.117.2.409. [DOI] [PubMed] [Google Scholar]
- Couly GF, Le Douarin NM. Mapping of the early neural primordium in quail-chick chimeras : I. Developmental relationships between placodes, facial ectoderm. 1985;110:422–439. doi: 10.1016/0012-1606(85)90101-0. [DOI] [PubMed] [Google Scholar]
- Couly GF, Le Douarin NM. Mapping of the early neural primordium in quail-chick chimeras II. The prosencephalic neural plate and neural folds: Implications for the genesis of cephalic human congenital abnormalities. Dev. Bio. 1987;120:198–214. doi: 10.1016/0012-1606(87)90118-7. [DOI] [PubMed] [Google Scholar]
- Lievre AL, Le Douarin N. The early development of cranial sensory ganglia and the potentialities of their component cells studied in quail-chick chimeras. Dev. Bio. 1982;94:291–310. doi: 10.1016/0012-1606(82)90349-9. [DOI] [PubMed] [Google Scholar]
- Kennedy A, Dickinson A. Median facial clefts in Xenopus laevis: roles of retinoic acid signaling and homeobox genes. Dev. Bio. 2012;365:229–240. doi: 10.1016/j.ydbio.2012.02.033. [DOI] [PubMed] [Google Scholar]
- Trainor P, Tam P. Cranial paraxial mesoderm and neural crest of the mouse embryo- codistribution in the craniofacial mesenchyme but distinct segregation in the branchial arches. Dev. 1995;121:2569–2582. doi: 10.1242/dev.121.8.2569. [DOI] [PubMed] [Google Scholar]
- Sive HL, Grainger RM, Harland RM. Early Development of Xenopus laevis. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2000. [Google Scholar]
- Tandon P, Showell C, Christine K, Conlon F. Morpholino injection in Xenopus. Methods Mol. Biol. 2012;843:29–46. doi: 10.1007/978-1-61779-523-7_4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nieuwkoop PD, Faber J. Normal Table of Xenopus laevis (Daudin) New York: Garland Publishing; 1994. [Google Scholar]