

Abstract
Exposure to heat stress causes physiological and epigenetic changes in plants, which may also be altered in the progeny. We compared the progeny of stressed and control Arabidopsis thaliana wild type and Dicer-like mutant dcl2, dcl3, and dcl4 plants for variations in physiology and molecular profile, including global genome methylation, mRNA levels, and histone modifications in the subset of differentially expressed genes at normal conditions and in response to heat stress. We found that the immediate progeny of heat-stressed plants had fewer, but larger leaves, and tended to bolt earlier. Transposon expression was elevated in the progeny of heat-stressed plants, and heat stress in the same generation tended to decrease global genome methylation. Progeny of stressed plants had increased expression of HSFA2, and reduction in MSH2, ROS1, and several SUVH genes. Gene expression positively correlated with permissive histone marks and negatively correlated with repressive marks. Overall, the progeny of heat stressed plants varied in both their physiology and epigenome and dcl2 and dcl3 mutants were partially deficient for these changes.
Keywords: heat stress, stress tolerance, transgenerational effects, Arabidopsis thaliana, epigenetic response, Dicer-like mutants dcl2, dcl3, and dcl4
Introduction
Plants frequently encounter stresses that negatively impact their growth, survival, and ultimately crop yield, and because they are stationary organisms, their ability to respond to these stresses is particularly crucial.1
Previous research on heat stress has identified changes in both plant physiology and the genome as being modified under stress, with many genes being heat-inducible, allowing for immediate plant response.2 Establishing acquired resistance requires many changes in gene expression and metabolite composition, but ultimately results in plants being able to rapidly modify their homeostasis, acclimating in response to a changing environment.3,4 Many stress-induced events seem to be unintentional, such as activation of transposons. ONSEN and TSI (transcriptionally silent information) are just a few of many transposable elements (TEs), which are one of the most important sources of mutations in genomes.5 An important function of epigenetic regulation is to silence TEs. However, instances of environmental stress may lead to situations where TE expression is increased, and therefore mutation rates are increased.6 For example, heat stress has been previously shown to activate ONSEN.7 By interacting and disrupting other genes, TEs can create new functions that may be beneficial, especially in times of stress that require new coping mechanisms. Often, changes in response to stress such as temperature are reversible, but in certain cases these modifications, including phenotypic and epigenetic, may be passed on to the progeny.8,9
Epigenetic systems have the dynamic ability to rapidly induce changes in response to stress, resulting in modifications that may be short or long-term.10-12 For example, environmental stress has been shown to cause hypo- or hypermethylation, altering the accessibility of chromatin to recombination machinery, and thus modifying variability in the genome at a time when it is most needed.13,14 These changes may be inherited by the progeny of stressed plants, as has been seen in dandelions, tobacco, and Arabidopsis.4,14,15 Recent work on pathogens in Arabidopsis showed that the systemic acquired resistance (SAR) to Pseudomonas syringae pv tomato DC3000 (PstDC3000), could be passed on to one generation of non-stressed progeny, including changes to DNA methylation and chromatin remodelling.16 Likewise, infection of Nicotiana tabacum plants with tobacco mosaic virus resulted in transgenerational changes in genome stability, tolerance to viral and fungal pathogens, metabolite profile, as well as global genome and locus-specific methylation changes.17-19
Small RNAs (sRNAs) are a core component of the epigenetic modifications that occur in response to stress in plants, and there is a dynamic relationship between sRNAs, DNA methylation and changes to histones.20 Dicer-like 2 (DCL2), DCL3, and DCL4 proteins are involved in the biogenesis of various types of siRNAs in Arabidopsis. The important role of small RNAs and dicers in epigenetic stress memory is emphasized by a previous study where parental herbivory primed progeny for enhanced insect resistance but the trait was not present in Arabidopsis dcl2/dcl3/dcl4 mutants.21 Similarly, DCL2 and DCL3 were shown to be required for transgenerational changes in homologous recombination frequency in response to several stresses including heat, cold, drought, and UVC exposure.4
Finally, histone modifications may have an impact on gene expression, both positive and negative. For example, in Arabidopsis acetylation at H3K9 correlates with high gene expression, while dimethylation leads to repression.22 Repressive chromatin mark H3K9me2 positively correlated with DNA methylation and negatively correlated with gene expression in the set of genes in the progeny of plants exposed to salt stress.23 Thus, epigenetic changes including DNA methylation, small RNA expression and histone modifications may all impact gene expression, as well as the epigenetic memory transmitted to the progeny of stressed plants.4,23
Our previous research suggests that transgenerational changes in response to stress may include differential expression of genes involved in DNA repair, regulation of chromatin structure, and DNA methylation.4,23 We demonstrated that the progeny of plants exposed to salt stress showed hypermethylation of promoters and decreased expression of several genes, including Mut-S homolog (MSH6), repressor of silencing 1 (ROS1), and Su(var)3–9 homologs (SUVH) group of genes.23
MSH6 is a DNA mismatch repair protein that, along with MSH2, is involved in the initial recognition of DNA errors.24 Reduction in MSH6 expression in the progeny of stressed plants potentially allows for more point mutations and higher frequency of homologous and homeologous recombination,25 as well as other genomic changes, as has been observed in the progeny of stressed plants.8,26
ROS1 encodes a DEMETER DNA glycosylase that catalyzes the excision of methylated cytosines, thereby triggering increased activity of DNA methyltransferases.27 Decreased ROS1 expression may lead to hypermethylation, including increased methylation of transposons in the progeny of stressed plants, as has been observed previously with salt stress.4
SUVH proteins in Arabidopsis are involved in H3K9 methylation and link it to DNA methylation and transposon repression.28 For example, SUVH2 controls histone methylation, as well as de novo and maintenance DNA methylation through DRM2.29 In addition to histone methylation, SUVH5 and SUVH6 control non-CG methylation, through CMT3, and work with SUVH4 to control transposon expression.30,31
Based on previous work on progeny of salt-stressed plants, we suspected that the progeny of heat-stressed plants would also show morphological and epigenetic variation under both normal and stressed conditions, in comparison to the progeny of control. Indeed, the progeny of heat-stressed plants have shown enhanced stress tolerance when exposed to the parental stress. Furthermore, we found the progeny of heat-stressed plants to have changes in leaf size and bolting as compared with the progeny of control plants. We also found that the expression of ROS1, MSH6, and SUVH genes and histone modifications at the loci harboring these genes were similar to those observed previously for the progeny of salt-stressed plants.23
To examine the potential role of DCL proteins in the transmission of heat stress memory, dcl2, dcl3, and dcl4 mutants were used for comparison to wild-type plants, with the same Columbia background. This work provides evidence of the transmission of both phenotypic and epigenetic modifications to progeny, as a result of heat stress and demonstrates partial requirement for DCL proteins in the process.
Results
Phenotypic transgenerational response to heat stress
In order to determine whether the progeny of stressed plants (S1) are phenotypically different from the progeny of control (C1) plants, heat stress was applied to wild type (line 15D8), dcl2, dcl3, and dcl4 mutants. Measurements were taken a week after the final day of heat stress. Changes to morphology such as seed size and leaf size and number are an indication of the general fitness of the progeny. In particular, leaf size and number is correlated with both biomass and the photosynthesis rate of a plant, providing another indicator of plant fitness and allowing us to examine if parental treatment impacted fitness of progeny under normal and stressed conditions.
Changes in leaf number and leaf size
The number of leaves present on the progeny of stressed plants grown under normal conditions (S1-) was consistently lower than on the progeny of control plants grown under normal conditions (C1-). In addition, the number of leaves on heat stressed plants that were progeny of control plants (C1+) was overall higher than heat stressed progeny of stressed plants (S1+), in particular for dcl plants. The number of leaves in progeny of stressed plants (S1) in comparison to progeny of control plants (C1), decreased for all dcl plants, regardless of F1 treatment (Fig. 1A).

Figure 1. Impact of heat stress on the number and the size of leaves in the progeny of heat stressed and control plants. Plants were stressed at 50 °C for 3 h, on 5 consecutive days. Measurements were taken for leaf number, length, and width. All confidence intervals are a result of bootstrap x 10,000, at 95% confidence (P = 0.05) (A-C) Leaf number (A), length (B), and width (C) in F1 progeny of wild type (15D8) and mutant (dcl2, dcl3, and dcl4) plants, as indicated by Legend, exposed to heat and control plants. “C1” – the progeny of plants grown at normal conditions in F0. “S1” – the progeny of plants exposed to heat in F0. “+” and “-” indicate exposure to stress or growth in uninduced conditions, respectively, each of which had approximately 24 plants. Asterisks (*) indicate a significant difference from wild type, with the same treatment group, or difference between treated and non-treated plants within the same F0 treatment (C1+ vs C1- and S1+ vs S1-) within the same mutant, as calculated using a t test (P ≤ 0.05). (D) Bars represent ratio of change in F1 plants in response to heat stress (S+/S- or C+/C-), and those that overlap with value of 1 indicate no significant change under heat stress. Asterisks (*) are used to indicate mutants that are significantly different from wild-type (15D8) plants with the same treatment. Legend indicates parental treatment.
In general, leaves that were stressed were longer than leaves grown under normal conditions. However, while heat stress increased leaf length for 15D8, parental heat stress also had an impact, as the progeny of stressed plants (S1) had longer leaves than the progeny of control (C1) 15D8 plants under both stressed and normal conditions. S1 dcl4 plants leaves were significantly shorter than those belonging to 15D8, regardless treatment, while C1 dcl4 were quite similar (Fig. 1B).
Leaf width tended to increase with F1 heat stress. However, the difference between the progeny of stressed and the progeny of control plants was not substantial, except for a decrease in leaf width for S1+ dcl3 and dcl4 progeny, in comparison to C1+. Also, while the leaf width of C1- dcl2 was significantly higher than C1- 15D8 plants, it was still significantly lower in the progeny of stressed plants grown under normal conditions (S1-). Leaf width decreased in S1- dcl3 plants, but increased in S1- 15D8 and S1- dcl4 plants (Fig. 1C).
Heat stress resulted in significantly fewer leaves in the progeny of stressed dcl2 and dcl3 plants, in contrast to 15D8 and dcl4. Leaf length was not significantly influenced by parental treatment. However, leaf width showed a significant difference between all other plant types and dcl3, which was wider under stress. In nearly all cases, leaf number and size increased in response to heat stress resulting in a ratio of stressed to non-stressed greater than 1 (Fig. 1D).
Changes in seed length
There was significant variation in the size of seeds produced by both progeny of control plants and progeny of stressed plants (Fig. 2A). dcl3 seeds were the largest in the group of S1 plants, in contrast to C1 seeds, where dcl2 was the largest. dcl2 seeds produced under stress were significantly smaller than those produced by control plants. In particular, seeds of heat stressed dcl2 plants were significantly smaller than those produced by any other mutant type. dcl4 seed size decreased with heat, where they were significantly different than 15D8. In response to heat stress, 15D8 and dcl3 plants produced significantly larger seeds than dcl2 and dcl4. 15D8 seeds increased in size in response to heat stress, while dcl2 and dcl4 mutants decreased, and dcl3 did not significantly change (Fig. 2B).
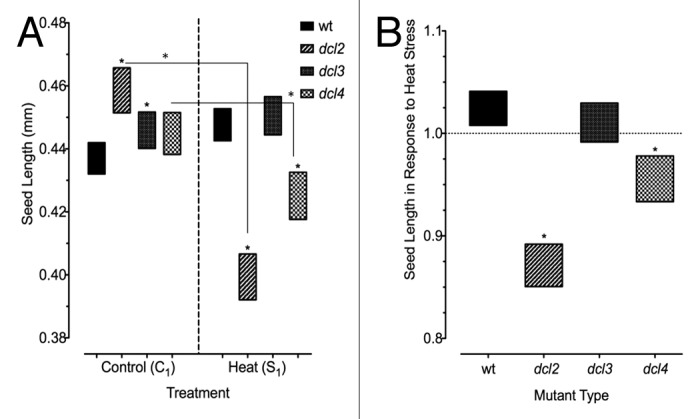
Figure 2. Size of seeds produced by heat stressed and control plants. Approximately 100–200 seeds per sample group. (A) Seed size in F1 progeny of wild type (15D8) and mutant (dcl2, dcl3, and dcl4) plants, as indicated by Legend, exposed to heat and control plants. “C1” – the progeny of plants grown at normal conditions in F0. “S1” – the progeny of plants exposed to heat in F0. Asterisks (*) indicate a significant difference from wild type, with the same treatment group, or difference between different treatments (C1 vs S1) within the same mutant, as calculated using a t test (P ≤ 0.05). (B) Bars represent ratio of change in seeds produced by plants that had been heat stressed (S1/C1), and those that overlap with value of 1 indicate no significant difference in seed size in response to heat stress. Asterisks (*) are used to indicate mutants that are significantly different from wild-type (15D8) plants with the same treatment, as calculated using a t test (P ≤ 0.05). Legend indicates parental treatment.
Analysis of bolting rates
Parental treatment impacted the bolting rate of plants under normal growth conditions. Specifically, the progeny of plants that were heat stressed, bolted faster under normal conditions (S1-) than the progeny of control (C1-), with the exception of dcl4, which bolted later (Fig. 3A). However, regardless of parental treatment, bolting percentages increased in response to heat stress (+), with the exception of dcl2 S1 (Fig. 3B).

Figure 3. Percentage of F1 plants that had bolted at approximately 4 wk of age. Approximately 24 plants were included in each treatment group. (A) Comparison of bolting in F1 plants when grown at normal conditions. (B) Comparison of bolting in F1 plants when grown under either normal (-) or stressed (+) conditions. Each treatment group is labeled according to treatment and mutant type, on the horizontal axis.
Epigenetic transgenerational response to heat stress
Changes in expression of transposons
ONSEN transposon showed significantly higher expression in the progeny of heat stressed plants (S1) than progeny of control plants (C1), regardless of F1 treatment. In most cases, F1 heat stress did not significantly alter expression, especially in the progeny of stressed plants (S1+ in comparison to S1-). There was an increase in ONSEN expression in heat stressed C1 dcl4 plants (Fig. 4A). Progeny of heat stressed plants also tended to have higher TSI expression than progeny of control plants. While stressing C1 15D8 plants increased expression, in most other cases, F1 heat stress did not impact TSI expression (Fig. 4B).
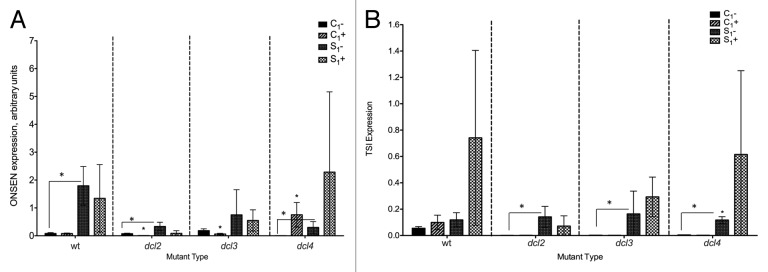
Figure 4. Expression of ONSEN (A) and TSI (B) transposons in F1 progeny of wild type (15D8) and mutant (dcl2, dcl3, and dcl4) plants exposed to heat and control plants. Y-axis shows arbitrary units of gene expression. “C1” – the progeny of plants grown at normal conditions in F0. “S1” – the progeny of plants exposed to heat in F0. “+” and “-” indicate exposure to stress or growth in uninduced conditions, respectively. Bars show standard deviation calculated from 3 technical repeats. Asterisks (*) indicate a significant difference between control (-) and stress (+) with the same parental treatment or between different treatments (C1 vs S1), as calculated using a t test (P ≤ 0.05).
The progeny of heat stressed plants grown under normal conditions had higher TSI and ONSEN expression than the progeny of control plants. When plants were heat stressed, the progeny of heat stressed plants (S1+) had a higher level of TSI and ONSEN expression than the progeny of controls (C1+), though the difference was not significant in most cases.
Changes in global genome methylation
In all cases, methylation decreased in stressed plants of the same generation, for example C1+ in comparison to C1-, and S1+ in comparison to S1-. The only possible exception to this was dcl3 C1+ in comparison to C1-. This difference was most substantial in S1+ for both dcl2 and dcl3 plants, while 15D8 maintained a similar decrease in methylation under stress, regardless of parental treatment (Fig. 5).
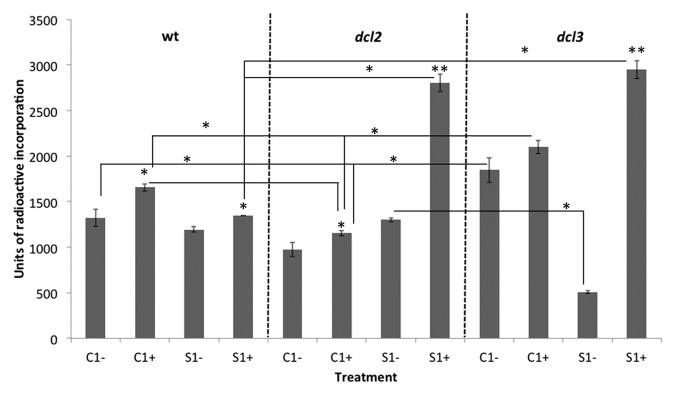
Figure 5. Analysis of global genome methylation in the F1 progeny of wild type (15D8) and mutant (dcl2 and dcl3) plants exposed to heat and control plants. Y-axis shows units of radioactive incorporation (dpm/µg). Higher incorporation indicates a lower degree of methylation. “C1” – the progeny of plants grown at normal conditions in F0. “S1” – the progeny of plants exposed to heat in F0. “+” and “-” indicate exposure to stress or growth in uninduced conditions, respectively. Error bars show standard deviation calculated from 2 technical repeats. Asterisks (*, **) indicate a significant difference between control (-) and stress (+) with the same parental treatment, between different treatments (C1 vs S1) or between different mutants, as calculated using a t test (P ≤ 0.05 or P ≤ 0.01, for 1 or 2 asterisks, respectively).
Interestingly, dcl3 showed the lowest level of methylation after 2 generations of growth in normal conditions (C1-), while dcl2 showed the highest. dcl3 showed an increase in methylation in the progeny of stressed plants, grown under normal conditions (S1-), in comparison to C1-, while dcl2 methylation decreased. The highest level of methylation for plants grown in normal conditions was seen in S1- dcl3 and the difference between S1- and C1- dcl3 plants was the most prominent as compared with 15D8 and dcl2 plants (Fig. 5). Results of cytosine extension assay were confirmed via HPLC (Fig. S1).
Changes in mRNA expression
HSFA2 is the dominant heat stress transcription factor in Arabidopsis, due to its high activator potential for transcription of genes encoding heat stress proteins and its continued accumulation during repeated cycles of heat stress and recovery, regardless of tissue.32,33 The progeny of heat-stressed wild type plants had significantly higher HSFA2 expression than the progeny of control plants. The progeny of stressed mutants did not have higher HSFA2 expression unless they were stressed. Wild type and dcl3 but not dcl2 plants had higher HSFA2 expression in the stressed progeny of control (C1+) plants than those plants grown under normal conditions (C1-) (Fig. 6A).
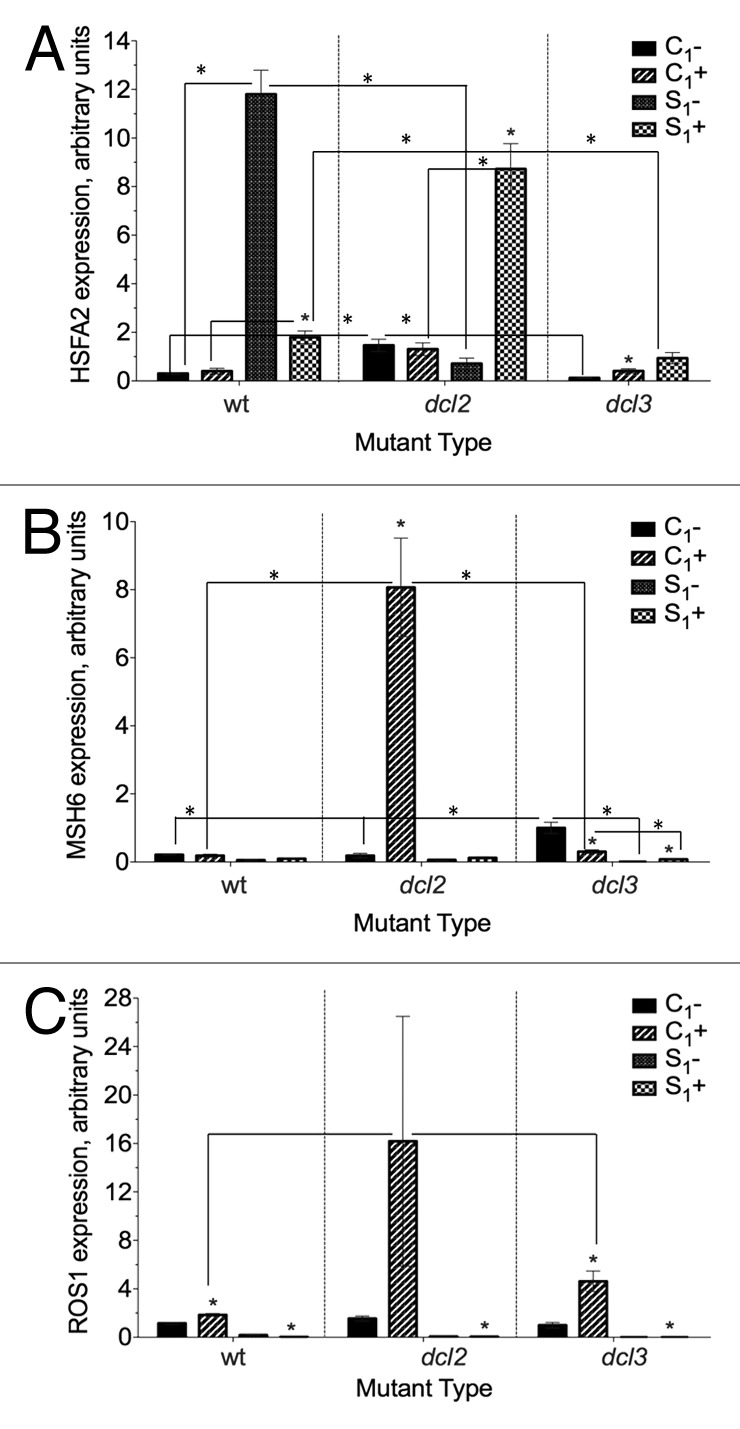
Figure 6. Expression of (A) HSFA2 (B) MSH6 (C) ROS1 in F1 progeny of wild type (15D8) and mutant (dcl2 and dcl3) plants exposed to heat and control plants. Y-axis shows arbitrary units of gene expression, standardized to tubulin. “C1” – the progeny of plants grown at normal conditions in F0. “S1” – the progeny of plants exposed to heat in F0. “+” and “-” indicate exposure to stress or growth in uninduced conditions, respectively. Bars show standard error of the mean (SEM) calculated from 3 technical repeats. Asterisks (*) indicate a significant difference between control (-) and stress (+) with the same parental treatment, between different treatments (C1 vs S1) or between different mutants, as calculated using a t test (P ≤ 0.05).
The offspring of heat-stressed plants had a significantly lower level of MSH6 expression than the progeny of control plants in both dcl2 and dcl3 mutants (Fig. 6B). Such differences were not significant for the wild type. Stressing C1 dcl2 plants resulted in significant increase in MSH6 expression (Fig. 6B). Similarly, expression of ROS1 was significantly lower in the progeny of stressed plants than the progeny of controls. Stressed C1 plants (+) had increased ROS1 expression in comparison to C1 plants grown under normal conditions. In contrast, stressed S1 plants had significantly lower ROS1 expression than S1- plants (Fig. 6C). There was no substantial difference between wild type and mutant plants, except that there was somewhat stronger induction of ROS1 expression in C1+ group.
The progeny of control plants had significantly higher SUVH2 expression than the progeny of stressed plants. However, stressing the progeny of control plants (C1+) significantly increased SUVH2 expression. In contrast, stressing the offspring of stressed plants (S1+) significantly decreased expression compared with normal conditions (S1-) for 15D8 and dcl3 (Fig. 7A).

Figure 7. Expression of (A) SUVH2 (B) SUVH5 (C) SUVH6 (D) SUVH8 in F1 progeny of wild type (15D8) and mutant (dcl2 and dcl3) plants exposed to heat and control plants. Y-axis shows arbitrary units of gene expression, standardized to tubulin. “C1” – the progeny of plants grown at normal conditions in F0. “S1” – the progeny of plants exposed to heat in F0. “+” and “-” indicate exposure to stress or growth in uninduced conditions, respectively. Bars show standard error of the mean (SEM) calculated from 3 technical repeats. Asterisks (*) indicate a significant difference between control (-) and stress (+) with the same parental treatment, between different treatments (C1 vs S1) or between different mutants, as calculated using a t test (P ≤ 0.05).
Among plants grown in normal conditions, SUVH5 expression was significantly lower in progeny of stressed plants (S1-), than progeny of control plants (C1-) for 15D8 and dcl2. In most cases, stressing plants with the same parental treatment increased SUVH5 expression, in comparison to those grown under normal conditions. The only exception was C1 dcl2 plants (Fig. 7B). Likewise, the progeny of control plants had significantly higher SUVH6 expression than the progeny of stressed plants. Similar to SUVH5, stressed progeny of control plants (C1+) had significantly higher levels of SUVH6 expression than the control progeny of control plants (C1-), but stressed progeny of stressed plants (C1+) had significantly lower SUVH6 for 15D8 and dcl3 (Fig. 7C).
The changes in SUVH8 expression among differing treatment groups were not consistent. In both dcl2 and dcl3 plants, expression was lowest for S1-, but for 15D8 SUVH8 expression was highest for the same treatment group. Expression of SUVH8 significantly increased in C1+ plants, compared with C1- plants for dcl2 and dcl3 (Fig. 7D).
Changes in histone modifications
Permissive acH3K9 mark showed significant enrichment in the progeny of 15D8 and dcl2 heat stressed plants, for heat-shock transcription factor HSFA2 coding region, whereas repressive me2H3K9 marks only showed a tendency to such increase. The progeny of heat stressed dcl3 plants had significantly lower levels of me2H3K9 in HSFA2 coding region, as compared with wt plants (Fig. 8A).

Figure 8. Analysis of H3K9me2 and H3K9ac histone modifications in HSFA2, SUVH2, SUVH5 genes. PCR analysis was done in the progeny of heat-stressed (S1) and control (C1) plants using primers specific to the gene body region of HSFA2 (A), promoter region of SUVH2 (B), gene body region of SUVH2 (C), and gene body region of SUVH5 (D). The y-axis shows the levels of H3K9me2/H3K9ac expression in average arbitrary units (calculated from 2 technical repeats). The x-axis indicates the mutant type. The asterisks (*) denotes a significant differences between the progeny of control and the progeny of stressed or between mutants for the same treatment, as determined with a t test (P ≤ 0.05).
The level of acH3K9 significantly decreased in the promoter region of SUVH2 in the progeny of heat stressed 15D8 plants, compared with the progeny of control plants. The changes to acH3K9 and me2H3K9 were similar among 15D8 and dcl2, with the opposite trend occurring for dcl3 (Fig. 8B).
The coding region of SUVH2 did not show any significant changes in the above mentioned chromatin marks, however changes in dcl3 again had an opposite trend to 15D8 and dcl2 (Fig. 8C).
In SUVH5 gene, the chromatin marks were not significantly changed in the progeny of stressed plants, in comparison to progeny of control in 15D8 or mutants tested (Fig. 8D).
Discussion
Changes to leaf phenotype
Changes to leaf number and size were observed in the progeny of heat stressed plants, compared with the progeny of controls. The variation was observed when progeny were grown under stress, or normal conditions, with several differences occurring among wild-type and plants deficient in dcl2, dcl3, or dcl4.
Heat stress can shorten the life cycle of a plant, by increasing the rate of plant development, and thereby decreasing the growth period as well as resulting seed and plant yield.34 It can also result in a significant increase in number of leaves.35,36 Interestingly, the progeny of heat-stressed plants (S1) had fewer leaves than the progeny of control plants (C1), under both normal and heat-stressed conditions. It is possible that the progeny of stressed plants received a form of epigenetic memory that allowed them to retard the growth process, making them better suited to conditions of heat stress, in which development is sped up. Recent report by Suter and Widmer (2013) also demonstrates a decrease in the leaf number in the progeny of plants exposed to heat for 3 consecutive generations.37
In contrast to leaf number, not only did leaf length generally increase in heat stressed plants, but the progeny of stressed wild-type plants also had longer leaves than the progeny of control, even under normal growth conditions. In conjunction with the aforementioned observation of leaf number, it appears that the progeny of stressed plants are able to increase leaf size, and therefore biomass, without accelerating the developmental process via the growth of new leaves.
Leaf width also tended to increase with heat stress but the variation among the progeny of stressed plants and the progeny of controls under heat stress was not significant in most cases. However, the progeny of stressed 15D8 and dcl4 plants had wider leaves under normal growth conditions than the progeny of non-stressed plants, indicating that the stress memory may in fact be more evident under conditions of normal growth, than those of stress.
Previous work has shown that heat can directly impact the appearance of a leaf, increasing both leaf elongation and leaf width.38 It is possible that some of these characteristics are passed onto progeny of stressed plants, resulting in larger leaves not only in response to high temperature, but also under normal growth conditions as well. In addition, work by Bos et al. (2000)38 showed that the leaf width is closely associated with specific leaf weight. It is possible that by increasing leaf width, the progeny of heat stressed plants increased plant biomass, providing a positive buffer against the decrease in biomass that is generally associated with heat stress and accelerated development.34
DCL2 and DCL3, proteins involved in RNA-directed DNA methylation (RdDM), might play an important role in epigenetic modifications that pass on stress memory to progeny.39 In response to heat stress, dcl2 and dcl3 plants often had a different response than 15D8 and dcl4. For example, there were significantly fewer leaves in the progeny of stressed dcl2 and dcl3 than 15D8 plants, in response to heat stress.
Previous work has found that dcl2 and dcl3 were impaired in stress-induced homologous recombination frequency and DNA methylation.4 For example, dcl3 was impaired in recombination increase in response to UV-C and cold. In addition, heat and UV-C lead to genome-wide hypermethylation in progeny of stressed wild-type plants. However, the progeny of heat and UV-C stressed dcl2 and dcl3 plants did not show significant changes in methylation in comparison to the progeny of control.4 An impaired ability to transmit epigenetic changes such as methylation in response to stress may correspond to the lack of transmission of morphological changes, as seen in our study.
Changes to seed phenotype
In our study, wild-type (15D8) seed length increased in response to heat stress (S1 seeds) but dcl2 and dcl4 were both significantly smaller than wild-type, as well as showing a significant decrease in seed size (in comparison to the C1 seeds produced by control plants). dcl3 seeds did not undergo a significant change in size in response to heat stress, nor did they vary significantly from 15D8.
Previous work has shown a significant decrease in seed size, taken as a measurement of seed weight, in response to heat stress.35,40 Heat stress decreased seed size and caused lower yields.41 The reduction in seed size caused by heat stress has also been correlated with decreased germination.42 It is possible that 15D8 plants were able to shift the balance toward larger seeds, in response to heat stress. This increase in seed size could potentially result in higher seedling survival of progeny, making them better adapted to withstand stress, a trait dcl plants may not have been able to transmit as effectively.
Changes to bolting time
Heat increased bolting time in all cases, regardless of parental treatment. However, under normal conditions the progeny of heat-stressed plants bolted earlier than the progeny of control plants, with the exception of dcl4.
A recent study in maize showed that heat stress decreased plant height by up to 30%.43 Heat stressed plants not only decrease height and biomass, but often have fewer flowers, and as a result, fewer seeds and fruit-set. Reduced fruit-set is often a result of fewer pollen grains and decreased pollen viability.44,45 As described previously, heat often increases plant growth development, and in this case it likely promotes bolting at an earlier age.
Earlier bolting in the progeny of heat-stressed plants, in comparison to the progeny of control, under normal growth conditions may be caused by the same phenomenon that triggered bolting earlier in heat-stressed plants. The transgenerational inheritance mechanism is unknown but it is possible that changes in the production of plant hormones that regulate the growth cycle may trigger the developmental change earlier, even in unstressed offspring.33 The work of Balasubramanian et al. (2006) showed induction of early flowering in plants exposed to 27 °C.46 Although the authors did not test bolting, it is likely that earlier bolting has also occurred in heat stressed plants. Similarly, more recent work showed that the progeny of plants exposed to heat for 3 generations also exhibited early flowering phenotype.37 Also, Brachi et al. (2012), found selection for early bolting in the progeny of water stressed plants, similar to our results for heat stress.9
Changes in transposon expression
Our analysis showed that expression of ONSEN and TSI had increased in the progeny of stressed plants, indicating that 5 days of 3 hours of heat stress at 50 °C was sufficient for the formation of an epigenetic memory of TE activation strong enough to persist into progeny. The lack of variation among single dcl mutants and wild-type expression is supported by previous work indicating that the Dicer family is redundant for ONSEN transposition.47
In recent work, heat-induced ONSEN expression was significantly higher in dcl3 plants than wild-type indicating that DCL3 may be partially responsible for restricting the accumulation of ONSEN following heat-stress. However, there were no new ONSEN insertions in the progeny of heat stressed dcl3 plants, indicating that DCL3 was not required for the transmission of transgenerational transposition of ONSEN. The progeny of heat-stressed wild-type plants had no transgenerational transposition of ONSEN.7 Our work showed the increased ONSEN expression in the progeny of heat-stressed plants, regardless of mutant type. The same may be true of TSI, which also increased expression in the progeny of heat-stressed plants, as well as other TEs. TE proliferation in somatic tissue has been associated with biotic and abiotic stress,48-50 so it is interesting that this change can be observed in the progeny.
Transposon expression is also activated in response to various other stresses. For example, Tto1 expression was shown to be upregulated by tissue culture stress in Arabidopsis,51 wounding in tobacco52 and treatment with methyl jasmonate in Arabidopsis.53
When exposed to stress, the progeny of stressed and control plants did not usually vary in TE expression levels. This indicates that although the natural level of transposon expression tended to be higher in the progeny of stressed plants, changes in TE expression caused by heat stress in S1 and C1 plants were not significant. Such a response provides evidence that the expression levels enabled by parental stress are not significantly different than those which would naturally occur in response to the stress.
Changes in global genome methylation
In F1 plants that were heat stressed, methylation decreased in comparison to plants with the same parental treatment when grown under normal conditions. Previous work showed that abiotic stress induced the accumulation of transcripts associated with active demethylation in plants, for example, exposure to water stress in pea and heavy metal stress in hemp and clover.54,55 Biotic stress, in the form of viral infection also caused hypomethylation of somatic tissue in Arabidopsis plants.56
The most substantial decrease in methylation was seen in the stressed progeny of stressed plants for dcl2 and dcl3, which resulted in the lowest overall levels of methylation. However, the response to heat stress did not significantly change with differing parental treatments for 15D8, indicating the inheritance of epigenetic stress memory in wild-type may vary from dcl plants due potentially to the role that DCL proteins play in DNA methylation.
Changes in mRNA expression
Progeny of heat-stressed plants had higher levels of heat shock transcription factor A2 (HSFA2) expression than progeny of control plants. This is interesting because HSFA2 expression is triggered by heat stress.32 These results indicate that a mechanism for transgenerational inheritance of increased HSFA2 expression in the progeny of heat-stressed plants exists.
In contrast to HSFA2, all other genes decreased expression in S1 plants, compared with C1, within the same mutant type. Previous work has shown that the progeny of salt-stressed plants had hypermethylation of many genes involved in the regulation of chromatin structure paralleled by lower expression levels, results consistent with the decrease in expression seen in our study.23 These results indicate that some responses to stress are common among the progeny of both salt and heat stressed plants.
Decreased ROS1 expression may lead to hypermethylation, including increased methylation of transposons in the progeny of stressed plants, as has been observed previously with salt stress.4 Hypermethylation should prevent expression from transposon loci. However, in our study, the progeny of stressed plants actually had higher levels of ONSEN and TSI expression, suggesting that hypermethylation may not occur in the entire genome.
SUVH2, SUVH5, SUVH6, and SUVH8 previously showed decreased mRNA levels in the progeny of salt-stressed Arabidopsis plants.23 These genes were also downregulated in the progeny of heat-stressed plants in this study. Potentially, decreased expression of these homologs may protect the progeny of stressed plants from genome-wide hypermethylation and contribute to TE activation because of their activity as histone methyltransferases and their link to DNA methylation.28
Changes in histone modifications
ChIP analysis was used to determine if changes in mRNA expression correlated with histone modifications. However, because of similar changes in expression among SUVH genes, ROS1 and MSH6, only a subset of the genes examined via mRNA expression were once again studied. SUVH2 and SUVH5 were both studied to ensure the ChIP trends were also consistent, and HSAF2 was examined due to its differing mRNA expression results.
The level of acH3K9 and me2H3K9 increased in the progeny of heat stressed 15D8 and dcl2 plants for HSFA2. In contrast, acH3K9 increased but me2H3K9 decreased in dcl3. These results are supported by mRNA data that showed a higher level of HSFA2 expression in S1 than C1 plants.
Changes in the chromatin marks in SUVH2 promoter regions of the wild type plants were similar to those found before in the progeny of salt stressed plants.23 In contrast changes at the coding region were different in the progeny of heat stressed plants as compared with salt stressed, suggesting that not all genes and gene regions have similar histone modifications associated with them in plants exposed to different stresses.23
In conclusion, we have shown that heat stress impacts plant epigenome and physiology, and these changes may be transmitted to progeny. Transgenerational inheritance appears to be at least partially reliant on DCL2 and DCL3, though some redundancy may exist. It will be interesting to determine how many generations these transgenerational changes are maintained in the absence of stress, and if similar epigenetic modifications are made in response to other environmental stresses.
Materials and Methods
Plant growth conditions
Arabidopsis thaliana lines 15d8, dcl2, dcl3, and dcl4 mutants (Columbia ecotype) were planted in soil with a ratio of 7:1, all-purpose potting soil to vermiculite. Line 15d8 plants carry in the genome luciferase-based homologous recombination substrate. All 3 mutants were previously crossed to 15d8 plants and plants homozygous for recombination substrate and for the deficiency for a respective DCL mutation were selected and used for the experiments.4 In the current work, the 3rd generation of these plants was used.
Soil was prepared using water containing a generic fertilizer to field capacity and then stratified for 96 h at 4 °C. Plants were then grown in growth chambers (BioChambers) at long day conditions of 16 h light (22 °C) and 8 h dark (18 °C), at high light conditions (328 µmol(photon)m−2s−1), under a constant humidity of 65%.
At approximately 5 d post germination (dpg) plants were transplanted into pots containing soil and fertilizer, for a total of 12 plants per pot, and 2 pots per sample group, resulting in approximately 24 plants per treatment group, including control and stressed group. Heat stress was applied at one-week dpg for 5 consecutive days, 3 hours at 50 °C. Seeds produced by control and stressed F0 plants were collected and combined, and named C1 and S1, respectively. The same growth conditions were used for F1 plants; C1 and S1 seeds were germinated and were heat-stressed or not, resulting in stressed progeny of stressed plants (S1+), stressed progeny of control plants (C1+), progeny of stressed plants grown under normal conditions (S1-) and progeny of control plants grown under normal conditions (C1-).
Seven days post-stress, morphological measurements were taken and tissue was collected. Tissue was taken from rosette leaves of all treatment groups, and flash-frozen using liquid nitrogen and then stored at -80 °C. Four samples of approximately 100 mg of tissue were collected from each treatment group. Seeds were collected from those plants from which tissue samples were not taken (~20 plants per group).
Morphological measurements in C1 and S1 plants
The number of true leaves on each plant was counted and measurements of the length and width were taken of the third emerging leaf (~24 plants). At approximately 4 weeks post germination bolting was assessed on each plant and expressed as a percentage.
Seeds were collected from the remaining plants (~20) from each treatment group and combined. Seeds were photographed under the microscope, and the seed length of approximately 100–200 seeds per treatment group was measured using Image J.
Real-Time qPCR analysis
Approximately 100 mg of plant tissue was ground in liquid nitrogen and transferred to a chilled 1.5-mL Eppendorf tube, and 800 μL of TRIzol reagent (Invitrogen) was added. The remainder of the extraction was performed according the manufacturer’s protocol. The quantity and quality of mRNA was measured in RNase-free double distilled water using a spectrophotometer. cDNA were prepared using the iScript Select cDNA synthesis kit (Bio-Rad) according to manufacturer’s protocol.
The transcriptional activity of ONSEN and TSI was determined by amplifying the genes from cDNA using qPCR with SsoFast EvaGreen Supermix (Bio-Rad, cat. #1725200). The transcriptional activity of several histone modifier genes (SUVH2, SUVH5, SUVH6, and SUVH8) as well as a demethylase ROS1, mismatch repair gene MSH6, and heat shock transcription factor HSFA2, were also measured, and Tubulin encoding gene was used as a control. Sequence of primers used for RT PCR is shown in Supporting information, Table S1, and were designed using Beacon Designer7 program.
Amplification was done under the following conditions: 1) 98 °C for 2 min for one cycle; 98 °C for 5 s, 48 °C for 5 s, for 40 cycles, and 2) melt-curve analysis - 65 °C to 95 °C for 5 s, with a 0.5 °C increment. The average of 3 reactions was obtained, and the normalized expression was calculated using 2-ΔΔCT method.
ChIP-qPCR analysis
ChIP analysis of histone modifications in the progeny of 15D8, dcl2 and dcl3 Arabidopsis plants grown under normal and heat-stressed conditions, was done according to previously described protocol57 with minor modifications previously described before.23 Approximately 6 plants were used per treatment group. For immunoprecipitation, ChIP grade antibodies against acetyl H3K9 (Millipore, cat. #- 17–658) and methyl 2 H3K9 (Abcam, cat. #- ab1220) were used.
A no-antibody negative control was used, and all precipitated gDNA fragments were amplified using qPCR with SsoFast EvaGreen Supermix (Bio-Rad, cat. #1725200) using either promoter- or gene- specific primers under the following conditions: (1) 98 °C for 2 min for one cycle; 98 °C for 5 s, 48 °C for 5 s, for 40 cycles, and (2) melt-curve analysis- 65 °C to 95 °C for 5 s, with a 0.5 °C increment. Primers used included gene body for SUVH2, SUVH5 and HSFA2 as well as SUVH2 promoter region (Table S1). The normalization was done against Tubulin. The average of 2 reactions per treatment group was obtained, and the normalized expression was calculated using 2-ΔΔCT method.
Cytosine extension assay
Approximately 100 mg of tissue, taken from rosette leaves of 15D8, dcl2 and dcl3 plants, per sample group was used for the extraction of genomic DNA. Due to the similarity between 15D8 and dcl4 in the morphological portion of this study, dcl4 plants were not examined. Genomic DNA quality and quantity were determined using a spectrophotomer. Aliquots of 1.0 µg genomic DNA per 1 µL nuclease-free water were prepared.
Global genome methylation changes were detected using the cytosine-extension assay, following the protocol as described by.58 One aliquot of 1.0 µg genomic DNA was incubated with a methylation-sensitive endonuclease, while a second aliquot was incubated without the restriction enzyme and served as a background control.
The incorporation of the radiosensitive label from samples incubated with the endonuclease was measured using PerkinElmer liquid scintillation analyzer Tri-carb 2910 TR (RIT). Incorporation was directly dependent on the completeness of the DNA digestion, and therefore, a higher level of methylation decreased the digestion rate and resulted in a lower incorporation rate. The readings taken from samples incubated without the restriction enzyme indicated the background radiolabel incorporation (RIB). The actual radiolabel incorporation (RIA) was calculated using the formula RIA = RIT – RIB. This was further confirmed by HPLC detection following Rozhon et al. (2008).59
Statistical treatment of the data
Statistical analysis of morphological data including leaf number, length, width, and seed length, were performed using R software. A 95% confidence interval was calculated for each measurement per treatment group, using a sample size of approximately 24 plants, using a bootstrap x 10,000. Confidence intervals were compared with bootstrap x 10,000 to determine if 2 intervals were significantly different from each other.
The response of plants under stress was determined by dividing the measurement of stressed plants by the measurement under normal conditions, in order to determine if a significant change occurred. If the resulting value overlapped with 1 at 95% confidence (P = 0.05) no significant change was observed. All ratios were calculated using bootstrap x 10,000. Comparisons were done between wild-type and dcl mutants in order to determine if the difference between the 2 responses was significant at a confidence interval of 95%. These results were graphed using Prism (Graphpad) software.
Gene expression results for transposons and genes involved in epigenetic regulation as well as ChIP results were graphed and analyzed using Prism (Graphpad) software. Standard deviations and standard errors of the mean were given from qPCR (BioRad) results and a t test was performed to determine differences, where significance was given to result with a P ≤ 0.05.
Supplementary Material
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We appreciate the help of John Sheriff in statistical analysis, Andriy Bilichak in ChIP work, and Rocio Rodriguezjuares in HPLC detection of global DNA methylation. Financial support was provided by Alberta Agricultural Research Institute and National Science and Engineering Research Council of Canada Discover grant to Igor Kovalchuk and NSERC MSc scholarship to Zoe Migicovsky.
Supplemental Materials
Supplemental materials may be found here: www.landesbioscience.com/journals/psb/article/27971
Glossary
Abbreviations:
- ChiP
chromatin immunoprecipitation
- DCL
Dicer-like
- Dpg
days post germination
- gDNA
genomic DNA
- “C1
” progeny of control plants
- “C1-“
progeny of control plants grown under normal conditions
- “C1+
” progeny of control plants grown under stress conditions
- “S1
” progeny of stress plants
- “S1-
” progeny of stressed plants grown under normal conditions
- “S1+
” progeny of stressed plants grown under stress conditions
- qPCR
quantitative PCR
- RdDM
RNA-directed DNA methylation
- SAR
systemic acquired resistance
- sRNAs
small RNAs
- TE
transposable elements
References
- 1.Mittler R, Blumwald E. Genetic engineering for modern agriculture: challenges and perspectives. Annu Rev Plant Biol. 2010;61:443–62. doi: 10.1146/annurev-arplant-042809-112116. [DOI] [PubMed] [Google Scholar]
- 2.Thomashow MF. PLANT COLD ACCLIMATION: Freezing Tolerance Genes and Regulatory Mechanisms. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:571–99. doi: 10.1146/annurev.arplant.50.1.571. [DOI] [PubMed] [Google Scholar]
- 3.Vinocur B, Altman A. Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Curr Opin Biotechnol. 2005;16:123–32. doi: 10.1016/j.copbio.2005.02.001. [DOI] [PubMed] [Google Scholar]
- 4.Boyko A, Blevins T, Yao Y, Golubov A, Bilichak A, Ilnytskyy Y, Hollunder J, Meins F, Jr., Kovalchuk I. Transgenerational adaptation of Arabidopsis to stress requires DNA methylation and the function of Dicer-like proteins. PLoS One. 2010;5:e9514. doi: 10.1371/journal.pone.0009514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.He F, Zhang X, Hu JY, Turck F, Dong X, Goebel U, Borevitz JO, de Meaux J. Widespread interspecific divergence in cis-regulation of transposable elements in the Arabidopsis genus. Mol Biol Evol. 2012;29:1081–91. doi: 10.1093/molbev/msr281. [DOI] [PubMed] [Google Scholar]
- 6.Mirouze M, Paszkowski J. Epigenetic contribution to stress adaptation in plants. Curr Opin Plant Biol. 2011;14:267–74. doi: 10.1016/j.pbi.2011.03.004. [DOI] [PubMed] [Google Scholar]
- 7.Ito H, Gaubert H, Bucher E, Mirouze M, Vaillant I, Paszkowski J. An siRNA pathway prevents transgenerational retrotransposition in plants subjected to stress. Nature. 2011;472:115–9. doi: 10.1038/nature09861. [DOI] [PubMed] [Google Scholar]
- 8.Boyko A, Kovalchuk I. Transgenerational response to stress in Arabidopsis thaliana. Plant Signal Behav. 2010;5:995–8. doi: 10.4161/psb.5.8.12227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brachi B, Aimé C, Glorieux C, Cuguen J, Roux F. Adaptive value of phenological traits in stressful environments: predictions based on seed production and laboratory natural selection. PLoS One. 2012;7:e32069. doi: 10.1371/journal.pone.0032069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Grant-Downton RT, Dickinson HG. Epigenetics and its implications for plant biology 2. The ‘epigenetic epiphany’: epigenetics, evolution and beyond. Ann Bot. 2006;97:11–27. doi: 10.1093/aob/mcj001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhu JK. Epigenome sequencing comes of age. Cell. 2008;133:395–7. doi: 10.1016/j.cell.2008.04.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chinnusamy V, Zhu JK. Epigenetic regulation of stress responses in plants. Curr Opin Plant Biol. 2009;12:133–9. doi: 10.1016/j.pbi.2008.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Sano H. Inheritance of acquired traits in plants: reinstatement of Lamarck. Plant Signal Behav. 2010;5:346–8. doi: 10.4161/psb.5.4.10803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Verhoeven KJ, Jansen JJ, van Dijk PJ, Biere A. Stress-induced DNA methylation changes and their heritability in asexual dandelions. New Phytol. 2010;185:1108–18. doi: 10.1111/j.1469-8137.2009.03121.x. [DOI] [PubMed] [Google Scholar]
- 15.Boyko A, Kovalchuk I. Genome instability and epigenetic modification--heritable responses to environmental stress? Curr Opin Plant Biol. 2011;14:260–6. doi: 10.1016/j.pbi.2011.03.003. [DOI] [PubMed] [Google Scholar]
- 16.Luna E, Bruce TJ, Roberts MR, Flors V, Ton J. Next-generation systemic acquired resistance. Plant Physiol. 2012;158:844–53. doi: 10.1104/pp.111.187468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Boyko A, Kathiria P, Zemp FJ, Yao Y, Pogribny I, Kovalchuk I. Transgenerational changes in the genome stability and methylation in pathogen-infected plants: (virus-induced plant genome instability) Nucleic Acids Res. 2007;35:1714–25. doi: 10.1093/nar/gkm029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kathiria P, Sidler C, Golubov A, Kalischuk M, Kawchuk LM, Kovalchuk I. Tobacco mosaic virus infection results in an increase in recombination frequency and resistance to viral, bacterial, and fungal pathogens in the progeny of infected tobacco plants. Plant Physiol. 2010;153:1859–70. doi: 10.1104/pp.110.157263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Mandal RK, George GP, Mittal RD. Association of Toll-like receptor (TLR) 2, 3 and 9 genes polymorphism with prostate cancer risk in North Indian population. Mol Biol Rep. 2012;39:7263–9. doi: 10.1007/s11033-012-1556-5. [DOI] [PubMed] [Google Scholar]
- 20.Simon SA, Meyers BC. Small RNA-mediated epigenetic modifications in plants. Curr Opin Plant Biol. 2011;14:148–55. doi: 10.1016/j.pbi.2010.11.007. [DOI] [PubMed] [Google Scholar]
- 21.Rasmann S, De Vos M, Casteel CL, Tian D, Halitschke R, Sun JY, Agrawal AA, Felton GW, Jander G. Herbivory in the previous generation primes plants for enhanced insect resistance. Plant Physiol. 2012;158:854–63. doi: 10.1104/pp.111.187831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Feng S, Jacobsen SE. Epigenetic modifications in plants: an evolutionary perspective. Curr Opin Plant Biol. 2011;14:179–86. doi: 10.1016/j.pbi.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bilichak A, Ilnystkyy Y, Hollunder J, Kovalchuk I. The progeny of Arabidopsis thaliana plants exposed to salt exhibit changes in DNA methylation, histone modifications and gene expression. PLoS One. 2012;7:e30515. doi: 10.1371/journal.pone.0030515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lario LD, Ramirez-Parra E, Gutierrez C, Casati P, Spampinato CP. Regulation of plant MSH2 and MSH6 genes in the UV-B-induced DNA damage response. J Exp Bot. 2011;62:2925–37. doi: 10.1093/jxb/err001. [DOI] [PubMed] [Google Scholar]
- 25.Li L, Jean M, Belzile F. The impact of sequence divergence and DNA mismatch repair on homeologous recombination in Arabidopsis. Plant J. 2006;45:908–16. doi: 10.1111/j.1365-313X.2006.02657.x. [DOI] [PubMed] [Google Scholar]
- 26.Yao Y, Kovalchuk I. Abiotic stress leads to somatic and heritable changes in homologous recombination frequency, point mutation frequency and microsatellite stability in Arabidopsis plants. Mutat Res. 2011;707:61–6. doi: 10.1016/j.mrfmmm.2010.12.013. [DOI] [PubMed] [Google Scholar]
- 27.Gong Z, Morales-Ruiz T, Ariza RR, Roldán-Arjona T, David L, Zhu JK. ROS1, a repressor of transcriptional gene silencing in Arabidopsis, encodes a DNA glycosylase/lyase. Cell. 2002;111:803–14. doi: 10.1016/S0092-8674(02)01133-9. [DOI] [PubMed] [Google Scholar]
- 28.Thorstensen T, Grini PE, Aalen RB. SET domain proteins in plant development. Biochim Biophys Acta. 2011;1809:407–20. doi: 10.1016/j.bbagrm.2011.05.008. [DOI] [PubMed] [Google Scholar]
- 29.Johnson LM, Law JA, Khattar A, Henderson IR, Jacobsen SE. SRA-domain proteins required for DRM2-mediated de novo DNA methylation. PLoS Genet. 2008;4:e1000280. doi: 10.1371/journal.pgen.1000280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ebbs ML, Bender J. Locus-specific control of DNA methylation by the Arabidopsis SUVH5 histone methyltransferase. Plant Cell. 2006;18:1166–76. doi: 10.1105/tpc.106.041400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ebbs ML, Bartee L, Bender J. H3 lysine 9 methylation is maintained on a transcribed inverted repeat by combined action of SUVH6 and SUVH4 methyltransferases. Mol Cell Biol. 2005;25:10507–15. doi: 10.1128/MCB.25.23.10507-10515.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Schramm F, Ganguli A, Kiehlmann E, Englich G, Walch D, von Koskull-Döring P. The heat stress transcription factor HsfA2 serves as a regulatory amplifier of a subset of genes in the heat stress response in Arabidopsis. Plant Mol Biol. 2006;60:759–72. doi: 10.1007/s11103-005-5750-x. [DOI] [PubMed] [Google Scholar]
- 33.Kotak S, Larkindale J, Lee U, von Koskull-Döring P, Vierling E, Scharf KD. Complexity of the heat stress response in plants. Curr Opin Plant Biol. 2007;10:310–6. doi: 10.1016/j.pbi.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 34.Porter JR. Rising temperatures are likely to reduce crop yields. Nature. 2005;436:174. doi: 10.1038/436174b. [DOI] [PubMed] [Google Scholar]
- 35.Prasad PV, Chaube SK, Shrivastav TG, Kumari GL. Development of colorimetric enzyme-linked immunosorbent assay for human chorionic gonadotropin. J Immunoassay Immunochem. 2006;27:15–30. doi: 10.1080/15321810500403649. [DOI] [PubMed] [Google Scholar]
- 36.Prasad PV, Rani A, Chaube SK, Rohil V, Shrivastav TG. Kinetic analysis of a human chorionic gonadotropin-beta epitope-paratope interaction. Growth Factors. 2008;26:331–42. doi: 10.1080/08977190802445354. [DOI] [PubMed] [Google Scholar]
- 37.Suter L, Widmer A. Phenotypic effects of salt and heat stress over three generations in Arabidopsis thaliana. PLoS One. 2013;8:e80819. doi: 10.1371/journal.pone.0080819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bos H, Andersen S, Rossing P, De Zeeuw D, Parving HH, De Jong PE, Navis G. Role of patient factors in therapy resistance to antiproteinuric intervention in nondiabetic and diabetic nephropathy. Kidney Int Suppl. 2000;75:S32–7. doi: 10.1046/j.1523-1755.2000.07506.x. [DOI] [PubMed] [Google Scholar]
- 39.Henderson IR, Zhang X, Lu C, Johnson L, Meyers BC, Green PJ, Jacobsen SE. Dissecting Arabidopsis thaliana DICER function in small RNA processing, gene silencing and DNA methylation patterning. Nat Genet. 2006;38:721–5. doi: 10.1038/ng1804. [DOI] [PubMed] [Google Scholar]
- 40.Morrison MJ, Stewart DW. Heat Stress during Flowering in Summer Brassica. Crop Sci. 2002;42:797–803. doi: 10.2135/cropsci2002.0797. [DOI] [Google Scholar]
- 41.Sadras VO. Evolutionary aspects of the trade-off between seed size and number in crops. Field Crops Res. 2007;100:125–38. doi: 10.1016/j.fcr.2006.07.004. [DOI] [Google Scholar]
- 42.Dornbos DL, Mullen RE. Influence of stress during soybean seed fill on seed weight, germination, and seedling growth rate. Can J Plant Sci. 1991;71:373–83. doi: 10.4141/cjps91-052. [DOI] [Google Scholar]
- 43.Cicchino JB, Aslin RN, Rakison DH. Correspondences between what infants see and know about causal and self-propelled motion. Cognition. 2011;118:171–92. doi: 10.1016/j.cognition.2010.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Prasad PV, Craufurd PQ, Summerfield RJ, Wheeler TR. Effects of short episodes of heat stress on flower production and fruit-set of groundnut (Arachis hypogaea L.) J Exp Bot. 2000;51:777–84. doi: 10.1093/jexbot/51.345.777. [DOI] [PubMed] [Google Scholar]
- 45.Peet M, Ramchand CN, Lee J, Telang SD, Vankar GK, Shah S, Wei J. Association of the Ban I dimorphic site at the human cytosolic phospholipase A2 gene with schizophrenia. Psychiatr Genet. 1998;8:191–2. doi: 10.1097/00041444-199800830-00010. [DOI] [PubMed] [Google Scholar]
- 46.Balasubramanian S, Sureshkumar S, Lempe J, Weigel D. Potent induction of Arabidopsis thaliana flowering by elevated growth temperature. PLoS Genet. 2006;2:e106. doi: 10.1371/journal.pgen.0020106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Matsunaga W, Kobayashi A, Kato A, Ito H. The effects of heat induction and the siRNA biogenesis pathway on the transgenerational transposition of ONSEN, a copia-like retrotransposon in Arabidopsis thaliana. Plant Cell Physiol. 2012;53:824–33. doi: 10.1093/pcp/pcr179. [DOI] [PubMed] [Google Scholar]
- 48.Wessler SR. Turned on by stress. Plant retrotransposons. Curr Biol. 1996;6:959–61. doi: 10.1016/S0960-9822(02)00638-3. [DOI] [PubMed] [Google Scholar]
- 49.Capy P, Gasperi G, Biémont C, Bazin C. Stress and transposable elements: co-evolution or useful parasites? Heredity (Edinb) 2000;85:101–6. doi: 10.1046/j.1365-2540.2000.00751.x. [DOI] [PubMed] [Google Scholar]
- 50.Grandbastien M-A. Acitvation of plant retrotransposons under stress conditions. Trends Plant Sci. 1998;3:181–7. doi: 10.1016/S1360-1385(98)01232-1. [DOI] [Google Scholar]
- 51.Okamoto H, Hirochika H. Efficient insertion mutagenesis of Arabidopsis by tissue culture-induced activation of the tobacco retrotransposon Tto1. Plant J. 2000;23:291–304. doi: 10.1046/j.1365-313x.2000.00779.x. [DOI] [PubMed] [Google Scholar]
- 52.Sugimoto K, Takeda S, Hirochika H. MYB-related transcription factor NtMYB2 induced by wounding and elicitors is a regulator of the tobacco retrotransposon Tto1 and defense-related genes. Plant Cell. 2000;12:2511–28. doi: 10.1105/tpc.12.12.2511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Takeda S, Sugimoto K, Kakutani T, Hirochika H. Linear DNA intermediates of the Tto1 retrotransposon in Gag particles accumulated in stressed tobacco and Arabidopsis thaliana. Plant J. 2001;28:307–17. doi: 10.1046/j.1365-313X.2001.01151.x. [DOI] [PubMed] [Google Scholar]
- 54.Labra M, Ghiani A, Citterio S, Sgorbati S, Sala F, Vannini C, Ruffini-Castiglione M, Bracale M. Analysis of cytosine methylation pattern in response to water deficit in pea root tips. Plant Biol. 2002;4:694–9. doi: 10.1055/s-2002-37398. [DOI] [Google Scholar]
- 55.Aina R, Sgorbati S, Santagostino A, Labra M, Ghiani A, Citterio S. Specific hypomethylation of DNA is induced by heavy metals in white clover and industrial hemp. Physiol Plant. 2004;121:472–80. doi: 10.1111/j.1399-3054.2004.00343.x. [DOI] [Google Scholar]
- 56.Yao Y, Bilichak A, Golubov A, Kovalchuk I. Local infection with oilseed rape mosaic virus promotes genetic rearrangements in systemic Arabidopsis tissue. Mutat Res. 2011;709-710:7–14. doi: 10.1016/j.mrfmmm.2011.02.014. [DOI] [PubMed] [Google Scholar]
- 57.Saleh A, Alvarez-Venegas R, Avramova Z. An efficient chromatin immunoprecipitation (ChIP) protocol for studying histone modifications in Arabidopsis plants. Nat Protoc. 2008;3:1018–25. doi: 10.1038/nprot.2008.66. [DOI] [PubMed] [Google Scholar]
- 58.Boyko A, Kovalchuk I. Detection of changes in global genome methylation using the cytosine-extension assay. Methods Mol Biol. 2010;631:33–9. doi: 10.1007/978-1-60761-646-7_4. [DOI] [PubMed] [Google Scholar]
- 59.Rozhon W, Baubec T, Mayerhofer J, Mittelsten Scheid O, Jonak C. Rapid quantification of global DNA methylation by isocratic cation exchange high-performance liquid chromatography. Anal Biochem. 2008;375:354–60. doi: 10.1016/j.ab.2008.01.001. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.