

Abstract
Iron (Fe) and copper (Cu) homeostasis are tightly linked across biology. Understanding crosstalk between Fe and Cu nutrition could lead to strategies for improved growth on soils with low or excess metals, with implications for agriculture and phytoremediation. Here, we show that Cu and Fe nutrition interact to increase or decrease Fe and/or Cu accumulation in leaves and Fe uptake processes. Leaf Cu concentration increased under low Fe supply, while high Cu lowered leaf Fe concentration. Ferric reductase activity, an indicator of Fe demand, was inhibited at insufficient or high Cu supply. Surprisingly, plants grown without Fe were more susceptible to Cu toxicity.
Keywords: iron, copper, mineral homeostasis, ferric-chelate reductase, uptake
Iron (Fe) and copper (Cu) are trace heavy metals that are required by plants for their roles in redox chemistry. These metals form the active sites in numerous enzymes involved in disparate processes, such as mitochondrial respiration, photosynthesis, oxidative stress protection, and various metabolic pathways.1-4 Low metal concentration leads to deficiency and inefficiencies in metabolism, while too much causes metal toxicity. Consequences of metal toxicity are oxidative damage to cellular components,5 or displacement of the correct metal from active sites in proteins.6 Certain soils may have too little or low availability of metal micronutrients naturally, whereas other soils may have excess metals naturally or due to human activities.7,8 Excess or deficient supply of one metal may lead to deficiencies or toxicities of other metals, thus there is a need for the plant to have crosstalk mechanisms to coordinate uptake, chelation, transport, or other gene expression mechanisms to maintain metal homeostasis.
Crosstalk between Fe and Cu has been shown to occur in previous reports. In chloroplasts, superoxide dismutases (SODs) scavenge reactive oxygen species, and both Fe-containing SODs (FeSODs) and Cu-containing (CuSODs) are present, with both functioning equivalently.9 We noted that CuSODs had increased expression under Fe deficiency, while FeSOD expression decreased.10,11 Cu concentrations in Arabidopsis and other species were higher in Fe-deficient leaves.11-14 Together, this suggested that accumulation of Cu under Fe deficiency allows replacement of FeSOD proteins with CuSOD proteins. A physiological role for this switch from FeSODs to CuSODs was demonstrated, because inability to make this switch resulted in decreased ability to counteract oxidative stress.11 Additionally, certain microRNAs that are known to be regulated by Cu and to target transcripts of Cu-containing proteins15-18 were also regulated by Fe.11
Fe-Cu crosstalk may also influence mineral transport and uptake. Rosette Cu concentration of Fe-deficient Arabidopsis plants had doubled within 24 h of switching plants to Fe-deficient nutrient solution.11 Several Cu and Fe related genes respond to both Fe and Cu deficiencies, such as COPT2, ZIP2, and the ferric-chelate reductase FRO3.10,11,19-26 Similarly, Cu deficiency upregulated ferric-chelate reductase activity in roots of a number of species.14,27
To test whether crosstalk between Fe and Cu supply influences accumulation of Fe and Cu in leaves, Arabidopsis plants were grown in hydroponics10 for 9 d on Fe and Cu treatments (Fig. 1A). When Cu was withheld, rosette Cu concentration was low and did not change across Fe supply. At 0.2 µM Cu, rosette Cu concentration was higher at 1 µM Fe than at 25 µM Fe, whereas at 0.5 µM Cu supply, rosettes accumulated significantly more Cu at 1 and 5 µM Fe than at 25 µM Fe. These results indicate that moderate-term low Fe supply increases Cu accumulation in Arabidopsis rosettes, and that this phenomenon is not limited to conditions of sudden withdrawal of Fe supply as performed previously.11 Iron and Cu treatments had no effect on overall Fe concentration in rosettes (Fig. 1B), indicating that total leaf Fe quantity was maintained, although it is possible that the Fe was partitioned to different compartments or proteins under low Fe supply.28 In the first leaf of cucumber plants (Fig. 1C), Fe deficiency (0 µM Fe supply) decreased Fe concentration to 42% of that of Fe replete leaves, and increased Cu concentration by 2.8-fold. This result supports our model that Fe-deficient plants require additional Cu in leaves to supply CuSOD proteins. In previous results, Fe-deficient Arabidopsis also accumulated Zn, but later than Cu and to lower relative levels,11 and Mn concentration was not increased, supporting the idea that Cu accumulation is specific and not a secondary effect of increased metal uptake.
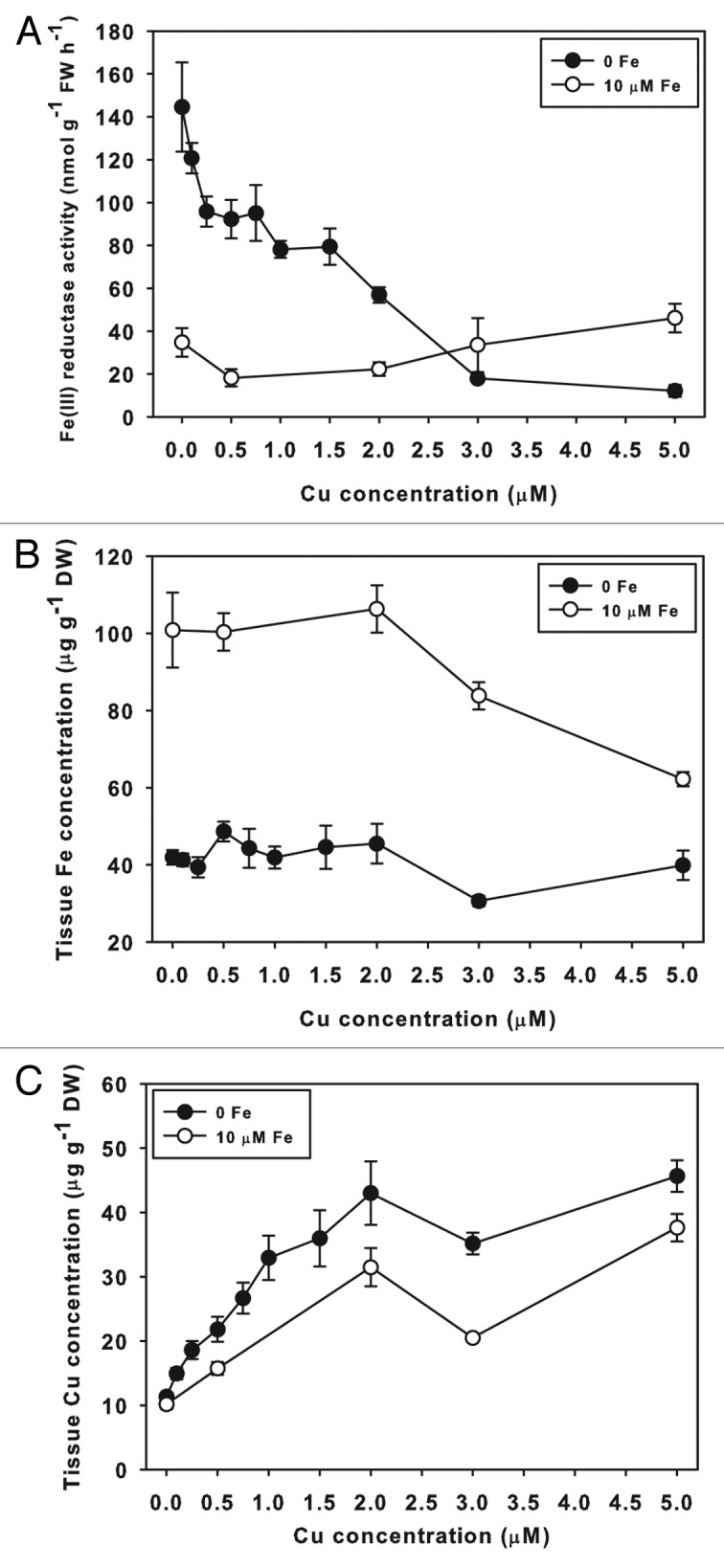
Figure 1. Iron and Cu concentrations in Arabidopsis and cucumber leaves. Arabidopsis rosette (A) Cu and (B) Fe concentration at different Fe and Cu supplies. Plants were grown for 9 d on hydroponic solution with Cu and Fe supply as indicated. (C) Cucumber first leaf Fe and Cu concentrations after 7 d growth on –Fe or +Fe (10 µM) solution. * indicates significant difference from +Fe (P ≤ 0.05) as determined by t-test.
Root ferric-chelate reductase activity is a reliable biomarker for plant Fe uptake activity, and thus an indicator of Fe sufficiency/deficiency status. To test Fe-Cu crosstalk effects on Fe uptake responses, we measured root ferric-chelate reductase activity in Arabidopsis thaliana plants treated with 0 or 25 µM Fe and a range of Cu for 3 d (Fig. 2A). Peak ferric-chelate reductase activity was observed at 0.5 µM, while at lower Cu supply, a decrease in Fe reductase activity was observed. As Cu supply increased above 0.5 µM, ferric-chelate reductase activity decreased. When Cu was at 0.75 µM, there was no elevation in ferric-chelate reductase in –Fe plants over +Fe, and at 1.0 and 1.5 µM, ferric-chelate reductase of –Fe plants was less than half that of +Fe plants. This was due to Cu toxicity in the –Fe plants, as the –Fe roots with 1.0 or 1.5 µM Cu were noticeably stunted and discolored, with a green-brown appearance, while +Fe plants treated with 1.5 µM Cu had a normal bright white coloration. We have grown Arabidopsis (with 25 µM Fe) at Cu supply of up to 5 µM with no signs of toxicity. Thus, Fe-deficient plants are substantially more sensitive to Cu toxicity than Fe replete plants.
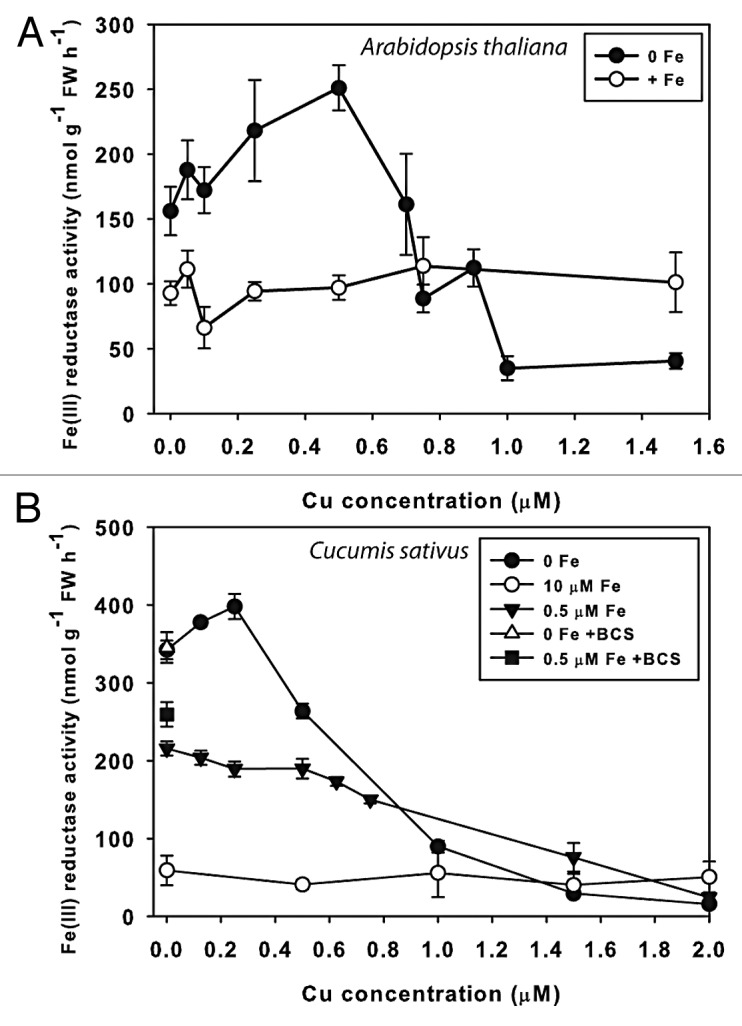
Figure 2. Effect of Cu supply on root ferric-chelate reductase activity. (A) Hydroponic Arabidopsis thaliana (Col-0) root ferric-chelate reductase activity in Fe sufficient or deficient plants, treated for 3 d with +Fe (25 μM) or –Fe and Cu concentrations as indicated. (B) Cucumber root ferric-chelate reductase activity in Fe sufficient or deficient plants, treated for 3 d with 10 μM, 0.5 μM, or –Fe, with indicated Cu concentrations in the hydroponic solution. Some treatments also included 5 μM BCS to test for Cu contamination.
A similar experiment was conducted using cucumber (Cucumis sativus cv Ashley, Jung Seed Co, Randolph, Wisconsin USA) seedlings (Fig. 2B). Cucumber plants were switched from complete nutrient solution to treatments of 0 or 0.5 µM Fe, and a range of Cu. Supplying cucumber plants with 0.5 µM Fe is sufficiently low to induce ferric-chelate reductase, but does not result in severe leaf chlorosis. In –Fe treatments, Fe reductase peaked at 0.25 µM Cu, and was lower at 0.1 µM and 0 Cu. Ferric-chelate reductase decreased at concentrations above 0.25 µM, until at 1.5 µM Cu the activity was not higher than in +Fe plants. Plants grown on 0.5 µM Fe had highest ferric-chelate reductase activity at 0 Cu. Activity declined as Cu supply increased, and at higher Cu concentrations 0 Fe, 0.5 µM Fe, and 10 µM Fe treatments had similar ferric-chelate reductase. To test whether there were trace amounts of Cu in our solution that might influence the results, we also included a treatment with the Cu chelator BCS (bathocuproinedisulfonic acid, Sigma Chemical Co) to bind potential trace Cu. This treatment resulted in no additional ferric-chelate reductase activity for 0 Fe plants, and a slight increase in 0.5 µM Fe plants.
Pumpkin (Cucurbita max cv Big Max, Eden Brothers, Dahlonega, Georgia USA) seedlings were grown in a hydroponic solution used previously for Cucurbita pepo.29 Ferric-chelate reductase activity in Fe-deficient pumpkin roots was highest at 0 Cu. As Cu supply increased, ferric-chelate reductase activity decreased (Fig. 3A), until at 3 µM Cu, activity was the same as in control plants, and activity was inhibited at 5 µM Cu. Roots of the 0 Fe, 5 µM Cu treatment looked similar to roots of the 10 µM Fe 5 µM Cu treatment and were not stunted and discolored like roots of Arabidopsis thaliana on Fe-deficient, high-Cu treatments, suggesting that pumpkin may be the more Cu tolerant species. Iron concentration in the first leaf of pumpkin did not vary across the Cu treatments in the –Fe treatment, but in +Fe treatments decreased at 3 and 5 µM Cu (Fig. 3B). This suggests that at high Cu supply, the plant decreased its demand for and subsequently accumulated less Fe. However, Cu concentrations steadily increased as Cu supply increased in both Fe treatments (Fig. 3C), until leveling off at 2 µM. Copper concentration was higher in the leaves of Fe-deficient plants at all levels above 0 µM Cu. This provides another example of Fe-Cu crosstalk, corroborating our results from Arabidopsis and cucumber, which together suggest that Fe status of plants modulates Cu uptake and accumulation in leaves.
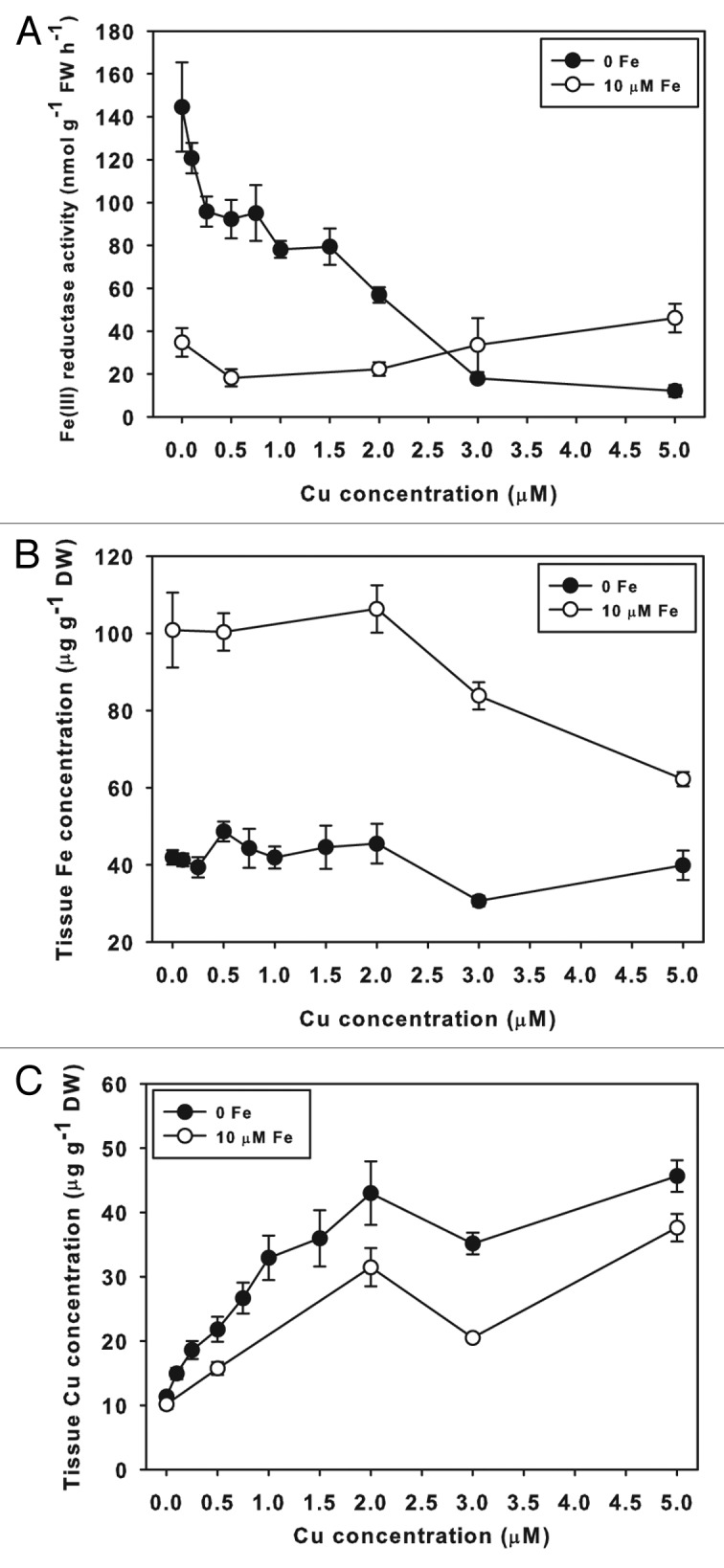
Figure 3. Effect of Cu supply on root ferric-chelate reductase activity and leaf Fe and Cu accumulation. (A) Pumpkin root ferric-chelate reductase activity in Fe sufficient or deficient plants, treated for 3d with –Fe or +Fe (10 μM) and different Cu concentrations in the hydroponic solution. (B) Iron concentration and (C) Cu concentration of first leaf of pumpkin plants from panel A.
The relationship between ferric-chelate reductase activity and Cu supply to Fe-deficient plants (Fig. 2) is complex, with an optimum Cu concentration of 0.5 and 0.25 µM for Fe-deficient Arabidopsis and cucumber, respectively. Along the Cu supply curve there were three phases. In the first phase, below optimal Cu, there was an inhibition of ferric-chelate reductase activity, suggesting that Cu may be required for some aspect of ferric-chelate reductase synthesis or activity. A similar inhibition for whole root ferric-chelate reductase activity by low Cu was observed for Plantago lanceolata.30 In pumpkin, and in cucumber supplied with 0.5 µM Fe, 0 Cu was optimal. As Cu was increased above optimal concentrations, lower ferric-chelate reductase activity occurred. It is possible that increased Cu results in more efficient or rapid synthesis of Cu proteins that replace Fe proteins, thus reducing the Fe demand and generating a feedback inhibition of ferric-chelate reductase activity or decreased shoot-to-root demand signal. For example, in Arabidopsis, FeSOD expression increased under Cu deficiency31,32 and decreased under high Cu supply.33 Indeed, ferric-chelate reductase activity decreased (Fig. 3A) as Cu concentration in leaves of Fe-deficient pumpkin increased (Fig. 3C). The third phase of the curve indicates Cu toxicity, which abolished ferric-chelate reductase activity. Similar inhibition of ferric-chelate reductase activity by high Cu has been observed in cucumber34 and Plantago lanceolata,30 but high Cu did not inhibit ferric-chelate reductase protein activity in Plantago membrane isolates, which suggested that Cu blocked expression rather than function of the ferric reductase.
In conclusion, our results show that Fe and Cu status and supply interact to influence uptake of both metals. The sensing mechanisms are not known, but the metal chelator nicotianamine is important for Fe homestasis35,36 and can bind both Fe and Cu.37 We also showed that under Fe deficiency, plants were strikingly more susceptible to Cu toxicity. Understanding this aspect of Fe-Cu crosstalk could have implications for agriculture and for phytoremediation of metal contaminated soils. The conditions described here provide an entry point into growth conditions that can be used to produce plants that are engaging in Fe-Cu crosstalk to further understand these mechanisms.
Acknowledgments
The authors thank Grace Troupe and Alec Hogan for technical assistance. This work was supported in part by the University of Nebraska UCARE program and NSF grant no. IOS-1257568 to Waters B.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1.Burkhead JL, Reynolds KAG, Abdel-Ghany SE, Cohu CM, Pilon M. Copper homeostasis. New Phytol. 2009;182:799–816. doi: 10.1111/j.1469-8137.2009.02846.x. [DOI] [PubMed] [Google Scholar]
- 2.Hänsch R, Mendel RR. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl) Curr Opin Plant Biol. 2009;12:259–66. doi: 10.1016/j.pbi.2009.05.006. [DOI] [PubMed] [Google Scholar]
- 3.Pilon M, Ravet K, Tapken W. The biogenesis and physiological function of chloroplast superoxide dismutases. Biochim Biophys Acta. 2011;1807:989–98. doi: 10.1016/j.bbabio.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 4.Puig S, Andrés-Colás N, García-Molina A, Peñarrubia L. Copper and iron homeostasis in Arabidopsis: responses to metal deficiencies, interactions and biotechnological applications. Plant Cell Environ. 2007;30:271–90. doi: 10.1111/j.1365-3040.2007.01642.x. [DOI] [PubMed] [Google Scholar]
- 5.Dietz KJ, Baier M, Krämer U. Free Radicals and Reactive Oxygen Species as Mediators of Heavy Metal Toxicity in Plants. In: Prasad MNV, Hagemeyer J. Heavy Metal Stress in Plants. Berlin, Germany: Springer, 1999:73-97. [Google Scholar]
- 6.Nishida S, Aisu A, Mizuno T. Induction of IRT1 by the nickel-induced iron-deficient response in Arabidopsis. Plant Signal Behav. 2012;7:329–31. doi: 10.4161/psb.19263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Clemens S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie. 2006;88:1707–19. doi: 10.1016/j.biochi.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 8.Nagajyoti PC, Lee KD, Sreekanth TVM. Heavy metals, occurrence and toxicity for plants: a review. Environ Chem Lett. 2010;8:199–216. doi: 10.1007/s10311-010-0297-8. [DOI] [Google Scholar]
- 9.Pilon M, Ravet K, Tapken W. The biogenesis and physiological function of chloroplast superoxide dismutases. Biochim Biophys Acta. 2011;1807:989–98. doi: 10.1016/j.bbabio.2010.11.002. [DOI] [PubMed] [Google Scholar]
- 10.Stein RJ, Waters BM. Use of natural variation reveals core genes in the transcriptome of iron-deficient Arabidopsis thaliana roots. J Exp Bot. 2012;63:1039–55. doi: 10.1093/jxb/err343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Waters BM, McInturf SA, Stein RJ. Rosette iron deficiency transcript and microRNA profiling reveals links between copper and iron homeostasis in Arabidopsis thaliana. J Exp Bot. 2012;63:5903–18. doi: 10.1093/jxb/ers239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chaignon V, Di Malta D, Hinsinger P. Fe-deficiency increases Cu acquisition by wheat cropped in a Cu-contaminated vineyard soil. New Phytol. 2002;154:121–30. doi: 10.1046/j.1469-8137.2002.00349.x. [DOI] [Google Scholar]
- 13.Waters BM, Troupe GC. Natural variation in iron use efficiency and mineral remobilization in cucumber (Cucumis sativus) Plant Soil. 2012;352:185–97. doi: 10.1007/s11104-011-0988-3. [DOI] [Google Scholar]
- 14.Welch RM, Norvell WA, Schaefer SC, Shaff JE, Kochian LV. Induction of iron(III) and copper(II) reduction in pea (Pisum sativum L.) roots by Fe and Cu staus: Does the root-cell plasmalemma Fe(III)-chelate reductase perform a general role in regulating cation uptake? Planta. 1993;1993:555–61. [Google Scholar]
- 15.Abdel-Ghany SE, Pilon M. MicroRNA-mediated systemic down-regulation of copper protein expression in response to low copper availability in Arabidopsis. J Biol Chem. 2008;283:15932–45. doi: 10.1074/jbc.M801406200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sunkar R, Kapoor A, Zhu JK. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell. 2006;18:2051–65. doi: 10.1105/tpc.106.041673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yamasaki H, Abdel-Ghany SE, Cohu CM, Kobayashi Y, Shikanai T, Pilon M. Regulation of copper homeostasis by micro-RNA in Arabidopsis. J Biol Chem. 2007;282:16369–78. doi: 10.1074/jbc.M700138200. [DOI] [PubMed] [Google Scholar]
- 18.Yamasaki H, Hayashi M, Fukazawa M, Kobayashi Y, Shikanai T. SQUAMOSA promoter binding protein-like7 is a central regulator for copper homeostasis in Arabidopsis. Plant Cell. 2009;21:347–61. doi: 10.1105/tpc.108.060137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Buckhout TJ, Yang TJW, Schmidt W. Early iron-deficiency-induced transcriptional changes in Arabidopsis roots as revealed by microarray analyses. BMC Genomics. 2009;10:147. doi: 10.1186/1471-2164-10-147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Colangelo EP, Guerinot ML. The essential basic helix-loop-helix protein FIT1 is required for the iron deficiency response. Plant Cell. 2004;16:3400–12. doi: 10.1105/tpc.104.024315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.del Pozo T, Cambiazo V, González M. Gene expression profiling analysis of copper homeostasis in Arabidopsis thaliana. Biochem Biophys Res Commun. 2010;393:248–52. doi: 10.1016/j.bbrc.2010.01.111. [DOI] [PubMed] [Google Scholar]
- 22.García MJ, Lucena C, Romera FJ, Alcántara E, Pérez-Vicente R. Ethylene and nitric oxide involvement in the up-regulation of key genes related to iron acquisition and homeostasis in Arabidopsis. J Exp Bot. 2010;61:3885–99. doi: 10.1093/jxb/erq203. [DOI] [PubMed] [Google Scholar]
- 23.Mukherjee I, Campbell NH, Ash JS, Connolly EL. Expression profiling of the Arabidopsis ferric chelate reductase (FRO) gene family reveals differential regulation by iron and copper. Planta. 2006;223:1178–90. doi: 10.1007/s00425-005-0165-0. [DOI] [PubMed] [Google Scholar]
- 24.Sancenón V, Puig S, Mira H, Thiele DJ, Peñarrubia L. Identification of a copper transporter family in Arabidopsis thaliana. Plant Mol Biol. 2003;51:577–87. doi: 10.1023/A:1022345507112. [DOI] [PubMed] [Google Scholar]
- 25.Wintz H, Fox T, Wu YY, Feng V, Chen WQ, Chang HS, Zhu T, Vulpe C. Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis. J Biol Chem. 2003;278:47644–53. doi: 10.1074/jbc.M309338200. [DOI] [PubMed] [Google Scholar]
- 26.Yang TJW, Lin W-D, Schmidt W. Transcriptional profiling of the Arabidopsis iron deficiency response reveals conserved transition metal homeostasis networks. Plant Physiol. 2010;152:2130–41. doi: 10.1104/pp.109.152728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Chen YX, Shi JY, Tian GM, Zheng SJ, Lin Q. Fe deficiency induces Cu uptake and accumulation in Commelina communis. Plant Sci. 2004;166:1371–7. doi: 10.1016/j.plantsci.2004.01.018. [DOI] [Google Scholar]
- 28.Page MD, Allen MD, Kropat J, Urzica EI, Karpowicz SJ, Hsieh SI, Loo JA, Merchant SS. Fe sparing and Fe recycling contribute to increased superoxide dismutase capacity in iron-starved Chlamydomonas reinhardtii. Plant Cell. 2012;24:2649–65. doi: 10.1105/tpc.112.098962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Waters BM, Blevins DG. Ethylene production, cluster root formation, and localization of iron(III) reducing capacity in Fe-deficient squash roots. Plant Soil. 2000;225:21–31. doi: 10.1023/A:1026513022280. [DOI] [Google Scholar]
- 30.Schmidt W, Bartels M, Tittel J, Fuhner C. Physiological effects of copper on iron acquisition processes in Plantago. New Phytol. 1997;135:659–66. doi: 10.1046/j.1469-8137.1997.00691.x. [DOI] [Google Scholar]
- 31.Bernal M, Casero D, Singh V, Wilson GT, Grande A, Yang H, Dodani SC, Pellegrini M, Huijser P, Connolly EL, et al. Transcriptome sequencing identifies SPL7-regulated copper acquisition genes FRO4/FRO5 and the copper dependence of iron homeostasis in Arabidopsis. Plant Cell. 2012;24:738–61. doi: 10.1105/tpc.111.090431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Perea-García A, Garcia-Molina A, Andrés-Colás N, Vera-Sirera F, Pérez-Amador MA, Puig S, Peñarrubia L. Arabidopsis copper transport protein COPT2 participates in the cross talk between iron deficiency responses and low-phosphate signaling. Plant Physiol. 2013;162:180–94. doi: 10.1104/pp.112.212407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Schiavon M, Zhang LH, Abdel-Ghany SE, Pilon M, Malagoli M, Pilon-Smits EAH. Variation in copper tolerance in Arabidopsis thaliana accessions Columbia, Landsberg erecta and Wassilewskija. Physiol Plant. 2007;129:342–50. doi: 10.1111/j.1399-3054.2006.00809.x. [DOI] [Google Scholar]
- 34.Alcántara E, Romera FJ, Canete M, Delaguardia MD. Effects of heavy-metals on both induction and function of root Fe(III) reductase in Fe-deficient cucumber (Cucumis sativus L) plants. J Exp Bot. 1994;45:1893–8. doi: 10.1093/jxb/45.12.1893. [DOI] [Google Scholar]
- 35.Cassin G, Mari S, Curie C, Briat JF, Czernic P. Increased sensitivity to iron deficiency in Arabidopsis thaliana overaccumulating nicotianamine. J Exp Bot. 2009;60:1249–59. doi: 10.1093/jxb/erp007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Klatte M, Schuler M, Wirtz M, Fink-Straube C, Hell R, Bauer P. The analysis of Arabidopsis nicotianamine synthase mutants reveals functions for nicotianamine in seed iron loading and iron deficiency responses. Plant Physiol. 2009;150:257–71. doi: 10.1104/pp.109.136374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.von Wiren N, Klair S, Bansal S, Briat JF, Khodr H, Shioiri T, Leigh RA, Hider RC. Nicotianamine chelates both FeIII and FeII. Implications for metal transport in plants. Plant Physiol. 1999;119:1107–14. doi: 10.1104/pp.119.3.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]