

Abstract
Synchronic measurements of spontaneous ultra-weak light emission from germinating wheat seedlings both in Brazil and after transportation to Japan, and with a simultaneous series of germinations with local seedlings in the Czech Republic, are presented. A series of tests was also performed with samples returned from Japan to Brazil and results compared with those from undisturbed Brazilian seedlings. Native seedlings presented semi-circadian rhythms of emission which correlated with the gravimetric tidal acceleration at their locality, as did seeds which had been transported from Brazil to Japan, and then returned to Brazil. Here, however, there were very small disturbances within the periodicity of emissions, perhaps as a result of similar tidal profiles at locations whose longitudes are 180° apart, as in this case, different from previous results obtained in Brazil–Germany tests with other longitude shift. This feature of the Brazil and Japan locations may have minimized the requirement for the acclimatization of the transported seed to their new location.
Keywords: germination, biophoton emission, gravimetric tides, chronobiology
It was shown that plants present circadian-like rhythms which are synchronised with the temporal variation of the local gravimetric tidal acceleration, or “Etide,” which stands as a proxy for the lunisolar gravitational “pull” experienced at the Earth’s surface.1,2 This was found also for the spontaneous, ultra-weak photon emission (UPE) from developing wheat seedlings.3 This spontaneous light emission is related to the timecourse and activity of metabolic processes during germination and early seedling growth, when rapid reproduction occurs promoting new tissues for leaflets and radicles;4 it can be used as a non-invasive, real-time probe of the living state.5,6
A previous report concerned germination tests that were performed simultaneously, at 2 locations, one in Brazil using local seeds, and a second in Germany using seed samples brought from Brazil. Local German seeds were also tested at the same time. In each location, the same test was performed at the same hour, UTC, but with the time adjusted to local time (i.e., UTC –4 h in Brazil, and UTC +1h in Germany). Indications of both a phase-lag with respect to the local tide, together with indications of some sort of acclimatization to the local gravimetric rhythm, were found in the Brazil seedlings transported to Europe.7
In order to test further our hypothesis of a close relationship between not only the rhythms of tidal acceleration and photon emission but also that these 2 rhythms are dependent upon the location, a further transcontinental test with wheat seedlings was performed. In this case, 3 widely separated locations were selected: one in Brazil, a second in Japan, and a third in the Czech Republic. Tests in Japan used seed samples brought from Brazil in order to investigate any possible mistuning or time delay with respect to the Etide/photon emission timecourses, as found in the above-mentioned Brazil–Germany transcontinental tests.7 Subsequently, Brazilian seeds, which had been tested in Japan, were brought back to Brazil from Japan and put to germinate. Their photon emissions were then measured in parallel with undisturbed seeds kept in Brazil from the beginning. UPE data of tests were analyzed by means of superimposing their timeseries upon that of the relevant local gravimetric tidal estimates.
In the data presented here, local seedlings of wheat growing both in Limeira (SP, Brazil) and in Prague (Czech Republic) present UPE data synchronous with the local tide, as expected.3,7 The samples transported for testing in Japan, and later brought back and tested in Brazil, also appeared almost in tune with both the local gravimetric tide and with the UPE timecourse from seedlings that had not been transported around the globe, and showed no significant phase-lag or mistuning, as found in the previous series of 2006.7 One possible explanation is that the geographical longitude of the test site in Japan (137° E) is diametrically opposite in the east–west direction, about 180° away from the test site in Brazil (47° W), and so Japanese and Brazilian gravimetric variations in time are almost in phase, though separated by a 12-h time displacement (UTC +9h for Japan, UTC –3h for Brazil). Latitudes vary as well, but basically the north–south difference alters the cycle’s amplitude (daily extrema) while its phase is minimally disturbed.
Thus, even flying for 24 h to reach their final destination, the transported seeds find the same gravimetric tidal time pattern upon arrival: they do not experience any difference in the gravimetric tide and so there is no requirement to adjust or “acclimatize” to the new locality.
The reason for performing simultaneous observations on seedling photon emissions in 2 widely different, transcontinental locations is to check whether the rhythms of UPE are an inherent feature of the biological material, regardless of its location, or whether they might be responses to some external factor which pervades the locality of the respective test laboratories.
This approach was already used by Brown et al.8 to study the circadian biorhythm of melanophore activity of marine-dwelling crabs. Here, crabs, with a melanophore rhythm coordinated with that of the marine tides at their usual location on the east coast of the US, were transported to a novel location in a laboratory at the west coast. At this latter location, the rhythm of activity was no longer that found at the east coast location but was now in harmony with the tidal rhythm at the new west coast location, even though there was no direct exposure to the marine tides at this location. This supported the hypothesis that the biorhythm was not inherent to the crab, either as a clock or memory, but was regulated externally, according to some characteristic of the current geographical location. Brown et al. (1955) believed that it was some factor, due to the sun and moon, such as the varying lunisolar gravitational acceleration which, besides regulating the marine tide, also regulated, in a more direct, but more subtle manner, the melanophore biorhythm. A similar transportation experiment was performed by Renner,9,10 but using honey bees trained to adhere to a single, constant feeding time during a 24-h cycle. Bees trained in Long Island, NY USA, were transported to Davis, CA USA. The time difference between localities was 3.25 h (solar time). Here, over a 3-day period of testing, the feeding time was shifted by up to 1.5 h on each day so that it eventually conformed to the same relative feeding time in the new locality.
Working with the hypothesis that some biorhythms, including that of growth and, hence, of biophoton emission also, are somehow linked to the lunisolar gravimetric tide, not only with respect to the general timekeeping of the 2 rhythms due to the geographical and temporal displacement of the 2 locations, but also in the relationships to the rate of change of the 2 rhythms, wheat seeds from a stock of seeds held in Brazil (BR) were used in simultaneous experiments in both Brazil (Limeira) and Japan (Hamamatsu), while native seeds were used in similar experiments in the Czech Republic (Prague). Further, the remainder of the stock of Brazilian seeds which had traveled to Japan were brought back to Brazil, where tests were performed in parallel with tests on seeds which had remained in the Brazilian location all the while.
Samples of Brazilian bread wheat (Triticum aestivum L. cv MaisVita) seeds (BR) were transported by air to Japan via Paris, departing from Brazil (São Paulo) on 09/19/2012 and arriving in Tokyo at 21/09/2012. Tests in Hamamatsu, Japan (JP), started one week after the arrival, on 09/28/2012. A similar series of tests started on the same date in Brazil (Limeira/SP), using samples from the same stock as transported to Japan. Simultaneously, a series of tests was made in Prague (Czech Republic) using local wheat seeds (Triticum aestivum L. cv Federer). After 3 sequential tests in Japan, on 10/08/2012, the remaining stock of BR seeds was returned to Brazil, again via Paris, arriving in Brazil on the following day. A test series, using these seeds, was commenced on 10/11/2012. Technical issues caused a 2-week interruption of one of the series of tests (the D series) for the JP → BR samples on 10/15/2012, but tests were resumed on 11/01/2012.
The UPE procedures in both Limeira and Prague were based on the same photon-counting unit (H7630, Hamamatsu KK). They used 25 wheat seeds placed in a petri dish (10 cm) with distilled water (10 mL) and filter paper, and prepared in dark. The Japanese set-up used only 6 seeds in 3mL of water (2.5-cm diameter petri dish + filter paper), and a sensitive, miniaturized apparatus (Hamamatsu Type 6100, Biophoton Assay System).
Thus, all experiments ran in parallel, under the same internal laboratory conditions, using similar photon counting equipment with low and stable background noise levels, on the same date. Whereas in Limeira and Prague, the lunisolar tidal acceleration was different at the time when the simultaneous germination tests and photon counting were performed, in Hamamatsu the tide was similar to that at Limeira because they are almost 180° longitudinally apart on the globe.
Figure 1 presents the normalized plots for the residue to linear fitting of each photon-count (PC) timecourses data (presented in detail at supplementary Figure S1), for each of the tests up to 10/23/2012: A1-A6 for tests in Brazil, B1-B3 (a, b and c) for tests in Japan, C1-C8 for Czech tests, and D1 for the initial test of samples returned to Brazil from Japan.
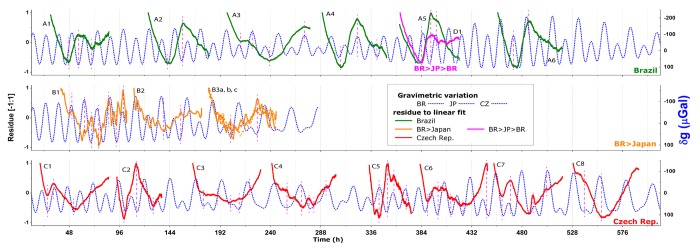
Figure 1. Normalized residue of Photon counts data (Fig. S1) to a linear growth fit: Brazilian samples in Limeira/BR (A series, top), traveled to Hamamatsu/Japan (B, middle) and back to Brazil (D1, top on A5), and Czech samples in Prague (C, bottom), plotted against the contemporaneous local gravimetric acceleration oscillation (δg). Vertical marks (dash lines) over singular PC points.
Data in these timecourses are superimposed upon the gravimetric tidal acceleration variations (δg) of the respective test location. Vertical marks are used to highlight singular points along the PC curves, i.e., the abrupt changes during the course of the first/second derivative, and possible coincidences with the points of local minima and maxima in the gravimetric rhythm.
Series A and C, both with local seeds, are clearly different in their temporal pattern of photon emission but present singular points in the PC timecourse which generally relate to δg inflexion points, coincident with, or very close to, gravimetric minima/maxima, as expected on the evidence of previous experience.3 An exception occurs at the end of test A5, just in the middle of a δg semi-cycle (at approx. 402 h), but this result could be discarded since test A5 is abnormal at this point, with a sudden decrease of PC values.
Tests in Japan – the B series – present PC curves that are less smooth than those of the other series. The PC timecourses display more peaks, but this may be because the UPE is from just 6 seedlings instead of 25 and, hence, there is less homogenization of the data. Despite this, the singular PC points are almost coincident with the local gravimetric tide. It is noticeable that in the 3rd test of this B series, where 3 samples were measured in parallel (a, b, and c), there is a simultaneous increase of PC in each sample. Also, in the upper curve, which shows the most intense emissions, there are 2 pronounced local PC peaks coincident with the local gravimetric tidal peaks (one at the 192 h, and the next approx. 12 h later).
The test D1, using Brazilian seeds returned from Japan, has similar features, the starts and finishes of the rises being synchronous with gravimetric tide inversion points. One noticeable fact is that, in this D1 test, the increase of the PCs commences one half-tide cycle later than the corresponding test in the local series A5, although the singular points in the PCs of both the A5 and D1 tests are synchronous with tidal inversion points. Unfortunately technical issues rendered incomplete the PC data of the sequential tests D2–D5 (not shown).
The remaining timecourses of tests for non-transported BR seeds performed in parallel to those that were transported to and from Japan (D6-D9), from 11/01/2012 to the end 11/14/2012, are presented together with the local gravimetric tide at Figure S2, and its normalized version at Figure 2, in similar manner as done for Figures 1 and S1.
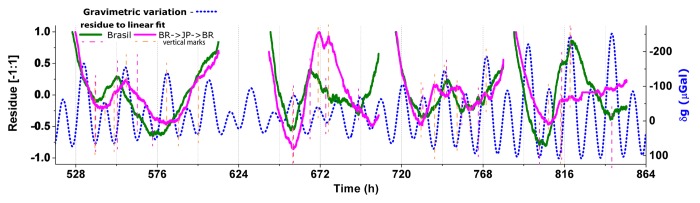
Figure 2. Normalized residue of Photon counts data (Fig. S2) to a linear growth fit: Brazilian samples in Limeira/BR and traveled back from Japan to Brazil plotted against the contemporaneous local gravimetric acceleration oscillation (δg). Vertical marks over singular PC points.
Ignoring the difference in absolute sensitivity between the photon-counting units of the set-ups in each locality, the 2 PC curves can be easily compared, as shown. It is noticeable that, for all 4 pairs of tests, the non-transported and the transported samples both present PC singular points which are almost coincident. Although there are some differences in curve shape, mainly for the 2nd and 4th pair of tests, most of the turning points seem coincident with the local minima or maxima of the gravimetric tidal pattern. The 3rd test, however, on 11/09/2012, has a curious feature: the transported sample shows PC singular points that appear to relate to earlier inflection points of δg than do the PC features of the local sample, ie., the PC pattern of the transported sample appears to be one cycle in advance of the undisturbed, non-transported sample, contrary to the PC pattern which in test D1, using the BR→JP→BR seed sample (Fig. 1, Fig. S1).
In Figures 1 and 2 not only are the PC singular points related to δg local peaks but also the plots of the residuals resemble the temporal patterns of the plotted δg time values, as can be seem for tests A1, A4, and A6 and, more clearly, for tests C1, C2, C5, and C7.
The normalized plots [0,1] for tests A2-A3 and B2-B3b, and for Brazilian and Japanese gravimetric tides are shown at Figure S3. The superimposed pair A2/B2 are practically coincident in their time evolution in the final 48h, after the initial decay (up to 120th hour), although the Japanese test had been started 6 hours before. In this case the δg of BR and of JP are almost synchronous but with alternating smaller and larger local minima.
In contrast, the pair A3/B3b keep their relative delay, with PC in Japan starting to grown around a local JP-δg minima (~197h) and the PC in Brazil starting just in the next BR-δg minima, 12 h latter. In this case the δg of BR and of JP are again in phase, but with a very pronounced difference in local amplitude.
To complement the information of Figure 2 the first time derivatives (δ/δτ) of its time plots were determined and their normalized [–1,1] plots are shown at Figure S4. In this case, local maxima and minima of curves of Figure 2 appear at Figure S4 as zero-crossing points. Important coincident zero-crossings of δ(PC)/δτ and of the δ(δg)/δτ curve are marked dashed (orange) ellipses. It is clear that, in some cases, both BR and BR→JP→BR δ(PC)/δτ data cross the zero line in synchronism with the δ(δg)/δτ crossings, while in others the data of BR→JP→BR is delayed and presents zero-crossing in the next δg singular point.
Brown et al.11 have shown 6-h and 12-h semi-circadian rhythms to occur regularly in the timecourses of oxygen consumption of carrots and potato tubers. These, and other, more minor, rhythmic components were also recognized by Ziegler et al.12 in laboratory-based timecourses of the vertical migration of caribbean spiny lobster larvae, a rhythmic process apparently driven ultimately by lunisolar tidal acceleration,13 but proximately modified by marine tidal rhythms and photoperiods, among other variables.14
With respect to the putative acclimatization of biological processes to the prevailing gravimetric tides, one should consider the factors which might need to be adjusted following the transportation of seed stocks from one locality to another. For example, the transcontinental air flight may admit some disturbance to the BR seeds. Changes in the barometric pressure during flight could be one such disturbance. It is known, for instance, that barometric pressure variations influence seed metabolism, even when the seeds are dormant.15 Another disturbing factor could be the incidence of cosmic radiation at the high altitude of the flight, in which case acclimatization may involve the repair of radiation damage to cellular organelles, such as nuclei and mitochondria within the embryo and its surrounding tissues, and that traces of these events are expressed in the subsequent photon emission patterns of the germinating seedlings.
Future work should concentrate not only on finding further correspondences between photon counts and the local gravimetric tides but also on using different longitudal distances for translocational experiments, such as those which we have here described. The advantages of these latter experiments are that, not only do they consolidate the gravimetric tide/photon count relationship, but they can also give evidence of whether or not there are endogenous rhythms in the seeds which are disturbed by the transportation, and which are subsequently corrected by an acclimatization period. The latter may or may not be mediated by the gravimetric tide itself, but at least the acclimatization seems to bring the seed into a new state of conformity with this tide.
An ideal approach would be to measure germination of local seeds transported simultaneously to and from places with intermediate (~45°) and large (~90°) longitudinal (east–west) differences. A consequence of this would be that the gravimetric tidal cycles would be de-tuned by one-quarter and one-half a cycle, respectively. Experiments at the 2 poles of the Earth could also be of interest, where the gravimetric tidal cycles are of a different structure.
Supplementary Material
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
Authors are grateful to Emile Klingelé for the gift of the Etide program, to FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo, Brazil, #04/10146–3 and #11/10763–6) and CNPq (Conselho Nacional de Pesquisa, Brazil, # 552258/2009–1) for partial lab and trip supports, and to Hamamatsu Photonics KK for supporting experiments in Japan. M.C. and K.Č. are supported by Czech Science Foundation, grant no. GP13–29294S, and are also thankful to Pavel Ill for donating the wheat seeds (Triticum aestivum cv Federer).
Supplemental Materials
Supplemental materials may be found here: www.landesbioscience.com/journals/psb/article/28671
Glossary
Abbreviations:
- PC
photon count
- UPE
ultraweak photon emission
- UTC
Universal coordinated time
References
- 1.Zürcher E, Cantiani MG, Sorbetti-Guerri F, Michel D. Tree stem diameters fluctuate with tide. Nature. 1998;392:665–666. doi: 10.1038/33570. [DOI] [Google Scholar]
- 2.Barlow PW, Fisahn J. Lunisolar tidal force and the growth of plant roots, and some other of its effects on plant movements. Ann Bot. 2012;110:301–18. doi: 10.1093/aob/mcs038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Moraes TA, Barlow PW, Klingelé E, Gallep CM. Spontaneous ultra-weak light emissions from wheat seedlings are rhythmic and synchronized with the time profile of the local gravimetric tide. Naturwissenschaften. 2012;99:465–72. doi: 10.1007/s00114-012-0921-5. [DOI] [PubMed] [Google Scholar]
- 4.Gallep CM. Ultra-weak, spontaneous photon emission in seedlings – toxicological and chronobiological applications. Luminescence. 2014 doi: 10.1002/bio.2658. Forthcoming. [DOI] [PubMed] [Google Scholar]
- 5.Devaraj B, Mashashi U, Inaba H. Biophotons: ultraweak light emission from living systems. Curr Opin Solid State Mater Sci. 1997;2:188–93. doi: 10.1016/S1359-0286(97)80064-2. [DOI] [Google Scholar]
- 6.Van Wijk R, Schamhart DH. Regulatory aspects of low intensity photon emission. Experientia. 1988;44:586–93. doi: 10.1007/BF01953306. [DOI] [PubMed] [Google Scholar]
- 7.Gallep CM, Moraes TA, Dos Santos SR, Barlow PW. Coincidence of biophoton emission by wheat seedlings during simultaneous, transcontinental germination tests. Protoplasma. 2013;250:793–6. doi: 10.1007/s00709-012-0447-x. [DOI] [PubMed] [Google Scholar]
- 8.Brown FA, Webb HM, Bennett MF. Proof for an endogenous component in persistent solar and lunar rhythmicity in organisms. Proc Natl Acad Sci U S A. 1955;41:93–100. doi: 10.1073/pnas.41.2.93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Renner M. Űber ein weiteres Versetzungsexperiment zur analyse des Zeitsinnes und der Sonnenorientierung der Honigbiene. Z Vgl Physiol. 1959;42:449–83. doi: 10.1007/BF00297804. [DOI] [Google Scholar]
- 10.Renner M. The contribution of the honey bee to the study of time-sense and astronomical orientation. Cold Spring Harb Symp Quant Biol. 1960;25:361–7. doi: 10.1101/SQB.1960.025.01.037. [DOI] [PubMed] [Google Scholar]
- 11.Brown FA, Freeland RO, Ralph CL. Persistent rhythms of O2-consumption in potatoes, carrots and the seaweed, Fucus. Plant Physiol. 1955;30:280–92. doi: 10.1104/pp.30.3.280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ziegler TA, Cohen JH, Forward RB., Jr. Proximate control of diel vertical migration in Phyllosoma larvae of the Caribbean spiny lobster Panulirus argus. Biol Bull. 2010;219:207–19. doi: 10.1086/BBLv219n3p207. [DOI] [PubMed] [Google Scholar]
- 13.Barlow PW. The daily rhythm of leaf movements reflects the rhythm of the lunisolar tidal acceleration. Plant Signal Behav. Forthcoming. [Google Scholar]
- 14.Cohen JH, Forward RB., Jr Zooplankton diel vertical migration – a review of proximate control. Oceanogr Mar Biol Annu Rev. 2009;47:77–110. [Google Scholar]
- 15.Graviou E. Analogies between rhythms in plant material, in atmospheric pressure, and solar lunar periodicities. Int J Biometeorol. 1978;22:103–11. doi: 10.1007/BF01552889. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.