

Abstract
In vitro experiments were conducted to evaluate the suitability of several mixtures of high tanniniferous non legumes with low tanniniferous legumes on in vitro gas production (IVGP), dry matter degradation, Ammonia-N, methane production and microbial population. Eight treatments were examined in a randomized complete block design using four non-legumes and two legumes (Carallia integerrima×Leucaena leucocephala (LL) (Trt 1), C. integerrima×Gliricidia sepium (GS) (Trt 2), Aporosa lindeliyana×LL (Trt 3), A. lindeliyana×GS (Trt 4), Ceiba perntandra×LL (Trt 5), C. perntandra×GS (Trt 6), Artocarpus heterophyllus×LL (Trt 7), A. heterophyllus×GS (Trt 8). The condensed tannin (CT) content of non legumes ranged from 6.2% (Carallia integerrima) to 4.9% (Ceiba perntandra) while the CT of legumes were 1.58% (Leucaena leucocephala) and 0.78% (Gliricidia sepium). Forage mixtures contained more than 14% of crude protein (CP) while the CT content ranged from 2.8% to 4.0% respectively. Differences (p<0.05) were observed in in vitro gas production (IGVP) within treatments over a 48 h period dominated by C. perntandra×G. sepium (Trt 6). The net gas production (p<0.05) was also high with Trt6 followed by A. heterophyllus×L. leucocephala (Trt 7) and A. heterophyllus×G. sepium (Trt 8). Highest (p>0.05) NH3-N (ml/200 mg DM) production was observed with the A. heterophyllus×G. sepium (Trt 8) mixture which may be attributed with it’s highest CP content. The correlation between IVGP and CT was 0.675 while IVGP and CP was 0.610. In vitro dry matter degradation (IVDMD) was highest in Trt 8 as well. Methane production ranged from 2.57 to 4.79 (ml/200 mg DM) to be synonimous with IVGP. A higher bacteria population (p<0.05) was found in C. perntandra×G. sepium (Trt 6) followed by Artocarpus heterophyllus+G. sepium (Trt 8) and the same trend was observed with the protozoa population as well. The results show that supplementing high tannin non leguminous forages by incremental substitution of legume forage increased gas production parameters, NH3-N, IVDMD and microbial population in the fermentation liquid. Methane production was not significantly affected by the presence of CT or different levels of CP in forage mixtures. Among non legumes, Ceiba perntandra and Artocarpus heterophyllus performed better in mixture with L. leucocephala and G. sepium.
Keywords: Legume, Condensed Tannins, Crude Protein, In vitro Gas Production, In vitro Dry Matter Degradation, Methane
INTRODUCTION
Goat production in Sri Lanka is becoming popular especially among small holders as a main livelihood activity mainly to produce meat. However, the level of production from local goats is generally low and this is primarily due to poor feeding practices especially during dry periods. In the traditional feeding systems non legume and legume foliages of trees and shrubs are the main feed resources for goats (Seresinhe and Marapana, 2011). Leguminous tree foliage is potential source of protein and minerals and could be employed as supplements to non legumes to increase the level of production. However, the presence of tannins in both legumes and non-legumes limits utilization of both species as they can reduce the feed intake, nutrient digestibility and protein availability (Silanikove et al., 2001). Nevertheless, some tanniniferous feeds have beneficial effects in ruminant diets by improving nitrogen utilization efficiency and amino acid absorption. Condensed tannins also have biological effects on the control of gastrointestinal parasites; possible direct effects could be mediated through CT-nematode interactions, which reduce nematode viability (Nguyen et al., 2005).
A management strategy to reduce negative effects of tannins in fodder trees could be to feed mixtures of low and high tannin content species, which could create positive effects on in vitro gas production, rumen degradation and digestibility of diets (Castro-Gonzáles and Alayon-Gamboa, 2008). A better understanding of the effects of low and high tannin foliage mixtures on nutrient digestibility and methane mitigation properties would improve management of such resources. This knowledge would be of considerable importance to Sri Lanka for the efficient utilization of tree forage and research must be established to develop feeding strategies to overcome undesirable effects when using tanniniferous foliage. This study evaluated the suitability of several mixtures of high tanniniferous non legume foliage mixed with low tanniniferous legume foliage on in vitro gas production and rumen degradability characteristics.
MATERIALS AND METHODS
Forages used and proximate analysis
Edible forage samples (leaves and tender stems) from plant species given in Table 1 were hand harvested. Standard methods as described by AOAC (1990) were used for determination of dry matter, ash and crude protein. Fiber components (neutral detergent fiber, NDF; acid detergent fiber, ADF) were determined by methods of Van Soest (1967). Acid detergent residue was treated with 72% H2SO4 for lignin estimation.
Table 1.
Leguminous and non-leguminous forage species used for the experiment
| Family | Common name | Botanical name |
|---|---|---|
| Rhizophoraceae | Dawata | Carallia integerrima |
| Euphorbiaceae | Kebella | Aporosa lindeliyana |
| Boraginaceae | Imbul | Ceiba perntandra |
| Moraceae | Jak fruit | Artocarpus heterophyllus |
| Leguminoseae | Ipil ipil | Leucaena leucocephala |
| Leguminoseae | Glirizidia | Gliricidia sepium |
Analyses of tannins
Tannins were analyzed by first weighing of 200 mg of feed into a 50 ml conical flask. The feed sample was extracted with 70% aqueous acetone in an ultrasonic bath for 2 h and the contents were centrifuged for 20 min at 5,000×g and the supernatant was collected for tannin analyses.
Total phenols were estimated by the Folin-Ciocalteu reaction (Makkar, 2003). For the condensed tannin (CT) fraction, the extract was treated with Butanol-HCl in the presence of ferric ammonium sulphate, and CT expressed as leucocyanidin equivalent as
Where A550nm is absorbance at 500 nm assuming that the effective E 1 cm, 550 nm of leucocyanidin is 460 (Porter et al., 1986).
Insoluble polyvinylpolypyrrolidone (PVPP; 100 mg) was weighed into 100 mm×12 mm test tubes. Distilled water, 1 ml, and then 1 ml tannin containing extract were added and vortexed. The tube was kept at 4°C for 15 min, vortexed again, then centrifuged (3,000×g) for 10 min and the supernatant collected. The phenolic content of the supernatant was measured by Folin-Ciocalteau reaction and this was regarded as the non tannin phenol (NTP).
Total tannin phenols (TTP) were calculated as the difference of TP and NTP. Hydrolysable tannins (HT) were calculated as the difference between TTP and CT.
Experimental design
Eight treatments were examined in a randomized complete block design using four non legumes with high tannins and two shrub legumes with low tannins at a ratio of 3:1. Treatment combinations of high tannin non-legume and low tannin legumes used for the experiment are given in Table 2.
Table 2.
Treatment combinations of high tannin non-legume and low tannin legumes used for the experiment
| Treatment | Combination |
|---|---|
| Treatment 1 | C. integerrima+L. leucocephala |
| Treatment 2 | C. integerrima+G. sepium |
| Treatment 3 | A.lindeliyana+L. leucocephala |
| Treatment 4 | A. lindeliyana+G. sepium |
| Treatment 5 | C. perntandra+L. leucocephala |
| Treatment 6 | C. perntandra+G. sepium |
| Treatment 7 | A. heterophyllus+L. leucocephala |
| Treatment 8 | A. heterophyllus+G. sepium |
In vitro gas production
In vitro gas production was determined as described by Menke and Steingass (1988). Rumen fluid was collected before feeding in the morning from two fistulated donor bulls at the experimental farm of the Faculty of Agriculture. Rumen fluid was strained through four layers of gauze into a pre-warmed, insulated bottle. All laboratory handling of rumen fluid was carried out under a continuous flow of CO2.
Samples (200 mg) consisting of 150 mg high tannin non-legume+50 mg low tannin legume) of the oven-dry feedstuffs were accurately weighed into 100-ml glass syringes fitted with plungers. Syringes were filled with 30 ml of medium consisting of 10 ml of rumen fluid and 20 ml of buffer solution as described by Menke and Steingass (1988). Two blank samples containing 30 ml of medium only were included. The syringes were placed in an incubator (39°C) and the syringes rotated during first 4 h. Gas production was recorded after 4, 8, 12, 24 and 48 h of incubation. In all experiments, each incubation was repeated on three different days so that each treatment was conducted in triplicate.
In vitro dry matter digestibility
At the end of the fermentation period, the fermented residues were filtered into pre-weighed filter dried for 24 h at 105°C and weighed and in vitro dry matter digestibility (IVDMD) was calculated using the standard formula.
Ammonia production
Ammonium concentration in fermentation liquid was determined using Kjeldhal method. Only distillation and titration steps were followed.
Methane production
Methane (CH4) was analyzed in the Dept of Animal Science laboratory of ETH, Zurich using Hewlett Packard Gas Chromatograph (Model 5890, Series II, Avondale, PA, USA).
Protozoa and bacteria counts
Protozoal and bacterial counts were counted with Bürker counting chambers (0.1 and 0.02 mm depth, respectively; Blau Brandw, Wertheim, Germany).
Statistical analysis
Analysis of variance (ANOVA) was performed on Chemical composition, in vitro digestibility and gas production data. The statistical significance of the differences between means was tested using the Duncan Multiple Range Test (DMRT). Correlation coefficients were calculated using MS EXCEL version 2007.
RESULTS AND DISCUSSION
Nutritive value of forages, in vitro gas production and dry matter degradability
The chemical composition of forages is presented in Table 3; the entire legume×non legume mixtures used in the current study had a CP content more than 14% as confirmed by Seresinhe et al. (2003). We observed that the CP content of both leguminous species used in this study was significantly higher than that of all non leguminous species (Table 3). However, in this study the low level (= 8%) of CP in non-legume species could be compensated by combination of legume species with high (= 20%) CP content as evident from NH3-N production (Table 5). It was shown that non leguminous species, C. perntandra and A. heterophyllus had higher (p<0.05) CP comparable with other non-legumes (Table 5). NH3-N concentrations in the present study ranged from 5.66 to 9.13 ml/200 mg DM (p>0.05). NH3 concentration is balanced between degradation of feed protein and uptake of ammonia for synthesis of microbial protein. Although not significant, the higher NH3-N concentration observed in A. heterophyllus +L. leucocephala (Trt7) is likely due to higher CP content.
Table 3.
Proximate composition of forages
| Botanical name | DM % | CP % | CT % | NDF % | ADF % | ADL % |
|---|---|---|---|---|---|---|
| Non-legumes | ||||||
| Carallia integerrima (Dawata) | 32.1a±1.02 | 8.0c±0.97 | 6.2a±0.35 | 56.7a±4.31 | 47.0b±3.21 | 25.8a±3.25 |
| Aporosa lindeliyana (Kebella) | 29.9b±0.78 | 9.0c±0.78 | 6.4b±0.01 | 50.2ab±3.18 | 51.3a±1.70 | 27.2a±1.48 |
| Ceiba perntandra (Imbul) | 27.7b±0.81 | 17.6b±0.98 | 4.9b±0.06 | 46.1b±6.43 | 33.8d±0.71 | 15.8c±0.78 |
| Artocarpus heterophyllus (Jak fruit) | 30.0ab±0.97 | 15.9b±0.07 | 5.4a±0.32 | 46.5b±1.77 | 43.8c±1.77 | 18.8b±0.49 |
| Legumes | ||||||
| Leucaena leucocephala (Ipil ipil) | 31.0a±0.85 | 26.8a±40.56 | 1.6 LT±0.06 | 33.5c±0.636 | 23.6e±0.71 | 8.5cd±0.35 |
| Gliricidia sepium (Glirizidia) | 26.0b±1.98 | 20.0a b±1.33 | 0.8 LTd±0.21 | 35.1c±0.98 | 35.9d±0.72 | 5.1d±0.55 |
| Sig. | *** | * | * | *** | *** | *** |
Data are mean values of four replicates. DM = Dry matter; CP = Crude protein; CT = Condensed tannin; NDF = Neutral detergent fiber; ADF = Acid detergent fiber; ADL = Acid detergent lignin. Means within the same column with differing superscripts (a, b, c and d) are significantly different (* p<0.05, *** p<0.001).
Table 5.
In vitro gas production (IVGP), ammonia production (%) and dry matter degradability (IVDMD %) of forage combinations
| Treatment | IVDMD % | Mean gas production (ml/200 mg DM)**** | NH3 -N (ml/200 mg DM)**** |
|---|---|---|---|
| Trt1: C. integerrima+L. leucocephala | 47.8 b±7.98 | 37.5bc±3.32 | 7.4 ±0.99 |
| Trt2: C. integerrima+G. sepium | 49.9 b±5.43 | 38.5bc±4.93 | 5.7±0.76 |
| Trt 3: A. lindeliyana+L. leucocephala | 38.3 b±4.67 | 34.5 c±3.87 | 5.9±0.72 |
| Trt 4: A. lindeliyana+G. sepium | 38.4 b±7.79 | 36.0 c±2.00 | 7.9±0.68 |
| Trt 5: C. perntandra+L. leucocephala | 46.6 b±5.25 | 41.3 ab ±4.57 | 8.3±0.06 |
| Trt 6: C. perntandra+G. sepium | 44.5 b±3.70 | 44.0 a±1.83 | 8.9±0.473 |
| Trt7: A. heterophyllus+L. leucocephala | 56.3 a±22.9 | 44.0 a±1.83 | 9.1±0.35 |
| Trt8: A. heterophyllus+G. sepium | 49.9 b±5.27 | 43.0 a±3.16 | 8.5±0.60 |
| Sig. | * | ** | NS |
Means in the same column with differing superscripts (a, b and c) are significant in IVDMD %, Mean gas production and NH3 -N.
p<0.05,
p<0.01. NS = Not significant. Data are mean values of three replicates (twenty four samples).
Amount of gas produced in ml as a result of incubation of 200 mg substrate of dry matter.
In contrast, non legume foliage contains significantly higher NDF, ADF and ADL contents as compared with leguminous foliage (Table 3). Among non legumes, C. integerrima and A. lindeliyana contained higher (p<0.05) NDF, ADF and ADL as compared with Ceiba perntandra and Artocarpus heterophyllus. Combinations of C. perntandra×LL and A. heterophyllus×LL mixtures contained higher (p<0.05) CP as compared with the same combinations with GS (Trt 6, Trt 8). The CP contents of mixtures of C. integerrima×LL and A. lindeliyana×LL (Trt 1, Trt 3) were also higher as compared with the same combinations with GS. Supplementation with tree foliage rich in CP and low in NDF, ADF and ADL aimed to compensate the limitations in nutrients in non leguminous species.
The condensed tannin content of non-legume and legume combinations ranged between 2.84 to 3.99 (Table 4). The beneficial effects of forage mixtures containing low levels of tannins could be due to the protection of proteins from microbial degradation thus increasing the amount of undegraded protein entering the small intestine (Barry et al., 1986). However, higher concentrations of tannins in the diet are associated with the reduction in organic matter digestibility. Feedstuffs that are inherent in certain anti-nutritive factors had been reported to be low in organic matter digestibility (Aregheore and Abdulrazak, 2005).
Table 4.
Proximate composition of forage mixtures
| Botanical name | DM % | CP % | Condensed CT % | NDF % | ADF % | ADL % |
|---|---|---|---|---|---|---|
| Carallia integerrima ×Leucaena leucocephala (Trt 1) | 31.5a±1.10 | 17.4c±0.89 | 3.9a±0.41 | 45.1a±3.67 | 35.3b±1.02 | 18.6a±2.67 |
| Carallia integerrima ×Gliricidia sepium (Trt 2) | 29.1a±0.98 | 14.0d±0.89 | 3.5a±0.32 | 39.8c±2.98 | 33.7c±2.05 | 15.5b v2.13 |
| Aporosa lindeliyana ×Leucaena leucocephala (Trt. 3) | 30.5a±0.86 | 17.9c±0.77 | 4.0a±0.40 | 39.8c±3.01 | 33.7c±2.78 | 17.8a±2.11 |
| Aporosa lindeliyana ×Gliricidia sepium (Trt 4) | 28.0a±0.89 | 14.5d±0.88 | 3.6a±0.35 | 40.6b±2.67 | 39.9ab±2.34 | 16.1ab±1.98 |
| Ceiba perntandra ×Leucaena leucocephala (Trt. 5) | 29.4a±0.97 | 22.2a±0.98 | 3.3b±0.33 | 41.8b±2.89 | 28.7d±2.56 | 12.1b±1.87 |
| Ceiba perntandra ×Gliricidia sepium (Trt. 6) | 26.9a±0.99 | 18.8b±0.77 | 2.8c±0.37 | 42.6ab±2.87 | 34.9bc±2.04 | 10.5d±1.09 |
| Artocarpus heterophyllus ×Leucaena leucocephala (Trt. 7) | 30.5a±0.86 | 21.4ab±0.86 | 3.5a±0.22 | 40.0c±2.67 | 37.5b±2.45 | 16.1ab±1.25 |
| Artocarpus heterophyllus ×Gliricidia sepium (Trt. 8) | 28.0a±0.79 | 18.0bc±0.95 | 3.1ab±0.31 | 40.8bc±2.97 | 43.6a±2.01 | 14.4b±1.45 |
| Sig. | * | * | * | * | *** | *** |
Data are mean values of four replicates. DM = Dry matter; CP = Crude protein; CT = Condensed tannin; NDF = Neutral detergent fiber; ADF = Acid detergent fiber ADF; ADL = Acid detergent lignin. Means within the same column with differing superscripts (a, b, c and d) are significantly different (* p<0.05, *** p<0.001).
There was a steady increase in the gas production for over a period of 48 h as well as significant differences between forage mixtures in net gas volume (Table 5). The highest net gas production was observed in C. perntandra+GS (Trt 6) and A. heterophyllus+LL (Trt 7) but not significantly different from either with C. perntandra+LL (Trt 5) or A. heterophyllus+GS (Trt 8). However, net gas production of C. integerrima and A. lindeliyana either with LL (Trt 1, Trt 2) or GS (Trt 3, Trt 4) mixtures were lower but not significant from each other. There are many factors that may determine the amount of gas produced during fermentation including the nature and level of fiber, the presence of secondary metabolites (Babayemi et al., 2004) and potency of the rumen liquor for incubation. It is possible to attain the potential gas production of feedstuffs if the donor animal from which rumen liquor is collected for incubation has met it’s nutrient requirements. Generally, gas production is a function of and a mirror of degradable carbohydrate therefore, the amount depends on the nature of carbohydrates (Blummel and Becker, 1997).
The correlation (R2) between in vitro gas production after 48 h incubation and condensed tannin content was 0.67. More than 60% variation in the in vitro gas production on incubation was explained by condensed tannins. The findings are consistent with Njidda and Ikhimioyza (2010), Tolera et al. (1997) and Getachew et al. (2002) who found strong correlations between CT and gas production. The results also suggest the relationship (R2 = 0.61) of CP content and in vitro gas production is also high (Figure 1). It has been reported that the high content of CP and low content of condensed tannin (CT) is associated with degradability of feed, resulting in the higher values for the potential gas production (Ahmed et al., 2007). Further they reported that a negative correlation of potential gas production with ADF and CT may be due to the reduction of microbial activity from increasingly adverse environmental conditions.
Figure 1.
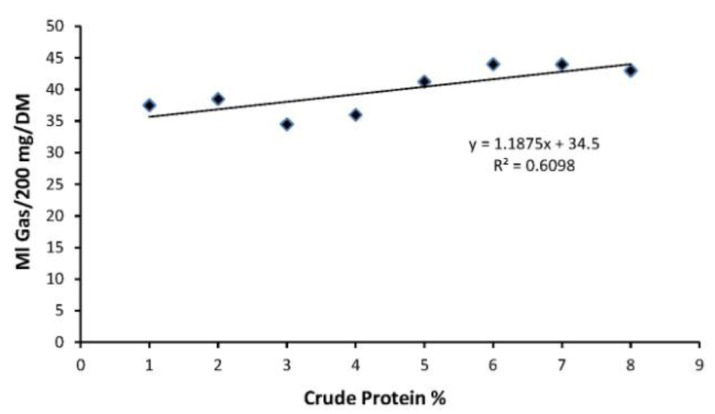
Relationship between in vitro gas production and crude protein percentage in forage combinations.
The results of the IVDMD are shown in Table 5. Highest (p<0.05) IVDMD was observed in Trt 7. The IVDMD of other treatments were not different among each other. In tree leaves, tannins are present in the NDF and ADF fractions and are tightly bound to the cell wall and cell protein and seem to be involved in decreasing digestibility (Reed et al., 1990). According to the present results it could be suggested that feeds containing high levels of CP and low levels of tannins could generate more methane in the rumen. Soliva et al. (2008) indicated that plants known to contain plant secondary metabolites e.g. tannins are able to suppress methanogenesis. Work done by Balogun et al. (1998), Seresinhe and Iben (2003) and Ammar et al. (2004) pointed out that there were significant (p<0.001) negative correlations between IVDMD and cell wall constituents (ADF and NDF) and also between IVDMD and CT (Kamalak, 2005). Moreover, Seresinhe and Iben (2003) reported the existence of a correlation between IVDMD and CP content which was further confirmed by Kamalak et al. (2005). Chenost et al. (2001) further confirmed the inherent direct relationship between CP and digestibility using seventy nine different forages with four replicates.
Methane production, bacteria and protozoa counts
Methane production (ml/200 mg DM) (Table 6) ranged from 2.57 to 4.79 ml among forage mixtures the least and highest being from A. lindeliyana+LL (Trt 4) to C. perntandra+LL (Trt 5). Also it is evident that the treatments show a high capacity for total gas production to be synonimous for high methane production. Methane production indicates an enenrgy loss to ruminants and many tropical feedstuffs have been implecated with increased methonogenensis. Carlos and Lascano (2003) confirmed that the inclusion of tannin-rich legumes such as Calliandra calothyrsus and Flemingia macrophylla in forage-based diets significantly reduced methane release but also negatively affected nutrient degradation and N turnover. However, in the present study the tannin content was reduced due to combining effect of high and low tannin forages but a more or less similar range of methane production was observed as compared with semi arid browses like Ficus polita and F. thonningii in Nigeria (Njidd and Ikhimioya, 2010). Carlos and Lascano (2003) further hypothesized that to take advantage of the methane suppressing effect of tannin-rich legumes without affecting nutrient degradation and N turnover it was necessary to combine them with legumes low or free of tannin. He further suggested that future work should concentrate in defining the optimal type and proportion of tannin rich legumes in these mixtures as we did in the present study. Further, Tjahyono and Santoso (2010) reported that the positive or negative effects of tannin on methane production may vary depending on the amount of tannin in the plant. Highest (p<0.05) bacteria population was found in C. perntandra+GS (Trt 6) followed by A. heterophyllus+LL (Trt 7) as compared with other treatments. Protozoa populations ranged between 20 and 55×103 ml. Although not significant, same trend as in the bacteria population was observed as well. The protozoa populations found in this study were in similar ranges as observed by Hariadi and Santoso (2009). The results confirm that lowest level of tannins in Trt 6 could increase the protozoa and bacteria populations in the fermentation liquid support the results of Hess et al. (2003) who reported that tannins may cause significant shifts in rumen microbial populations.
Table 6.
Methane production, bacteria and protozoa counts after 48 h (incubation)
| Treatment | CH4 production (ml/200 mg DM)**** | Bacteriax103 (Ml−1) | Protozoax103 (Ml−1) |
|---|---|---|---|
| Trt1: C. integerrima+L. leucocephala | 3.3±0.12 | 225b±52.50 | 35.0±3.52 |
| Trt2: C. integerrima+G. sepium | 3.7±0.33 | 150c±39.68 | 20.0±1.16 |
| Trt 3: A. lindeliyana+L. leucocephala | 2.6±0.18 | 175 b±67.22 | 40.0±5.57 |
| Trt 4: A. lindeliyana+G. sepium | 2.6±0.09 | 240b ±22.00 | 30.0±1.73 |
| Trt 5: C. perntandra+L. leucocephala | 4.8±0.09 | 155c±99.21 | 40.0±6.66 |
| Trt 6: C. perntandra+G. sepium | 4.2±0.11 | 300a ±17.81 | 55.0±1.15 |
| Trt7: A. heterophyllus+L. leucocephala | 4.5±0.38 | 272a ±26.33 | 48.0±3.46 |
| Trt8: A.heterophyllus+G. sepium | 3.8±0.16 | 217 bc±13.12 | 40.0±4.58 |
| Sig. | NS | * | NS |
Means in the same column with differing superscripts (a, b and c) are significant in Bacteria count.
* p<0.05, ** p<0.01, NS = Not significant. Data are mean values of three replicates (twenty four samples).
Supplementing high tannin non leguminous foliage by incremental substitution of L. leucocephala and G. sepium foliage increased gas production parameters, IVDMD, and microbial populations in the fermentation liquid due to reduction in tannins as well as an increase in CP content of non-legume and legume combinations. Methane production was low in all treatments but no significant effect was found due to the presence of tannins or higher levels of CP.
ACKNOWLEDGEMENTS
Authors greatly acknowledge the financial and laboratory assistance and guidance given by Prof. Michael Kreuzer and Dr. Carla Soliva in the Department of Agricultural and Food Science, ETH Zurich, Switzerland. Authors also appreciate the assistance of Messieurs D. S. Wijewardhana, S. Karunathilaka and M. P. Kapila Premakumara.
REFERENCES
- AOAC. Official methods of analysis. 15th ed. Association of Official Analytical Chemists; Arlington, VA, USA: 1990. [Google Scholar]
- Ahmed GN, Abdel Nasir MA, Fadel E. Chemical composition and in vitro gas production characteristics of six fodder trees leaves and seeds. J Agric Biol Sci. 2007;3:983–986. [Google Scholar]
- Ammar H, Lopez S, Gonzalez JS, Ranilla M. Comparison between Analytical methods and biological assays for the assessment of tannin-related antinutritive effects in some Spanish browse species. J Sci Food Agric. 2004;84:1349–1356. [Google Scholar]
- Aregheore EM, Abdulrazak SA. Estimation of organic matter digestibility and metabolizable energy content of agro-industrial wastes using in vitro gas production Nigerian. J Anim Prod. 2005;32:79–87. [Google Scholar]
- Babayemi OJ, Demeyer D, Fievez V. Nutritive value and qualitative assessment of secondary compounds in seeds of eight tropical browse, shrub and pulse legumes. Commun Agric Appl Biol Sci. 2004;69:103–110. [PubMed] [Google Scholar]
- Balogun RO, Jones RJ, Holmes JHG. Digestibility of some tropical browse species varying in tannin content. Anim Feed Sci Technol. 1998;76:77–88. [Google Scholar]
- Barry TN, Manley TR, Duncan SJ. The role of condensed tannins in the nutritive value of Lotus peduculatus for sheep 4 sites of carbohydrate and protein digestion as influenced by dietary reactive tannin concentration. Br J Nutr. 1986;55:123–137. doi: 10.1079/bjn19860016. [DOI] [PubMed] [Google Scholar]
- Blummel M, Becker K. The degradability characteristics of fifty-four roughages and roughage neutral detergent fibers as described by in vitro gas production and the relationship to voluntary feed intake. Br J Nutr. 1997;77:757–768. doi: 10.1079/bjn19970073. [DOI] [PubMed] [Google Scholar]
- Carlos E, Lascano . Tropical grasses and legumes: Optimizing genetic diversity for multipurpose use (Project IP5) Centro Internacional de Agricultura Tropical (CIAT); 2003. [Google Scholar]
- Castro-Gonzáles A, Alayon-Gamboa JA. Affects of Brosimum alicastrum and Lysolima latisiliquum mixtures on voluntary intake, nutrient digestibility and nitrogen balance in sheep fed tropical pastures. Anim Feed Sci Technol. 2008;141:246–258. [Google Scholar]
- Chenost M, Aufrère J, Macheboeuf D. The gas-test technique as a tool for predicting the energetic value of forage plants. Anim Res. 2001;50:349–364. [Google Scholar]
- Getachew G, Makkar HPS, Becker K. Tropical browses: Contents of phenolic compounds in vitro gas production and stoichiometric relationship between short chain Fatty acid and in vitro gas production. J Agric Sci. 2002;139:341–352. [Google Scholar]
- Hariyadi BT, Santoso B. Evaluation of tropical plants containing tannin on in vitro methanogenesis and fermentation parameters using rumen fluid. J Sci Food Agric. 2010;90:456–461. doi: 10.1002/jsfa.3839. [DOI] [PubMed] [Google Scholar]
- Hess HD, Monsalve lM, Lascano CE, Carulla CE, Diaz TE, Kreuzer M. Supplementation of a tropical grass diet with forage legumes and Sapindus saponaria fruits: effects of in vitro ruminal nitrogen turnover and methanogenensis. Aust J Agric Res. 2003;54:703–713. [Google Scholar]
- Kamalak A. Chemical composition and in vitro dry matter digestibility of leaves of. Vitis Vinifera Livest Res Rural Dev. 2005;17:68. [Google Scholar]
- Kamalak A, Canbolat O, Gurbuz Y, Ozzy O, Ozkose CO. Chemical composition and its relationship to in vitro gas production of several tannin-containing trees and shrub leaves. Asian-Aust J Anim Sci. 2005;18:203–208. [Google Scholar]
- Makkar HPS. Quantification of tannins in tree and shrub foliage. A Laboratory Manual Kluwer Academic Publishers; Amsterdam, The Netherlands: 2003. [Google Scholar]
- Menke KH, Steingass H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim Res Dev. 1988;28:7–55. [Google Scholar]
- Nguyen TM, Van Binh D, Ørskov ER. Effect of foliages containing condensed tannins and gastrointestinal parasites. Anim Feed Sci Technol. 2005;122:77–87. [Google Scholar]
- Njidda AA, Ikhimioya I. In vitro gas production and dry matter digestibility of semi-arid browses of north eastern Nigeria. Slovak J Anim Sci. 2010;43:154–159. [Google Scholar]
- Porter IP, Hrstich IN, Chan BG. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry. 1986;25:223–230. [Google Scholar]
- Reed JD, Soller H, Woodwar A. Fodder tree and straw diets for sheep: intake, growth, digestibility and the effects of phenolic on nitrogen utilization. Anim Feed Sci Technol. 1990;30:39–50. [Google Scholar]
- Seresinhe T, Marapana RAUJ. Goat farming systems in the southern province of Sri Lanka: Feeding and management strategies. World J Agric Sci. 2011;7:383–390. [Google Scholar]
- Seresinhe YDJW, Seresinhe T, Pathirana KK. Proceedings of the second academic session. University of Ruhuna; Matara, Sri Lanka: 2003. Jan 23, Assessment of the feeding value of five shrub legumes of local interest; pp. 43–48. [Google Scholar]
- Seresinhe T, Iben C. In vitro quality assessment of two tropical shrub legumes in relation to their extractable tannins content. J Anim Physiol Anim Nutr. 2003;87:109–115. doi: 10.1046/j.1439-0396.2003.00422.x. [DOI] [PubMed] [Google Scholar]
- Silanikove N, Gilboa N, Nitsan Z. Effects of condensed tannins in carob leaves (Ceratonia siliqua) on rumen volume and passage rate of liquid and particulate matter along the digestive tract in goats. Small Rumin Res. 2001;40:95–99. doi: 10.1016/s0921-4488(00)00209-1. [DOI] [PubMed] [Google Scholar]
- Soliva CR, Zeleke AB, Clement C, Hess HD, Fievez V, Kreuzer M. In vitro screening of various tropical foliages, seeds, fruits and medicinal plants for low methane and high ammonia generating potentials in the rumen. Anim Feed Sci Technol. 2008;147:53–71. [Google Scholar]
- Tjahyono B, Santoso HB. Evaluation of tropical plants containing tannin on in vitro methanogenesis and fermentation parameters using rumen fluid. J Sci Food Agric. 2010;90:456–461. doi: 10.1002/jsfa.3839. [DOI] [PubMed] [Google Scholar]
- Tolera A, Khazaal K, Ørskov ER. Nutritive evaluation of some browse species. Anim Feed Sci Technol. 1997;67:181–195. [Google Scholar]
- Van Soest PJ. Development of a comprehensive system of feed analysis and its application to forages. J Anim Sci. 1967;26:119–128. [Google Scholar]