

Abstract
Mitochondrial encephalomyopathy, lactic acidosis, and stroke-like episodes (MELAS) syndrome is one of the most common mitochondrial disorders. Although the pathogenesis of stroke-like episodes remains unclear, it has been suggested that mitochondrial proliferation may result in endothelial dysfunction and decreased nitric oxide (NO) availability leading to cerebral ischemic events. This study aimed to assess NO production in subjects with MELAS syndrome and the effect of the NO precursors arginine and citrulline. Using stable isotope infusion techniques, we assessed arginine, citrulline, and NO metabolism in control subjects and subjects with MELAS syndrome before and after arginine or citrulline supplementation. The results showed that subjects with MELAS had lower NO synthesis rate associated with reduced citrulline flux, de novo arginine synthesis rate, and plasma arginine and citrulline concentrations, and higher plasma asymmetric dimethylarginine (ADMA) concentration and arginine clearance. We conclude that the observed impaired NO production is due to multiple factors including elevated ADMA, higher arginine clearance, and, most importantly, decreased de novo arginine synthesis secondary to decreased citrulline availability. Arginine and, to a greater extent, citrulline supplementation increased the de novo arginine synthesis rate, the plasma concentrations and flux of arginine and citrulline, and NO production. De novo arginine synthesis increased markedly with citrulline supplementation, explaining the superior efficacy of citrulline in increasing NO production. The improvement in NO production with arginine or citrulline supplementation supports their use in MELAS and suggests that citrulline may have a better therapeutic effect than arginine. These findings can have a broader relevance for other disorders marked by perturbations in NO metabolism.
Keywords: Stroke, Endothelial dysfunction, Nitric oxide synthase, Mitochondrial diseases, Stable isotope, Clinical research
1. Introduction
Mitochondrial encephalomyopathy, lactic acidosis, and stroke-like episodes (MELAS) syndrome is one of the most common mitochondrial disorders, with an estimated prevalence of 60:100,000 [1]. MELAS syndrome is primarily caused by mutations in mitochondrial DNA, with the most frequent being the m.3243A>G mutation in the MTTL1 gene that encodes tRNALeu/(UUR) [2]. It has a broad spectrum of manifestations, including stroke-like episodes, exercise intolerance, muscle weakness, epilepsy, dementia, migraine headaches, short stature, sensorineural hearing loss, lactic acidosis, and diabetes [3]. MELAS syndrome typically affects a young population, with 75% of cases presenting before 20 years of age, and causes significant morbidity, with the life-time prevalence of stroke-like episodes approaching 99% [3]. However, no specific consensus approach to its treatment has been established [4].
When stroke-like episodes occur in MELAS syndrome, the affected areas do not correspond to classical vascular territories but rather have an irregular distribution, suggesting that events reflect a small-vessel etiology [5]. The pathogenesis of these stroke-like episodes is not clear; however, it has been suggested that these events are due to ischemic insults, based upon SPECT (single photon emission computed tomography) scanning studies showing hypoperfusion in the affected regions during early stages of stroke-like episodes [6].
Energy depletion due to mitochondrial dysfunction in MELAS syndrome can stimulate mitochondrial proliferation in various tissues, including vascular endothelial cells [7,8]. It has been suggested that endothelial dysfunction, resulting from mitochondrial proliferation, can lead to impaired cerebral blood flow and stroke-like episodes in MELAS syndrome [5,9]. One of the functions of the vascular endothelium is to produce nitric oxide (NO), which, amongst a host of properties, plays a major role in smooth muscle relaxation that is needed to maintain the patency of small blood vessels [10,11]. Thus, endothelial dysfunction may result in impaired NO production. Furthermore, mitochondrial proliferation in endothelial cells is associated with increased cytochrome c oxidase (COX) activity, which can bind to and sequester NO. Thus, decreased NO availability, either by impaired production or sequestration by COX, can potentially play a major role in the mechanism of stroke-like episodes observed in MELAS syndrome [5,9]. This hypothesis is supported by the observation of lower NO metabolite concentrations during stroke-like episodes in subjects with MELAS syndrome [6].
NO is formed from arginine via the enzyme nitric oxide synthase (NOS), which catalyzes the conversion of arginine to citrulline. The NO is rapidly oxidized to nitrite and nitrate, which are more stable molecules. Citrulline is a nonessential amino acid for which the main source is the de novo synthesis in small intestine enterocytes through a number of mitochondrial enzymes. Citrulline can be converted to arginine via argininosuccinate synthase (ASS) and argininosuccinate lyase (ASL). Arginine is derived from the diet, protein breakdown, and the de novo synthesis from citrulline, the latter constituting 5–15% of arginine production (Fig. 1) [12–14]. Along with arginine availability, NO synthesis is also regulated by asymmetric dimethylarginine (ADMA) which is an endogenous inhibitor of NOS. ADMA is generated by the methylation of arginine residues in proteins and subsequently released by protein hydrolysis within cells to be transported in plasma. ADMA enters cells by the same cationic amino acid transporters (CATs) used by arginine, and thus can also limit intracellular arginine availability [15]. Elevated ADMA levels can potentially contribute to endothelial dysfunction in a number of human diseases, including chronic renal disease, hypertension, and diabetes [16–20].
Fig. 1.
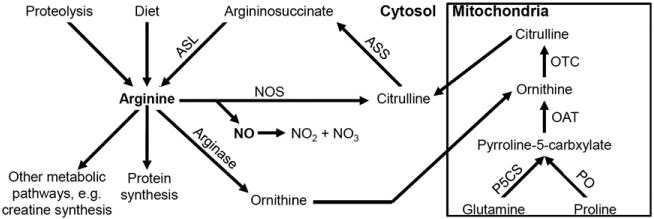
Schematic presentation of arginine, citrulline, and NO metabolic pathways (ASS: argininosuccinate synthase, ASL: argininosuccinate lyase, NOS: nitric oxide synthase, PO: proline oxidase, P5CS: pyrroline 5 carboxylate synthase, OAT: ornithine aminotransferase, OCT: ornithine transcarbamylase).
Both arginine and citrulline potentially act as NO precursors, and it has been proposed that their administration may result in increased NO availability and hence can have therapeutic effects in stroke-like episodes in MELAS syndrome [9]. This hypothesis is supported by the demonstration that oral and intravenous arginine administration to subjects with MELAS syndrome led to improvement in the clinical symptoms associated with stroke-like episodes and a decrease in the frequency and severity of these episodes [6,21] No previous clinical studies have been performed to assess NO production and the effect of arginine on NO production in MELAS syndrome. Likewise, no studies have been conducted to evaluate the effect of citrulline, which can theoretically be a more effective NO precursor than arginine, as arginase may compete with NOS for the intracellular arginine pool. In addition, there is evidence that ASS and ASL co-localize and are co-induced with inducible NOS (iNOS) in various cell types [22]. Such co-localization can result in substrate channeling of citrulline to more efficiently produce NO in certain sub-cellular compartments.
We hypothesized that subjects with MELAS syndrome have impaired NO production, administration of L-arginine or L-citrulline would improve NO production, and L-citrulline would improve NO production to a higher degree than L-arginine supplementation. To test these hypotheses we measured whole body NO synthesis rates, among other variables, in control subjects and subjects with MELAS syndrome before and after arginine or citrulline oral supplementation. In this study, we have used well-established stable isotope infusion protocols to provide a quantitative assessment of whole body in vivo arginine, citrulline, and NO metabolism. Such methods have not been previously employed in this syndrome or other mitochondrial diseases. Therefore, this is the first study to assess the in vivo NO metabolism in subjects with MELAS syndrome.
2. Subjects and methods
2.1. Subjects
Ten subjects with MELAS syndrome and ten control subjects participated in this study. Subjects with MELAS syndrome were adults aged 18–57 years, diagnosed clinically with MELAS syndrome and harbored the m.3243A>G mutation in the MTTL1 gene (Supplementary Table 1). Control subjects were healthy adults aged 20–46 years. We aimed to obtain a control cohort that was age-, gender, and BMI-matched to the cohort of subjects with MELAS. The study was approved by the Institutional Review Board (IRB) at Baylor College of Medicine (BCM) and the General Clinical Research Center (GCRC) Scientific Advisory Committee (SAC) at Texas Children’s Hospital (TCH).
2.2. Study design
Subjects with MELAS syndrome were admitted twice to the GCRC at TCH. During the hospital stay, they were placed on a low-nitrite and nitrate, protein-controlled diet containing 0.8 g·kg−1·d−1 protein. After 48 h on the diet, the first stable isotope infusion was administered as described below. Following the first infusion, oral L-arginine supplementation was initiated at a dose of 10 g·m2 body surface area−1·d−1 divided every 4 h for 48 h. A second stable isotope infusion was then performed. After an interval of at least 1 week, the subjects with MELAS syndrome were admitted for a second time, where a similar procedure was conducted, but rather than L-arginine, L-citrulline was given at the same dose and frequency. Control subjects were admitted once and had one stable isotope infusion after being on the controlled diet for 48 h (Fig. 2).
Fig. 2.
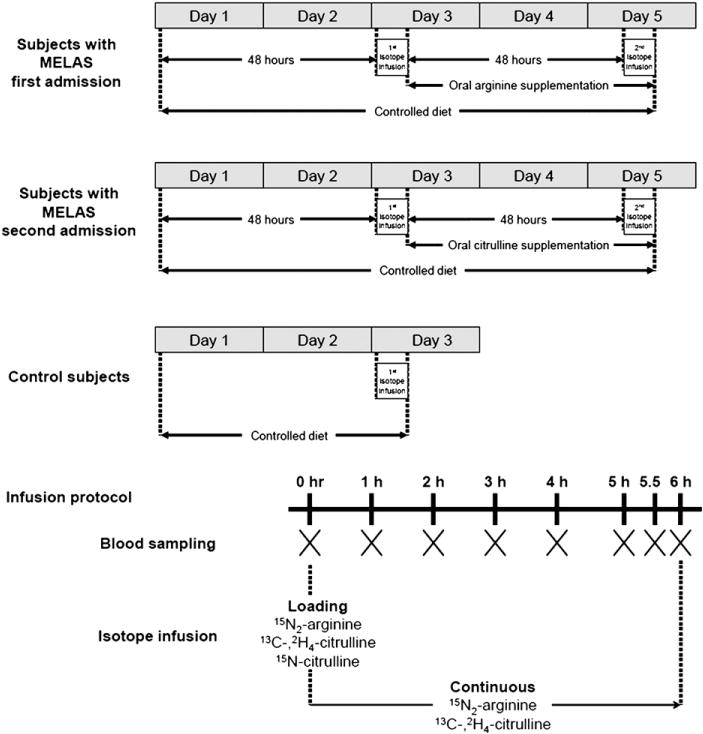
Study design for subjects with MELAS syndrome during the first and second admission and for control subjects, and the infusion protocol. Subjects with MELAS syndrome were admitted twice to the GCRC at TCH. The flux of different amino acids can be affected by dietary protein intake; therefore, research subjects were placed on a protein-controlled diet containing 0.8 g·kg−1·d−1 protein. To ensure a steady state protein metabolism, the first stable isotope infusion was administered after 48 h of the initiation of this diet. Following the first infusion, oral L-arginine supplementation was initiated at a dose of 10 g·m2 body surface area−1·d−1 divided every 4 h for 48 h to achieve a steady state. A second stable isotope infusion was then performed. The controlled diet was also low in nitrate and nitrite to limit the effect of enteral intake on plasma nitrite and nitrate measurements. With each stable isotope infusion, research subjects received four small meals composed of the complete nutritional supplement Ensure (Abbott Nutrition, Columbus, Ohio), given every 2 h starting 2 h before the initiation of the stable isotope infusion. The amount of the nutritional supplement contained one third of the daily intake of protein divided in four equal portions. The remaining two thirds of the protein intake were consumed throughout the rest of the day. The L-arginine supplementation was maintained during the second infusion but was administered at half of its dose (0.833 g·m2 body surface area−1·dose−1) every 2 h. After an interval of at least 1 week, the subjects with MELAS syndrome were admitted for a second time, where a similar procedure was conducted, but rather than L-arginine, L-citrulline was given at the same dose and frequency. Control subjects were admitted once and had one stable isotope infusion after being on the controlled diet for 48 h. With each stable isotope infusion, blood samples were drawn before and during the isotope infusion at times 0, 1, 2, 3, 4, 5, 5.5, and 6 h.
2.3. Stable isotope infusion protocol
Solutions of 15N2-guanidino arginine (98%), 13C-,2H4-citrulline (99%), and 15N-citrulline (98%) (Cambridge Isotope Laboratories, Woburn, MA) were prepared in normal saline at the TCH investigational pharmacy. Two intravenous catheters were established in opposite arms, with one catheter used for isotope infusion and the other for blood sampling. The isotope infusion was initiated by the administration of a bolus containing 15N2-arginine (5 μmol·kg−1), 13C-,2H4-citrulline (1 μmol·kg−1), and 15N-citrulline (0.16 μmol·kg−1); followed by a continuous infusion of 15N2-arginine (5 μmol·kg−1·h−1) and 13C-2H4-citrulline (1 μmol·kg−1·h−1) for 6 h. Blood samples were drawn before and during the isotope infusion (Fig. 2).
2.4. Sample analyses
The blood samples were drawn into heparinized tubes and centrifuged immediately. The plasma was transferred and stored at −70 °C for later analyses. The plasma arginine and citrulline isotopic enrichments were measured by liquid chromatography–tandem mass spectroscopy (LC-MS/MS) after conversion to their DANS [5-(dimethylamino)-1-napthalene sulfonamide] derivatives as previously described [23]. The isotopic enrichment of plasma NO metabolites (nitrite and nitrate; NOx) was determined by negative chemical ionization gas chromatography–mass spectrometry (NCI GC-MS). Using cadmium and acetic acid, nitrate was reduced to nitrite which was subsequently converted to its pentafluorobenzyl derivative and extracted with toluene, as previously described [24,25].
Plasma amino acid concentrations were measured by standard high-pressure liquid chromatography (HPLC). Plasma NOx and ADMA concentrations were measured by in vitro isotope dilution. To measure the NOx concentration, plasma samples were spiked with a known quantity of Na15NO3, the nitrate was reduced to nitrite, and the isotopic enrichment of the nitrite was measured as previously described [24,25]. To measure the ADMA concentration, plasma samples were spiked with a known quantity of 2H7-ADMA, converted into its DANS derivative, and the isotopic enrichment of the ADMA measured as previously described [23].
2.5. Calculations
The isotopic enrichments of arginine and citrulline reached a plateau in plasma during the final 3 h of the infusion; therefore, the rate of appearance or flux (Q) of arginine and citrulline was calculated from the steady-state equation:
where IEinf is the isotopic enrichment of infused arginine or citrulline, IEplateau is the isotopic enrichment of arginine or citrulline in plasma at the steady state, and i is the infusion rate of 15N2-arginine or 13C-,2H4-citrulline.
Under steady-state conditions, the rate of appearance of arginine or citrulline equals the rate of disappearance. Therefore, arginine or citrulline clearance rates in mL·kg−1·min−1 were calculated by dividing the flux by the plasma concentration.
De novo arginine synthesis rate was calculated from the rate of conversion of citrulline to arginine as previously described [26]:
where IEarg is the plateau plasma enrichment of the M+5 isotopomer of arginine (13C-,2H4-arginine derived from 13C-,2H4-citrulline), IEcit is the plateau plasma enrichment of the M+5 isotopomer of citrulline (13C-,2H4-citrulline), and icit is the infusion rate of 13C-2H4-citrulline.
The NO synthesis rate was calculated from the rate of conversion of arginine to citrulline as previously described [26]:
where IEcit is the plateau plasma enrichment of the M+1 isotopomer of citrulline (15N-citrulline derived from 15N2-arginine), IEarg is the plateau plasma enrichment of the M+2 isotopomer of arginine (15N2-arginine) and iarg is the rate of infusion of 15N2-arginine.
NO synthesis rate was also calculated according to the precursor-product equation, as previously described [27]:
where [IE NOxt6−IE NOxt3] is the increase in isotopic enrichment of NOx over the period from 3 to 6 h (t6−t3) of the infusion and IEarg is the plateau isotopic enrichment of plasma arginine.
The absolute synthesis rate (ASR) of plasma NOx was calculated as the product of NOx concentration by FSR and expressed as μmol·L plasma−1·h−1.
2.6. Statistical analysis
Results were expressed as means±SEMs. The results of the two initial infusions administered during the first (before arginine) and second (before citrulline) admissions for subjects with MELAS syndrome were averaged and compared to the values of the control subjects using the unpaired Student’s t test. Differences in subjects with MELAS syndrome before and after arginine or citrulline supplementation were assessed by the paired Student’s t test. Tests were considered statistically significant if p<0.05. NO synthesis rate was considered the primary analysis of the study, whereas different plasma concentrations and flux measurements were considered secondary analyses.
3. Results
Subjects with MELAS syndrome had a lower mean body weight and height (p<0.05), but no significant differences in age, gender, and body mass index (Table 1). Subjects with MELAS syndrome were found to have lower plasma arginine (p<0.001) and citrulline (p<0.05) concentrations. They also had higher concentrations of plasma alanine (p<0.001) and branched-chain amino acids (BCAA) valine (p<0.005), leucine (p<0.05), and isoleucine (p<0.05). Other amino acids did not show significant differences between the two groups. Although NOx showed a trend to be lower in subjects with MELAS syndrome, the difference did not reach a statistically significant level. The plasma ADMA concentration was significantly higher in subjects with MELAS syndrome (p<0.005) (Table 2).
Table 1.
Characteristics of the research subjects, presented as mean±SD.
| Parameter | Control subjects (n=10) |
Subjects with MELAS (n=10) |
P |
|---|---|---|---|
| Age | 29.7±9.6 | 31.8±11.8 | NS |
| Gender (male/female) | 6/4 | 7/3 | NS |
| Weight (kg) | 68.0±11.9 | 56.3±10.1 | <0.05 |
| Height (cm) | 172.8±9.0 | 162.0±8.6 | <0.05 |
| BMI (kg·m2 body surface area−1) | 22.6±2.7 | 21.4±3.0 | NS |
(BMI: body mass index, NS: not statistically significant).
Table 2.
Plasma amino acids, NOx, and ADMA concentrations; arginine and citrulline kinetics; and NO synthesis rate in subjects with MELAS syndrome and control subjects. Values are expressed as means±SEMs.
| Control subjects (n=10) | Subjects with MELAS (n=10) | P | |
|---|---|---|---|
| Amino acids lower in subjects with MELAS | |||
| Arginine (μM) | 77.8±4.4 | 57.4±2.4 | <0.001 |
| Citrulline (μM) | 28.1±1.1 | 23.2±1.6 | <0.05 |
| Amino acids higher in subjects with MELAS | |||
| Alanine (μM) | 358.2±25.0 | 534.0±19.3 | <0.001 |
| Valine (μM) | 181.8±7.1 | 228.1±9.7 | <0.005 |
| Leucine (μM) | 114.3±8.3 | 143.4±8.2 | <0.05 |
| Isoleucine (μM) | 62.9±5.4 | 80.5±6.7 | <0.05 |
| NOx (μM) | 17.3±1.4 | 15.2±1.4 | NS |
| ADMA (μM) | 0.38±0.02 | 0.53±0.04 | <0.005 |
| Arginine and citrulline flux and clearance | |||
| Arginine flux (μmol·kg−1·h−1) | 56.4±3.0 | 49.8±2.9 | NS |
| Arginine clearance (mL·kg−1·min−1) | 13.4±0.5 | 16.2±1.0 | <0.05 |
| Citrulline flux (μmol·kg−1·h−1) | 8.2±0.6 | 5.7±0.4 | <0.005 |
| Citrulline clearance (mL·kg−1·min−1) | 5.6±0.4 | 5.0±0.4 | NS |
| De novo arginine | |||
| De novo arginine synthesis rate (μmol·kg−1·h−1) | 5.5±0.6 | 3.5±0.3 | <0.005 |
| Ratio of de novo arginine synthesis to arginine flux (%) | 9.9±1.0 | 7.1±0.6 | <0.05 |
| NO synthesis rates | |||
| Arginine-to-citrulline flux (μmol·kg−1·h−1) | 0.100±0.013 | 0.067±0.008 | <0.05 |
| ASR of NOx (μmol·L plasma−1·h−1) | 0.49±0.03 | 0.36±0.05 | <0.05 |
Arginine flux was not statistically different between subjects with MELAS syndrome and control subjects. However, arginine clearance was higher in subjects with MELAS syndrome (p<0.05). Citrulline clearance was not different between the two groups. Subjects with MELAS syndrome had lower citrulline flux (p<0.005), de novo arginine synthesis rate (p<0.005), percentage of arginine flux derived from de novo synthesis (p<0.05), and NO synthesis rate, measured by arginine-to-citrulline flux (p<0.05) and the ASR of NOx (p<0.05) (Table 2).
The plasma arginine concentration increased after oral supplementation with arginine (p<0.001) or citrulline (p<0.001). Similarly, plasma citrulline increased after supplementation with arginine (p<0.05) or citrulline (p<0.005). However, citrulline supplementation led to greater increases in both arginine and citrulline plasma concentrations. The BCAA were lower after both arginine and citrulline supplementation. The differences were significant for valine (p<0.001), leucine (p<0.05), and isoleucine (p<0.05) after arginine supplementation, and for valine (p<0.005) after citrulline supplementation. The alanine concentration showed a statistically significant decrease after arginine (p<0.05). However, a greater decrement in plasma alanine level was observed after citrulline supplementation (p<0.001). Plasma NOx concentration showed a trend to being higher after arginine and citrulline supplementation, but these differences did not reach statistical significance. ADMA levels did not change significantly with arginine or citrulline supplementation (Table 3).
Table 3.
The effect of arginine and citrulline supplementation on plasma amino acids, NOx, and ADMA concentrations in subjects with MELAS syndrome (n=10). Values are expressed in μM as means±SEMs.
| L-Arginine supplementation
|
L-Citrulline supplementation
|
|||||
|---|---|---|---|---|---|---|
| Before | After | P | Before | After | P | |
| Argininea | 57.1±3.2 | 143.8±9.9 | <0.001 | 57.6±2.1 | 182.0±14.4 | <0.001 |
| Citrullinea | 22.6±1.7 | 26.6±2.4 | <0.05 | 23.8±1.8 | 115.6±20.1 | <0.005 |
| Alaninea | 531.8±22.6 | 495.6±25.6 | <0.05 | 536.3±18.8 | 433.9±27.7 | <0.001 |
| Valine | 234.8±12.1 | 204.0±8.0 | <0.001 | 221.4±9.7 | 196.4±11.1 | <0.005 |
| Leucine | 148.5±11.2 | 125.9±7.1 | <0.05 | 138.3±6.6 | 124.9±10.1 | NS |
| Isoleucine | 83.0±7.2 | 70.4±4.5 | <0.05 | 78.0±7.2 | 70.3±9.1 | NS |
| NOx | 15.0±1.5 | 16.5±1.8 | NS | 15.6±1.7 | 16.9±2.1 | NS |
| ADMA | 0.54±0.04 | 0.54±0.03 | NS | 0.51±0.04 | 0.56±0.04 | NS |
The differences are statistically higher after citrulline supplementations <0.05).
Arginine flux increased after arginine (p<0.001) and citrulline (p<0.001) supplementation. Citrulline flux also increased after arginine (p<0.05) and citrulline (p<0.001) supplementation. The increases in both arginine flux and citrulline flux were more significant after citrulline supplementation. The clearance of arginine and citrulline did not show significant differences with arginine or citrulline supplementation. The de novo arginine synthesis rate increased after arginine supplementation (p<0.05), however, the increase was greater after citrulline supplementation (p<0.001). The contribution of de novo arginine synthesis to arginine flux decreased after arginine supplementation (p<0.001) and increased after citrulline supplementation (p<0.001). The NO synthesis rate, measured by the two methods, arginine-to-citrulline flux and ASR of NOx, increased after arginine (p<0.001 and p<0.05, respectively) and, to a greater extent, citrulline supplementation (p<0.05 and p<0.05, respectively) (Fig. 3).
Fig. 3.
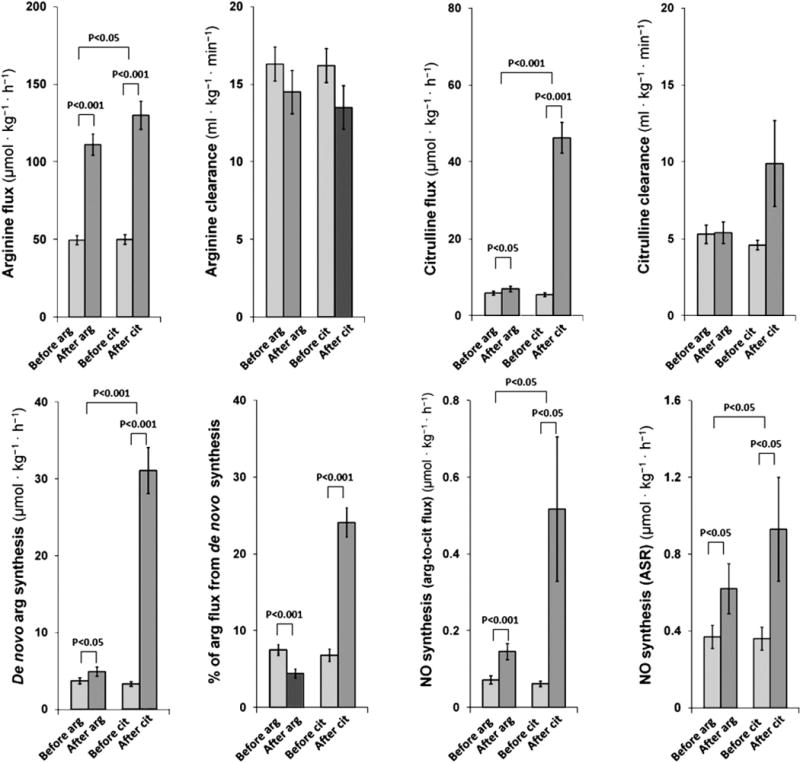
The effect of arginine and citrulline supplementation. Values are expressed as means±SEMs. Arginine flux increased after arginine (49.6±3.0→111.1± 6.9 μmol·kg−1·h−1, p<0.001) and citrulline (50.0±3.1→130.0±9.1 μmol·kg−1·h−1, p<0.001) supplementation. Arginine clearance did not show significant differences after arginine (16.3±1.1→14.5±1.4 mL·kg−1·min−1) and citrulline (16.2±1.1→13.5±1.5 ml·kg−1·min−1) supplementation. Citrulline flux increased after arginine (5.9±0.5→7.0±0.7 μmol·kg−1·h−1, p<0.05) and citrulline (5.5±0.5→46.3±4.0 μmol·kg−1·h−1, p<0.001) supplementation. Citrulline clearance did not show significant differences after arginine (5.3±0.6→5.4±0.7 mL·kg−1·min−1) and citrulline (4.6±0.3→9.9±2.8 ml·kg−1·min−1) supplementation. De novo arginine synthesis rate increased after arginine (3.7±0.4→4.9±0.6 μmol·kg−1·h−1, p<0.05) and citrulline (3.3±0.3→31.1±3.0 μmol·kg−1·h−1, p<0.001) supplementation. Percentage of arginine flux derived from de novo synthesis decreased after arginine supplementation (7.5±0.7→4.4±0.6%, p<0.001) and increased after citrulline supplementation (6.8±0.8→24.1±1.9%, p<0.001). The NO synthesis rate measured by arginine-to-citrulline flux increased after arginine (0.072±0.011→0.146±0.021 μmol·kg−1·h−1, p<0.001) and citrulline (0.062±0.007→0.517±0.188 μmol·kg−1·h−1, p<0.05) supplementation. The NO synthesis rate measured by ASR of NOx increased after arginine (0.37±0.06→0.62±0.13 μmol·L plasma−1·h−1, p<0.05) and citrulline (0.36±0.06→0.93±0.27 μmol·L plasma−1·h−1, p<0.05) supplementation.
4. Discussion
This study aimed to compare NO synthesis rates, amongst other related variables, between subjects with MELAS syndrome and control subjects, and to assess the effect of arginine and citrulline supplementation. The use of stable isotope infusion protocols in subjects with this syndrome provided a quantitative assessment of whole body arginine, citrulline, and NO metabolism.
Both impaired NO production due to endothelial dysfunction and post-production sequestration by increased COX have been suggested as possible mechanisms for decreased NO availability in MELAS syndrome [5,9]. A recently published study has showed increased protein nitration in mitochondrial diseases suggesting that oxidative stress results in decreased NO availability by inhibiting NOS and directing NO to reactive nitrogen species formation [28]. This study provides direct evidence for a defect in whole body NO production by showing that subjects with MELAS syndrome have lower NO synthesis rate when compared to control subjects. The results of this study also suggest that several factors play major roles in the impaired NO production observed in subjects with MELAS syndrome, including elevated ADMA levels, higher arginine clearance, and, most importantly, decreased de novo arginine synthesis secondary to decreased citrulline availability.
The results demonstrate that subjects with MELAS syndrome have lower plasma arginine and citrulline concentrations, consistent with previous reports [9,21]. In subjects with MELAS, the arginine flux was not significantly different from control subjects. However, the arginine clearance rate was higher, suggesting that increased clearance rather than decreased production may be responsible for the lower plasma arginine concentrations. It is not clear why subjects with MELAS syndrome may have an increased arginine clearance but one possible mechanism could be increased renal losses. Although the mechanism of hypocitrullinemia in MELAS is similarly unknown, it has been proposed that mitochondrial dysfunction results in impaired synthesis of citrulline, for which the main source is the de novo synthesis through a number of mitochondrial enzymes (Fig. 1) [9,29]. As citrulline is a non-protein amino acid with negligible content in the diet, its flux represents primarily the de novo synthesis rate [30]. Therefore, the lower citrulline flux observed in subjects with MELAS syndrome reflects a defect in citrulline synthesis. Most of the citrulline flux is directed towards arginine synthesis [26], therefore, the observed decreased de novo arginine synthesis rate in subjects with MELAS syndrome is likely secondary to lower citrulline availability. Based on these results, the lower NO production in subjects with MELAS syndrome may reflect decreased arginine availability due to two factors: i) increased arginine clearance resulting in a lower plasma arginine concentration, and ii) decreased citrulline synthesis, leading to decreased citrulline availability for the de novo arginine synthesis and hence lower intracellular arginine.
The higher ADMA concentration in subjects with MELAS syndrome can impair NOS activity thereby contributing to the reduced NO production. It is unclear why individuals with MELAS syndrome have higher plasma ADMA concentration. The mitochondrial dysfunction observed in this syndrome may result in increased reactive oxygen species production and oxidative stress that may impair the activity of dimethylarginine dimethylaminohydrolase (DDAH), the enzyme metabolizing ADMA. Alternatively, oxidative stress can stimulate cell injury and increased proteolysis resulting in increased ADMA production [15].
As one would expect, plasma arginine increased after arginine supplementation, however, the increase in plasma arginine was greater after administrating citrulline at the same dose as arginine. One possibility is that intestinal absorption of citrulline may be greater than that of arginine due to the action of intestinal arginase II on the ingested arginine [22]. While the citrulline concentration showed a modest increase after arginine supplementation, not surprisingly citrulline supplementation resulted in about a 5-fold increase in plasma citrulline concentration.
The observed increase in NO production after arginine or citrulline supplementation was accompanied by increased de novo arginine synthesis and plasma arginine concentration, suggesting that the increase in NO production was driven by increased arginine availability. The increase in de novo arginine synthesis results in increased intracellular arginine in sub-cellular compartments that contain NOS, ASS and ASL, which have been shown to co-localize in the cell. In contrast, plasma arginine needs to be transported into the cell and escape arginase degradation to finally reach the sub-cellular compartment that contains NOS [22]. Therefore; we propose that de novo-synthesized arginine plays a more important role in driving NO synthesis than plasma arginine. This would explain why citrulline led to a higher NO production than that associated with arginine supplementation. Citrulline supplementation resulted in a higher plasma arginine than arginine supplementation (~315% vs ~250%). However, more robust effects of citrulline supplementation were observed with regard to citrulline concentration, citrulline flux, and the de novo arginine synthesis rate. The latter was increased tenfold after citrulline supplementation, accounting for 24% of total arginine production, whereas it was only increased by 1.3-fold after arginine supplementation, accounting for only 5% of total arginine production. These findings suggest that compared to arginine, citrulline supplementation results in a marked increase in citrulline availability, leading to a higher de novo arginine synthesis, which in turn results in higher intracellular arginine and hence a greater increase in NO synthesis.
The higher plasma alanine level found in subjects with MELAS syndrome is expected due to the lactic acidemia that is one of the cardinal manifestations of MELAS syndrome [3]. Lactic acidemia likely results from an inability of dysfunctional mitochondria to carry out oxidative metabolism, leading to shunting of pyruvate to lactate [5]. Endothelial dysfunction and NO deficiency may also result in decreased blood perfusion and therefore may aggravate lactic acidemia. Interestingly, arginine and, to a greater extent, citrulline led to a reduction in plasma alanine concentrations, suggesting that such supplementations may improve lactic acidemia in MELAS syndrome by increasing NO production and improving perfusion. However, measurement of lactic acid concentrations was not part of our study design.
The metabolism of BCAA takes place in mitochondria via the branched-chain ketoacid dehydrogenase (BCKD) complex [31]. Elevated BCAA in subjects with MELAS may reflect another aspect of mitochondrial dysfunction. Interestingly, the BCAA concentrations decreased after arginine or citrulline supplementation. It is unclear how the improved NO production may result in reduced BCAA concentrations, but improved mitochondrial metabolism modulating BCKD activity may be a possible mechanism.
This study provides evidence that subjects with MELAS syndrome have lower NO production that improves after arginine and, to a greater extent, after citrulline supplementation, supporting their clinical use and suggesting that citrulline may have a better therapeutic effect than arginine. Although arginine supplementation or restriction has been proposed and trialled in several disorders [32], citrulline has not yet received much attention. Increasing NO availability will potentially improve perfusion in all microvasculature compartments, including muscle tissue. Therefore, the effect of arginine and citrulline supplementation may not be limited to improving stroke-like episodes, but may lead to improvement in other clinical features associated with MELAS syndrome, including migraine headaches, muscle weakness, exercise intolerance, and diabetes. Additional assessments of the clinical effects of arginine or citrulline supplementation on different aspects of MELAS syndrome are needed. Although we studied subjects with MELAS syndrome, the same principles could potentially be applied to other human diseases where endothelial dysfunction and NO deficiency play a role in their pathogenesis.
Although previous studies have demonstrated that arginine supplementation improves clinical symptoms associated with stroke-like episodes in subjects with MELAS syndrome [6,21], no studies had been conducted to assess NO production in MELAS syndrome. This study provides an in vivo assessment of whole body arginine, citrulline, and NO metabolism in subjects with MELAS syndrome. The use of well-established stable isotope infusion protocols has enabled us to quantitatively demonstrate impaired NO production in subjects with MELAS syndrome and to assess the effect of arginine and citrulline supplementation in restoring NO production. Although perturbation of NO metabolism has been suggested to play a major role in mitochondrial disorders, to the best of our knowledge, this is the first study that provides an in vivo whole body assessment of NO metabolism in subjects with a mitochondrial disorder.
5. Conclusions
In conclusion, lower NO production in subjects with MELAS syndrome may result from several mechanisms, including higher plasma ADMA concentrations and lower arginine availability due to increased arginine clearance and decreased de novo arginine synthesis. Based on our finding that citrulline supplementation results in higher NO production than arginine supplementation, we propose that a major determinant of NO production is the de novo arginine synthesis, which is substantially increased with citrulline supplementation. The improvement in NO production with arginine or citrulline supplementation supports their use in MELAS and suggests that citrulline may have a better therapeutic effect than arginine.
Acknowledgments
This study was funded by the Society for Inherited Metabolic Disorders (SIMD) through the SIMD/Hyperion fellowship award that Ayman El-Hattab has received. We also acknowledge the patients and their families for their participation, the National Institutes of Health for funding the General Clinical Research Center (GCRC) (M01-RR0188), the GCRC staff at Texas Children’s Hospital, the United Mitochondrial Disease Foundation (UMDF), and the Pfeiffer Foundation.
Footnotes
Supplementary materials related to this article can be found online at doi:10.1016/j.ymgme.2012.01.016.
References
- 1.Chinnery PF, Turnbull DM. Epidemiology and treatment of mitochondrial disorders. Am J Med Genet. 2011;106:94–101. doi: 10.1002/ajmg.1426. [DOI] [PubMed] [Google Scholar]
- 2.Wallace DC. Diseases of the mitochondrial DNA. Annu Rev Biochem. 1992;61:1175–1212. doi: 10.1146/annurev.bi.61.070192.005523. [DOI] [PubMed] [Google Scholar]
- 3.Hirano M, Pavlakis SG. Mitochondrial myopathy, encephalopathy, lactic acidosis, and strokelike episodes (MELAS): current concepts. J Child Neurol. 1994;9:4–13. doi: 10.1177/088307389400900102. [DOI] [PubMed] [Google Scholar]
- 4.Scaglia F, Northrop JL. The mitochondrial myopathy encephalopathy, lactic acidosis with stroke-like episodes (MELAS) Syndrome. A review of treatment options. CNS Drugs. 2006;20:443–464. doi: 10.2165/00023210-200620060-00002. [DOI] [PubMed] [Google Scholar]
- 5.Sproule DM, Kaufmann P. Mitochondrial encephalopathy, lactic acidosis, and strokelike episodes: basic concepts, clinical phenotype, and therapeutic management of MELAS syndrome. Ann N Y Acad Sci. 2008;1142:133–158. doi: 10.1196/annals.1444.011. [DOI] [PubMed] [Google Scholar]
- 6.Koga Y, Akita Y, Nishioka J, et al. L-Arginine improves the symptoms of strokelike episodes in MELAS. Neurology. 2005;64:710–712. doi: 10.1212/01.WNL.0000151976.60624.01. [DOI] [PubMed] [Google Scholar]
- 7.Ohama E, Ohara S, Ikuta F, et al. Mitochondrial angiopathy in cerebral blood vessels of mitochondrial encephalomyopathy. Acta Neuropathol. 1987;74:226–233. doi: 10.1007/BF00688185. [DOI] [PubMed] [Google Scholar]
- 8.Goto Y, Horai S, Matsuoka T, et al. Mitochondrial myopathy, encephalopathy, lactic acidosis, and stroke-like episodes (MELAS): a correlative study of the clinical features and mitochondrial DNA mutation. Neurology. 1992;42:545–550. doi: 10.1212/wnl.42.3.545. [DOI] [PubMed] [Google Scholar]
- 9.Naini A, Kaufmann P, Shanske S, et al. Hypocitrullinemia in patients with MELAS: an insight into the “MELAS paradox”. J Neurol Sci. 2005;229:87–193. doi: 10.1016/j.jns.2004.11.026. [DOI] [PubMed] [Google Scholar]
- 10.Green DJ, Maiorana A, O’Driscoll G, Taylor R. Effect of exercise training on endothelium-derived nitric oxide function in humans. J Physiol. 2004;561:1–25. doi: 10.1113/jphysiol.2004.068197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Toda N, Okamura T. The pharmacology of nitric oxide in the peripheral nervous system of blood vessels. Pharmacol Rev. 2003;55:271–324. doi: 10.1124/pr.55.2.3. [DOI] [PubMed] [Google Scholar]
- 12.Palmer RM, Rees DD, Ashton DS, Moncada S. L-arginine is the physiological precursor for the formation of nitric oxide in endothelium-dependent relaxation. Biochem Biophys Res Commun. 1988;153:1251–1256. doi: 10.1016/s0006-291x(88)81362-7. [DOI] [PubMed] [Google Scholar]
- 13.Moncada S, Higgs A. The L-arginine-nitric oxide pathway. N Engl J Med. 1993;329:2002–2012. doi: 10.1056/NEJM199312303292706. [DOI] [PubMed] [Google Scholar]
- 14.Wu G, Morris SM., Jr Arginine metabolism: nitric oxide and beyond. Biochemical. 1998;336:1–17. doi: 10.1042/bj3360001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Teerlink T, Luo Z, Palm F, Wilcox CS. Cellular ADMA: regulation and action. Pharmacol Res. 2009;60:448–460. doi: 10.1016/j.phrs.2009.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Vallance P, Leone A, Calver A, et al. Accumulation of an endogenous inhibitor of nitric oxide synthesis in chronic renal failure. Lancet. 1992;339:572–575. doi: 10.1016/0140-6736(92)90865-z. [DOI] [PubMed] [Google Scholar]
- 17.Surdacki A, Nowicki M, Sandmann J, et al. Reduced urinary excretion of nitric oxide metabolites and increased plasma levels of asymmetric dimethylarginine in men with essential hypertension. J Cardiovasc Pharmacol. 1999;33:652–658. doi: 10.1097/00005344-199904000-00020. [DOI] [PubMed] [Google Scholar]
- 18.Wang D, Strandgaard S, Iversen J, Wilcox CS. Asymmetric dimethylarginine, oxidative stress, and vascular nitric oxide synthase in essential hypertension. Am J Physiol Regul Integr Comp Physiol. 2009;296:195–200. doi: 10.1152/ajpregu.90506.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Asagami T, Abbasi F, Stuelinger M, et al. Metformin treatment lowers asymmeteric dimethylarginine concentrations in patients with type 2 diabetes. Metabolism. 2003;51:843–846. doi: 10.1053/meta.2002.33349. [DOI] [PubMed] [Google Scholar]
- 20.Tarnow L, Hovind P, Teerlink T, Stehouwer CD, Parving HH. Elevated plasma asymmetric dimethylarginine as a marker of cardiovascular morbidity in early diabetic nephropathy in type 1 diabetes. Diabetes Care. 2004;27:765–769. doi: 10.2337/diacare.27.3.765. [DOI] [PubMed] [Google Scholar]
- 21.Koga Y, Akita Y, Nishioka J, et al. MELAS and L-arginine therapy. Mitochondrion. 2007;7:133–139. doi: 10.1016/j.mito.2006.11.006. [DOI] [PubMed] [Google Scholar]
- 22.Mori M, Gotoh T. Regulation of nitric oxide production by arginine metabolic enzymes. Biochem Biophys Res Commun. 2000;3:715–719. doi: 10.1006/bbrc.2000.3169. [DOI] [PubMed] [Google Scholar]
- 23.Kao CC, Bandi V, Guntupalli KK, et al. Arginine, citrulline and nitric oxide metabolism in sepsis. Clin Sci (Lond) 2009;117:23–30. doi: 10.1042/CS20080444. [DOI] [PubMed] [Google Scholar]
- 24.Tsikas D, Boger RH, Bode-Boger SM, et al. Quantification of nitrite and nitrate in human urine and plasma as pentafluorobenzyl derivatives by gas chromatography–mass spectrometry using their 15N-labelled analogs. J Chromatogr B Biomed Appl. 1994;661:185–191. doi: 10.1016/0378-4347(94)00374-2. [DOI] [PubMed] [Google Scholar]
- 25.Jahoor F, Badaloo A, Villalpando S, et al. Arginine flux and intravascular nitric oxide synthesis in severe childhood undernutrition. Am J Clin Nutr. 2007;86:1024–1031. doi: 10.1093/ajcn/86.4.1024. [DOI] [PubMed] [Google Scholar]
- 26.Castillo L, Chapman TE, Sanchez M, et al. Plasma arginine and citrulline kinetics in adults given adequate and arginine-free diets. Proc Natl Acad Sci U S A. 1993;90:7749–7753. doi: 10.1073/pnas.90.16.7749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Villalpando S, Gopal J, Balasubramanyam A, et al. In vivo arginine production and intravascular nitric oxide synthesis in hypotensive sepsis. Am J Clin Nutr. 2006;84:197–203. doi: 10.1093/ajcn/84.1.197. [DOI] [PubMed] [Google Scholar]
- 28.Vattemi G, Mechref Y, Marini M, et al. Increased protein nitration in mitochondrial diseases: evidence for vessel wall involvement. Mol Cell Proteomics. 2011;10 doi: 10.1074/mcp.M110.002964. M110.002964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Parfait B, de Lonlay P, von Kleist-Retzow JC, et al. The neurogenic weakness, ataxia and retinitis pigmentosa (NARP) syndrome mtDNA mutation (T8993G) triggers muscle ATPase deficiency and hypocitrullinaemia. Eur J Pediatr. 1999;55:55–58. doi: 10.1007/s004310051009. [DOI] [PubMed] [Google Scholar]
- 30.Moinard C, Cynober L. Citrulline: a new player in the control of nitrogen homeostasis. J Nutr. 2007;137:1621S–1625S. doi: 10.1093/jn/137.6.1621S. [DOI] [PubMed] [Google Scholar]
- 31.Harris RA, Joshi M, Jeoung NH, Obayashi M. Overview of the molecular and biochemical basis of branched-chain amino acid catabolism. J Nutr. 2005;135:1527S–1530S. doi: 10.1093/jn/135.6.1527S. [DOI] [PubMed] [Google Scholar]
- 32.Coman D, Yaplito-Lee J, Boneh A. New indications and controversies in arginine therapy. Clin Nutr. 2008;27:489–496. doi: 10.1016/j.clnu.2008.05.007. [DOI] [PubMed] [Google Scholar]