

Abstract
We investigated the effect of the amount of body condition loss in the dry period and early lactation in 42 high-yielding Holstein-Friesian cows on milk yield and the share of fatty acids in milk fat. Energy reserves were estimated based on the body condition scoring (BCS) and backfat thickness (BFT). Milk yield and milk composition were determined over 305-d lactation. From d 6 to 60 of lactation, the concentrations of 43 fatty acids in milk fat were determined by gas chromatography. Cows were categorized based on body condition loss from the beginning of the dry period to the lowest point of the BCS curve in early lactation into three groups: low condition loss group (L) ≤0.5 points (n = 14); moderate condition loss group (M) 0.75 to 1.0 points (n = 16) and high condition loss group (H) >1.0 points (n = 12). Cows whose body energy reserves were mobilized at 0.8 BCS and 11 mm BFT, produced 12,987 kg ECM over 305-d lactation, i.e. 1,429 kg ECM more than cows whose BCS and BFT decreased by 0.3 and 5 mm, respectively. In group H, milk yield reached 12,818 kg ECM at body fat reserve mobilization of 1.3 BCS and 17 mm BFT. High mobilization of body fat reserves led to a significant (approx. 5%) increase in the concentrations of monounsaturated fatty acids-MUFA (mostly C18:1 cis-9, followed by C18:1 trans-11), a significant decrease in the levels of fatty acids adversely affecting human health, and a drop in the content of linoleic acid, arachidonic acid and docosahexaenoic acid in milk fat. In successive weeks of lactation, an improved energy balance contributed to a decrease in the concentrations of unsaturated fatty acids (UFA) and an increase in the conjugated linoleic acid (CLA) content of milk fat.
Keywords: Dairy Cows, Body Condition Loss, Backfat Thickness, Oleic Acid, CLA
INTRODUCTION
Intensive genetic selection has resulted in dairy cows that yield large amounts of milk in early lactation while mobilizing body reserves extensively. The cow’s milk output increases faster than its feed intake capacity, reaching their maxima 4 to 7 wks and 8 to 22 wks after calving, respectively (Ingvartsen, 2006). Negative energy balance (NEB) is partially caused by reduced dry matter intake around the time of calving. The rate and amount of changes in body condition score (BCS) and backfat thickness are good indicators of body energy reserve mobilization in early lactation (Grummer et al., 2004; Schroeder and Staufenbiel, 2006; Berry et al., 2007; Nogalski and Górak, 2008; Ayres et al., 2009). Energy stores are, therefore, a key component of milk production. Research in this area has implicated increased milk production with increased activity of lipolytic enzymes in adipose tissue, and more recently with greater expression of genes involved in body fat mobilization (Sumner and McNamara, 2007).
Milk fat is the main energy component of milk and the most easily digested animal fat in the human diet. Milk fat contains 400 to around 500 fatty acids (Reklewska et al., 2002). Due to its high content of saturated fatty acids and the presence of cholesterol, milk fat is considered a risk factor for atherosclerosis by some doctors and dieticians. However, it remains a rich source of many fatty acids (FA) known for their health-promoting properties (Reklewska et al., 2002; Khanal and Dhiman, 2007). Milk FA originate from four major pathways: directly from the diet, de novo synthesis in the mammary gland, formation in the rumen by biohydrogenation or bacterial degradation, and release from body fat stores (Stoop et al., 2009). Changes in milk fat composition over lactation imply shifts in the activity of these pathways and are related to changes in the energy status of the cow (Gross et al., 2011). Van Knegsel et al. (2005) suggested that during NEB, de novo synthesis of FA (C6:0 to C14:0) was reduced and body fat reserves used. Lactation stage and energy balance significantly contribute to variation in milk fat composition and alter the activity of different fatty acid pathways. The majority of studies conducted to date have focused on cattle herds with an average milk yield of up to 10,000 kg. Therefore, it seems interesting to determine whether milk production and the fatty acid profile of milk from high-yielding cows are affected by NEB.
The objective of this study was to investigate the effect of the amount of body reserve loss in the dry period and early lactation in high-yielding cows on milk yield and the fatty acid profile of milk fat.
MATERIALS AND METHODS
Cows
A total of 42 animals from a herd of 330 Holstein-Friesian cows were selected for the experiment based on the following criteria: a similar calving date to eliminate the effect of calving season (the selected cows calved between 16 January 2008 and 14 March 2008), yield for the previous lactation (not lower than 10,000 kg of milk during 305-d lactation), for heifers-yield estimated based on pedigree information, lactation number-proportionally for the entire herd (first lactation 15 cows, second lactation 15 cows and third lactation 12 cows). A herd of cows kept in a free-stall system was investigated in this experiment. The cows were milked in a parallel parlor three times a day. The animals were fed a total mixed ration (TMR) based on maize silage, grass and alfalfa haylage, and brewer’s spent grain. In the complete diet, the concentrate was supplemented with protein, minerals, vitamins and milk production enhancers (protein and rumen-protected fat, active yeast cultures and other energy supplements). Cows were fed a TMR formulated to produce 45 to 50 L of milk (7.15 MJ/kg net energy (lactation) and 18% crude protein). Dry cows and heifers on pre-calving days 60 to 45 were kept in deep litter free-stall facilities, and they were divided into two groups: i) dry cows: 6 to 8 wks ap (ante partum) to 3 wks ap; ii) transition cows: 3 wks ap to 1 wk pp (post partum). Dry cows were fed a TMR of grass and straw haylage supplemented with minerals in the dry period. Transition cows were administered lactation rations with an increasing share of concentrate. Feed was offered twice daily, using mixer wagons, and it was gathered up several times per day to ensure ad libitum access.
Study design and measurements
Energy reserves were estimated based on the body condition scoring (BCS) scale of 1 (thin) to 5 (obese) points with 0.25 intervals (Wildman et al., 1982), and backfat thickness (BFT). Ultrasound measurements of BFT were performed with the use of the Mysono 201 device (Medison Co.) equipped with a convex probe operating in the 2 to 5 MHz frequency range, in the area between the ischial tuberosity, the coxal tuber and the sacrum. Prior to the measurements, the skin was shaved and covered with gel. Evaluations were carried out at two-week intervals, from 5 wks ap to 20 wks pp. Milk yield in kg ECM and the average content of fat and protein over 305-d lactation were calculated for each cow, as follows.
Energy corrected milk: milk with a standardized energy content (Sjaunja et al., 1990),
Between lactation days 6 and 60, at five-day intervals, milk samples were collected from clinically healthy cows during morning milking. The proximate chemical composition of milk was determined by infrared spectrophotometry, using the Milco-scan 133 B analyzer (Foss Electric). Milk fat was extracted by the method proposed by Röse Gottlieb (AOEC, 1990), and the concentrations of 43 fatty acids were determined by gas chromatography, in the Varian CP 3800 chromatograph equipped with a split/splitless injector and a flameionization detector (FID). One μl ester samples were placed on a capillary column with a length of 100 m and an internal diameter of 0.25 mm, with the CP-sil88 stationary phase. Helium was used as the carrier gas, injector temperature was 260°C, and the total time of a single analysis was 68 min. The fatty acids and fatty acid groups analyzed in the study are presented in Table 1.
Table 1.
The fatty acids and fatty acid groups analyzed in the study
| Groups of fatty acids | Fatty acids included |
|---|---|
| SCFA | C 4 to C 10 |
| MCFA | C 11 to C 17 |
| LCFA | C 18 to C 22 |
| SFA | C 4:0, C 6:0, C 7:0, C 8:0, C 10:0, C 11:0, C 12:0, C 13:0, C 14:0, C 15:0, C 16:0, C 17:0 |
| UFA | C 10:1, C 12:1, C 14:1, C 16:1, C 17:1, C18:1 trans 6+9, C18:1 trans 10+11, C18:1 cis 9, C18:1 cis 11, C18:1 cis 12, C18:1 cis 13, C18:1 trans 16, C18:2 cis 9 trans 13, C 18:2, C18:3, C 18:2 cis 9 trans 11, C 20:1, C 20:2, C 20:4, C 20:5, C 22:5 |
| MUFA | C 10:1, C 12:1, C 14:1, C 16:1, C 17:1, C18:1 trans 6+9, C18:1 trans 10+11, C18:1 cis 9, C18:1 cis 11, C18:1 cis 12, C18:1 cis 13, C18:1 trans 16, C 20:1 |
| PUFA | C18:2 cis 9 trans 13, C 18:2, C18:3, C 18:2 cis 9 trans 11, C 20:2, C 20:4, C 20:5, C 22:5 |
| CLA | C 18:2 cis 9 trans 11 |
| n-3 | C 18:3, C 20:5 |
| n-6 | C 18:2, C 20:4 |
| NE | C 12:0, C 14:0, C 16:0, |
SCFA = Short-chain fatty acids; MCFA = Medium-chain fatty acids; LCFA = Long-chain fatty acids; SFA = Saturated fatty acids.
UFA = Unsaturated fatty acids; MUFA = Monounsaturated fatty acids; PUFA = Polyunsaturated fatty acids.
CLA = Conjugated linoleic acid; NE = Acids with a negative effect on human health.
In the herd, regular health examinations were performed by the same investigator once a week. Animal handling and sampling procedures performed for the needs of this study have been fully approved by the local ethics committee in Olsztyn, Poland.
Statistical analysis
The results were processed using Statistica 9.0 software (Statsoft 2010). The experimental factor was the amount of body condition loss from the beginning of the dry period to the lowest point of the BCS curve in early lactation. Cows were categorized based on body condition loss into three groups: low condition loss group (L) ≤0.5 points (n = 14); moderate condition loss group (M) 0.75 to 1.0 points (n = 16) and high condition loss group (H) >1.0 points (n = 12). The effect of body condition loss on milk yield and the share of fatty acids in milk fat was evaluated by the least squares analysis of variance, according to the below model:
where:
Yij = values of the analyzed parameters
μ = population mean
Ai = effect of the ith body condition loss
eij = random error.
Differences between means were estimated using Tukey’s test.
RESULTS
The depth and length of the NEB phase are illustrated by changes in the BCS profile and BFT (Figure 1). Cows with a higher BCS in the dry period were characterized by more intense fat reserve mobilization. Between 5 wks ap to the point of the lowest BCS, i.e. until wk 9 pp, the body condition of group H cows deteriorated by 1.3 points on average, which corresponded to an average change of 17 mm in BFT. In group H cows, intense mobilization of fat reserves began already at the end of the dry period. During the dry period and in the first three weeks of lactation, changes in their BCS exceeded 1 point. In cows whose BCS decreased to a lower degree, the energy demand and dietary energy intake were already balanced in wk 5 pp (group L).
Figure 1.
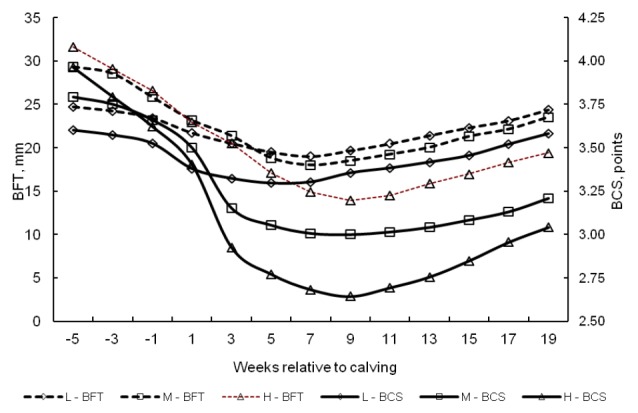
Changes in the body condition score (BCS) and backfat thickness (BFT) of cows in the dry period and early lactation. L = Low condition loss, M = Moderate condition loss, H = High condition loss.
In group M, the average total output over 305-d lactation was 12,987 kg ECM (Table 2). The difference between groups Ma and L was significant, at 1,429 kg (p<0.05). At peak lactation, the daily milk yield was 50 to 55 kg (Figure 2). The fat content of milk was very high in the first weeks of lactation, and it continued to decrease from wk 4. Milk fat concentrations until d 60 of lactation and the average fat content of milk during 305-d lactation were higher in cows intensively mobilizing their body fat reserves. The protein content of milk was highest in group L cows, but the noted differences were statistically non-significant (Table 2).
Table 2.
Characteristics of 305-d lactation (Means±SE)
| Parameter | Low condition loss group (n = 14) |
Moderate condition loss group (n = 16) |
High condition loss group (n = 12) |
|---|---|---|---|
| ECM (kg) | 11,558±330.3a | 12,987±403.4b | 12,818±304.2b |
| Fat (%) | 4.13±0.07 | 4.16±0.08 | 4.21±0.07 |
| Protein (%) | 3.27±0.03 | 3.23±0.04 | 3.22±0.03 |
p≤0.05.
Figure 2.
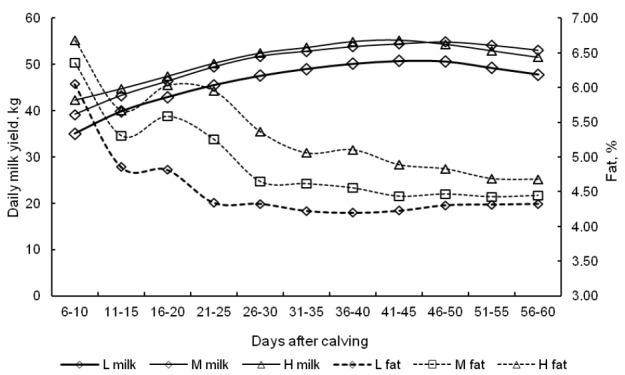
Changes in the daily yield and the fat content of milk in early lactation. L = Low condition loss, M = Moderate condition loss, H = High condition loss.
The rate of body energy reserve mobilization had a significant effect of the fatty acid profile of milk fat during the first two months of lactation (Table 3). Milk fat in group L cows had a significantly higher proportion of short-chain fatty acids and medium-chain fatty acids, and a lower proportion of long-chain fatty acids, compared with milk fat in the other groups. Most UFA are LCFA, which is why UFA concentrations were significantly (p<0.01) lower in group L. The difference in the levels of MUFA, in particular oleic acid, exceed 5 g/100 g total fat, in comparison with groups M and H (Figure 3). UFA had the highest share of the total fatty acid pool in the second week of lactation (Figure 4), and their proportion decreased on successive days. The n-3 fatty acid content of milk fat was significantly higher in group L cows, compared with the remaining groups, and it increased slightly with the progress of lactation (Figure 5).
Table 3.
The effect of body condition loss from d 6 to 60 of lactation on the concentrations of selected fatty acid groups in milk fat
| Fatty acid g/100 g milk fat | Low condition loss (L)
|
Moderate condition loss (M)
|
High condition loss (H)
|
|||
|---|---|---|---|---|---|---|
| LSM | SE | LSM | SE | LSM | SE | |
| SFCA | 7.97A | 0.183 | 7.07B | 0.162 | 7.09B | 0.205 |
| MCFA | 45.53a | 0.645 | 43.18b | 0.560 | 43.12b | 0.614 |
| LCFA | 46.50a | 0.851 | 49.75b | 0.705 | 49.73b | 0.772 |
| SFA | 63.20A | 0.721 | 58.27B | 0.647 | 58.18B | 0.758 |
| UFA | 36.79A | 0.772 | 41.73B | 0.654 | 41.76B | 0.753 |
| MUFA | 32.78A | 0.725 | 37.56B | 0.638 | 37.53B | 0.705 |
| PUFA | 4.14 | 0.122 | 4.17 | 0.113 | 4.23 | 0.158 |
| n-3 | 0.40a | 0.009 | 0.37b | 0.006 | 0.36b | 0.009 |
| n-6 | 2.57 | 0.047 | 2.50 | 0.028 | 2.47 | 0.048 |
| n-6/n-3 | 6.46 | 0.092 | 6.75 | 0.142 | 6.85 | 0.101 |
Means within a row with different superscripts differ.
p≤0.01;
p≤0.05.
SCFA = Short-chain fatty acids; MCFA = Medium-chain fatty acids; LCFA = Long-chain fatty acids; SFA = Saturated fatty acids.
UFA = Unsaturated fatty acids; MUFA = Monounsaturated fatty acids; PUFA = Polyunsaturated fatty acids.
Figure 3.
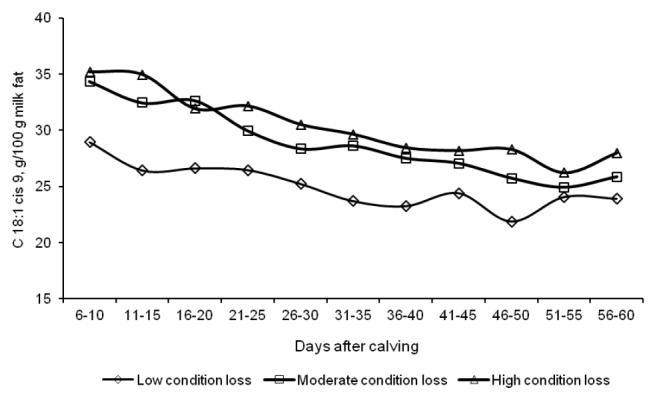
Changes in the oleic acid (C 18:1 cis 9) content of milk fat from d 6 to 60 of lactation.
Figure 4.
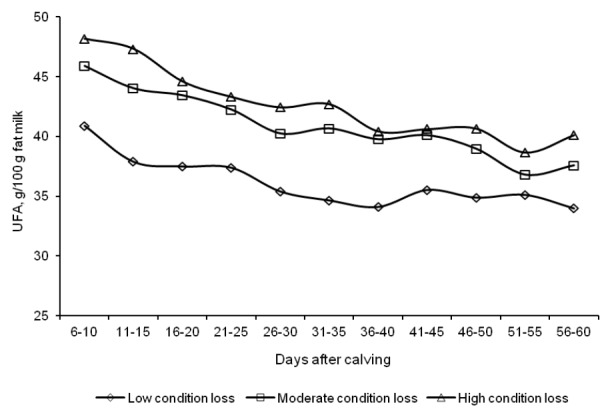
Changes in the unsaturated fatty acid (UFA) content of milk fat from d 6 to 60 of lactation.
Figure 5.

Changes in the n-3 fatty acid content of milk fat from d 6 to 60 of lactation.
The concentrations of functional fatty acids in milk were significantly influenced by the amount and rate of body energy reserve mobilization (Table 4). The content of short-chain butyric acid in milk was higher in group L than in the other groups, and it oscillated around 2.9 g/100 g total fat throughout the experiment (Figure 6). The rate of body fat reserve mobilization in early lactation had no significant effect on the CLA content of milk, which was found to increase in successive weeks along with an increase in daily milk yield (Figure 7). The concentrations of TVA and OA in milk from group L cows were lower, compared with the other groups (Table 4). A significant difference to the advantage of group L cows was noted with respect to the levels of LNA, AA and DHA.
Table 4.
The concentrations of functional fatty acids in milk fat from d 6 to 60 of lactation
| Fatty acid g/100 g milk fat | Low condition loss (L)
|
Moderate condition (M) loss
|
High condition loss (H)
|
|||
|---|---|---|---|---|---|---|
| LSM | SE | LSM | SE | LSM | SE | |
| C 4:0 (BA) | 2.931 | 0.053 | 2.799 | 0.043 | 2.839 | 0.066 |
| C 18:1 trans 11 (TVA) | 1.038Aa | 0.036 | 1.188A | 0.024 | 1.132b | 0.030 |
| C 18:1 cis (OA) | 25.53A | 0.687 | 29.43B | 0.653 | 29.68B | 0.677 |
| C 18:2 (LA) | 2.392 | 0.044 | 2.356 | 0.028 | 2.312 | 0.048 |
| C 18:2 cis 9 trans 11 (CLA) | 0.357 | 0.013 | 0.335 | 0.010 | 0.347 | 0.012 |
| C 18:3 (LNA) | 0.357Aa | 0.008 | 0.335b | 0.006 | 0.322B | 0.008 |
| C 20:4 (AA) | 0.176A | 0.007 | 0.145B | 0.004 | 0.160 | 0.008 |
| C 20:5 (EPA) | 0.043 | 0.003 | 0.036 | 0.002 | 0.041 | 0.003 |
| C 22:6 (DHA) | 0.076A | 0.003 | 0.059B | 0.002 | 0.065B | 0.003 |
Means within a row with different superscripts differ.
p≤0.01;
p≤0.05.
BA = Butyric acid; TVA = Trans-vaccenic acid; OA = Oleic acid; LA = Linoleic acid; CLA = Conjugated linoleic acid.
LNA = Linolenic acid; AA = Arachidonic acid; EPA = Eicosapentaenoic acid; DHA = Docosahexaenoic acid
Figure 6.
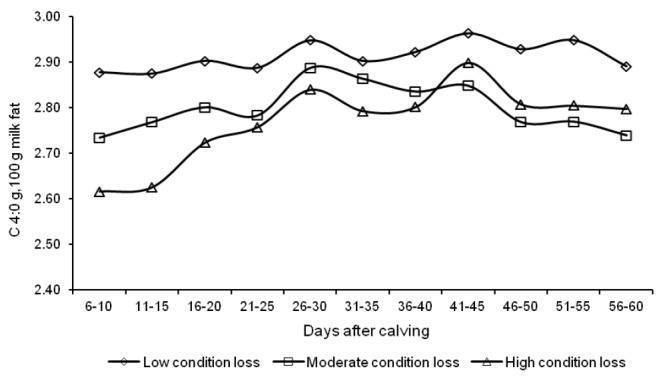
Changes in the butyric acid (C 4:0) content of milk fat from d 6 to 60 of lactation.
Figure 7.
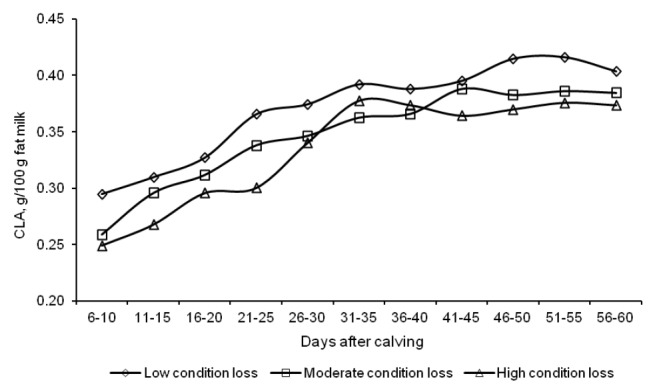
Changes in the conjugated linoleic acid (CLA) content of milk fat from d 6 to 60 of lactation.
Cows characterized by low body fat mobilization, in comparison with cows of the remaining groups, produced milk with a significantly higher content of fatty acids having an adverse effect on human health (Table 5). The differences in the levels of myristic acid between groups L and M, and L and H were significant (p>0.01), at 14.3% and 15.1%, respectively. The proportion of saturated fatty acids C 12:0, 14:0 and 16:0 increased with progressing lactation (Figure 8).
Table 5.
The concentrations of fatty acids adversely affecting human health in milk fat from d 6 to 60 of lactation
| Fatty acid g/100 g milk fat | Low condition loss (L)
|
Moderate condition (M) loss
|
High condition loss (H)
|
|||
|---|---|---|---|---|---|---|
| LSM | SE | LSM | SE | LSM | SE | |
| C 12:0 | 2.31a | 0.107 | 1.86b | 0.089 | 1.82b | 0.094 |
| C 14:0 | 8.05A | 0.258 | 6.90B | 0.224 | 6.83B | 0.248 |
| C 16:0 | 29.98a | 0.302 | 28.74b | 0.245 | 28.93b | 0.361 |
Means within a row with different superscripts differ.
p≤0.01;
p≤0.05.
Figure 8.

The concentrations of fatty acids adversely affecting human health in milk fat from d 6 to 60 of lactation.
DISCUSION
BCS at calving is positively correlated with a decrease in dry matter intake (Drackley 1999) and with BCS reduction after calving, and thus it is associated with the extent of NEB balance in early lactation (Berry et al., 2007). In our study, cows characterized by the highest drop in BCS (group H) already began to mobilize their fat reserves from 3 to 5 wks ap, while in the other cows the process started from 1 to 3 wks ap. The curves illustrating changes in BCS and BFT were similar, thus suggesting that a subjective assessment of body condition is a good indicator of the energy status of dairy cattle. Cows which mobilized approximately 11 mm subcutaneous fat (group M), and not 17 mm (group H), were characterized by the highest milk yield throughout lactation, which does not agree with the findings of Roche et al. (2009) who reported that body condition loss was positively correlated with milk production during lactation and peak milk yield. However, the cited authors analyzed cows producing 4,000 to 9,500 kg milk per lactation, kept in different systems and fed different diets. In our study, all cows were fed exclusively a TMR, and they produced nearly 13,000 milk per 305-d lactation on average. Under such conditions, the most intense mobilization of body energy reserves was not associated with the highest productivity, which could result from the fact that an excessive NEB makes the cow more susceptible to metabolic disorders (Santschi et al., 2011).
The fatty acid profile of milk is determined by shifts in the activity of fatty acid pathways (Stoop et al., 2009). Under severe NEB, the concentrations of short-chain and medium-chain fatty acids decrease, while the content of long-chain fatty acids increases. SFCA are an important source of energy for milk consumers as they are rapidly synthesized in the digestive tract and metabolized in the liver. Thus, they should have a high share of the total fatty acid pool. We found that high mobilization of body fat reserves adversely affected SFCA levels. The desirable, high UFA content of milk in groups M and H resulted from enhanced fat reserve mobilization and increased supply of non-esterified fatty acids (NEFA) to the udder. Gross et al. (2011) also observed a decrease in UFA concentrations in milk fat in successive weeks of lactation, as the energy balance improved. In our experiment, differences in MUFA content were mostly due to the elevated levels of oleic acid in the milk fat of cows intensively mobilizing their body energy reserves. Oleic acid (C 18:1 cis 9) is the predominant FA in adipocytes. It is released primarily through lipolysis during NEB (Rukkwamsuk et al., 2000). Plasma NEFA and triglycerides are utilized by the mammary gland for milk FA synthesis (Gross et al., 2011). In the present study, a high transfer rate of oleic acid from plasma into milk fat was confirmed by an elevated proportion of C 18:1 cis 9 in the milk fat of group M and H cows. Long-chain FA are derived from plasma and incorporated into milk fat, and they inhibit de-novo synthesis of short-chain FA by the mammary gland (Palmquist et al., 1993). The observed increase in butyric acid with progressing lactation in the present study is consistent with the decreasing adipose tissue mobilization. Van Haelst et al. (2008) suggested that an elevated proportion of oleic acid may be an interesting trait for the prediction of subclinical ketosis, particularly since this FA was elevated in milk fat before diagnosis of ketosis. The correlation between energy balance and the proportion of C 18:1 cis 9 was 0.77 (Gross et al., 2011).
The elevated levels of n-3 fatty acids in milk from cows characterized by low mobilization of body fat reserves could be due to increased supply of LNA to the rumen. LNA acts a substrate for both TVA, which is converted to CLA, and long-chain fatty acids formed by further desaturation and elongation. In our study, the overall n-6/n-3 fatty acid ratio was higher than recommended (Reklewska et al., 2002). The concentrations of n-6 fatty acids are higher in milk from cows fed a TMR, and so is the n-6/n-3 fatty acid ratio (Nałęcz-Tarwacka, 2006).
The CLA content of milk fat increased with progressing lactation, with an improved energy balance, which corroborates the findings of Gross et al. (2011) who noted an increase in CLA levels in successive weeks of lactation in both cows fed a restricted ration (reduced energy intake) and control group cows.
Higher concentrations of lauric, myristic and palmitic acids in milk from cows with lower mobilization of body fat reserves, observed in our study, resulted from differences in the rate of fatty acid synthesis in the mammary gland. As demonstrated by Schroeder et al. (2003), the concentrations of lauric acid and myristic acid increased in milk from cows whose BCS improved by 0.1 on average in early lactation. This indicates that body fat reserve mobilization is associated with an increase in the share of UFA in the total fatty acid pool in milk, mostly due to a high supply of NEFA accompanied by a lower SFA content. According to Nałęcz-Tarwacka (2006), the proportion of fatty acids which adversely affect human health in the total fatty acid pool may be reduced through pasture grazing and avoiding mono diets.
CONCLUSIONS
Under conditions of high milk production, the highest mobilization of body energy reserves by cows with the highest BCS in the dry period was not accompanied by the highest productivity, which could result from their greater susceptibility to subclinical disease. Our findings do not indicate a clear association between high milk production and intensive feeding (TMR) and deterioration of the functional quality of milk fat. High mobilization of body fat reserves was associated with elevated levels of MUFA (mostly oleic acid), reduced concentrations of fatty acids adversely affecting human health and a decreased content of some functional fatty acids in milk. With progressing lactation, an improved energy balance contributed to a decrease in UFA concentrations and an increase in the CLA content of milk.
REFERENCES
- Ayres H, Ferreira RM, de Souza Torres-Júnior JR, Demétrio CGB, de Lima CG, Baruselli PS. Validation of body condition score as a predictor of subcutaneous fat in Nelore (Bos indicus) cows. Livest Sci. 2009;123:175–179. [Google Scholar]
- AOAC. Official methods of analysis of the Associated Official Analytical Chemists. Washington DC, USA: 1990. Chepter 32. [Google Scholar]
- Berry DP, Buckley F, Diplom P. Body condition score and live weight effects on milk production in Irish Holstein-Friesian dairy cows. Animal. 2007;1:1351–1359. doi: 10.1017/S1751731107000419. [DOI] [PubMed] [Google Scholar]
- Drackley JK. Biology of dairy cows during the transition period: the final frontier? J Dairy Sci. 1999;82:2259–2273. doi: 10.3168/jds.s0022-0302(99)75474-3. [DOI] [PubMed] [Google Scholar]
- Gross J, van Dorland HA, Bruckmaier RM, Schwarz FJ. Milk fatty acid profile related to energy balance in dairy cows. J Dairy Res. 2011;78:479–488. doi: 10.1017/S0022029911000550. [DOI] [PubMed] [Google Scholar]
- Grummer RR, Mashek DG, Hayirli A. Dry matter intake and energy balance in the transition period. Veterinary Clinics of North America-Food Animal Practice. 2004;20:447–470. doi: 10.1016/j.cvfa.2004.06.013. [DOI] [PubMed] [Google Scholar]
- Ingvartsen KL. Feeding- and management-related disease in the transition cow: Physiological adaptation around calving and strategies to reduce feeding-related diseases. Anim Feed Sci Technol. 2006;126:175–213. [Google Scholar]
- Khanal RC, Dhiman TR. Status of milk fat conjugated linoleic acid (CLA) in selected commercial dairies. Asian-Aust J Anim Sci. 2007;20:1525–1538. [Google Scholar]
- Nałęcz-Tarwacka T. Treatises and Monographs. Publications of Warsaw Agricultural University; 2006. Effect of selected factors on the functional component content of milk fat in dairy cows; p. 108. [Google Scholar]
- Nogalski Z, Górak E. Effects of the body condition of heifers at calving and at the first stage of lactation on milk performance. Med Weter. 2008;64:322–326. [Google Scholar]
- Palmquist DL, Beaulieu AD, Barbano AD. Feed and animal factors influencing milk fat composition. J Dairy Sci. 1993;76:1753–1771. doi: 10.3168/jds.s0022-0302(93)77508-6. [DOI] [PubMed] [Google Scholar]
- Reklewska B, Oprządek A, Reklewski Z, Panicke L, Kuczyńska B, Oprządek J. Alternative for modifying the fatty acid composition and decreasing the cholesterol level in the milk of cows. Livest Prod Sci. 2002;76:135–243. [Google Scholar]
- Roche JR, Friggens NC, Kay JK, Fisher MW, Stafford KJ, Berry DP. Invited review: Body condition score and its association with dairy cow productivity, health, and welfare. J Dairy Sci. 2009;92:5769–5801. doi: 10.3168/jds.2009-2431. [DOI] [PubMed] [Google Scholar]
- Rukkwamsuk T, Geelen MJH, Kruip TAM, Wensing T. Interrelation of fatty acid composition in adipose tissue, serum, and liver of dairy cows during the development of fatty liver postpartum. J Dairy Sci. 2000;83:52–59. doi: 10.3168/jds.S0022-0302(00)74854-5. [DOI] [PubMed] [Google Scholar]
- Santschi DE, Lefebvre DM, Cue RI, Girard CL, Pellerin D. Incidence of metabolic disorders and reproductive performance following a short (35-d) or conventional (60-d) dry period management in commercial Holstein herds. J Dairy Sci. 2011;94:3322–3330. doi: 10.3168/jds.2010-3595. [DOI] [PubMed] [Google Scholar]
- Schroeder GF, Delahoy JE, Vidaurreta I, Bargo F, Gagliostro GA, Muller LD. Milk fatty acid composition of cows fed a total mixed ration or pasture plus concentrates replacing corn with fat. J Dairy Sci. 2003;86:3237–3248. doi: 10.3168/jds.S0022-0302(03)73927-7. [DOI] [PubMed] [Google Scholar]
- Schroeder UJ, Staufenbiel R. Invited review: Methods to determine body fat reserves in the dairy cow with special regard to ultrasonographic measurement of backfat thickness. J Dairy Sci. 2006;89:1–14. doi: 10.3168/jds.S0022-0302(06)72064-1. [DOI] [PubMed] [Google Scholar]
- Sjaunja LO, Baevre B, Junkkarinen L, Pedersen J, Setala J. Proc. Paris: 1990. A Nordic proposal for an energy corrected milk (ECM) formula; pp. 156–157. (27th Session of the ICRPMA). July 2–6. [Google Scholar]
- Stoop WM, Bovenhuis H, Heck JML. Effect of lactation stage and energy status on milk fat composition of Holstein-Friesian cows. J Dairy Sci. 2009;92:1469–1478. doi: 10.3168/jds.2008-1468. [DOI] [PubMed] [Google Scholar]
- Sumner JM, McNamara JP. Expression of lipolytic genes in the adipose tissue of pregnant and lactating Holstein dairy cattle. J Dairy Sci. 2007;90:5237–5246. doi: 10.3168/jds.2007-0307. [DOI] [PubMed] [Google Scholar]
- Van Haelst YNT, Beeckman A, van Knegsel ATM, Fievez V. Short communication: Elevated concentrations of oleic acid and long-chain fatty acids in milk fat of multiparous subclinical ketotic cows. J Dairy Sci. 2008;91:4683–4686. doi: 10.3168/jds.2008-1375. [DOI] [PubMed] [Google Scholar]
- Van Knegsel ATM, van den Brand H, Dijkstra HJ, Tamminga S, Kemp B. Effect of dietary energy source on energy balance, production, metabolic disorders and reproduction in lactating dairy cattle. Reprod Nutr Dev. 2005;45:665–688. doi: 10.1051/rnd:2005059. [DOI] [PubMed] [Google Scholar]
- Wildman EE, Jones GM, Wagner PE, Boman RL, Troutt HF, Lesch TN. A dairy cow body condition scoring system and its relationship to selected production characteristics. J Dairy Sci. 1982;65:495–501. [Google Scholar]