

Abstract
A polysaccharide named GSP-2 with a molecular size of 32 kDa was isolated from the fruiting bodies of Ganoderma sinense. Its structure was well elucidated, by a combined utilization of chemical and spectroscopic techniques, to be a β-glucan with a backbone of (1→4)– and (1→6)–Glcp, bearing terminal- and (1→3)–Glcp side-chains at O-3 position of (1→6)–Glcp. Immunological assay exhibited that GSP-2 significantly induced the proliferation of BALB/c mice splenocytes with target on only B cells, and enhanced the production of several cytokines in human peripheral blood mononuclear cells and derived dendritic cells. Besides, the fluorescent labeled GSP-2 was phagocytosed by the RAW 264.7 cells and induced the nitric oxide secretion from the cells.
Introduction
Lingzhi is a well-known anticancer fungus. Although it is the general name of Ganoderma species, the name Lingzhi usually means a single species G. lucidum, which is one of the most studied mushrooms in the world [1]. Its anti-cancer effects are associated with not only the cytotoxic triterpenoids, but also the immunomodulating polysaccharides via the inhibition of DNA polymerase and post-translational modification of the Ras oncoprotein, or the stimulation of cytokine production [2]–[5].
While Ganoderma sinense is also called the same name “Lingzhi” like G. lucidum, as recorded in Chinese Pharmacopoeia 2010, it is only distributed in China and contains little ganoderic acid like triterpenoids [6]–[7]. Both official Lingzhi species are rich in polysaccharides and possess multiple biological activities [1], such as antimicrobials [6]–[7], immunomodulation [8], and antitumor effect [9]. Since G. sinense is traditionally used in the form of decoction, the water-soluble polysaccharides are also considered its major active ingredients, having immune-balancing [10], antioxidant [11]–[12] and antitumor activities [13]. It was reported that the protein-bound polysaccharide from the fruiting bodies of G. Sinense could significantly induce the proliferation of human peripheral blood mononuclear cells (PBMCs), in the meanwhile enhance the secretion of tumor necrosis factor-α, interleukin (IL)-10 and IL-12 and transforming growth factor-β from PBMCs and enhance the population of CD14+ cell [13]. Unfortunately, few polysaccharides have been purified from G. sinense and little is known about their chemistry and bioactivity [1].
In our previous study, two purified polysaccharides GSP-6B and GSP-4 were reported from the fruiting bodies of G. sinense [14]–[15]. Their chemical structures were partially elucidated to be a hyperbranched β-glucan and a heteropolysaccharide, respectively. Additionally, both polysaccharides exhibited immunomodulating effects in vitro without cytotoxicity. However, they might not fully explain the chemistry and immunomodulating effects of G. sinense because of their low yield. In this study, we report on the structure elucidation and immunomodulatory activities of another new polysaccharide (GSP-2) which is isolated as a major polysaccharide fraction from the fruiting bodies of G. sinense.
Experimental Section
Materials and Chemicals
Dried fruiting bodies of G. sinense were purchased from herbal store in Hong Kong and authenticated by Professor Zhu-Liang Yang at Kunming Institute of Botany, Chinese Academy of Sciences. The voucher specimen was deposited at the Institute of Chinese Medicine, the Chinese University of Hong Kong, with the voucher specimen number 2010-3271. DEAE Sepharose CL-6B was purchased from GE Healthcare (UK). Reference monosaccharides, T-series dextrans, trifluoroacetic acid (TFA), dimethyl sulfoxide (DMSO), Griess Reagent and lipopolysaccharide (LPS) were purchased from Sigma (St. Louis, MO, USA). RPMI 1640 medium, fetal bovine serum (FBS) and penicillin/streptomycin (100 U/ml) were obtained from Invitrogen (NY, USA). The Raw 264.7 cells were purchased from ATCC, the Global Bioresource Center (Manassas, VA, USA). The fresh human buffy coats were supplied by Hong Kong Red Cross Blood Transfusion Service. The use of human buffy coat for experiment was approved by The Joint Chinese University of Hong Kong-New Territories East Cluster Clinical Research Ethics Committee. The IL-1β, IL-2, IL-4, IL-10, IL-12, IFN-γ, TNF-α ELISA test kits were purchased from BD Pharmingen (CA, USA). GM-CSF was the product of Norvatis (Switzerland).
General methods
The UV-vis spectra were tested on a Beckman DU650 ultraviolet and visible spectrophotometer. Gas chromatography mass spectrometry (GC-MS) tests for sugar composition analysis were performed on a Shimadzu QP-2010 instrument equipped with a DB-5 column (30 m×0.25 mm×0.25 µm) and a quadrupole rods mass detector (225 °C), the column temperature was increased from 140 °C to 225 °C in a rate of 2 °C/min then hold on 5 min. Gas GC-MS tests for methylation analysis were measured with a DB-5 column (30 m×0.25 mm×0.25 µm), and at temperatures programmed from 170–225°C at 2°C/min and then hold on 10 min. The FT-IR spectra (KBr pellets) were recorded on SPECORD in a range of 400–4000 cm−1. The absolute configuration test of the sugar residues were conducted under the method of Gerwig, Kamerling and Vliegenthart [16]–[17].
Extraction and purification of the polysaccharide
The extraction and preparation of crude polysaccharide has been reported before [14]–[15], after that one in five of the crude polysaccharides (1.0 g) dissolved in 50 mL water was loaded on a DEAE Sepharose CL-6B column (5.0×70.0 cm), and eluted with a 6-step gradient of distilled water, and sodium hydroxide. Guided by the colorimetric total carbohydrate test using the phenol-sulfuric acid method, after all the crude polysaccharide was separated by the DEAE Sepharose CL-6B column, the 0.1 M sodium chloride eluting fractions were combined, dialyzed, lyophilized, and futher purified by a series of Sepharcyl S-300 and Sepharcyl S-400 gel-permeation chromatographic process (2.6 cm×60 cm), eluted with water to afford a purified polysaccharide (GSP-2, 656 mg).
Determination of homogeneity and molecular weight
The homogeneity and molecular weight of GSP-2 were determined on an Agilent 1100 system equipped with an ELSD detector. Samples (2 mg/mL, 10 µL) were applied to TSK GMPWXL gel filtration columns (7.8×300 mm×2) and eluted with 20 mM CH3COONH4 at 0.6 mL/min with column temperature maintained at 40°C. Commercially available T-series dextrans (MW 2000, 670, 410, 270, 150, 80, 50, 12, 5 and 1 kD) were used as standard molecular markers.
Monosaccharide composition analysis
The identification and quantification of the monosaccharides of GSP-2 was achieved by GC-MS analysis. GSP-2 (10 mg) was hydrolyzed with 2 M trifluoroacetic acid (TFA) at 100 °C for 3 h [18]. The monosaccharides were conventionally converted into their completely acetylated derivation and analyzed by GC-MS according to the method of Lawrence and Lyengar [19].
Methylation analysis
GSP-2 (10 mg) was methylated three times according to the method reported by Needs [20]. Complete methylation was confirmed by the disappearance of the OH band (3200–3700 cm-1) in the IR spectrum. The methylated products were hydrolyzed, reduced and acetylated [21]. The partially methylated alditol acetates were analyzed by GC-MS.
Partial acid hydrolysis
50 mg of GSP-2 was partially hydrolyzed with 0.1 M TFA at 60 °C for 10 h [18]. The resulting hydrolysate was then dialyzed against distilled water for 24 h by a dialysis bag with 7000 Da cut-off. The non-dialysate was collected and coded as GSP-2-P for sugar composition and methylation analysis under the same protocol of the native polysaccharide GSP-2.
NMR studies
GSP-2 (25 mg) was dried in vacuum under the presence of P2O5 for 72 h, and then exchanged with deuterium by lyophilizing with D2O for three times. The deuterium exchanged polysaccharide was put in a 5-mm NMR tube and dissolved in 1.0 mL 99.96% D2O. All 1D and 2D NMR spectrums were acquired at 298 K on a Bruker Avance 500 MHz NMR spectrometer. Tetramethylsilane (TMS) was used as external standard for the 13C NMR spectrum, and D2O was used as internal standard for 1H NMR spectrum.
Proliferation of mouse splenocytes, splenic T, B cells after GSP-2 treatment
Female BALB/c mice, 8–10 weeks old (18–22 g), were purchased from the Experimental Animal Division of Peking University Health Sciences Center, Beijing, China. All animals were maintained at the specific pathogen free (SPF) laboratory of Experimental Animal Division of Peking University Health Science Center,with stable temperature, food and water supplying. All animal experiments were carried out with the permission of Beijing Experimental Animal Management Authority, Beijing, China, at the animal facilities of this department. The mice were sacrificed by the method of cervical dislocation,all efforts were made to minimize suffering,after that the spleens were collected. Spleens were gently smashed by pressing with the flat surface of a syringe plunger against stainless steel sieve (200 mesh). Red blood cells were lysed by brief treatment with distilled water. The splenocytes were washed twice and then resuspended in complete RPMI 1640 medium supplemented with 10% (v/v) FBS, penicillin/streptomycin (100 U/mL), L-glutamine (2 mM).
Freshly prepared splenocytes resuspended in RPMI 1640 medium at 107 cells/mL were plated into 100 mm tissue culture dishes and incubated for 4 h at 37°C in a CO2 incubator. The non-adherent cells were collected, washed twice in PBS, and then incubated with MACS magnetic microbeads coated with rat anti-mouse CD19 mAb (Myltenyi Biotec, Italy) at a density of 10 µL antibody solution/107 splenocytes for 30 min at 4°C. The labeled cells were washed twice with PBS and then applied to a MACS separation column. The effluent was collected as non-B cells. After further washes, the column was removed from the magnet separator and the B cells were flushed out of the column using a plunger. To increase the purity of T cells in the non-B cells, the non-B cell fraction was reapplied to a separation column and the effluent was collected as T cells. The isolated B cells were reacted with FITC-anti mouse CD19 antibody and T cells were reacted with FITC-anti mouse CD3 antibody for 30 min at 4°C. The purities of B cells and T cells were greater than 95% as evaluated by flow cytometry analysis.
Freshly prepared mouse splenocytes (4×105 cells/well), T cells (2×105 cells/well), and B cells (2×105 cells/well), were cultured in flat-bottomed 96-well plates (Nunc, Denmark) in a volume of 200 µl/well. Different concentrations of GSP-2 (10, 30, 100 µg/mL) or Dextran (50 µg/mL) or LPS (2 µg/mL) or ConA (2 µg/mL) were added to the wells. The cells were incubated at 37°C and 5% CO2 for 48 h. In the last 8 h of incubation, [3H] thymidine ([3H]TdR, 0.2 µCi/well) was added into each well. The cells were then harvested, using a 96-well plate harvester (Tomtec, USA), onto fiberglass filters and radioactivity on the filtermatt was counted in a MicroBeta Trilux LSC counter (EG&G Wallac, USA). For inhibition experiments Polymixin B (PMB) (10 ug/ml) was added into the culture system.
Nitric oxide production and phagocytosis of GSP-2 on macrophages
Macrophages RAW264.7 (4×105 cell/well) were seeded in 24-well plate overnight. Various concentration of GSP-2 (0, 100, 200, 400, 800 µg/mL) or LPS (10 µg/mL) were added and incubated for 24 h. Culture supernatant was added to Griess Reagent in the ratio of 1∶1 in a 96-well plate and the plate was incubated in darkness for 10 min. The plates were then read at a wavelength of 540 nm spectrophotometrically. Nitrite standard curve was plotted with standard NaNO2 solution with Griess treatment to determine the concentration of nitrite in culture supernatant samples.
In order to detect the phagocytosis of polysaccharide in macrophages, the polysaccharide GSP-2 was labeled with Alexa Fluor 488 succinimidyl ester (Molecular Probes, Eugene, OR) according to the reported protocol [22]. Briefly, a diaminopropane (DAP) moiety was added to the reducing terminus of the polysaccharides chain of GSP-2 by sodium borohydride reduction to produce a DAP-GSP-2. Then the amine reactive AlexaFluor 488 succinimidyl ester was attached to the DAP moiety located at the reducing terminus of GSP-2 to produce the fluorescent labeled polysaccharide GSP-2. For experiments, the RAW 264.7 cells (5×104 cells/well) were seeded at 24-well culture plates and incubated overnight. The cells were treated with 10 µg (in 1 mL PBS) labeled GSP-2 for 24 h. After incubation, the cells were collected, washed with PBS twice and resuspended in PBS. The fluorescence of the samples was detected by flow cytometry (Becton Dickinson FACSCanto II). The cells were also visualized and imaged under inverted fluorescent microscope (Olympus IX-71).
Cytokine production of human peripheral blood mononucelar cells and dendritic cells after GSP-2 treatment
The cytokine productions of GSP-2 treated-human peripheral blood mononuclear cells (PBMCs) and monocyte-derived dendritic cells (moDCs) were assessed to evaluate the immunomodulatory activities of GSP-2. The PBMCs were isolated from fresh human buffy coat by density centrifugation using Ficoll-Paque™ Plus (GE Healthcare) as previously reported 7. For generation of moDC, monocytes (2×106 cells/mL) were first selected from PBMC by attachment on culture flask at 37°C, 5% CO2 for 45 min. The non-adherent cells were removed by washing 2 to 3 times with a gentle stream of medium. The adherent cells were cultured at 1×106 cells/mL in RPMI-1640 medium plus 10% (v/v) FBS supplemented with 50 ng/mL GM-CSF and 40 ng/mL IL-4. Immature dendritic cells (DC) were harvested after 6 days, and maturation was induced for 2 days by adding 10 ng/mL lipopolysaccharide (LPS) in culture medium. The cells were incubated at 37°C in a humidified atmosphere of 5% CO2.
The isolated human PBMCs and moDCs (2×106 cells/ml) were then seeded in 96-well flat bottom microplates and incubated with GSP-2 from 0.3–1000 ng/ml. Lipopolysaccharide (LPS) (8 ng/ml) and a beta-glucan from Euglena gracilis (Sigma) (0.3–1000 ng/ml) were used as positive controls. After incubations of 18 h (for PBMCs) or 48 h (for moDCs) with the polysaccharides, the cell free supernatants were collected for cytokine ELISA experiments. The cytokines concentrations of IL-10 and IL-12 (from both PBMCs and moDCs) and IL-1, TNF-α (from PBMCs) were determined by ELISA kits according to the manufacturer instruction with detection limits ranged from 3.1 to 7.8 pg/ml.
Statistical Analysis
All the experiments described here were repeated at least 2 times. Results are presented as mean ± standard error of the mean (SEM). Comparison of the data was performed using the one way analysis of variance (ANOVA) followed by Bonferroni post-test, as appropriate, were performed using GraphPad PRISM software version 5.0 (GraphPad Software, USA). Significance was defined as a * P value of <0.05, ** P<0.01, *** P<0.001.
Result and Discussion
The structure elucidation
GSP-2, with high homogeneity as indicated by the single and symmetrical peak in the HPLC-GPC-ELSD analysis shown in Fig. 1A, has an apparent molecular weight of 32 kDa as calculated from the calibrated standard curve. Sugar composition analysis indicated that GSP-2 was mainly composed of glucose as its sugar constituents, and absolute configuration study further revealed that all the glucose residues in GSP-2 are of D configuration. Besides, amino acid composition analysis revealed that 5.2% of this fraction is protein, which was composed of 16 kinds of amino acids as shown in the Table 1. Since the de-protein process has been repeated for ten times, it is supposed that these amino acids mainly remained on the sugar chain of GSP-2 by covalent bonds. The characterization will make this fraction very soft and flexible and facilitate to be recognized by the polysaccharide receptors on the surface of the immune cells, such as Toll-like receptor 4 on the surface of dentritic cells [23]–[24].
Figure 1. HPLC profile, methylation analysis result and 1D NMR spectrums of GSP-2.

A: HPLC profile of GSP-2. Samples (2 mg/mL, 10 µL) were analyzed on an Agilent 1100 system equipped with an ELSD detector and TSK GMPWXL gel filtration columns (7.8×300 mm×2), with 20 mM CH3COONH4 as mobile phase at 0.6 mL/min and column temperature at 40°C. Commercially available T-series dextrans (MW 2000, 670, 410, 270, 150, 80, 50, 12, 5 and 1 kD). B: Methylation analysis result of GSP-2. GC-MS tests for methylation analysis were measured with a DB-5 column (30 m×0.25 mm×0.25 µm), and at temperatures programmed from 170–225 °C at 2 °C/min and then hold on 10 min, in the figure, a: T-Glcp, b: 1,3-linked Glcp, c: 1,4-linked Glcp, d: 1,6-linked Glcp, e: 1,3,6-linked Glcp. C-E: 1D NMR (1H, 13C, DEPT) spectra of GSP-2 in D2O with TMS as external standard, obtained on a Bruker AM 500 spectrometer with a dual probe in the FT mode at room temperature.
Table 1. Amino acid composition of the polysaccharide (GSP-2) isolated from the fruiting bodies of Ganoderma sinense a .
| Amino acid | Contnet µg/mg |
| Aspartic acid | 2.73 |
| Threonine | 10.72 |
| Serine | 8.71 |
| Glutamic acid | 3.79 |
| Glycine | 3.00 |
| Alanine | 6.50 |
| Vakine | 3.38 |
| Methiomine | 0.56 |
| Isoleucine | 1.18 |
| Leucine | 2.55 |
| Phenylalanine | 3.09 |
| Lysine | 0.61 |
| Histidine | 0.71 |
| Arginine | 0.30 |
| Proline | 3.28 |
| Tyrosine | 0.55 |
All the result were tested by a HITACHI automatic amino acid analyzer.
Methylation analysis revealed the exsitence of five types of sugar residues in GSP-2, i.e., non-reducing terminal glucopyranosyl (Residue A, 27%), (1→3)–linked glucopyranosyl (Residue B, 16%), (1→4)–linked glucopyranosyl (Residue C, 15%), (1→6)–linked glucopyranosyl (Residue D, 22%), (1→3, 6)–linked glucopyranosyl (Residue E, 20%), (Fig. 1B, Table 2). In the FT-IR spectrum of GSP-2, the strong band at 3412.2 cm−1 was attributed to the hydroxyl stretching vibration of the polysaccharide, and that at 2917.27 cm−1 was due to the C–H stretching vibration absorption. Characteristically, the bands at 1000–1100 cm−1 suggested the presence of pyranose form of the glucosyl residue in the GSP-2 (Data not shown).
Table 2. GC–MS test result for the methylated sugar moieties of the polysaccharide GSP-2 a , b .
| Residue No. | Methylated sugars | Type of linkage | Molar ratio |
| A | 2,3,4,6-Me4-Glc | Terminal Glcp | 27% |
| B | 2,4,6-Me3- Glc | 1,3-Linked Glcp | 16% |
| C | 2,3,6-Me3-Glc | 1,4-Linked Glcp | 15% |
| D | 2,3,4Me3- Glc | 1,6-Linked Glcp | 22% |
| E | 2,4-Me2-Glc | 1,3,6-Linked Glcp | 20% |
The result were tested on DB-5 GC-MS column.
All the sugar residues were primarily identified by their MS spectrum and further confirmed by their relative retention time to 2,3,4,6-Me4-Glc.
As the first step to understand the sequence of the sugar residues existing in GSP-2, a partial acidic hydrolysis was conducted and produced a degraded polysaccharide GSP-2-P with a yield of 63.5%. Methylation analysis by GC-MS revealed that GSP-2-P, with a molecular weight of 25 kDa, was mainly composed of the (1→6)–linked glucopyranosyl and (1→4)–linked glucopyranosyl sugar residues. Compared with the original polysaccharide, the increased molar ratio of 1,6-linked Glcp and 1,4-linked Glcp indicated that these two residues are existing in the backbone of GSP-2. In the meanwhile, the significant decrease of the molar ratio of 1,3,6-linked-Glcp and 1,3-linked Glcp revealed that 1,3-linked Glcp is located on the side chain position, the backbone of GSP-2 has branches at O-3 (not O-6) position of 1,3,6- linked-Glcp.
NMR analysis
In the 1H NMR spectrum of GSP-2, due to the overlap of the D2O (δ 4.70-4.78), only one group of anomeric proton signals can be identified at δ 4.40-4.45. In the high field region, a group of sugar ring proton signals are observed at the field of δ 3.00 to δ 4.30 (Fig. 1C). In the 13C NMR spectrum of GSP-2, only one group of irresoluble anomeric signals are shown at δ103.3, in the high field region, a group of sugar ring carbon signals are shown in the field of δ 60.0 to δ 85.0 (Fig. 1D, 1E). As shown in 1H and 13C NMR spectra of all the five glucopyranosyl residues (Res A-E), the appearance of the anomeric carbon signals at δ 103.3 and proton signals ranged from δ 4.45 to δ4.72 revealed that all the residues are of β anomeric configuration [25]-[26].
To attribute the 1H and 13C NMR signals of GSP-2, a HSQC technique was employed to identify the correlation between the proton and carbon signals. From the spectrum, two correlations can be found in the anomeric region of the HSQC spectrum, ie: δ 4.70-4.72 (1H)/δ 103.3 (13C) and δ 4.40-4.45 (1H)/δ 103.3 (13C), as showed in the Fig. 2A, these two correlation signals can be employed as the start point to completely assigned all the 1H and 13C signals of GSP-2. Based on the 2D NMR data (H-H COSY, NOESY and HMBC), as listed in the Table 3, it can be deduced that the C-3 of residues B and E, C-4 of residue C were downfield to δ 84.4 and δ 78.5 due to the α effect of glycosylation, for the same reason, the C-6 signals of residues D and E were downfield to δ 69.2, which also can be confirmed by the reversed signal in the DEPT spectrum of GSP-2 (Fig. 1E) [27]-[28].
Figure 2. 2D NMR spectrums and proposed structure of the polysaccharides from the fruiting bodies of G. sinense.

(A: HSQC spectrum, B: HMBC spectrum, C: NOESY spectrum, D proposed strcuture).
Table 3. 1H NMR and 13C NMR-chemical shifts of GSP-2 a , b .
| Glycosyl residue | H-1/C-1 | H-2/C-2 | H-3/C-3 | H-4/C-4 | H-5/C-5 | H-6a,H-6b/C-6 |
| β-D-Glcp-(1→ | 4.72 | 3.22 | 3.43 | 3.44 | 3.40 | 3.62, 3.82 |
| A | 103.3 | 73.8 | 75.8 | 70.2 | 76.0 | 60.9 |
| →3)-β-D- Glcp-(1→ | 4.70 | 3.42 | 3.65 | 3.35 | 3.38 | 3.62, 3.82 |
| B | 103.3 | 73.8 | 84.4 | 70.0 | 76.3 | 60.9 |
| →4)-β-D -Glcp-(1→ | 4.42 | 3.22 | 3.36 | 3.58 | 3.60 | 3.61, 3.80 |
| C | 103.3 | 74.0 | 73.9 | 78.5 | 75.1 | 60.9 |
| →6)-β-D-Glcp-(1→ | 4.41 | 3.22 | 3.43 | 3.44 | 3.58 | 3.77, 4.12 |
| D | 103.3 | 73.8 | 76.0 | 68.8 | 75.8 | 69.2 |
| →3,6)-β-D-Glcp-(1→ | 4.45 | 3.48 | 3.65 | 3.43 | 3.58 | 3.77, 4.12 |
| E | 103.3 | 73.9 | 84.4 | 68.8 | 76.0 | 69.2 |
Measured in 500/125 MHz, δ in ppm.
Values of 13C chemical shift were record with reference to TMS as internal standard.
In this study, the specific linkages of glycosyl residues were further determined by the HSQC, HMBC and NOESY (Fig. 2A-C) experiments. The inter-residue HMBC correlations from H-1 of Residue C ( C ) to C-6 of Residue E ( E ), and H-6 of Residue E to C-1 of Residue C, in collaboration with the inter-residue NOESY effects of H-1 (C) with H-6 (E), established the linkage of residues C-1 of Residue C to O-6 position of Residue E. The inter-residue HMBC correlations from H-1 of Residue E to C-6 of Residue D, and the H-6 of Residue D to C-1 of Residue E, in collaboration with the inter-residue NOESY effects of H-1 (E) with H-6 (D), established the linkage of residues C-1 of Residue E to O-6 position of Residue D. The inter-residue HMBC correlations from H-1 of Residue D to C-4 of Residue C, and the H-4 of Residue C to C-1 of Residue D, in collaboration with the inter-residue NOESY effects of H-1(D) with H-4 (C), established the linkage of residues C-1 of Residue D to O-4 position of Residue C. The observed HMBCs from H-1 (B) to C-3 (E), together with NOESY effect of H-1 (B) with H-3 (E), constructed the branch substitution located at C-3 of Residue E. Similarly, the HMBCs from H-1 (A) to C-3 (B), in combination with the corresponding inter-residue NOESY effects of H-1 (A) with H-3 (B), revealed that the terminal Residue A attached at C-3 of Residue B.
After comprehensive composition analysis, methylation analysis and NMR experiments, it thus can be concluded that GSP-2 is a protein-bound polysaccharide having a backbone composed of (1→4)- and (1→6)-linked β-D-glucopyranosyl residues, bearing the side chains composed of (1→3)- and terminal-linked β-D-glucopyranosyl residues at O-3 position of (1→6)-linked β-D-glucopyranosyl residues in the backbone (Fig. 2D).
Immunomodulatory effect
As shown in 3H thymidine incorporation assay, GSP-2 significantly stimulated the proliferation of the mouse splenocyte in a dose-dependent manner from 10 to100 µg/mL for 48 h. The influence induced by the accidently contamination of the endotoxin was excluded by adding a specific inhibitor of lipopolysaccharide (Polymixin B, PMB, 10 µg/mL) to the medium (Fig. 3A). Further study on isolated cells indicated that both GSP-2 (30 µg/mL) and LPS (2 µg/mL) induced vigorous proliferation of mouse splenic B cells, while GSP-2 showed no effect on splenic T cells (Fig. 3B).
Figure 3. Stimulating effect of GSP-2 on the proliferation of the mouse splenocytic B and T cells.
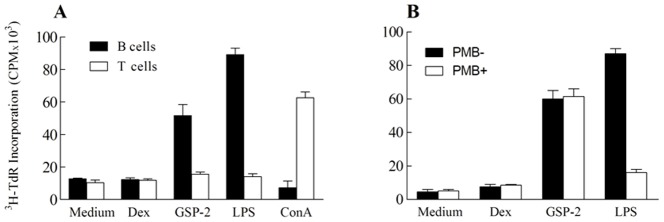
Freshly fractionated splenocyte, splenic B (solid bars) and T (open bars) cells were stimulated with, or without (Medium), dextran or GSP (30 µg/ml). LPS (2 µg/ml) and ConA (2 µg/ml) were included as controls. In a parallel experiment, mouse splenocytes were stimulated with, or without (Medium), GSP (30 µg/ml), dextran (30 µg/ml) or LPS (10 µg/ml) in triplicate wells in the presence, or absence, of PMB. 3H-TdR was added to the cultures for the last 8 hrs of incubation and then 3H-TdR incorporation (CPM) of each well counted. A: Parallel experiment to exclude the influence of the endotoxin contamination; B: Stimulating effect of GSP-2 to the mouse splenocytic B and T cells. All results are presented as mean ± SEM, *, P<0.05; **, P<0.01; ***, P<0.001 for difference from culture without treatment. (n = 9, repeated 3 times).
The immunomodulatory activities GSP-2 were also evaluated in mouse macrophage like cell line (RAW 264.7). As shown in Fig. 4A, GSP-2 significantly stimulated the production of nitric oxide (NO) in RAW 264.7 cells in a dose-dependent manner. As well known that NO is an important chemokine secreted by macrophage that can oxidize the pathogens, the secretion of NO is related to the swallowing of the cells to the pathogens, such as bacteria and virus [29] Some biological macromolecules, such as polysaccharides also can be identified as non-self and phagocytosed by macrophages, and the cell will further degrade the polysaccharides to small deribs and present to the other immune cells. In order to investigate the phagocytosis of the macrophages to our polysaccharide fraction, a fluorescent tag was specifically labeled on the reducing end of polysaccharide GSP-2 backbones, and then added to the macrophage culture medium. The cells were visualized and imaged under inverted fluorescent microscope after 1 h and 24 h (Fig. 4B). The number of fluorescent-labeled cells was increased after 24 h incubation with labeled GSP-2. The fluorescence of the labeled and unlabeled cell samples was also detected by flow cytometry as shown in Fig. 4C. These results demonstrated that polysaccharide GSP-2 could be phagocytosed by RAW 264.7 cells, in which biological response (such as NO production) was induced.
Figure 4. Nitric oxide production and phagocytosis of GSP-2-treated RAW 264.7 cells.

A: The levels of NO production of RAW264.7 cells were assessed after incubation with GSP-2 or LPS for 24 h using Griess assay. All data are expressed as mean ± SEM of three individual experiments (n = 12). Significant difference: *, P<0.05; **, P<0.01; ***, P<0.001 for difference from culture without treatment. B: The viable RAW 264.7 cell populations were gated (left) and the FITC-positive population was shown in fluorescent-GSP-2-labeled cells (lower right histogram). C: RAW 264.7 cells were incubated with 10 µg GSP-2 (fluro-labeled) for 24 h. The cells were imaged at the 1st and 24th hour (lower histograms). After 24 h incubation, the cells were collected, washed and resuspended in PBS and the fluorescence of the samples was detected by flow cytometry. (Upper histograms: viable RAW 264.7 cells without fluorescent labeled GSP-2, lower histograms, 1st hour: viable RAW 264.7 cells with fluorescent labeled GSP-2, 24th hour).
It is suggested that GSP-2 is a typical B cell and macrophage dependent immune-modulator. Polysaccharides from medicinal materials could exhibit different immunopotentiating activities. For example, polysaccharides found in the roots of Acanthopanax koreanum, Acanthopanax senticosus, Platyloden grandiflorum and angelan are also able to activate mouse B cells and macrophages but not T cells B cells [30]-[31]. Polysaccharides from P. linteus can stimulate B cells, T cells and macrophages [32], while lentinan and β(1→3)-glucan isolated from Lentinus edodes is a stimulator of T cells and macrophages, but not B cells [33]. The relationship between polysaccharide origin, structure and their immuno-modulation activity remains to be further characterized.
Regarding the receptor of these immune-activity polysaccharides on B cells and macrophages, some have been generally acknowledged, like mannose receptor, CR3 (CD11b/CD18), lactosylceramide, dectin-1, Toll-like receptors (TLR) and so on. Polysaccharides from the roots of Acanthopanax koreanum and Platyloden grandiflorum stimulate B cells through TLR2, TLR4 and also CD19 and CD79b [31], [34]. Ando et al reported that safflower polysaccharides activated NF-κB signaling pathway via TLR4 in macrophages [35]. Polysaccharides may employ molecules such as CD14, RP105, MD-1 or MD2 as a bridge to indirectly interact with TLR4. Ongoing experiments in our laboratory will search for potential receptor of GSP-2 on B cells and macrophages. Apart from the immuno-stimulating effects of GSP-2 in mouse immune cells, the immunomodulatory activities of GSP-2 in human PBMCs and moDCs were also elucidated so that the clinical potential of the polysaccharide could be revealed. As shown in Fig. 5, the productions of TNF-α, IL-1β, IL-12 and GM-CSF were increased dose-dependently in GSP-2-treated PBMCs. Besides, GSP-2 could also stimulate the IL-10 and IL-12 productions of human monocyte-derived dendritic cells in a dose dependent manner (Fig. 6). Dendritic cells (DCs) are the antigen presenting cells that initiate and direct adaptive immune responses, capable of inducing protective adaptive immune responses [36]. And IL-10 and IL-12 are two important mediators secreted by DCs: IL-10 is a pleiotropic cytokine with anti-inflammatory and immunosuppressive properties [37], which can act as a feedback regulatory role to constrict the immune response when DC is activated [38]; And IL-12 can polarizes the immune system toward a primary Th1 cell response [38]–[39]. These result indicated that GSP-2 could robust the response of the dentritic cells to the pathogens.
Figure 5. Cytokine productions of GSP-2-treated PBMCs.

Culture supernatants were collected 24-2 and the cytokines concentrations were specifically determined by ELISA. All data are expressed as mean ± SEM of three individual experiments (n = 12). Differences between the treated and untreated control groups were compared using one-way ANOVA. * P<0.05, ** P<0.01, *** P<0.001 for difference from culture without treatment.
Figure 6. Cytokine productions of GSP-2-treated moDC.

Culture supernatants were collected 48-2 and the cytokines concentrations were specifically determined by ELISA. Lines represented mean percentage ± S.E.M. of duplicates (n = 8). LPS and Beta-glucan from E. gracilis were used as positive control. Significant difference: *, P<0.05; **, P<0.01; ***, P<0.001 for difference from culture without treatment.
It is know that the beta glucan from different source might own different structure. Beta-glucan from oat are of a linear mainly structure of 1,4-β-Glcp residues, while mushroom sourced beta glucan often bear a backbone structure of 1,3-β-Glcp residues or 1,6-β-Glcp residues, and branched by the side chain of 1,3-β-Glcp residues. In this study, GSP-2 has a very novel backbone structure of 1,6-, 1,4-β-Glcp residues. As GSP-2 is highly water-soluble, we suppose that it might share some similar characters with the well studied yeast water-soluble beta-glucan (PGG), which could be identified by the toll-like receptor-4 on the surface of the cells. The influence of the structural factors such as the length of the backbone, degree of polymerization, degree of branching and the length of the side chain to the interaction between this fraction and the toll-like receptors deserve further studies [40].
In summary, a water-soluble protein-bound glucan GSP-2 was isolated from the medical mushroom G. sinense, with an apparent molecular weight of 32 kDa. Structural analysis revealed that GSP-2 contains a backbone composed of (1→4)- and (1→6)-linked β-D-glucopyranosyl residues, bearing the side chains of (1→3)- and terminal-linked β-D-glucopyranosyl residues at O-3 position of (1→6)-linked β-D-glucopyranosyl residues in the backbone. In addition, GSP-2 was found to be a B cell dependent immunomodulator without any in vitro stimulating effect to the mouse splenetic T cells, and could significantly stimulating cytokine secretion from PBMCs and derived dendritic cells. More interestingly, the fluorescent labeled GSP-2 could be phagocytosis by mouse macrophage like RAW264.7 cells, in the meanwhile, enhanced the nitric oxide production of the cells, suggesting that the endocytosis function of the innate immune cells might partially explain the absorption of protein-bound polysaccharides.
Supporting Information
ARRIVE Checklist.
(DOC)
Funding Statement
The chemical research work of this study is funded by the Innovation and Technology Fund (ITS/311/09 and InP/108/10) of the Government of the Hong Kong Special Administrative Region, Hong Kong Baptist University (FRG2/11-12/048, FRG1/12-13/018, FRG2/12-13/006, and RC-start up grant). The immune study is funded by the National Natural Science Foundation of China (81303210, 81102253). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Russell R, Paterson M (2006) Ganoderma-A therapeutic fungal biofactory. Phytochemistry 67: 1985–2001. [DOI] [PubMed] [Google Scholar]
- 2. Sliva D (2006) Ganoderma lucidum in cancer research. Leuk Res 30: 767–768. [DOI] [PubMed] [Google Scholar]
- 3.Ito H, Naruse S, Shimura K (1977) Studies on antitumor activity of basidiomycete polysaccharides. Vii. Antitumor effect of the polysaccharide preparations from Ganoderma lucidum on mouse sarcoma 180. Subject Strain Bibliography, Paper 1188. Available: http://mouseion.jax.org/ssbb1977/1188.
- 4. Ooi VEC, Liu F (1999) A review of pharmacological activities of mushroom polysaccharides. Int J Med Mushr 1: 195–206. [Google Scholar]
- 5. Nie SP, Zhang H, Li WJ, Xie MY (2013) Current development of polysaccharides from Ganoderma. Isolation, structure and bioactivities. Bioact. . Carbohydr Diet Fibr 1: 10–20. [Google Scholar]
- 6. Da J, Wu WY, Hou JJ, Long HL, Yao S, et al. (2012) Comparison of two officinal Chinese pharmacopoeia species of Ganoderma based on chemical research with multiple technologies and chemometrics analysis. J Chromatogr A 1222: 59–70. [DOI] [PubMed] [Google Scholar]
- 7. Liu JQ, Wang CF, Li Y, Luo HR, Qiu MH (2012) Isolation and bioactivity evaluation of terpenoids from the medicinal fungus Ganoderma sinense. Planta Med 78: 368–376. [DOI] [PubMed] [Google Scholar]
- 8. Cheng CH, Leung AY, Chen CF (2010) The effects of two different ganoderma species (Lingzhi) on gene expression in human monocytic THP-1 cells. Nutr Cancer 62: 648–658. [DOI] [PubMed] [Google Scholar]
- 9. Yue GG, Fung KP, Tse GM, Leung PC, Lau CB (2006) Comparative studies of various ganoderma species and their different parts with regard to their antitumor and immunomodulating activities in vitro. J Altern Complement 12: 777–789. [DOI] [PubMed] [Google Scholar]
- 10. Chen XP, Chen Y, Li SB, Chen YG, Lan JY, et al. (2009) Free radical scavenging of Ganoderma lucidum polysaccharides and its effect on antioxidant enzymes and immunity activities in cervical carcinoma rats. Carbohydr Polym 77: 389–393. [Google Scholar]
- 11. Lv GP, Zhao J, Duan JA, Tang YP, Li SP (2012) Comparison of sterols and fatty acids in two species of Ganoderma. Chem Cent J 6: 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Sato N, Zhang Q, Ma CM, Hattori M (2009) Triterpene-Farnesyl hydroquinone conjugates from Ganoderma sinense. . Chem Pharm Bull 57: 1076–1080. [DOI] [PubMed] [Google Scholar]
- 13. Yue GG, Chan BC, Han XQ, Cheng L, Wong EC, et al. (2013) Immunomodulatory Activities of Ganoderma sinense Polysaccharides in Human Immune Cells. Nutr Cancer 65: 765–774. [DOI] [PubMed] [Google Scholar]
- 14. Han XQ, Chan BC, Yu H, Yang YH, Hu SQ, et al. (2012) Structure characterization, and immunomodulating activity of a hyperbranched polysaccharide from the fruiting bodies of Ganoderma sinense . Int J Biol Macromol 51: 597–603.22750578 [Google Scholar]
- 15. Han XQ, Yu H, Chan BC, Dong CX, Yang YH, et al. (2012) Isolation, structure characterization and immunomodulating activity of a hyperbranched polysaccharide from the fruiting bodies of Ganoderma sinense. . J Agric Food Chem 60: 4276–4281. [DOI] [PubMed] [Google Scholar]
- 16. Mandal EK, Maity K, Maity S, Gantait SK, Behera B, et al. (2012) A heteropolysaccharide from aqueous extract of an edible mushroom, Pleurotus ostreatus cultivar: structural and biological studies. Carbohydr Res 347: 172–177. [DOI] [PubMed] [Google Scholar]
- 17. Bhanja SK, Nandan CK, Mandal S, Bhunia B, Maiti TK, et al. (2012) Isolation and characterization of the immunostimulating β-glucans of an edible mushroom Termitomyces robustus . Carbohydr Res 357: 83–89. [DOI] [PubMed] [Google Scholar]
- 18. Maji PK, Sen IK, Devi KSP, Maiti TK, Sikdar SR, et al. (2013) Structural elucidation of a biologically active heteroglycan isolated from a hybrid mushroom of Pleurotus florida and Lentinula edodes . Carbohydr Res 368: 22–28. [DOI] [PubMed] [Google Scholar]
- 19. Lawrence JF, Lyengar JR (1985) Gas chromatographic determination of polysaccharide gums in foods after hydrolysis and derivatization. J Chromatogr 350: 237–244. [DOI] [PubMed] [Google Scholar]
- 20. Needs PW, Selvendran RR (1993) Avoiding oxidative degradation during sodium hdroxide/methyl iodide-mediated carbohydrate methylation in dimethyl sulfoxide. Carbohydr Res 245: 1–10. [Google Scholar]
- 21. Sweet DP, Shapiro RH, Albersheim P (1975) Quantitative analysis by various g.l.c. response factor theories for partially methylated and partially ethylated alditol acetates. Carbohydr Res 40: 217–225. [Google Scholar]
- 22. Rice PJ, Adams EL, Ozment-Skelton T, Gonzalez AJ, Goldman MP, et al. (2005) Oral delivery and gastrointestinal absorption of soluble glucans stimulate increased resistance to infectious challenge. J Pharmacol Exp Ther 314: 1079–1086. [DOI] [PubMed] [Google Scholar]
- 23. Jiménez E, Garcia-Lora A, Martinez M, Garrido F (2005) Identification of the protein components of protein-bound polysaccharide (PSK) that interact with NKL cells.Cancer Immunol Immunother. 54: 395–399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Staub AM (1965) Removal of protein-Sevag method. Methods Carbohydr Chem 5: 5–6. [Google Scholar]
- 25. Dey B, Bhunia SK, Maity KK, Patra S, Mandal S, et al. (2010) Heteroglycan from an alkaline extract of a somatic hybrid mushroom (PfloVv1aFB) of Pleurotus florida and Volvariella volvacea: structural characterization and study of immunoenhancing properties. Carbohydr Res 345: 2736–2741. [DOI] [PubMed] [Google Scholar]
- 26. Mandal S, Maity KK, Bhunia SK, Dey B, Patra S, et al. (2010) Chemical analysis of new water-soluble (1→6)-, (1→4)-α, β-glucan and water-insoluble (1→3)-, (1→4)-β-glucan (Calocyban) from alkaline extract of an edible mushroom, Calocybe indica (Dudh Chattu). Carbohydr Res 345: 2657–2663. [DOI] [PubMed] [Google Scholar]
- 27. Bhunia SK, Dey B, Maity KK, Patra S, Mandal S, et al. (2011) Isolation and characterization of an immunoenhancing glucan from alkaline extract of an edible mushroom, Lentinus squarrosulus (Mont.) Singer. Carbohydr Res 346: 2039–2044. [DOI] [PubMed] [Google Scholar]
- 28. Ojha AK, Chandra K, Ghosh K, Islam SS (2010) Glucans from the alkaline extract of an edible mushroom, Pleurotus florida, cv Assam Florida: isolation, purification, and characterization. Carbohydr Res 345: 2157–63. [DOI] [PubMed] [Google Scholar]
- 29. Liu YW, Gao JL, GuanJ, Qian ZM, Feng K, et al. (2009) Evaluation of antiproliferative activities and action mechanisms of extracts from two species of Ganoderma on tumor cell lines. J Agric Food Chem 57: 3087–3093. [DOI] [PubMed] [Google Scholar]
- 30. Han SB, Yoon YD, Ahn HJ, Lee HS, Lee CW, et al. (2003) Toll-like receptor-mediated activation of B cells and macrophages by polysaccharide isolated from cell culture of Acanthopanax senticosus. Int Immunopharmacol 3: 1301–12. [DOI] [PubMed] [Google Scholar]
- 31. Han SB, Park SH, Lee KH, Lee CW, Lee SH, et al. (2001) Polysaccharide isolated from the radix of Platycodon grandiflorum selectively activates B cells and macrophages but not T cells. Int Immunopharmacol 1: 1969–78. [DOI] [PubMed] [Google Scholar]
- 32. Kim HM, Han SB, Oh GT, Kim YH, Hong DH, et al. (1996) Stimulation of humoral and cell mediated immunity by polysaccharide from mushroom Phellinus linteus. Int J Immunopharmacol 18: 295–303. [DOI] [PubMed] [Google Scholar]
- 33. Liu M, Li J, Kong F, Lin J, Gao Y (1998) Induction of immunomodulating cytokines by a new polysaccharide-peptide complex from culture mycelia of Lentinus edodes. Immunopharmacology 40: 187–98. [DOI] [PubMed] [Google Scholar]
- 34. Han SB, Park SK, Ahn HJ, Yoon YD, Kim YH, et al. (2003) Characterization of B cell membrane receptors of polysaccharide isolated from the root of Acanthopanax koreanum. Int Immunopharmacol 3: 683–91. [DOI] [PubMed] [Google Scholar]
- 35. Ando I, Tsukumo Y, Wakabayashi T, Akashi S, Miyake K, et al. (2002) Safflower polysaccharides activate the transcription factor NF-kappa B via Toll-like receptor 4 and induce cytokine production by macrophages. Int Immunopharmacol 2: 1155–62. [DOI] [PubMed] [Google Scholar]
- 36. Rout D, Mondal S, Chakraborty I, Islam SS (2008) The structure and conformation of a water-insoluble (1→3)-,(1→6)-β-D-glucan from the fruiting bodies of Pleurotus florida. Carbohydr Res 343: 982–987. [DOI] [PubMed] [Google Scholar]
- 37. Delamarre L, Mellman I (2011) Harnessing dendritic cells for immunotherapy. Semin Immunol 23: 2–11. [DOI] [PubMed] [Google Scholar]
- 38. Lin YL, Liang YC, Tseng YS, Huang HY, Chou SY, et al. (2009) Reishi polysaccharides induce immunoglobulin production through the TLR4/TLR2-mediated induction of transcription factor blimp-1. J Leukoc Biol 86: 877–889.19498044 [Google Scholar]
- 39. Chan CFG, Chan WK, Sze MYD (2009) The effects of β glucan on human immune and cancer cells. J Hematol Oncol 25: 1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Liu JJ, Gunn L, Hansen R, Yan J (2009) Combined yeast-derived β glucan with anti-tumor monoclonal antibody for cancer immunotherapy. Exp Mol Pathol 86: 208–214. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
ARRIVE Checklist.
(DOC)