

Abstract
Norovirus (NoV) is an important cause of nosocomial gastroenteric outbreaks. This 5-month study was designed to characterize NoV contamination and airborne dispersal in patient rooms during hospital outbreaks. Air vents, overbed tables, washbasins, dust, and virus traps designed to collect charged particles from the air were swabbed to investigate the possibility of NoV contamination in patient rooms during outbreaks in seven wards and in an outbreak-free ward. Symptomatic inpatients were also sampled. Nucleic acid extracts of the samples were examined for NoV RNA using genogroup I (GI) and GII real-time reverse transcription-PCR (RT-PCR). The NoV strains were characterized by RT-PCR, sequencing, and phylogenetic analysis of the RNA-dependent RNA-polymerase-N/S capsid-coding region (1,040 nucleotides [nt]). Patient strains from two outbreaks in one ward were sequenced across the RNA-dependent-RNA-polymerase major capsid-coding region (2.5 kb), including the hypervariable P2 domain. In the outbreak wards, NoV GII was detected in 48 of 101 (47%) environmental swabs and 63 of 108 patients (58%); NoV genotype II.4 was sequenced from 18 environmental samples, dust (n = 8), virus traps (n = 4), surfaces (n = 6), and 56 patients. In contrast, NoV GII was detected in 2 (GII.4) of 28 (7%) environmental samples and in 2 (GII.6 and GII.4) of 17 patients in the outbreak-free ward. Sequence analyses revealed a high degree of similarity (>99.5%, 1,040 nt) between NoV GII.4 environmental and patient strains from a given ward at a given time. The strains clustered on 11 subbranches of the phylogenetic tree, with strong correlations to time and place. The high nucleotide similarity between the NoV GII.4 strains from patients and their hospital room environment provided molecular evidence of GII.4 dispersal in the air and dust; therefore, interventional cleaning studies are justified.
INTRODUCTION
Norovirus (NoV) is a common cause of recurring nosocomial outbreaks of acute gastroenteritis and can inflict potentially life-threatening infections in the immunocompromised, the elderly, and newborns (1–3). Characterized by the sudden onset of projectile vomiting and diarrhea, NoV infections are usually of short duration in healthy individuals. These small, nonenveloped, noncultivable, genomically diverse, and single-stranded positive-sense RNA viruses with an approximately 7.5-kb genome are highly infectious and remarkably robust in different environmental settings (4–6). On ingestion, NoV passes through the gastric juices, multiplies in the small intestine, and is excreted in the feces at high concentrations (7). Therefore, NoV is readily transmitted by the fecal-oral route: through hand-to-hand contact, fecal contamination of food and water (8–10), and also, importantly, by aerosol dispersal from vomitus, with subsequent heavy contamination of the surrounding environment (11, 12).
The significance of such contamination is strongly indicated in reports in which NoV RNA was detected in environmental samples from hospitals (13), cruise ships (14, 15), restaurants (16), and houseboats (17) during recurring or ongoing outbreaks. The high viral load in vomitus and feces (7), low infectious dose (18), and short-term immunity (19) that typify NoV gastroenteritis increase the risk of prolonged outbreaks and environmental spread in semiclosed units, in which strains of NoV genotype II.4 (GII.4) or GII.3 tend to dominate (20). With no effective antiviral or vaccine therapy to counteract infection and transmission of NoV GII strains, preventative measures are based on strict hygiene control and isolation of infected patients (21).
Although contamination from aerosols derived from vomitus is strongly suggested in reports of NoV outbreaks affecting hotel restaurant diners (16), airplane travelers and crew (22), and nursing staff (23), the absence of a suitable culture system makes it difficult to demonstrate the presence of infectious virus in the air or other environmental samples (4). However, advances in molecular analyses permit a more detailed examination of such samples for NoV RNA (14). In this study of the environmental distribution of NoV during hospital outbreaks, molecular methods were used to investigate the nature of NoV contamination, including airborne dispersal of NoV in dust from patient rooms, as well as the molecular epidemiology of the outbreaks.
MATERIALS AND METHODS
Study outline.
Patient and environmental sampling were carried out in eight wards in four separate buildings of Sahlgrenska University Hospital, Gothenburg, Sweden (Table 1). Infection control nurses provided ward outbreak reports throughout the 5-month study. The ward nursing staff members were also investigated in studies to be reported elsewhere.
TABLE 1.
Norovirus infections in hospital patients and environmental contamination in eight wards and an administration office room during nosocomial outbreaks
| Building | Ward | Floor | Care unit | Patient samples |
Environmental samples |
||||
|---|---|---|---|---|---|---|---|---|---|
| Total no. (n = 125)a | No. of NoV GII-positive samples (n = 65)b | Median NoV GII CT (CT range) (IQR)c | Total no. (n = 135)a | No. of NoV GII-positive samples (n = 50)b | Median NoV GII CT (CT range) (IQR)c | ||||
| A | A1d | 5 | Hematology/immunosuppression | 17 | 12 | 20.6 (15.9–27.6) (7.25) | 26 + 10e | 12 + 10e | 33.1 (25.4–39.0) (4.6) |
| A2d | 5 | Medical | 12 | 5 | 22.6 (17.7–36.8) (22.6) | 12 | 1 | 29.2 | |
| B | B1 | 7 | Geriatric general medical | 32 | 13 | 19.3 (13.0–30.6) (9) | 10 | 5 | 30.6 (29.5–34.1) (2.8) |
| B2 | 6 | Geriatric orthopedic | 9 | 8 | 17.5 (14.2–26.7) (8.3) | 10 | 5 | 33.2 (31.1–38.6) (5.4) | |
| B3 | 6 | Geriatric general medical | 14f | 7 | 22.1 (18.2–28.8) (8.1) | 12 | 3 | 36.2 (31.2–39.5) (8.3) | |
| C | C1 | 6 | Dementia | 12 | 9 | 23.6 (16.4–37.8) (5.5) | 9 | 7 | 33.7 (31.2–39.5) (2.5) |
| C2 | 4 | Substance abuse and dementia | 12 | 9 | 20.6 (16.4–33.8) (6.6) | 12 | 5 | 37.7 (37.1–39.5) (1.8) | |
| D | Dg | 2 | Cardiology/lung transplant | 17 | 2 | 27.8 (20.9–34.7) (13.8) | 18 + 10e | 2e | 32.0 (28.2–35.8) (7.5) |
| E | 2 | Administration office | NAh | NA | NA | 6 | 0 | NA | |
Total number of samples examined in real-time RT-PCR assays designed for the detection of human norovirus GI and GII, adenovirus, astrovirus, rotavirus, and sapovirus.
Number of samples with a norovirus genogroup II (NoV GII)-positive real-time RT-PCR result.
CT, cycle threshold value in NoV GII-specific real-time RT-PCR assay; IQR, interquartile range.
Interconnected wards with shared dining room and shower facilities.
Denotes dust samples.
Includes one patient with acute gastroenteritis testing positive for rotavirus in real-time RT-PCR.
Outbreak-free ward with no nosocomial infections.
NA, not applicable.
In this study of NoV hospital infections and the dispersal of NoV in patient rooms, an outbreak ward was defined as a ward in which two or more cases of suspect or verified NoV gastroenteritis occurred in patients and the infection spread within the ward (24). Patient rooms were predominantly two- to four-bed rooms with communal patient toilets in the ward corridors; these toilets were accessible to visitors. Each ward also had two to four single rooms with individual toilets.
The sampling of symptomatic inpatients and their hospital room environment began in January 2012, the onset of seasonal nosocomial NoV outbreaks, and continued through May (see Table S1 in the supplemental material). Seven outbreak wards and one outbreak-free ward with two isolated cases of acute NoV gastroenteritis occurring 7 weeks apart were examined. As a control, an administration office room in another hospital building (E) with no wards was sampled for the presence of airborne virus particles (Table 1). The environmental and patient samples were processed separately, at different time points, and in two different laboratories dedicated to environmental or patient sample preparation. Strict precautions were taken to prevent cross-contamination of the samples and PCR products, including the use of UV work stations, dedicated pipettes, and water controls (25).
Patient samples.
Feces and/or vomitus were obtained from inpatients (n = 125) who exhibited routine diagnostic indications of acute gastroenteritis. A total of 125 patient samples were examined, with 108 from outbreak wards and 17 from an outbreak-free ward (Table 1). Suspensions (10%) were prepared in phosphate-buffered saline (PBS) (pH 7.4) (26), and 250 μl was mixed with 2 ml of NucliSENS lysis buffer (bioMérieux, Marcy-l'Étoile, France) before total nucleic acid (TNA) extraction (110 μl) on the NucliSENS easyMAG extractor (bioMérieux).
Environmental samples.
One investigator used standardized procedures to collect samples (total n = 135) (Table 1) from the eight wards and the administration office room. Sterile cotton-tipped swabs prewetted with PBS were drawn back and forth once across a 25-cm surface; circular sweeps were used to swab the washbasin drains and air vents. The swabs were transported to the laboratory in 2 ml NucliSENS lysis buffer and vortexed vigorously prior to TNA extraction (110 μl) on the NucliSENS easyMAG extractor.
Swabs (n = 91) from overbed tables, washbasins, and air vents were collected from patient rooms in eight wards (Table 1). Dust was sampled from rooms in two wards in May, in outbreak ward A1 and outbreak-free ward D (Table 1). One dust swab per room (n = 10) was collected from the locker tops in nine patient rooms and the nurses' office in ward A1. Dust (n = 10) was also sampled from the locker tops in 10 patient rooms in ward D.
Virus trap sampling (VTs) was based on an ionizer prototype manufactured by Airpoint Ab, Stockholm, Sweden. The device was modified by installing a disposable plastic cup with a 47-mm-diameter conductive surface as the collector plate (GP Plastic, Gislaved, Sweden). At 12 V, the ionizer generates negative ions, and surface molecules of virus particles in the air become negatively charged and are thus attracted to the positively charged collector plate. The ionizer was placed on a shelf above the bed 2 to 3 m from the patient's head and exposed for 3 h at room temperature. The sample cup was washed with PBS (1 ml) and rubbed gently with a sterile swab. PBS aliquots (250 μl) were transferred to 2 ml NucliSENS lysis buffer prior to TNA extraction (110 μl). VTs (n = 12) were collected from six patient rooms, two samples per room, in four outbreak wards: A1, B1, B2, and B3 (see Table S1 in the supplemental material). Three patient rooms in the outbreak-free ward, D, were also examined by VTs (n = 6). Control VTs (n = 6) was carried out in an administration office in building E (with no wards) on three consecutive days, with two samples per day, during the study period.
Real-time reverse transcription-RT-PCR.
Validated real-time reverse transcription RT-PCR (rRT-PCR) assays were used to detect NoV GI and NoV GII, rotavirus (RoV), human astrovirus (HuAstV) (6, 27), human sapovirus (HuSaV) (28), and human adenovirus (HuAdV) (29) in duplicate TNA extracts (10 μl) of environmental and patient samples, as described previously (6, 27). The rRT-PCR controls included virus-positive feces and a plasmid construct designed to control viral amplification. Cycle threshold (CT) values of ≤39.5 were recorded as positive.
NoV classic RT-PCR and nucleotide sequencing.
NoV rRT-PCR-positive samples were amplified in seminested RT-PCR of the NoV RNA-dependent RNA-polymerase (RdRp)-N/S capsid-coding region (1,040 nucleotides [nt]) as described previously for NoV GII strains (9). The amplicons were purified and cycle sequenced in unilateral master mixes with NoV GII primers and BigDye kits (version 3.1 or version 1.3; Applied Biosystems, Foster City, CA) (9).
NoV GII.4 patient strains from two outbreaks in ward A1 (in January and May) were amplified and sequenced across the RdRp-N/S major capsid-coding region (2.5 kb), as described for long-fragment analysis of NoV GII strains (9). This region of the NoV GII genome included the partial RdRp of open reading frame 1 (ORF1), the entire ORF2 encoding the major capsid protein, and the partial minor capsid protein-coding region in ORF3 (2.5 kb).
Comparative sequence and phylogenetic analyses.
The NoV sequences were analyzed and aligned with GenBank reference strains by using Sequencher 4.9 (Gen Codes Corp., Ann Arbor, MI) and Basic Local Alignment Search Tool (BLAST) (NCBI). Phylogenetic analysis included neighbor-joining methods using the MacVector 7.2 software (Accelrys, Inc., San Diego, CA).
Nucleotide sequence accession numbers.
The sequences described in the study have been deposited in GenBank under accession no. KF768469 to KF768544.
RESULTS
Outbreaks of acute gastroenteritis occurred in seven of eight wards monitored for nosocomial NoV infections and environmental contamination during the 5-month study (Table 1). Samples from inpatients and their hospital room environment were examined in rRT-PCR assays designed for laboratory diagnosis of enteric virus infections (6, 27). The NoV GII detection results according to building, ward, patient, room, day of outbreak sampling, CT values, and sequence-based genotyping are detailed in Table S1 in the supplemental material.
Patients.
NoV GII infections were detected in 63 of 108 (58%) patients sampled in the outbreak wards (CT range, 13.0 to 37.8; median, 20.0; interquartile range [IQR], 5.3). Sequence-based genotyping revealed NoV GII.4 infections in 56 of the 63 NoV GII-positive patients; seven strains could not be sequenced because of poor amplification or inadequate sample material (see Table S1 in the supplemental material). One case of RoV gastroenteritis was detected in ward B3.
In outbreak-free ward D, NoV infection was detected in 2 of 17 patients (12%) (Table 1). The first patient (P27; GII CT, 34.7) developed NoV gastroenteritis within 24 h of admission in mid-March (see Table S1 in the supplemental material). NoV strain P27 was found through sequencing to belong to GII.6. The short time delay between admission and onset of diarrheal symptoms indicated that this patient might have been admitted early in the acute stage of infection. No NoV infections were recorded in ward D prior to the study period, and no secondary spread was noted between March and May. The second case, P61, occurred early in May, 49 days after the first. Strain P61 (CT, 20.9) was found through sequencing to belong to GII.4. No further cases of NoV gastroenteritis were reported from ward D.
The epicurve showed the onset of NoV GII infections in outbreak wards A1, A2, B1, B2, B3, C1, and C2 during the 5-month study, including the single cases of NoV GII infections (GII.6 and GII.4) that occurred in outbreak-free ward D (Fig. 1). Evidence of NoV GII outbreaks over time was noted particularly for wards A1, A2, and B1. A1 outbreaks occurred on weeks 1 to 2 (January) and again on weeks 18 to 19 (May), A2 outbreaks occurred in late February and May, and B1 outbreaks were reported on four occasions, in January, February, March, and April.
FIG 1.
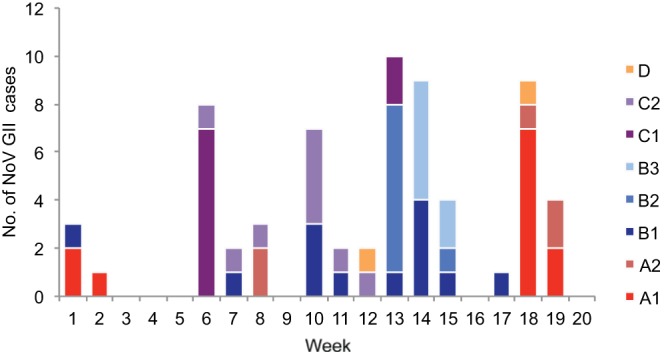
Epicurve showing distribution of NoV GII cases (n = 65) in eight wards, A1, A2, B1, B2, B3, C1, C2, and D, during seasonal nosocomial outbreaks, weeks 1 to 20, 2012. NoV GII.4 was detected in 57 of 58 NoV GII cases in which the strains were sequenced; the one exception was a case of GII.6 infection detected in outbreak-free ward D, week 12.
Environment.
NoV GII RNA was detected in 48 of 101 (47.5%) environmental swabs from seven outbreak wards (CT range, 25.4 to 39.5; median, 33.7; IQR, 4.4), and in 2 of 28 (7.1%) samples (CT, 28.0 and 35.8) from the outbreak-free ward.
NoV GII RNA (CT range, 26.7 to 39.5; median, 34.3; IQR, 5.1) was detected in 29 (37%) of 79 environmental swabs from overbed tables, washbasins, and air vents in the seven outbreak wards (A1, A2, B1, B2, B3, C1, and C2) (Table 1; see also Table S1 in the supplemental material). Sequence-based genotyping (1,040 nt) revealed NoV GII.4 strains in 6 of 10 rRT-PCR-positive swabs from wards B1 and C1. NoV was not detected in the surface samples (n = 12) from three rooms in the outbreak-free ward.
All dust samples (n = 10) collected on outbreak day 131 from 10 rooms (one sample per room) in ward A1 tested positive in NoV GII rRT-PCR (CT range, 25.4 to 35.8; median, 30.6; IQR, 5.5). Nine of these samples were from the patient rooms; the tenth was from the nurses' office. The results were verified by repeat extraction and assay of the dust. Sequencing revealed NoV GII.4 RNA (1,040 nt) in dust from 8 of the rooms, mixed with HuAstV contamination in 2 rooms (CT, 33.7 and 35.9), HuSaV in another 2 rooms (CT, 35.2 and 39.1), and HuAdV in 3 rooms (CT, 38.0, 38.0, and 38.9) (see Table S1 in the supplemental material).
NoV GII (CT, 28.0 and 35.8) was detected in dust from the locker tops in 2 of 10 rooms in the outbreak-free ward. These dust samples were collected on day 59 after the onset of symptoms in the first case of NoV gastroenteritis to occur in the ward (P27; NoV GII.6) and 10 days after the single case of GII.4 infection (P61). GII RNA detected in dust from these 2 rooms was found through sequencing (100 nt) to belong to GII.4; NoV GI (CT, 31.8) was also detected in dust from one of these rooms (see Table S1 in the supplemental material).
VTs (n = 12) from six patient rooms (two samples per room) in outbreak wards A1, B1, B2, and B3 were examined for NoV RNA (see Table S1 in the supplemental material). Nine of 10 VTs from five of these rooms were positive in NoV GII rRT-PCR assays (CT range, 29.5 to 39.0; median, 32.1; IQR, 6.6). Sequencing revealed NoV GII.4 RNA (1,045 nt) in the VTs from four of these rooms. The patients in these five rooms had symptomatic laboratory-confirmed NoV infections on the day of VT sampling. The VTs (n = 2) from the sixth room were NoV rRT-PCR negative. The patients in this room (R12 in ward B3) were symptom-free on the day of VT sampling (see Table S1).
VTs (n = 6) from three patient rooms in outbreak-free ward D tested negative for NoV when sampled after point cleaning with 1% Rely+On Virkon (DuPont, United Kingdom), following patient discharge (24). One of these rooms had been occupied by P27, a patient with NoV GII.6 infection (see Table S1 in the supplemental material). VT controls (n = 6) from the administration office in building E, which had no wards, were NoV rRT-PCR negative.
Comparative sequence and phylogenetic analyses.
Sequence analyses of the RdRp-N/S capsid-coding region (1,040 nt) showed a predominance of NoV GII.4 strains in the environmental and patient samples from the seven outbreak wards. The NoV GII.4 sequences detected in environmental swabs and patient samples collected from a given ward during an ongoing outbreak were identical or had a high degree of similarity (>99.5%, 1,040 nt), (Fig. 2). This level of similarity also held on analyses of the RdRp-N/S major capsid-coding region (2.5 kb) and of the 484-nt hypervariable P2 subdomain in patient strains from each of the ward A1 outbreaks (January and May).
FIG 2.

Neighbor-joining tree based on nucleotide analysis of the RNA-dependent RNA polymerase N/S capsid-coding region of the NoV genome (1,040 nt). P, patient strain sequence; A1, A2, B1, B2, B3, C1, C2, D, wards; R, patient room; Rx, patient room not defined; VT, virus trap; ptab, patient overbed table; whb, washbasin; airv, air vent. Hospital buildings are represented by four colors: red, building A; blue, building B; purple, building C; orange, building D. Wards are indicated by the symbols ◼, ●, and ◆. Environmental samples are indicated in green type. Relevant GenBank reference strains (accession no. GU445325, GQ845367, and JX459902) are included. The scale bar indicates the genetic distance per nucleotide per site per year.
Sequence-based genotyping showed that all strains but one (NoV GII.6; P27, ward D) were most similar to the pandemic recombinant NoV GII.4 variant New Orleans 2009. The nucleotide similarity of the GII.4 sequences detected in this study ranged between 97.9 and 98.9% (1,040 nt) compared to New Orleans 2009 (GenBank accession no. GU445325) and 97.0 and 98.8% compared to NoV Berowra 2012 (GenBank accession no. JX459902). The variants clustered in a similar pattern when the trees were constructed based on either the RdRp region or the major capsid-coding region of the sequences (data not shown).
Eleven NoV GII.4 variants were detected in wards, as shown on the phylogenetic tree (Fig. 2). Ten variants were from outbreak wards, and one was from the outbreak-free ward; the variant (P61) found in the outbreak-free ward was from an isolated case of GII.4 acute gastroenteritis occurring late in the season. These 11 variants showed 0.7 to 3.7% nt variation across the RdRp-N/S capsid-coding region (1,040 nt). The nucleotide diversity (1.7 to 1.9%) over the longer 2.5-kb sequence was comparable to the diversity across the 1,040-nt region (1.2 to 1.4%) for the two variants found in the patients from ward A1 outbreaks (in January and May). As expected, a slightly higher nt diversity (2.2 to 2.6%) was noted across the hypervariable P2 subdomain of these variants.
When each of the 11 variants was compared to its nearest-neighboring variant cluster, one to three putative amino acid substitutions were detected, except for variant P13 with five silent point mutations across the RdRp-N/S major capsid-coding region (1,040 nt). The two patient variants found in A1 outbreaks (in January and May) showed two amino acid changes at residues 1521 and 1642 in the partial RdRp region. Analyses of the longer A1 sequences spanning the RdRp-N/S major capsid- and partial minor capsid-coding region (nt 4313 to 6820; GenBank accession no. GU445325) revealed seven additional amino acid changes, one in the shell (amino acid [aa] 174), five in the P2 subdomain (aa 282, 291, 294, 297, and 372), and one in the partial minor capsid protein at position 23.
Compared to the New Orleans 2009 strain (GenBank accession no. GU445325), the ward A1 strains from the January and May outbreaks showed 6 and 3 amino acid changes, respectively. Substitutions were found in the A1 January variant at residues I1521V and A1642S in RdRp, N282S, R297H, and D372N in the P2 subdomain of the major capsid protein, and N23S in the minor capsid protein. The A1 May strains showed changes in the major capsid protein at residues S174P, T291I, and P294S. Substitutions at amino acid sites 294, 297, and 372 were located over the putative blocking epitope A (30).
All 10 GII.4 variants from the outbreak wards showed high correlations with time and place of outbreak: ward A1 in January 2012, ward A2 in February, ward B1 in January, ward B1 in February, ward B1 in March, ward B1 in April, ward B2 in April, and ward B3 in April (Fig. 2). However, the outbreak strains detected in wards A1 and A2 during May belonged to the same variant, possibly due to the shared dining and shower facilities (Fig. 2 and Table 1). The GII.4 strains detected during February and March in wards C1 and C2 on floors 4 and 6 of building C also clustered as one variant.
DISCUSSION
Nosocomial outbreaks of NoV GII.4 present a recurring problem of far-reaching consequence for patients and medical staff. Concerned that an explosive spread of virus in semiclosed communities could not be accounted for by fecal-oral spread alone, Caul (31) suggested airborne transmission of virus from aerosolized vomitus as a potential source of NoV infection, although a classical respiratory spread of infections was not documented. Molecular methods are now used to monitor NoV contamination on touch surfaces (14, 32), but the question of airborne contamination remains to be elucidated. We undertook this study to further our understanding of NoV contamination and airborne dispersal of NoV in patient rooms during nosocomial outbreaks.
Aware that restrictive measures do not always contain the spread of nosocomial infections, we investigated environmental contamination in the patient rooms by surface swabbing, including dust from the patient locker tops, and a VT ionizing device designed to capture airborne virus particles. Substantial NoV GII contamination was detected in rRT-PCR studies of dust (median CT, 30.6), and VT (median CT, 32.1) in outbreak wards, as measured by the low CT values registered in environmental samples: in patient samples, CT values of 30 cycles have been estimated to approximate a viral load of 106 genome copies per gram of feces (9, 27). Environmental contamination was markedly higher in the outbreak wards (47% [total n = 101]) than in the outbreak-free ward (7% [total n = 28]). The achievement of identifying long contiguous sequences from dust, VT, and surface samples argues for the detection of intact and potentially infectious virus and against the detection of fragmented viral RNA. In the absence of cell culture, molecular analysis of genomic NoV RNA is the method currently available for investigating environmental contamination (4). Therefore, volunteer studies or more refined molecular tools, such as the rRT-PCR RNase protection assay used to assess virus capsid integrity (33), may be required to determine viral infectivity and validate the relevance of our findings in environmental samples.
Genomic diversity and the prevalence of NoV GII.4 add to the complexity of nosocomial infections (34), as seen in this study, in which 11 NoV GII.4 variants (differing by 0.7 to 3.7%, 1,040 nt) were detected in eight wards in four separate buildings during the 5-month study (Fig. 1). The phylogenetic tree showed that these 11 variants correlated with the time of onset of a given outbreak in a given ward, as in wards A1 and B1, in which a different GII.4 variant was implicated in each of the outbreaks over time (Fig. 2). NoV GII.4 variants from patients clustered on 11 subbranches of the phylogenetic tree, and environmental strains placed on five of these branches (Fig. 2). Comparative sequence analyses revealed identical or high similarity (>99.5%, 1,040 nt) between the NoV GII.4 strain detected at a given time point in dust, VTs, overbed tables, washbasins, or air vents from a given ward room and the GII.4 strain detected in the symptomatic patient/s occupying the room at that time. These molecular epidemiological studies strengthen indices showing that dust in patient rooms might be a repository for NoV GII.4 virus during outbreaks, particularly as the transmission of infection by aerosolized vomitus has been indicated in hospital (23) and hotel dining room outbreaks (16). Therefore, we suggest that airborne dispersal of NoV, as detected in dust particles and in VT samples from patient rooms, may be a source of contamination in nosocomial outbreaks.
NoV sequence variability over the hypervariable P2 subdomain (454 nt) has been suggested as a marker for tracing outbreak strains in nosocomial settings (35, 36). However, in this study, the sequencing of a longer region (1,040 nt) that revealed variant-typical sites was as informative as the P2 subdomain for tracing NoV GII.4 patient and environmental strains (36).
Various theories have been presented for the prevalence of NoV GII.4 strains in which different emerging subtypes replace one another over time (19, 37) through the suggested greater epidemiological fitness of GII.4 over other circulating genotypes, such as the GII.6 strain observed here in one patient from the outbreak-free ward. Recombination and mutation feature strongly in the evolving pandemic NoV GII.4 lineage (38, 39), and GII.4 strains from this study showed a high degree of similarity (≥97.0%) to the recombinant strains New Orleans 2009 and Berowra 2012. NoV GII.4 variation was evident in the analyses of the 11 ward-related variants reported here. When each of the 11 variants was compared to its nearest neighboring cluster, one to three amino acid substitutions were detected, except for the B1 February variant (P13), which had the least distance difference of five silent point mutations from its neighboring cluster, B3 April (Fig. 2). Point mutations found in the longer sequences from the A1 January and A1 May variants were scattered across the entire sequence studied (2.5 kb), resulting in amino acid substitutions in the RdRp, shell, P2 subdomain, and partial minor capsid protein. Amino acid substitutions in the hypervariable P2 subdomain of the A1 variants were located close to and across the putative epitope A, a blockade epitope recognized as interacting with potential neutralizing antibodies, and one of several P domain epitopes with potential antigenic and histo-blood group antigen binding functions reported as changing between epidemic GII.4 strains (37, 40–42).
Our molecular findings raise important questions about the effectiveness of current hospital ward cleaning procedures in which contract cleaning has replaced the dedicated ward cleaner (43). A comparative study of environmental contamination with gastroenteric viruses in pediatric wards during outbreak-free periods indicated that environmental surveillance improved staff awareness of the problems, and changes in cleaning protocols reduced environmental contamination, although breakthrough incidents still occurred (32). Unfortunately, major cleaning including “all surfaces above head” is often performed just once a year. Such changed emphasis in cleaning policies may contribute to the problem of recurring nosocomial infections, a problem not unique to hospitals. In a 3-year study of cruise ship outbreaks, the importance of thoroughness of disinfection cleaning (TDC) in reducing outbreaks is emphasized, with low TDC scores being predictive of subsequent NoV outbreaks (44). In the present study, point cleaning (24) of patient rooms in the outbreak-free ward following the discharge of the single case of GII.6 infection did appear to reduce the level of environmental dispersal of NoV, as measured by the negative findings on VT and surface sampling (see Table S1 in the supplemental material). However, avoiding the rapid spread of NoV GII.4 strains that characterized NoV transmission in the other wards may require a more intense cleaning response. We suggest that the high sequence similarity between NoV GII.4 strains detected in symptomatic patients and in dust and other environmental samples from patient rooms indicates the need for interventional studies of hospital ward cleaning.
In this study of seasonal outbreaks of nosocomial infections, dust and virus trap sampling provided molecular evidence supporting airborne dispersal of patient-related NoV GII.4 in hospital rooms during outbreaks, representing a novel approach for investigating noncultivable human NoV. Although questions of virus infectivity and potential spread to patients from sources such as dust remain to be determined, we suggest that interventional studies of cleaning policies are justified.
Supplementary Material
ACKNOWLEDGMENTS
This study was supported by grants from Swedish Council for Working Life and Social Research (FAS 82010-0895).
We thank Birgitta Bidefors and Katarina Lindström Johansson, Virus Detection Unit, Department of Virology, Sahlgrenska University Hospital, Gothenburg, for skilled assistance.
Footnotes
Published ahead of print 23 April 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JCM.00266-14.
REFERENCES
- 1. Roddie C, Paul JP, Benjamin R, Gallimore CI, Xerry J, Gray JJ, Peggs KS, Morris EC, Thomson KJ, Ward KN. 2009. Allogeneic hematopoietic stem cell transplantation and norovirus gastroenteritis: a previously unrecognized cause of morbidity. Clin. Infect. Dis. 49:1061–1068. 10.1086/605557 [DOI] [PubMed] [Google Scholar]
- 2. Mattner F, Sohr D, Heim A, Gastmeier P, Vennema H, Koopmans M. 2006. Risk groups for clinical complications of norovirus infections: an outbreak investigation. Clin. Microbiol. Infect. 12:69–74. 10.1111/j.1469-0691.2005.01299.x [DOI] [PubMed] [Google Scholar]
- 3. Turcios-Ruiz RM, Axelrod P, St John K, Bullitt E, Donahue J, Robinson N, Friss HE. 2008. Outbreak of necrotizing enterocolitis caused by norovirus in a neonatal intensive care unit. J. Pediatr. 153:339–344. 10.1016/j.jpeds.2008.04.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Duizer E, Schwab KJ, Neill FH, Atmar RL, Koopmans MP, Estes MK. 2004. Laboratory efforts to cultivate noroviruses. J. Gen. Virol. 85:79–87. 10.1099/vir.0.19478-0 [DOI] [PubMed] [Google Scholar]
- 5. Duizer E, Bijkerk P, Rockx B, De Groot A, Twisk F, Koopmans M. 2004. Inactivation of caliciviruses. Appl. Environ. Microbiol. 70:4538–4543. 10.1128/AEM.70.8.4538-4543.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Nenonen NP, Hannoun C, Olsson MB, Bergstrom T. 2009. Molecular analysis of an oyster-related norovirus outbreak. J. Clin. Virol. 45:105–108. 10.1016/j.jcv.2009.04.011 [DOI] [PubMed] [Google Scholar]
- 7. Kageyama T, Kojima S, Shinohara M, Uchida K, Fukushi S, Hoshino FB, Takeda N, Katayama K. 2003. Broadly reactive and highly sensitive assay for Norwalk-like viruses based on real-time quantitative reverse transcription-PCR. J. Clin. Microbiol. 41:1548–1557. 10.1128/JCM.41.4.1548-1557.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Pether JV, Caul EO. 1983. An outbreak of food-borne gastroenteritis in two hospitals associated with a Norwalk-like virus. J. Hyg. (Lond.) 91:343–350. 10.1017/S0022172400060368 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Nenonen NP, Hannoun C, Larsson CU, Bergström T. 2012. Marked genomic diversity of norovirus genogroup I strains in a waterborne outbreak. Appl. Environ. Microbiol. 78:1846–1852. 10.1128/AEM.07350-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. de Wit MA, Widdowson MA, Vennema H, de Bruin E, Fernandes T, Koopmans M. 2007. Large outbreak of norovirus: the baker who should have known better. J. Infect. 55:188–193. 10.1016/j.jinf.2007.04.005 [DOI] [PubMed] [Google Scholar]
- 11. Caul EO. 1994. Small round structured viruses: airborne transmission and hospital control. Lancet 343:1240–1242. 10.1016/S0140-6736(94)92146-6 [DOI] [PubMed] [Google Scholar]
- 12. Evans MR, Meldrum R, Lane W, Gardner D, Ribeiro CD, Gallimore CI, Westmoreland D. 2002. An outbreak of viral gastroenteritis following environmental contamination at a concert hall. Epidemiol. Infect. 129:355–360. 10.1017/S0950268802007446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Green J, Wright PA, Gallimore CI, Mitchell O, Morgan-Capner P, Brown DW. 1998. The role of environmental contamination with small round structured viruses in a hospital outbreak investigated by reverse-transcriptase polymerase chain reaction assay. J. Hosp. Infect. 39:39–45. 10.1016/S0195-6701(98)90241-9 [DOI] [PubMed] [Google Scholar]
- 14. Boxman IL, Dijkman R, te Loeke NA, Hagele G, Tilburg JJ, Vennema H, Koopmans M. 2009. Environmental swabs as a tool in norovirus outbreak investigation, including outbreaks on cruise ships. J. Food. Prot. 72:111–119 [DOI] [PubMed] [Google Scholar]
- 15. Verhoef L, Boxman IL, Duizer E, Rutjes SA, Vennema H, Friesema IH, de Roda Husman AM, Koopmans M. 2008. Multiple exposures during a norovirus outbreak on a river-cruise sailing through Europe, 2006. Euro Surveill. 13:pii=18899 http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=18899 [PubMed] [Google Scholar]
- 16. Marks PJ, Vipond IB, Carlisle D, Deakin D, Fey RE, Caul EO. 2000. Evidence for airborne transmission of Norwalk-like virus (NLV) in a hotel restaurant. Epidemiol. Infect. 124:481–487. 10.1017/S0950268899003805 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Jones EL, Kramer A, Gaither M, Gerba CP. 2007. Role of fomite contamination during an outbreak of norovirus on houseboats. Int. J. Environ. Health Res. 17:123–131. 10.1080/09603120701219394 [DOI] [PubMed] [Google Scholar]
- 18. Teunis PF, Moe CL, Liu P, Miller SE, Lindesmith L, Baric RS, Le Pendu J, Calderon RL. 2008. Norwalk virus: how infectious is it? J. Med. Virol. 80:1468–1476. 10.1002/jmv.21237 [DOI] [PubMed] [Google Scholar]
- 19. Lindesmith LC, Donaldson EF, LoBue AD, Cannon JL, Zheng DP, Vinje J, Baric RS. 2008. Mechanisms of GII.4 norovirus persistence in human populations. PLoS Med. 5:e31. 10.1371/journal.pmed.0050031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Sukhrie FH, Beersma MF, Wong A, van der Veer B, Vennema H, Bogerman J, Koopmans M. 2011. Using molecular epidemiology to trace transmission of nosocomial norovirus infection. J. Clin. Microbiol. 49:602–606. 10.1128/JCM.01443-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Greig JD, Lee MB. 2012. A review of nosocomial norovirus outbreaks: infection control interventions found effective. Epidemiol. Infect. 140:1151–1160. 10.1017/S0950268811002731 [DOI] [PubMed] [Google Scholar]
- 22. Kirking HL, Cortes J, Burrer S, Hall AJ, Cohen NJ, Lipman H, Kim C, Daly ER, Fishbein DB. 2010. Likely transmission of norovirus on an airplane, October 2008. Clin. Infect. Dis. 50:1216–1221. 10.1086/651597 [DOI] [PubMed] [Google Scholar]
- 23. Chadwick PR, McCann R. 1994. Transmission of a small round structured virus by vomiting during a hospital outbreak of gastroenteritis. J. Hosp. Infect. 26:251–259. 10.1016/0195-6701(94)90015-9 [DOI] [PubMed] [Google Scholar]
- 24.Vårdhygien. 2011. Handlingsplan vid misstänkt/verifierad virusgastroenterit på akutintag och vårdenhet på Sahlgrenska Universitetssjukhuset, Frölunda Specialistsjukhus, Kungälvs sjukhus och Capio Lundby sjukhus. Sahlgrenska Universitetssjukhuset, Gothenburg, Sweden: (In Swedish.) http://www.sahlgrenska.se/upload/SU/omrade_4/laboratoriemedicin/klinisk_mikrobiologi/vardhygien/gastroenterit.pdf [Google Scholar]
- 25. Kwok S, Higuchi R. 1989. Avoiding false positives with PCR. Nature 339:237–238. 10.1038/339237a0 [DOI] [PubMed] [Google Scholar]
- 26. Nenonen NP, Hannoun C, Horal P, Hernroth B, Bergström T. 2008. Tracing of norovirus outbreak strains in mussels collected near sewage effluents. Appl. Environ. Microbiol. 74:2544–2549. 10.1128/AEM.02477-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Gustavsson L, Westin J, Andersson LM, Lindh M. 2011. Rectal swabs can be used for diagnosis of viral gastroenteritis with a multiple real-time PCR assay. J. Clin. Virol. 51:279–282. 10.1016/j.jcv.2011.05.025 [DOI] [PubMed] [Google Scholar]
- 28. Oka T, Katayama K, Hansman GS, Kageyama T, Ogawa S, Wu FT, White PA, Takeda N. 2006. Detection of human sapovirus by real-time reverse transcription-polymerase chain reaction. J. Med. Virol. 78:1347–1353. 10.1002/jmv.20699 [DOI] [PubMed] [Google Scholar]
- 29. Heim A, Ebnet C, Harste G, Pring-Akerblom P. 2003. Rapid and quantitative detection of human adenovirus DNA by real-time PCR. J. Med. Virol. 70:228–239. 10.1002/jmv.10382 [DOI] [PubMed] [Google Scholar]
- 30. Lindesmith LC, Costantini V, Swanstrom J, Debbink K, Donaldson EF, Vinjé J, Baric RS. 2013. Emergence of a norovirus GII.4 strain correlates with changes in evolving blockade epitopes. J. Virol. 87:2803–2813. 10.1128/JVI.03106-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Caul EO. 1996. Viral gastroenteritis: small round structured viruses, caliciviruses and astroviruses. Part I. The clinical and diagnostic perspective. J. Clin. Pathol. 49:874–880 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Gallimore CI, Taylor C, Gennery AR, Cant AJ, Galloway A, Xerry J, Adigwe J, Gray JJ. 2008. Contamination of the hospital environment with gastroenteric viruses: comparison of two pediatric wards over a winter season. J. Clin. Microbiol. 46:3112–3115. 10.1128/JCM.00400-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Topping JR, Schnerr H, Haines J, Scott M, Carter MJ, Willcocks MM, Bellamy K, Brown DW, Gray JJ, Gallimore CI, Knight AI. 2009. Temperature inactivation of feline calicivirus vaccine strain FCV F-9 in comparison with human noroviruses using an RNA exposure assay and reverse transcribed quantitative real-time polymerase chain reaction-A novel method for predicting virus infectivity. J. Virol. Methods 156:89–95. 10.1016/j.jviromet.2008.10.024 [DOI] [PubMed] [Google Scholar]
- 34. Morter S, Bennet G, Fish J, Richards J, Allen DJ, Nawaz S, Iturriza-Gómara M, Brolly S, Gray J. 2011. Norovirus in the hospital setting: virus introduction and spread within the hospital environment. J. Hosp. Infect. 77:106–112. 10.1016/j.jhin.2010.09.035 [DOI] [PubMed] [Google Scholar]
- 35. Xerry J, Gallimore CI, Cubitt D, Gray JJ. 2010. Tracking environmental norovirus contamination in a pediatric primary immunodeficiency unit. J. Clin. Microbiol. 48:2552–2556. 10.1128/JCM.00066-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Sukhrie FH, Teunis P, Vennema H, Bogerman J, van Marm S, Beersma MF, Koopmans M. 2013. P2 domain profiles and shedding dynamics in prospectively monitored norovirus outbreaks. J. Clin. Virol. 56:286–292. 10.1016/j.jcv.2012.12.006 [DOI] [PubMed] [Google Scholar]
- 37. Debbink K, Lindesmith LC, Donaldson EF, Costantini V, Beltramello M, Corti D, Swanstrom J, Lanzavecchia A, Vinjé J, Baric RS. 2013. Emergence of new pandemic GII.4 Sydney norovirus strain correlates with escape from herd immunity. J. Infect. Dis. 208:1877–1887. 10.1093/infdis/jit370 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Bull RA, Tanaka MM, White PA. 2007. Norovirus recombination. J. Gen. Virol. 88:3347–3359. 10.1099/vir.0.83321-0 [DOI] [PubMed] [Google Scholar]
- 39. Eden JS, Tanaka MM, Boni MF, Rawlinson WD, White PA. 2013. Recombination within the pandemic norovirus GII.4 lineage. J. Virol. 87:6270–6282. 10.1128/JVI.03464-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Bok K, Abente EJ, Realpe-Quintero M, Mitra T, Sosnovtsev SV, Kapikian AZ, Green KY. 2009. Evolutionary dynamics of GII.4 noroviruses over a 34-year period. J. Virol. 83:11890–11901. 10.1128/JVI.00864-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Allen DJ, Gray JJ, Gallimore CI, Xerry J, Iturriza-Gómara M. 2008. Analysis of amino acid variation in the P2 domain of the GII-4 norovirus VP1 protein reveals putative variant-specific epitopes. PLoS One 3:e1485. 10.1371/journal.pone.0001485 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Lindesmith LC, Beltramello M, Donaldson EF, Corti D, Swanstrom J, Debbink K, Lanzavecchia A, Baric RS. 2012. Immunogenetic mechanisms driving norovirus GII.4 antigenic variation. PLoS Pathog. 8:e1002705. 10.1371/journal.ppat.1002705 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Dancer SJ, White LF, Lamb J, Girvan EK, Robertson C. 2009. Measuring the effect of enhanced cleaning in a UK hospital: a prospective cross-over study. BMC Med. 7:28. 10.1186/1741-7015-7-28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Carling PC, Bruno-Murtha LA, Griffiths JK. 2009. Cruise ship environmental hygiene and the risk of norovirus infection outbreaks: an objective assessment of 56 vessels over 3 years. Clin. Infect. Dis. 49:1312–1317. 10.1086/606058 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.