

Abstract
This study aims to understand the extent to which modulation of the Na+-K+-2Cl− cotransporter NKCC2 differential splicing affects NaCl delivery to the macula densa. NaCl absorption by the thick ascending limb and macula densa cells is mediated by apical NKCC2. A recent study has indicated that differential splicing of NKCC2 is modulated by dietary salt (Schieβl IM, Rosenauer A, Kattler V, Minuth WW, Oppermann M, Castrop H. Am J Physiol Renal Physiol 305: F1139–F1148, 2013). Given the markedly different ion affinities of its splice variants, modulation of NKCC2 differential splicing is believed to impact NaCl reabsorption. To assess the validity of that hypothesis, we have developed a mathematical model of macula densa cell transport and incorporated that cell model into a previously applied model of the thick ascending limb (Weinstein AM, Krahn TA. Am J Physiol Renal Physiol 298: F525–F542, 2010). The macula densa model predicts a 27.4- and 13.1-mV depolarization of the basolateral membrane [as a surrogate for activation of tubuloglomerular feedback (TGF)] when luminal NaCl concentration is increased from 25 to 145 mM or luminal K+ concentration is increased from 1.5 to 3.5 mM, respectively, consistent with experimental measurements. Simulations indicate that with luminal solute concentrations consistent with in vivo conditions near the macula densa, NKCC2 operates near its equilibrium state. Results also suggest that modulation of NKCC2 differential splicing by low salt, which induces a shift from NKCC2-A to NKCC2-B primarily in the cortical thick ascending limb and macula densa cells, significantly enhances salt reabsorption in the thick limb and reduces Na+ and Cl− delivery to the macula densa by 3.7 and 12.5%, respectively. Simulation results also predict that the NKCC2 isoform shift hyperpolarizes the macula densa basolateral cell membrane, which, taken in isolation, may inhibit the release of the TGF signal. However, excessive early distal salt delivery and renal salt loss during a low-salt diet may be prevented by an asymmetric TGF response, which may be more sensitive to flow increases.
Keywords: differential splicing, epithelial transport, low-salt diet, mathematical modeling, tubuloglomerular feedback, salt homeostasis, blood pressure
the kidney plays an essential role in blood pressure regulation by means of short-term and long-term control of water and salt balance. About 25% of the filtered NaCl is reabsorbed by the thick ascending limb (TAL) of the loop of Henle. Because TAL cells are largely water impermeable, the reabsorption of NaCl is not accompanied by water reabsorption, resulting in a decrease in luminal NaCl concentration ([NaCl]) from ∼250 mM at the loop bend of a superficial nephron to as low as 20 mM (38, 44), together with a substantial reduction in tubular fluid osmolality, at the end of the cortical TAL. There, luminal [NaCl] is detected by the macula densa cells, which form a cluster of specialized sensor cells that detect changes in distal tubular fluid composition and convey signals to the glomerular arterioles. An increase in luminal [NaCl] at the macula densa generates a signaling cascade that ultimately induces vasoconstriction of the afferent arteriole and reduces glomerular filtration rate. This feedback mechanism, called tubuloglomerular feedback (TGF), balances TAL NaCl load with transport capacity and stabilizes NaCl delivery to the distal nephron downstream, where fine regulation of NaCl homeostasis takes place (42).
Sodium reabsorption along the TAL was initially thought to be passively driven by the favorable transepithelial electric potential gradient (4). Several scenarios were later proposed to explain the coupling between the active transport of Cl− and that of Na+ (5). The first descriptions of Na+-K+-Cl− cotransport activity in the TAL were published in the early 1980s (15, 17). We now understand that NaCl reabsorption in this segment is mediated on the apical side by the Na+-K+-2Cl− cotransporter NKCC2, which carries Na+, K+, and Cl− from the lumen into the cell. Na+ is subsequently extruded across the basolateral membrane by Na+-K+-ATPase pumps, whereas Cl− exits via basolateral K+-Cl− (KCC) cotransporters or Cl− channels (14, 21, 22). K+ is partly recycled by apical renal outer medullary K+ (ROMK) channels. Similarly, NKCC2 provides the primary mean for NaCl entry into the macula densa cells (25).
Three splice variants of NKCC2 are known to exist: A, B, and F. These isoforms differ markedly in their ion affinities. Variations among studies and species notwithstanding, NKCC2-B has a higher affinity for Cl− than NKCC2-A, whose affinity in turn exceeds that of NKCC2-F (10, 11, 32). The distribution of these isoforms varies along the different segments of the TAL, which may optimize the efficiency of NaCl uptake, given the large decrease in luminal [NaCl] along the TAL. In the rat kidney, isoform-specific RT-PCR indicates that NKCC2-F is localized in the medullary TAL, NKCC2-A in the medullary and cortical TAL as well as in macula densa cells, and NKCC2-B in the cortical TAL and macula densa cells (51).
In a recent study, Schieβl et al. (36) observed that the differential splicing of NKCC2 is modulated by dietary salt intake. A low-salt diet elicits a shift from NKCC2-A towards NKCC2-B, primarily in the cortical TAL and macula densa cells. The goal of the present study was to understand the extent to which modulation of NKCC2 differential splicing affects NaCl uptake by the TAL. To accomplish that goal, we developed a mathematical model of epithelial transport in the TAL, including the macula densa, and used that model to examine the regulation of NaCl transport in those segments. Specifically, we aimed to assess the validity of the hypothesis that the differential splicing of NKCC2 contributes to the capacity of the kidney to adapt to changes in reabsorptive needs.
MATHEMATICAL MODEL
We extended an epithelial cell-based model of the TAL of the loop of Henle of a superficial nephron in a rat kidney to include the macula densa plaque. The model TAL consists of a 2.5-mm medullary TAL (mTAL) segment and a 3.5-mm cortical TAL (cTAL) segment. The cTAL is assumed to be contiguous with a cluster of macula densa cells.
TAL model.
The transport of water and solutes across the mTAL and cTAL is determined following the approach of Weinstein (46) and Weinstein and Krahn (48). The model accounts for 13 solutes: Na+, K+, Cl−, HCO3−, H2CO3, CO2, NH3, NH4+, HPO42−, H2PO4−, H+, Ca2+, and urea. The (fixed) peritubular concentrations are homogeneous in the cortex and vary along the cortico-medullary axis in the outer medulla; see Table 1. In each simulation, we also specify the composition of the luminal fluid at the mTAL inlet. At a given axial position along the mTAL and cTAL, the flow rate (or cysotolic volume), electric potential, and solute concentrations in the lumen and cells are obtained by solving steady-state conservation equations for volume, charge, and mass, respectively. The Weinstein model is modified in two respects: the total length of the TAL is increased to 6 mm; we also account for the transport of Ca2+, which is reabsorbed only via the paracellular pathway in the TAL. The permeability of the mTAL and cTAL to Ca2+ is taken as 4.2 and 8.4 × 10−5 cm/s, respectively. All other TAL parameter values can be found in Table 1 in Ref. 48. They are identical in mTAL and cTAL, except for the density of NKCC2 (see below).
Table 1.
mTAL inflow and peritubular concentrations
| mTAL Inflow (Lumen) | OM-IM (Peritubular) | C-M (Peritubular) | MD (Peritubular) | |
|---|---|---|---|---|
| Na+ | 250 | 284 | 144 | 144 |
| K+ | 10 | 10 | 5 | 5 |
| Cl− | 232.54 | 270.94 | 123.88 | 122.88 |
| HCO3− | 25 | 25 | 25 | 25 |
| H2CO3 | 4.41 × 10−3 | 4.41 × 10−3 | 4.41 × 10−3 | 4.41 × 10−3 |
| CO2 | 1.5 | 1.5 | 1.5 | 1.5 |
| HPO4− | 6 | 3 | 2 | 2 |
| H2PO4− | 1.8 | 0.9 | 0.6 | 0.6 |
| Urea | 10 | 20 | 5 | 5 |
| NH3 | 57.3 × 10−3 | 57.3 × 10−3 | 29.4 × 10−3 | 14.7 × 10−3 |
| NH4+ | 3.84 | 3.84 | 1.97 | 0.99 |
| Ca2+ | 3.75 | 2.50 | 1.25 | 1.25 |
Concentrations are in mM. mTAL, medullary thick ascending limb; OM-IM, outer-inner medullary junction; C-M, cortico-medullary junction; MD, macula densa. Peritubular concentrations are taken to vary linearly between the OM-IM and C-M junctions; peritubular [NH4+] and [Cl−] also vary linearly between the C-M junction and the MD.
Macula densa model.
The cluster of macula densa cells is represented by one model macula densa cell (Fig. 1), which differs from the model cTAL cell in the following respects: it does not include a basolateral KCC4 cotransporter, since no studies have reported its presence in the macula densa; the basolateral K+ conductance is reduced by a factor of 10; and the abundance of apical K+ (ROMK) channels is increased by a factor of 2 relative to the cTAL (19).
Fig. 1.

Schematic diagram of the model macula densa (MD) cell. Not shown are basolateral Na+-3HCO3− and 2Na+-HPO42− cotransporters and solute background permeabilities. Luminal and peritubular concentrations are chosen to reflect normal-flow conditions. The model computes intracellular concentrations and transcellular fluxes (shown in parentheses).
In view of the measured effects of ouabain in the macula densa of rabbit nephrons, Bell et al. (3) suggested that macula densa cells regulate their cytosolic Na+ concentration by means of an apical, nongastric H+-K+-ATPase that can also act as a Na+/K+ exchanger, rather than with basolateral Na+-K+-ATPase pumps. In rat macula densa cells, however, significant levels of Na+-K+-ATPase have been detected (31, 49). Moreover, studies in mice have shown that TGF responses are conditional upon the activity of the Na+/K+-ATPase, not that of the Na+/H+ exchanger (NHE) or the nongastric H+-K+-ATPase (27). In the present rat model, we assume that Na+-K+-ATPase expression is 20% lower in macula densa cells than in cTAL cells, so as to account for lower detection levels in the former (49). The Na+ affinity of the pump is also taken to differ, since the rat macula densa, in contrast to the cTAL, expresses significant levels of the γ-subunit of the Na+-K+-ATPase (49), which is characterized by higher antagonistic effects of intracellular K+ on the binding of Na+ to cytoplasmic sites. The affinity of the Na+-K+-ATPase to Na+ in macula densa cells is taken as (33)
| 1 |
where [K+]i denotes the intracellular (cytosolic) concentration of K+.
In addition, macula densa cells are significantly permeable to water. The osmotic permeability of the apical and basolateral cell membrane is taken as 130 and 1,700 μm/s, respectively, so the net transepithelial permeability equals 125 μm/s (12). In comparison, the osmotic permeability of the collecting duct in the presence of vasopressin is ∼500 μm/s (34).
Parameter values for macula densa cells are summarized in Table 2. The distribution of NKCC2 isoforms A, B, and F is given in Table 3. It should be noted that apical Na+/H+ exchange is mediated by NHE2 in macula densa cells and by NHE3 in TAL cells (31). The apparent affinity of NHE2 to Na+ is ∼10 times higher than that of NHE3 (52). Nevertheless, in the absence of a full kinetic model of NHE2, we use that of NHE3 instead.
Table 2.
Transport parameters of macula densa cells
| Transporter | Value | Unit | Reference |
|---|---|---|---|
| Na+/H+ exchanger density (apical) | 6 | 10−9 mmol/cm2 | (48) |
| Na+-K+-ATPase density | 1,040 | 10−9 mmol/s/cm2 | Estimated |
| K+ apical permeability | 40 | 10−5 cm/s | (19) |
| K+ basolateral permeability | 0.2 | 10−5 cm/s | Estimated |
| Cl− basolateral permeability | 5.0 | 10−5 cm/s | Estimated |
| HCO3− basolateral permeability | 0.5 | 10−5 cm/s | Estimated |
| Pf, apical membrane | 133 | 10−4 cm/s | (12) |
| Pf, tight junction | 700 | 10−4 cm/s | (48) |
| Pf, basolateral membrane | 1,687 | 10−4 cm/s | (12) |
Pf, osmotic permeability. Other transport parameters are identical to those of the TAL and are listed in Table 1 in Ref 48.
Table 3.
NKCC2 expression levels
| Segment | NKCC2-A | NKCC2-B | NKCC2-F |
|---|---|---|---|
| Normal-salt conditions | |||
| mTAL | 8 | 0 | 50 |
| cTAL | 10 | 12 | 0 |
| MD | 20 | 20 | 0 |
| Low-salt conditions | |||
| mTAL | 5 | 0 | 70 |
| cTAL | 5 | 42 | 0 |
| MD | 10 | 70 | 0 |
Levels are given in pmol/cm2. NKCC2, Na+-K+-2Cl− cotransporter; cTAL, cortical thick ascending limb.
To validate the macula densa model, we compared its predictions with experimental results. Under basal ex vivo conditions, the peritubular fluid was assumed to contain the following (in mM): 145 NaCl, 5 NaHCO3, 5 KCl, 1.5 CaCl2, 1.6 Na2HPO4, 0.4 NaH2PO4, and 5 glucose. The composition of the luminal fluid differed in that luminal [NaCl] was lowered to 25 mM and isoosmotically replaced with an impermeant.
RESULTS
Macula densa model.
We first validated the macula densa model by comparing its predictions to experimental results. Note that most published measurements were obtained in rabbits, whereas our model applies to the rat. Under ex vivo conditions, with 25 mM [NaCl] in the lumen and 145 mM [NaCl] in the bath, the predicted basolateral transmembrane potential difference (Vbl) is −79 mV and the predicted intracellular concentrations of Na+, K+, and Cl− are, respectively, 22, 152, and 8 mM (Table 4). Whereas the luminal and cytosolic concentrations of Na+ and Cl− have the same order of magnitude, the lumen-to-cell K+ concentration ratio is much lower than 1; thus the NKCC2 transporter is predicted to mediate the extrusion of Na+, K+, and Cl− into the lumen (see below for simulations with modified luminal fluid composition that better mimics in vivo conditions).
Table 4.
Model predictions at macula densa under ex vivo conditions
| Conditions | [Na+]i | [K+]i | [Cl−]i | Vbl | pHi | JNKCC2 | JNHE |
|---|---|---|---|---|---|---|---|
| Lumen [NaCl] = 25 mM (reference) | 21.7 | 152.3 | 7.6 | −79.0 | 6.87 | −20.6 | 54.6 |
| Lumen [NaCl] = 145 mM | 61.2 | 112.9 | 27.7 | −51.6 | 6.89 | 121.7 | 48.0 |
| Bath [NaCl] = 25 mM | 27.2 | 146.8 | 6.5 | −54.2 | 6.84 | 52.8 | 52.1 |
| Lumen and bath [NH4Cl] = 0.1 mM | 22.4 | 151.1 | 7.5 | −79.2 | 6.85 | −20.6 | 57.6 |
| Lumen and bath [NH4Cl] = 1.0 mM | 31.4 | 135.9 | 7.0 | −81.2 | 6.70 | −21.2 | 93.9 |
| NKCC2 × 2.0 | 21.6 | 152.3 | 7.5 | −79.2 | 6.87 | −21.1 | 54.9 |
| NKCC2 × 0.5 | 21.8 | 152.2 | 7.7 | −78.7 | 6.87 | −19.6 | 54.3 |
| NHE × 2.0 | 23.8 | 151.3 | 7.3 | −79.3 | 6.96 | −27.7 | 72.2 |
| NHE × 0.5 | 19.9 | 152.9 | 7.9 | −78.7 | 6.78 | −14.4 | 39.3 |
Reference luminal and peritubular concentrations under ex vivo conditions are given in mathematical model. JNKCC2 and JNHE, respectively, represent the Na+ fluxes across NKCC2 and apical Na+/H+ exchanger (NHE). Fluxes are given in pmol·min−1·mm−1, concentrations are in mM, and the basolateral membrane potential (Vbl) is in mV. The last 4 rows correspond to a twofold increase or decrease in the density of NKCC2 or NHE, respectively.
The effects of increasing luminal [NaCl] under isoosmotic conditions are shown in Fig. 2. With 145 mM [NaCl] in the lumen and bath, the intracellular concentration of Na+ ([Na+]i) is calculated as 61 mM, ∼10 mM lower than experimental values (30), and that of Cl− ([Cl−]i) as 28 mM. Our model predicts a 27.4-mV depolarization of the basolateral membrane when luminal [NaCl] is raised from 25 to 145 mM; published measurements range between 20 and 30 mV (3, 37). Elevations in luminal [NaCl] stimulate transport across NKCC2, thereby raising the intracellular concentrations of Na+, K+, and Cl−. The [Cl−]i increase in turn stimulates the electrogenic efflux of Cl− via the basolateral Cl− conductance and makes Vbl less negative, which affects the TGF signal. The present model does not explicit represent the TGF signaling mechanism. However, as further discussed below, depolarization of the macula densa basolateral membrane has been proposed as a mechanism for initiation of the TGF signal (1). The apical NHE flux is predicted to decrease owing to the high [Na+]i. Note that the predicted [Na+]i does not reach the plateau that has been observed in rabbit macula densa cells (30).
Fig. 2.
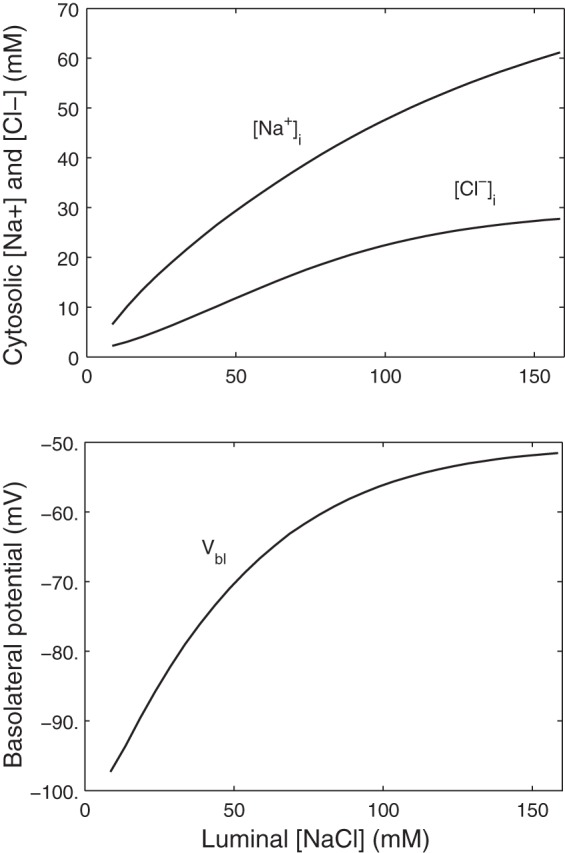
Effects of luminal NaCl concentration ([NaCl]) variations on the basolateral membrane potential (Vbl) and the intracellular concentrations of Na+ ([Na+]i) and Cl− ([Cl−]i) in macula densa cells.
We then simulated variations in peritubular [Na+], [Cl−], or [NaCl] under isoosmotic conditions. While such a maneuver is of limited physiological relevance in vivo, it has been used in isolated preparations of the juxtaglomerular apparatus. As shown in Table 4, our model predicts a 24.8-mV depolarization when peritubular [NaCl] is decreased from 145 to 25 mM, slightly less than the measured value of 29 mV (24). In accordance with experimental findings (24, 37), a 120-mM decrease in peritubular [Cl−] is predicted to depolarize Vbl, whereas a 120 mM decrease in peritubular [Na+] is predicted to hyperpolarize Vbl. Indeed, lowering peritubular [NaCl] or [Cl−] enhances the driving force for basolateral Cl− exit, thereby reducing [Cl−]i and depolarizing the basolateral membrane. Our model suggests that, in contrast, lowering peritubular [Na+] stimulates Na+ extrusion via (electrogenic) Na+-K+-ATPase pumps, thus hyperpolarizing the cell.
We also examined the effects of varying the density of NKCC2 or apical NHE transporters. As shown in Table 4, with 25 mM [NaCl] in the lumen and 145 mM [NaCl] in the peritubular fluid, an isolated, twofold increase in NKCC2 expression is predicted to slightly augment both Na+ extrusion via NKCC2 and Na+ import via NHE, to decrease [Cl−]i to a small extent, and to hyperpolarize the basolateral membrane by 0.2 mV. An isolated twofold increase in apical NHE expression has comparable effects. The increase in Na+ import via NHE is partly counterbalanced by an increase in Na+ extrusion via NKCC2. The extrusion of Cl− increases concomitantly, and Vbl is reduced by 0.3 mV.
In vivo, the luminal fluid at the macula densa is significantly hyposmotic relative to the interstitial fluid. Moreover, the luminal concentration of NH4+ in the early distal tubule is >1 mM (20, 39), whereas the previous simulations were done in the presence of low NH4+ (0.010 mM). Based on the results of our TAL model (described below), we computed macula densa transcellular fluxes and cytosolic concentrations when the luminal concentrations of Na+, K+, and Cl− are, respectively, 44, 2.5, and 34 mM. In addition, luminal pH was set to 6.60, and the luminal concentrations of HCO3−, ammonia species, phosphate species, and urea were taken as 4, 1.7, 9, and 12 mM, respectively. As shown in Fig. 1, under these conditions NKCC2 mediates a small amount of NaCl reabsorption. The Na+ flux across NKCC2 (∼20 pmol·min−1·mm−1) is about four times lower than that across the apical NHE. The model also predicts that, with so little K+ in the luminal fluid, NKCC2 extrudes K+ into the tubular fluid. In other words, a small fraction of the cotransporter secretes K+, Na+, and Cl− in the lumen, whereas the rest carry NH4+, Na+, and Cl− from lumen to cell.
Vallon et al. (43) provided experimental evidence that luminal [K+], in addition to [Cl−], constitutes the rate-limiting luminal TGF signal. To test that hypothesis, we varied luminal [K+] and computed Vbl at the macula densa. Specifically, as in the previous simulation, luminal concentrations were set based on the results of our TAL model, with the exception of [K+], which was set to 1.5, 2.5 (base case), and 3.5 mM. These luminal K+ concentrations yielded Vbl values of −90.3, −82.7, and −77.2 mV, respectively. Thus an increase in luminal [K+] depolarizes the macula densa basolateral membrane. That effect is analogous to variations in luminal [Cl−], suggesting that luminal [K+] may indeed be a positive regulator of the TGF response.
The macula densa expresses the NKCC2 isoforms A and B in a 1:1 ratio when rats are fed a standard salt diet (36). Under the conditions shown in Fig. 1, NKCC2-A is predicted to transport about a third less Na+, K+, Cl−, and NH4+ than NKCC2-B. Indeed, even though NKCC2-B has the lowest transport capacity, its affinity to Na+, Cl−, K+, and NH4+ is higher than that of NKCC2-A (11, 29, 32).
The effects of luminal ammonium on intracellular concentrations and Vbl in macula densa cells are shown in Fig. 3. In these simulations, the luminal fluid contains 35 mM Na+, 35 mM Cl−, and either 2 or 5 mM K+. Peritubular concentrations remain unchanged. At both K+ concentrations, increasing luminal [NH4+] acidifies the cell, thereby stimulating proton extrusion via the Na+/H+ exchanger, which raises [Na+]i and increases the Na+-K+-ATPase pumping rate. The more acidic pHi also reduces the driving force across the basolateral Cl−/HCO3− exchanger and decreases [Cl−]i. The Na+-K+-ATPase stimulation and [Cl−]i decrease both contribute to making the basolateral membrane potential more negative, which may inhibit the TGF signal. Note that the [Na+]i variations are less pronounced at high luminal [K+], because the contribution of NH4+ to the net NKCC2 flux is then smaller. That is, the impact of NH4+ on the transcellular transport of Na+ is reduced when there is more ambient K+.
Fig. 3.
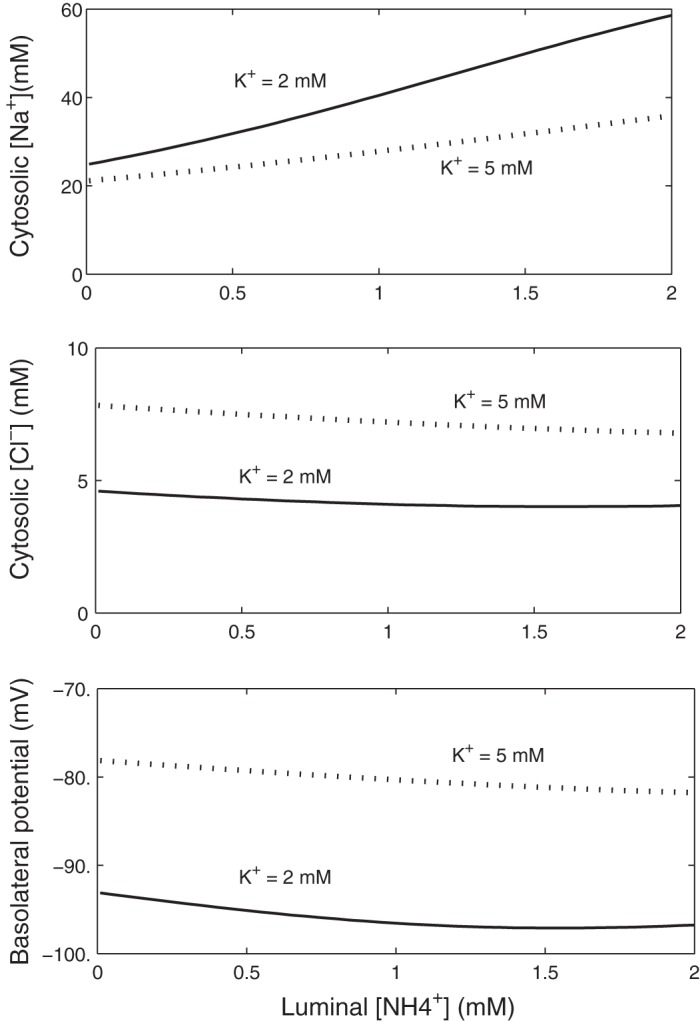
Effects of luminal [NH4+] variations on [Na+]i, [Cl−]i, and Vbl in macula densa cells. Luminal [NaCl] is set to 35 mM, and luminal [K+] equals 2 or 5 mM.
TAL model.
We then simulated the transport of water and solutes along the entire TAL. Luminal [Na+], [K+], [Cl−], and [NH4+] along the TAL are shown in Fig. 4. The changes in the slopes of the profiles, seen at the cortico-medullary junction, arise from the abrupt change in epithelial transport properties at that boundary. In particular, the distribution of NKCC2 isoforms, and the total amount of transporter, vary significantly between the medullary and cortical TAL (Table 2).
Fig. 4.

Luminal concentrations of selected solutes along the thick ascending limb (TAL).
Solute concentrations generally decrease along the flow direction, except for those of H+ and H2PO4−. NHE mediates acidification of the luminal fluid along the model TAL, consistent with experimental evidence (13). The increase in luminal [H2PO4−] is due to the buffering of secreted H+ by HPO42−.
Basolateral Na+-K+-ATPase provides the driving force for Na+ reabsorption, diluting its luminal concentration from 250 mM at the loop bend to 44.3 mM at the macula densa. The luminal concentration of the major anion Cl− decreases correspondingly from 233 to 33.5 mM. In the outer medulla, paracellular fluxes account for a substantial fraction of the NaCl reabsorptive fluxes; in the upper cortex, however, an unfavorable electrochemical gradient develops and the paracellular NaCl fluxes reverse direction. That prediction is consistent with a previous modeling study (28). It is noteworthy that luminal [Cl−] exhibits a significant decreasing slope at the macula densa, an indication that the system has not reached a static head, when transcellular Cl− reabsorption is nearly balanced by paracellular backleak. A nearly flat [Cl−] profile near the macula densa would render TGF ineffective.
The transepithelial membrane potential (Vte) increases from 2.7 mV at loop bend to 11.4 mV at the macula densa. Owing to the recycling of K+ and NH4+ across the luminal membrane, Vte is lumen positive, which partially compensates for the unfavorable transepithelial [Na+] gradient established in the cortex.
Low-salt-induced differential splicing of NKCC2 increases TAL NaCl reabsorption.
Schieβl et al. (36) reported that dietary salt intake modulates the differential splicing of NKCC2. During salt depletion (i.e., a prominent negative salt balance, and not just a low-salt diet), TAL tubular fluid flow is believed to decrease, due to lower glomerular filtration rate and higher proximal tubular reabsorption (7, 9, 45, 50). To study the effects of varying NKCC2 expression levels on TAL NaCl reabsorption, we conducted simulations in which the loop-bend fluid flow was reduced from 6.0 to 4.6 nl/min. This value was estimated by reducing the baseline flow in inverse proportion to the measured increase in fractional reabsorption by the proximal tubule in salt-depleted mice (50). Inflow solute concentrations were unchanged. The expression levels of the three NKCC2 isoforms in each segment were modified as shown in Table 3.
Note first that at low flow, NKCC2 is predicted to transport NaCl in the reverse direction at the macula densa, owing to reduced luminal concentrations (see below). That is, the majority of NKCC2 cotransporters carry Na+, Cl−, and K+ from the cytosol to the lumen, while a smaller fraction carries Na+, Cl−, and NH4+ into the cell. Since the NKCC2 flux is smaller than the NHE flux, the macula densa cell still mediates net Na+ reabsorption. The NKCC2 Cl− flux is not counterbalanced by any other apical Cl− transporter, however, and Cl− is thus secreted under these conditions.
At low flow, the model predicts substantially lower luminal [Na+] and [Cl−] at the macula densa, as illustrated in Table 5. With base-case NKCC2 isoform expression levels, [Na+] and [Cl−] are 37.8 and 24.0 mM, respectively, which corresponds to 14.7 and 28.4% decreases from values obtained with baseline flow. When “low-salt” NKCC2 isoform expression levels are assumed, macula densa luminal [Na+] and [Cl−] are predicted to be 36.4 and 21.0 mM, respectively, which correspond to an additional reduction of 3.7 and 12.5%, respectively, in delivery to the distal tubule. In other words, our model predicts that the shift from NKCC2-A to NKCC2-B enhances NaCl reabsorption along the TAL.
Table 5.
Model predictions at macula densa under normal- and low-salt conditions
| Conditions | Normal | Low Salt | Low Salt | Low Salt | Low Salt | Low Salt |
|---|---|---|---|---|---|---|
| NKCC2 Isoform | Base Case | Base Case | Switch | Base Case | Switch | Switch |
| Na+-K+-ATPase | Base Case | Base Case | Base Case | +50% | +50% | Segment Specific* |
| Lumen [Na+] | 44.3 | 37.8 | 36.4 | 35.8 | 35.1 | 34.5 |
| Lumen [Cl−] | 33.5 | 24.0 | 21.0 | 20.2 | 17.7 | 17.9 |
| Lumen [K+] | 2.66 | 3.13 | 3.50 | 3.11 | 3.42 | 3.36 |
| Lumen [NH4+] | 1.66 | 1.78 | 1.14 | 1.86 | 1.26 | 1.27 |
| Vbl | −82.7 | −86.8 | −88.3 | −88.6 | −90.7 | −90.9 |
| NHE Na+ flux | 105 | 93.1 | 105 | 92.8 | 113 | 114 |
| NKCC2 Na+ flux | 21.5 | −1.66 | −11.4 | −3.44 | −11.7 | −11.3 |
| NKCC2 K+ flux | −72.1 | −74.8 | −103 | −67.0 | −104 | −105 |
| NKCC2 NH+4 flux | 93.7 | 73.2 | 91.8 | 63.6 | 92.0 | 93.5 |
See text.
A low-salt diet not only modulates the relative abundance of NKCC2 isoforms but also increases the overall expression levels of NKCC2 (Table 3). As discussed by Weinstein (47), substantial modulation of transepithelial fluxes may require the coordinated regulation of luminal and peritubular transporters. It is therefore likely that low-salt-induced increases in NKCC2 abundance are accompanied by the upregulation of basolateral transporters, such as the Na+-K+-ATPase. When Na+-K+-ATPase expression was raised by 50% throughout the TAL, luminal [Na+] and [Cl−] at the macula densa were predicted to decrease to 35.1 and 17.7 mM, respectively. When base-case NKCC2 expression was assumed, a 50% increase in Na+-K+-ATPase expression alone yielded higher luminal concentrations, i.e., 35.8 mM [Na+] and 20.2 mM [Cl−].
It is reasonable to speculate that the expressions of NKCC2 and Na+-K+-ATPase are upregulated proportionally. Thus, in another simulation, we assumed that in each segment, the abundance of Na+-K+-ATPase increases half as much as that of NKCC2. Accordingly, Na+-K+-ATPase expression was raised by 14.7, 56.7, and 50.0% in the mTAL, cTAL, and macula densa cells, respectively. With this configuration, the luminal [Na+] and [Cl−] at the macula densa were predicted as 34.5 and 17.9 mM, respectively. Note that even with a substantially lower increase in mTAL Na+-K+-ATPase expression, these concentrations are similar to the case where Na+-K+-ATPase expression is increased uniformly by 50%. Results are summarized in Table 5.
To assess the robustness of our prediction that splicing changes affect NaCl delivery to the macula densa, we then conducted a sensitivity study. We varied the values of selected model parameters and computed the differences in macula densa luminal [Na+] and [Cl−] with baseline and low-salt induced NKCC2 isoform expressions. In separate sets of simulations, we varied by ±25% the apical NHE density, the apical permeability to K+, or Na+-K+-ATPase expression in the mTAL; else we varied by ±25 mM the loop-bend luminal [NaCl]; else we assumed a 2-mm mTAL (reduced from a base-case length of 2.5 mm) with a 3.5- or 4-mm cTAL. Our results suggest that the finding that the low-salt-induced differential splicing of NKCC2 increases TAL NaCl reabsorption is robust: together these simulations predicted that the isoform switch decreases macula densa luminal [Na+] and [Cl−] by 2.8–5.9 and 10.9–15.0%, respectively.
Effects of NKCC2 differential splicing on TGF.
We then considered the effects of NKCC2 differential splicing on TGF, with a focus on the Vbl of macula densa cells, since the TGF signaling mechanism is not explicitly represented in the model. We conducted simulations for a range of mTAL inflow rates, from 3.6 to 8.0 nl/min, for base-case and low-salt NKCC2 isoform expressions at the macula densa. With NKCC2 isoform distribution along the TAL assumed to reflect low-salt-induced NKCC2 splicing in both cases, luminal [Cl−] at the macula densa ranged from 18 mM at 3.6 nl/min to 48 mM at 8.0 nl/min. The predicted Vbl as a function of mTAL inflow rates is shown in Fig. 5. The low-salt diet elicits an increase in both the amount and relative proportion of NKCC2-B at the macula densa. Since NKCC2-B has a higher affinity to NH4+ compared with NKCC2-A, the model predicts that the shift acidifies the macula densa cell, thus stimulating Na+ entry via the Na+/H+ exchanger; the subsequent activation of Na+-K+-ATPase pumps slightly hyperpolarizes the basolateral membrane. Our results also suggest that the impact of the isoform shift on macula densa Vbl decreases with increasing flow rate, that is, with increasing luminal NaCl. The Vbl reduction is blunted as the flow rate is raised because luminal and cytosolic [Na+] and [NH4+] are then higher; the isoform switch thus has lesser effects on ionic fluxes across NKCC2, NHE and Na+-K+-ATPase. Together, these results indicate that for the same luminal [Cl−] at the macula densa, the low-salt-induced shift from NKCC2-A to NKCC2-B at the macula densa hyperpolarizes the basolateral membrane.
Fig. 5.
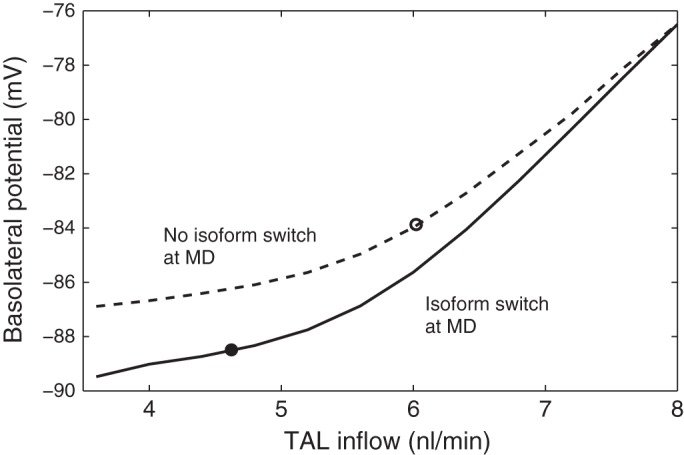
Basolateral membrane potential as a function of TAL inflow. Solid line, baseline Na+-K+-2 Cl− cotransporter NKCC2 isoform expression at the MD; dashed line, low-salt-induced expression at the MD. As the TAL inflow is raised from 3.6 to 8.0 nl/min, luminal [Cl−] increases from 18 to 48 mM. ● and ○: operating points for low- and normal-salt flow, respectively. The location of the low-salt operating point may prevent excessive salt loss.
In Fig. 5, the operating point corresponding to the low-salt case with the NKCC2 isoform switch in both the TAL and macula densa is assumed to be at 4.6 nl/min and is marked by the closed circle; the operating point for normal-salt flow, which is assumed to be at 6 nl/min, is marked by the open circle. The normal-salt operating point lies in the steeper part of the response curve, so that TGF is similarly sensitive to both increases and decreases in tubular flow. In contrast, the low-salt operating point lies in the relatively flat portion of the curve, which is concave upward. The implication for hemodynamic control is that, in the low-salt case, further reductions in tubular flow induce a relatively small degree of hyperpolarization, thereby preventing excessive dilation and increases in filtration. Conversely, significant increases in tubular flow result in a pronounced constrictive response. The impact of a low-salt diet on TAL NaCl transport and TGF signal is summarized in Fig. 6.
Fig. 6.
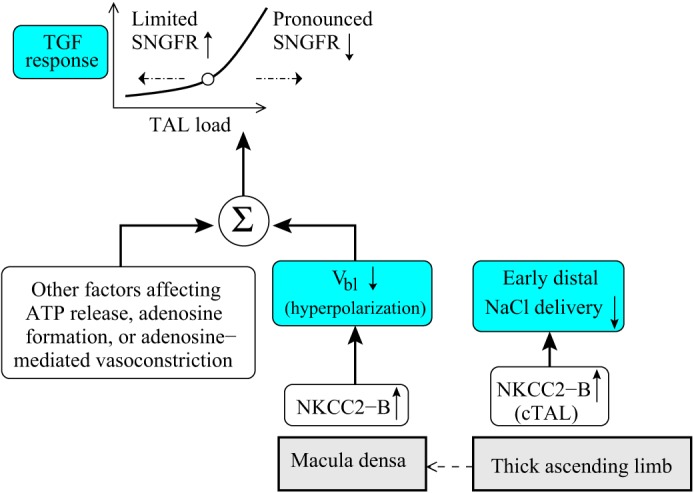
Impact of a low-salt diet on thick ascending limb NaCl transport and tubuloglomerular feedback (TGF) signal. The low-salt-induced elevation in NKCC2-B expression along the cortical thick ascending limb (cTAL) decreases early distal NaCl delivery. The NKCC2 isoform shift also hyperpolarizes the basolateral membrane (Vbl) of macula densa cells, which, taken in isolation, may inhibit the TGF signal (basolateral ATP release). A low-salt diet may have other effects on the TGF signal (e.g., enhanced adenosine formation via 5′-ectonucleotidase; facilitation of adenosine-mediated vasoconstriction by angiotensin II), which eventually yield a lower glomerular filtration rate and tubular load. An asymmetric TGF response, such that further reductions in flow induce limited vasodilation, whereas elevations in flow result in pronounced decreases in filtration, may prevent excessive salt loss. SNGFR, single nephron glomerular filtration rate.
DISCUSSION
Macula densa model.
We expanded the TAL model developed by Weinstein (46) and Weinstein and Krahn (48) to include the macula densa. Our model of rat macula densa cells reproduces experimental measurements in rabbit macula densa cells over a range of conditions. Predicted variations in basolateral membrane potential (Vbl) and intracellular concentrations, following changes in bath or lumen composition, generally agree well with measured values. Under ex vivo conditions, the predicted [Na+]i is ∼10 mM lower than measured values in rabbits. This discrepancy may be due to interspecies differences; as noted above, the abundance of Na+-K+-ATPase pumps may vary significantly between rabbits and rodents.
Our simulations indicate that under normal- and low-flow conditions, the rate of NaCl transport across NKCC2 is limited at the macula densa, that is, NKCC2 operates near its equilibrium state. At normal-flow rates, with luminal [K+] and [NH4+] ∼2 mM, the model predicts that the cotransporter mediates the net secretion of K+ in parallel with the net reabsorption of Na+, Cl−, and NH4+ and this across both isoforms A and B. At low flow (i.e., when luminal [NaCl] is ∼10 mM lower), NKCC2-A and NKCC2-B are predicted to carry Na+, Cl−, and K+ from the cytosol into the lumen and NH4+ in the opposite direction.
Adaptation to low-salt diet.
A major motivation behind this work was to assess whether the observed modulation of the differential splicing of NKCC2 by dietary salt intake contributes to the adaptative capacity of the kidney. A low-salt diet produces a shift from NKCC2-A to NKCC2-B both in the outer medulla and the cortex (36). Because NKCC2-B has the highest affinity to Cl− among the three isoforms, the shift should in principle enhance NaCl reabsorption by the TAL and promote salt retention. We found that per se, low-salt diet-induced variations in NKCC2 expression significantly increase transepithelial NaCl transport along the TAL; by themselves, these variations are predicted to reduce the delivery of Na+ and Cl− to the macula densa by 3.7 and 12.5%, respectively.
NKCC2 is likely not the only rate-limiting factor in salt reabsorption. As noted, a low-salt diet not only alters the relative distribution of NKCC2 isoforms, it also increases the overall abundance of the transporter (36). While this net increase in NKCC2 expression (about 30% in the mTAL and 100% in the cTAL) augments NaCl reabsorption in the TAL, the basolateral sodium pump also represents a rate-limiting step for transport. It has been argued that transporter density changes that are restricted to the luminal membrane cannot generate significant variations in transcellular fluxes (47). In the distal nephron, for example, aldosterone regulates Na+ transport by controlling the transcription of both apical epithelial Na+ (ENaC) channels and basolateral Na+-K+-ATPase pumps (8). When we assumed that the expression of Na+-K+-ATPase increases in parallel with that of NKCC2, the model predicted larger reductions in the delivery of Na+ and Cl− to the macula densa (Table 5).
TGF signal.
TGF is a negative feedback process in which the Cl− concentration of the tubular fluid flowing past the macula densa is sensed and the single nephron glomerular filtration rate is adjusted accordingly. Aspects of the TGF signaling pathway remain controversial. Nonetheless, there is strong evidence that ATP is released by the macula densa cells while adenosine, via activation of adenosine A1 receptors on mesangial cells and/or afferent arteriole smooth muscle cells, induces final vasoconstriction (2, 6, 40, 41). ATP can be released via maxi-anion channels present on the basolateral membrane of macula densa cells (2). The molecular nature of these channels and the mechanisms that control the opening and closure of this ATP pathway remain unclear; one proposed mechanism is the depolarization of the macula densa basolateral membrane (1). Simulations using the present model suggest that low-salt-induced variations in the differential splicing of NKCC2 hyperpolarize the basolateral membrane, in particular at low-flow rates (see Fig. 5), a result that, per se, may indicate reduced ATP release.
Komlosi et al. (23) observed similar ATP release from macula densa cells around the expected operating points in rabbits fed a low-salt vs. normal-salt diet, assuming that ambient [NaCl] at the macula densa under these conditions is 20 vs. 20–40 mM, respectively. Whether a low-salt diet or salt depletion alters the effects of Vbl on basolateral ATP release is not known. These conditions, however, are expected to lower macula densa neuronal nitric oxide (NO) synthase activity and NO formation. Further, local NO formation in isolated glomeruli has been shown to suppress the activity of 5′-ectonucleotidase (35), which contributes to the critical local formation of adenosine as the final mediator of TGF (6, 18). Therefore, a low-salt diet and salt depletion may enhance 5′-ectonucleotidase activity in the juxtaglomerular apparatus, which is expected to facilitate local adenosine formation. Moreover, higher local angiotensin II concentrations will enhance the vasoconstrictor response to the generated adenosine (16). Another explanation for the discrepancy between the experimentally observed similar ATP release and the model's predicted basolateral hyperpolarization is the possibility that a low-salt diet may affect the expression or regulation of the basolateral ATP-releasing pathway. Because the molecular nature of the ATP release mechanism is not known, this explanation is speculative.
Furthermore, an analysis of operating points on the TGF response curve (see Fig. 5) suggests that further salt loss is avoided, inasmuch as a decrease in tubular flow results in limited TGF-mediated vasodilation, whereas an increase in tubular flow induces marked vasoconstriction. This result is consistent with the observation of Komlosi et al. (23) that ATP release is not significantly different between low-salt and normal-salt conditions at ambient signals (see Fig. 2 of Ref. 23; compare the furo-2 excitation ratio at 20 mM luminal [NaCl] for low salt with values at 20 and 40 mM for normal salt). What is different, however, is the markedly larger increase in ATP release in the low-salt diet group as luminal [NaCl] increases, compared with normal-salt diet group. That finding is coherent with our model's prediction that an increase in NaCl delivery to the macula densa elicits a larger relative change in Vbl (depolarization) with low-salt NKCC2 expression at the macula densa.
Under low-flow conditions, the model predicts a decrease in luminal [Na+] and [Cl−] at the macula densa, as observed experimentally. However, luminal [K+] is predicted to increase, in contrast to experimental findings by Vallon et al. (43). The source of this discrepancy has yet to be clearly determined, but there are some differences between the experimental conditions and model simulations. For instance, luminal [NH4+] differs between the present model and the experiments by Vallon et al. Given that K+ and NH4+ compete for the same binding site on NKCC2, the presence of NH4+, or the lack thereof, significantly affects K+ transport. It is also noteworthy that a previous modeling study, which uses an epithelial-based model of the TAL (28), predicted lower macula densa luminal [K+] at low tubular flow. However, that model does not account for all the transporters included in the present model; it also uses some paracellular permeabilities that differ substantially from measured values.
Our model predicts that increasing luminal [K+] depolarizes the basolateral membrane of the macula densa cells, thereby possibly inducing a constrictive TGF signal. This result suggests that if macula densa luminal [K+] varies in tandem with TAL tubular flow, then K+ can serve as a rate-limiting, positive regulator, or one of the regulators, in the TGF mechanism, as previously proposed and supported by micropuncture studies by Vallon et al. (43).
In summary, our results suggest that with luminal solute concentrations consistent with in vivo conditions near the macula densa, NKCC2 operates near its equilibrium state and reverses direction when TAL flow is sufficiently reduced. Furthermore, the differential splicing of NKCC2 induced by a low-salt diet maintains salt balance by increasing TAL NaCl reabsorption. One limitation of this model is that it is formulated for steady state and, therefore, does not capture the TGF-mediated flow oscillations that have been observed experimentally (26). In addition, the model represents an isolated TAL, surrounded by an interstitial fluid with a fixed composition, and thus does not consider the interactions between the TAL and neighboring tubules. Nonetheless, combined with an epithelial-based model of the proximal tubule and descending limb, the present model could be used as an essential component in models of integrated renal hemodynamics and transport regulation.
GRANTS
This research was supported by National Institute of Diabetes and Digestive and Kidney Diseases Grants R01-DK-56248 and P30-DK-079337 (to V. Vallon) and R01-DK-089066 (to A. T. Layton) and National Heart, Lung, and Blood Institute Grant R01-HL-94728 (to V. Vallon); Deutsche Forschungsgemeinschaft Grant SFB 699/A4 (to H. Castrop); and Department of Veterans Affairs (to V. Vallon).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: A.E., V.V., and A.T.L. conception and design of research; A.E. and A.T.L. performed experiments; A.E., H.C., K.L., V.V., and A.T.L. analyzed data; A.E., H.C., K.L., V.V., and A.T.L. interpreted results of experiments; A.E. and A.T.L. prepared figures; A.E. and A.T.L. drafted manuscript; A.E., H.C., K.L., V.V., and A.T.L. edited and revised manuscript; A.E., H.C., K.L., V.V., and A.T.L. approved final version of manuscript.
REFERENCES
- 1.Bell PD, Komlosi P, Zhang ZR. ATP as a mediator of macula densa cell signalling. Purinergic Signal 5: 461–471, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bell PD, Lapointe JY, Sabirov R, Hayashi S, Peti-Peterdi J, Manabe KI, Kovacs G, Okada Y. Macula densa cell signaling involves ATP release through a maxi anion channel. Proc Natl Acad Sci USA 100: 4322–4327, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bell PD, Lapointe JY, Peti-Peterdi J. Macula densa cell signaling. Annu Rev Physiol 65: 481–500, 2003 [DOI] [PubMed] [Google Scholar]
- 4.Bourdeau JE, Burg MB. Voltage dependence of calcium transport in the thick ascending limb of Henle's loop. Am J Physiol Renal Fluid Electrolyte Physiol 236: F357–F364, 1979 [DOI] [PubMed] [Google Scholar]
- 5.Burg MB. Thick ascending limb of Henle's loop. Kidney Int 22: 454–464, 1982 [DOI] [PubMed] [Google Scholar]
- 6.Castrop H, Huang Y, Hashimoto S, Mizel D, Hansen P, Theilig F, Bachmann S, Deng C, Briggs J, Schnermann J. Impairment of tubuloglomerular feedback regulation of GFR in ecto-5′-nucleotidase/CD73-deficient mice. J Clin Invest 114: 634–642, 2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dixit MP, Xu L, Xu H, Bai L, Collins JF, Ghishan FK. Effect of angiotensin-II on renal Na+/H+ exchanger-NHE3 and NHE2. Biochim Biophys Acta 1664: 38–44, 2004 [DOI] [PubMed] [Google Scholar]
- 8.Dooley R, Angibaud E, Yusef YR, Thomas W, Harvey BJ. Aldosterone induced ENaC and basal Na+/K+-ATPase trafficking via protein kinase D1-phosphatidylinositol 4-kinaseIIIbeta trans Golgi signalling in M1 cortical collecting duct cells. Mol Cell Endocrinol 372: 86–95, 2013 [DOI] [PubMed] [Google Scholar]
- 9.du Cheyron D, Chalumeau C, Defontaine N, Klein C, Kellermann O, Paillard M, Poggioli J. Angiotensin II stimulates NHE3 activity by exocytic insertion of the transporter: role of PI 3-kinase. Kidney Int 64: 939–949, 2003 [DOI] [PubMed] [Google Scholar]
- 10.Gagnon E, Forbush B, Caron L, Isenring P. Functional comparison of renal Na-KCl cotransporters between distant species. Am J Physiol Cell Physiol 284: C365–C370, 2003 [DOI] [PubMed] [Google Scholar]
- 11.Gimenez I, Isenring P, Forbush B. Spatially distributed alternative splice variants of the renal Na-K-Cl cotransporter exhibit dramatically different affinities for the transported ions. J Biol Chem 277: 8767–8770, 2002 [DOI] [PubMed] [Google Scholar]
- 12.Gonzalez E, Salomonsson M, Muller-Suur C, Persson AE. Measurements of macula densa cell volume changes in isolated and perfused rabbit cortical thick ascending limb. II. Apical and basolateral cell osmotic water permeabilities. Acta Physiol Scand 133: 159–166, 1988 [DOI] [PubMed] [Google Scholar]
- 13.Good DW, Watts BA., 3rd Function roles of apical membrane Na+/H+ exchange at the medullary thick ascending limb. Am J Physiol Renal Fluid Electrolyte Physiol 270: F691–F699, 1996 [DOI] [PubMed] [Google Scholar]
- 14.Greger R, Bleich M, Schlatter E. Ion channels in the thick ascending limb of Henle's loop. Renal Physiol Biochem 13: 37–50, 1990 [DOI] [PubMed] [Google Scholar]
- 15.Greger R, Schlatter E. Presence of luminal K+, a prerequisite for active NaCl transport in the cortical thick ascending limb of Henle's loop of rabbit kidney. Pflügers Arch 392: 92–94, 1981 [DOI] [PubMed] [Google Scholar]
- 16.Hansen PB, Hashimoto S, Briggs J, Schnermann J. Attenuated renovascular constrictor responses to angiotensin II in adenosine 1 receptor knockout mice. Am J Physiol Regul Integr Comp Physiol 285: R44–R49, 2003 [DOI] [PubMed] [Google Scholar]
- 17.Hebert SC, Culpepper RM, Andreoli TE. NaCl transport in mouse medullary thick ascending limbs. I. Functional nephron heterogeneity and ADH-stimulated NaCl cotransport. Am J Physiol Renal Fluid Electrolyte Physiol 241: F412–F431, 1981 [DOI] [PubMed] [Google Scholar]
- 18.Huang DY, Vallon V, Zimmermann H, Koszalka P, Schrader J, Osswald H. Ecto-5′-nucleotidase (cd73)-dependent and -independent generation of adenosine participates in the mediation of tubuloglomerular feedback in vivo. Am J Physiol Renal Physiol 291: F282–F288, 2006 [DOI] [PubMed] [Google Scholar]
- 19.Hurst AM, Lapointe JY, Laamarti A, Bell PD. Basic properties and potential regulators of the apical K+ channel in macula densa cells. J Gen Physiol 103: 1055- 1070, 1994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jaeger P, Karlmark B, Giebisch G. Ammonium transport in the rat cortical tubule: relationship to potassium metabolism. Am J Physiol Renal Fluid Electrolyte Physiol 245: F593–F600, 1983 [DOI] [PubMed] [Google Scholar]
- 21.Kieferle S, Fong P, Bens M, Vandewalle A, Jentsch TJ. Two highly homologous members of the ClC chloride channel family in both rat and human kidney. Proc Natl Acad Sci USA 91: 6943–6947, 1994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kobayashi K, Uchida S, Mizutani S, Sasaki S, Marumo F. Intrarenal and cellular localization of CLC-K2 protein in the mouse kidney. J Am Soc Nephrol 12: 1327–1334, 2001 [DOI] [PubMed] [Google Scholar]
- 23.Komlosi P, Peti-Peterdi J, Fuson AL, Fintha A, Rosivall L, Bell PD. Macula densa basolateral ATP release is regulated by luminal [NaCl] and dietary salt intake. Am J Physiol Renal Physiol 286: F1054–F1058, 2004 [DOI] [PubMed] [Google Scholar]
- 24.Lapointe JY, Bell PD, Hurst AM, Cardinal J. Basolateral ionic permeabilities of macula densa cells. Am J Physiol Renal Fluid Electrolyte Physiol 260: F856–F860, 1991 [DOI] [PubMed] [Google Scholar]
- 25.LaPointe JY, Laamarti A, Bell PD. Ionic transport in macula densa cells. Kidney Int 54: S58–S64, 1998 [DOI] [PubMed] [Google Scholar]
- 26.Leyssac PP, Baumbach L. An oscillating intratubular pressure response to alterations in Henle loop flow in the rat kidney. Acta Physiol Scand 117: 415–419, 1983 [DOI] [PubMed] [Google Scholar]
- 27.Lorenz JN, Dostanic-Larson I, Shull GE, Lingrel JB. Ouabain inhibits tubuloglomerular feedback in mutant mice with ouabain-sensitive alpha1 Na,K-ATPase. J Am Soc Nephrol 17: 2457–2463, 2006 [DOI] [PubMed] [Google Scholar]
- 28.Nieves-Gonzalez A, Clausen C, Layton AT, Layton HE, Moore LC. Transport efficiency and workload distribution in a mathematical model of the thick ascending limb. Am J Physiol Renal Physiol 304: F653–F664, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nieves-Gonzalez A, Clausen C, Marcano M, Layton AT, Layton HE, Moore LC. Fluid dilution and efficiency of Na+ transport in a mathematical model of a thick ascending limb cell. Am J Physiol Renal Physiol 304: F634–F652, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Peti-Peterdi J, Bebok S, Lapointe JY, Bell PD. Novel regulation of cell [Na+] in macula densa cells: apical Na+ recycling by H-K-ATPase. Am J Physiol Renal Physiol 282: F324–F329, 2002 [DOI] [PubMed] [Google Scholar]
- 31.Peti-Peterdi J, Chambrey R, Bebok Z, Biemesderfer D, St John PL, Abrahamson DR, Warnock DG, Bell PD. Macula densa Na+/H+ exchange activities mediated by apical NHE2 and basolateral NHE4 isoforms. Am J Physiol Renal Physiol 278: F452–F463, 2000 [DOI] [PubMed] [Google Scholar]
- 32.Plata C, Meade P, Vazquez N, Hebert SC, Gamba G. Functional properties of the apical Na+-K+-2Cl− cotransporter isoforms. J Biol Chem 277: 11004–11012, 2002 [DOI] [PubMed] [Google Scholar]
- 33.Pu HX, Cluzeaud F, Goldshleger R, Karlish SJ, Farman N, Blostein R. Functional role and immunocytochemical localization of the gamma-a and gamma-b forms of the Na,K-ATPase gamma subunit. J Biol Chem 276: 20370–20378, 2001 [DOI] [PubMed] [Google Scholar]
- 34.Sands JM, Layton HE. Urine concentrating mechanism and its regulation. In: The Kidney: Physiology and Pathophysiology (3d ed), edited by Seldin DW, Giebisch G. Philadelphia, PA: Lippincott Williams & Wilkins, 2000, p. 1175–1216 [Google Scholar]
- 35.Satriano J, Wead L, Cardus A, Deng A, Boss GR, Thomson SC, Blantz RC. Regulation of ecto-5′-nucleotidase by NaCl and nitric oxide: potential roles in tubuloglomerular feedback and adaptation. Am J Physiol Renal Physiol 291: F1078–F1082, 2006 [DOI] [PubMed] [Google Scholar]
- 36.Schieβl IM, Rosenauer A, Kattler V, Minuth WW, Oppermann M, Castrop H. Dietary salt intake modulates differential splicing of the Na-K-2Cl cotransporter NKCC2. Am J Physiol Renal Physiol 305: F1139–F1148, 2013 [DOI] [PubMed] [Google Scholar]
- 37.Schlatter E, Salomonsson M, Persson AEG, Greger R. Macula densa cells sense luminal NaCl concentration via furosemide sensitive Na+,2Cl−,K+ cotranport. Pflügers Arch 414: 286–290, 1989 [DOI] [PubMed] [Google Scholar]
- 38.Schnermann J, Briggs J, Schubert G. In situ studies of the distal convoluted tubule in the rat. I. Evidence for NaCl secretion. Am J Physiol Renal Fluid Electrolyte Physiol 243: F160–F166, 1982 [DOI] [PubMed] [Google Scholar]
- 39.Simon E, Martina D, Buerkert J. Contribution of individual superficial nephron segments to ammonium handling in chronic metabolic acidosis in the rat. J Clin Invest 76: 855–864, 1984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sun D, Samuelson LC, Yang T, Huang Y, Paliege A, Saunders T, Briggs J, Schnermann J. Mediation of tubuloglomerular feedback by adenosine: evidence from mice lacking adenosine 1 receptors. Proc Natl Acad Sci USA 98: 9983–9988, 2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Thomson S, Bao D, Deng A, Vallon V. Adenosine formed by 5′-nucleotidase mediates tubuloglomerular feedback. J Clin Invest 106: 289–298, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Thurau K, Schnermann J. The sodium concentration in the macula densa cells as a regulating factor for glomerular filtration (micropuncture experiments). Klin Wochenschr 43: 410–413, 1965 [DOI] [PubMed] [Google Scholar]
- 43.Vallon V, Osswald H, Blantz RC, Thomson S. Potential role of luminal potassium in tubuglomerular feedback. J Am Soc Nephrol 8: 1831–1837, 1997 [DOI] [PubMed] [Google Scholar]
- 44.Vallon V, Richter K, Blantz RC, Thomson S, Osswald H. Glomerular hyperfiltration in experimental diabetes mellitus: potential role of tubular reabsorption. J Am Soc Nephrol 10: 2569–2576, 1999 [DOI] [PubMed] [Google Scholar]
- 45.Wang T, Chan YL. Mechanism of angiotensin II action on proximal tubular transport. J Pharmacol Exp Ther 252: 689–695, 1990 [PubMed] [Google Scholar]
- 46.Weinstein AM. A mathematical model of rat ascending Henle limb. III. Tubular function. Am J Physiol Renal Physiol 298: F543–F556, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Weinstein AM. Potassium excretion during antinatriuresis: perspective from a distal nephron model. Am J Physiol Renal Physiol 302: F658–F673, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Weinstein AM, Krahn TA. A mathematical model of rat ascending Henle limb. II. Epithelial function. Am J Physiol Renal Physiol 298: F525–F542, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wetzel RK, Sweadner KJ. Immunocytochemical localization of Na-K-ATPase α- and β-subunits in rat kidney. Am J Physiol Renal Physiol 281: F531–F545, 2001 [DOI] [PubMed] [Google Scholar]
- 50.Wulff P, Vallon V, Huang DY, Volkl H, Yu F, Richter K, Jansen M, Schlunz M, Klingel K, Loffing J, Kauselmann G, Bosl MR, Lang F, Kuhl D. Impaired renal Na+ retention in the sgk1-knockout mouse. J Clin Invest 110: 1263–1268, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yang T, Huang YG, Singh I, Schnermann J, Briggs JP. Localization of bumetanide and thiazide-sensitive Na-K-Cl cotransporters along the rat nephron. Am J Physiol Renal Fluid Electrolyte Physiol 271: F931–F939, 1996 [DOI] [PubMed] [Google Scholar]
- 52.Yu FH, Shull GE, Orlowski J. Functional properties of the rat Na/H exhanger NHE-2 isoform expressed in Na/H exchanger-deficient Chinese hamster ovary cells. J Biol Chem 268: 35536–25541, 1993 [PubMed] [Google Scholar]