

Abstract
In the thymus, T cell maturation is influenced by cholinergic signaling, and the predominantly expressed receptor is the α3-subunit of nicotinic acetylcholine receptors, encoded by the chrna3 gene. We here determined its cellular distribution utilizing an appropriate eGFP-expressing reporter mouse strain. Neither T cells (CD4, CD8) nor mesenchymal cells (desmin-positive) expressed eGFP. In the thymic medulla, eGFP-positive cells either were scattered or, more frequently, formed small clusters resembling Hassall's corpuscles. Immunolabeling revealed that these cells were indeed terminally differentiated epithelial cells expressing keratin 10 (K10) but neither typical cortical (K8, K18) nor medullary keratins (K5, K14). These labeling patterns reflected those in the epidermis of the skin, where overlap of K10 and eGFP expression was seen in the stratum granulosum, whereas underlying basal cells displayed K5-immunoreactivity. A substantial portion of thymic eGFP-positive cells was also immunoreactive to chromogranin A, a peptide previously reported in epidermal keratinocytes in the stratum granulosum. Its fragment catestatin has multiple biological activities, including suppression of proinflammatory cytokine release from macrophages and inhibition of α3β4 nAChR. The present findings suggest that its thymic production and/or release are under cholinergic control involving nAChR containing the α3-subunit.
1. Introduction
The thymus is the site of step-wise maturation of naïve T cells from immature thymocytes which occurs along with positive and negative selection processes while thymocytes migrate from the cortex to the medulla. These processes are influenced by cholinergic signaling [1–4], and acetylcholine (ACh) is endogenously synthesized in the thymus [5–7].
Signaling via nicotinic ACh receptors (nAChR) has received particular attention. These receptors are pentamers composed of various subunit combinations. The “muscle type” nAChR originally identified at the motor endplate consists of two α1-, one β1-, one ε- (or γ- at fetal stage), and one δ-subunit, and these are also expressed by myoid [8, 9] and epithelial cells of the thymic medulla [10–13]. Thymic expression and presentation of muscle-type nAChR subunits have been associated with a frequent (85%) variant of myasthenia gravis, a disease of the motor endplate, where autoantibodies against such subunits are formed, as the thymus frequently shows abnormal structure in this condition and thymectomy is beneficial for the patients [14].
“Neuronal” nAChR are homo- or heteromers of α-subunits 2–7 and 9-10 (α8 is expressed only in chicken) and β-subunits 2–4 [15–17]. Despite their designation as “neuronal” they are widely expressed outside the nervous system including the thymus [18, 19]. Amongst them, subunits α3, α5, and β4 exhibit highest expression in early postnatal mouse thymus, reaching 7–15% of mRNA content found in brain as standard [19]. Their genes—chrna3, chrna5, and chrnb4—are clustered on chromosome 9 in mice, and, when coexpressed, the translated proteins assemble to functional α3(α5)β4 nAChR with either 2 α- and 3 β-subunit or 3 α- and 2 β-subunit chains [15, 20]. The α3-subunit is essential for receptor function and may occur with or without an additional α5-subunit in these receptors while the α5-subunit does not form functional receptors without additional α-subunits [15]. Messenger RNAs coding for these subunits have been detected in isolated thymocytes and cultured thymic epithelial cells (TEC) obtained from children undergoing corrective cardiac surgery [18], but it is still unclear which specific thymic cell types express these receptors in situ.
In view of the reported specificity problems associated with immunohistochemical detection of nAChR subunits [21], we utilized a reporter mouse strain expressing eGFP under the control of the chrna3 promoter coding for the essential α3-subunit [22]. To further characterize and identify eGFP-positive cells in the thymus, tissue sections were subjected to immunohistochemistry with marker antibodies for subsets of thymocytes (CD4, CD8), myoid and mesenchymal (desmin) and thymic epithelial cells. Thymic epithelial cells are heterogeneous and can be classified according to various criteria. A broad classification divides them into four general types (with further subpopulations [23, 24]): (a) subcapsular/paraseptal/perivascular, (b) cortical, (c) medullary (mTEC), and (d) terminally differentiated mTEC, usually arranged in Hassall's corpuscles [25–28]. These types differ in their intermediate filament content, with cortical and a small population of mTEC expressing keratins typical for simple epithelia (K8, K18) [23, 28, 29], the majority of mTEC keratins characteristic for immature basal cells of stratified epithelia, that is, K5 and K14 [24, 28, 30], and terminally differentiated cells of Hassall's corpuscles (in human) and Hassall's corpuscles-like structures (in mice) expressing K10 [31].
2. Materials and Methods
2.1. Animals and Tissue Collection
Transgenic mice expressing eGFP under chrna3 promoter [22] were killed with an overdose of isoflurane (Abbott, Wiesbaden, Germany). Animals of either gender (N = 9) were transcardially perfused with heparin-containing rinsing solution [32] followed by either 4% paraformaldehyde in 0.1 M phosphate buffer (pH = 7.4) or Zamboni fixative (2% paraformaldehyde, 15% saturated picric acid in 0.1 M phosphate buffer, pH 7.4). Thymi and hairy skin of the head were dissected and fixed overnight by immersion in the same fixative used for perfusion. In 5 additional animals, tissues were immersion-fixed overnight in Zamboni fixative without prior perfusion. Specimens were washed in 0.1 M phosphate buffer, pH = 7.4, for 30 h, incubated overnight in 18% sucrose in 0.1 M phosphate buffer, and frozen in OCT compound (Sakura Finetek, Staufen, Germany) using liquid nitrogen.
2.2. Immunohistochemistry
Thymi and hairy skin of the head were cut with a cryostat into 4–10 μm thick sections and air-dried for 1 h. Nonspecific protein binding sites were saturated with 10% horse serum, 0.5% Tween, and 0.1% BSA in PBS (0.005 M phosphate buffer, pH = 7.4, with 0.45% NaCl). Sections were incubated for 16 h with primary antibodies diluted in PBS with doubled salt concentration and containing 0.01% NaN3. Each antibody was applied to thymi from at least 6 animals. Antibody dilutions and sources are specified in Table 1. After washing in PBS, sections were covered with Cy3-conjugated donkey anti-rabbit IgG (1 : 2000; Chemicon, Darmstadt, Germany) or donkey anti-rat IgG (1 : 1000; Dianova, Hamburg, Germany) for 1 h, washed, postfixed in 4% paraformaldehyde in 0.1 M phosphate buffer, washed again, and mounted in carbonate-buffered glycerol (1 : 1, pH = 8.6). Controls were run by replacing primary antibodies with unrelated isotypes from the same species and by omission of first antibodies. Sections were evaluated with an Axioplan 2 epifluorescence microscope equipped with an AxioCam MRm camera system (Zeiss, Jena, Germany). Except overall adjustment of brightness, no further manipulations of digital images were performed.
Table 1.
Primary antibodies used in the study.
| Target | Immunogen | Host | Clone | Dilution | Catalog number | Company |
|---|---|---|---|---|---|---|
| CD4 | Not specified | Rat | Monoclonal, clone RM4-5 |
1 : 400 | IH93-0042-91 | eBioscience, Frankfurt am Main, Germany |
|
| ||||||
| CD8 | Not specified | Rat | Monoclonal, clone 53-6.7 |
1 : 400 | IH93-0081-91 | eBioscience, Frankfurt am Main, Germany |
|
| ||||||
| Chromogranin A | Synthetic peptide corresponding to residues near the C-terminus of human chromogranin A | Rabbit | Polyclonal | 1 : 25 | 1782-1 | Epitomics, Cambridge, UK |
|
| ||||||
| Desmin | Synthetic peptide corresponding to C-terminus of human desmin | Rabbit | Monoclonal, clone Y66 |
1 : 200–1 : 800 | 04-585 | Merck Millipore, Darmstadt, Germany |
|
| ||||||
| Keratin 5 | Synthetic peptide corresponding to C-terminus of human keratin 5 | Rabbit | Monoclonal, clone SP27 |
1 : 50–1 : 200 | SPB-M3270 | Spring, Pleasanton, CA, USA |
|
| ||||||
| Keratin 8 | Synthetic peptide corresponding to C-terminus of human keratin 8 | Rabbit | Monoclonal, clone SP102 |
1 : 50 | SPB-M4020 | Spring, Pleasanton, CA, USA |
|
| ||||||
| Keratin 10 | C-terminus of the mouse keratin 10 | Rabbit | Polyclonal | 1 : 400 | PRB-159P | Covance, Münster, Germany |
|
| ||||||
| Keratin 14 | Synthetic peptide corresponding to C-terminus of human keratin 14 | Rabbit | Monoclonal, clone SP53 |
1 : 400 | SPB-M3534 | Spring, Pleasanton, CA, USA |
|
| ||||||
| Keratin 18 | Synthetic peptide corresponding to C-terminus of human keratin 18 | Rabbit | Monoclonal, clone SP69 |
1 : 200 | SPB-M3694 | Spring, Pleasanton, CA, USA |
3. Results
The major nicotinic receptor mediating synaptic transmission in autonomic ganglia is a heteropentamer of α3(α5)β4 subunits [15, 33]. Accordingly, autonomic nerve fibers surrounding thymic arteries exhibited intense eGFP fluorescence (Figure 1(a)). Besides these nerve fibers, positive cells were observed in the thymic medulla with some preference to the corticomedullary junction, but practically sparing the external cortex (Figures 1(b) and 1(c)). Such cells typically formed clusters. The larger clusters showed characteristic morphology of murine Hassall's corpuscle-like structures. Infrequently observed singular eGFP-positive cells and those forming groups of two or three, displayed variable shape, ranging from round to oval to elongated with processes emerging from the cell body (Figures 1(d)–1(i)). According to this complex shape, these eGFP-positive cells were not thymocytes and did not express CD4 or CD8 (Figure 2).
Figure 1.
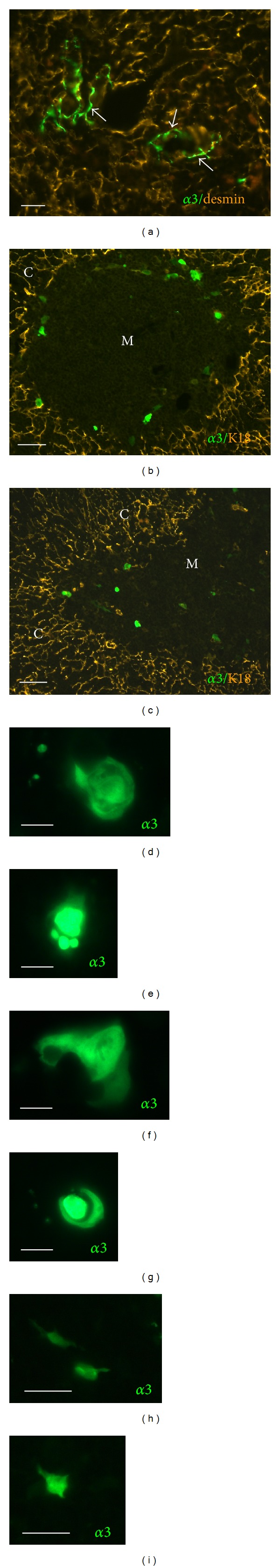
Chrna3-driven eGFP expression in the thymic medulla. (a) Perivascular nerve fibers (arrows) are α3-subunit-positive. Mesenchymal elements labeled with desmin antibody for better orientation (scale bar, 20 μm). (b), (c) α3-subunit-expressing cells are located in the thymic medulla and at the corticomedullary junction. Cortex delineated by immunolabeling with K18-antibodies (scale bar, 50 μm). (d)–(g) eGFP-Positive cells either form Hassall's corpuscles-like aggregates (scale bar, 10 μm) or lie singularly (h), (i) (scale bar, 20 μm). M: medulla; C: cortex.
Figure 2.
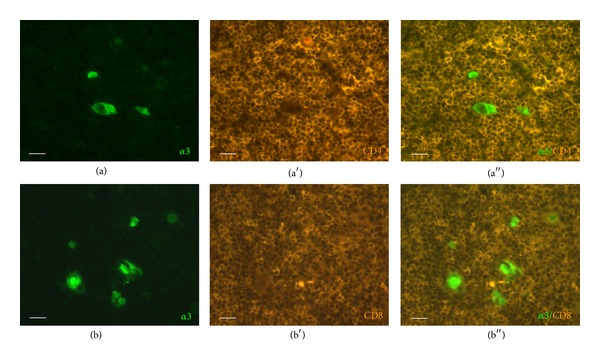
eGFP-positive cells are not thymocytes. Immunostaining for CD4 (a)–(a′′) and CD8 (b)–(b′′) revealed no colocalization with eGFP signal. Scale bar, 20 μm.
Desmin is a marker for thymic myoid cells and mesenchymal cells [9]. Accordingly, immunolabeling revealed a network of fine processes in the thymic medulla and a few positive cells bodies. Colocalization with eGFP fluorescence was not observed (Figure 3).
Figure 3.
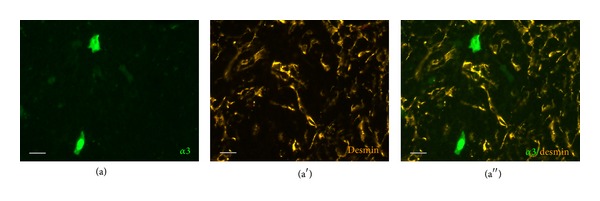
Alpha3-subunit-expressing cells are neither myoid nor mesenchymal cells, since there is no colocalization with desmin-immunoreactivity (scale bar, 20 μm).
K8- and K18-immunoreactive epithelial cell processes formed a dense network in the cortex, and a small population of less branched mTEC was also K8- and K18-immunoreactive, as previously described [34]. Neither of these antibodies, however, labeled eGFP-positive cells (Figures 4(a) and 4(b)). In the medulla, a dense mesh of K5- and K14-immunoreactive cell processes was noted. Such processes surrounded eGFP-positive cells which were larger than keratin-immunoreactive cell bodies and were K5/K14-negative (Figures 4(c) and 4(d)). This spatial arrangement resembled that of the epidermis where K5 is expressed by the basal cells and its expression ceases when keratinocytes terminally differentiate in the stratum granulosum [35, 36]. Accordingly, we observed eGFP fluorescence in the K5-negative stratum granulosum of hairy skin and K5-positive cells in the underlying stratum basale and the lowermost stratum spinosum (Figure 5(a)). In the epidermis, keratinocytes switch to K10 expression in the stratum spinosum which is kept in the stratum granulosum. This partly overlaps with eGFP fluorescence which is first seen in the stratum granulosum and extends into the stratum corneum (Figure 5(b)). The same pattern of extensive but not complete overlap of eGFP- and K10-immunoreactivity was found in the thymus (Figure 5(c)).
Figure 4.
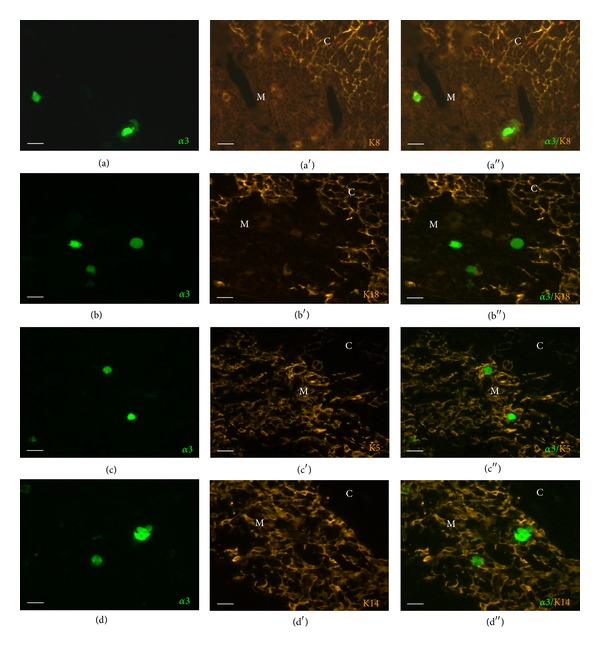
Neither cortex- (K8, K18) nor medulla-specific (K5, K15) cytokeratins are expressed by α3-subunit-positive cells. Immunostaining with K8 (a)–(a′′), K18 (b)–(b′′), K5 (c)–(c′′), and K14 (d)–(d′′) antibodies. Scale bar, 20 μm.
Figure 5.
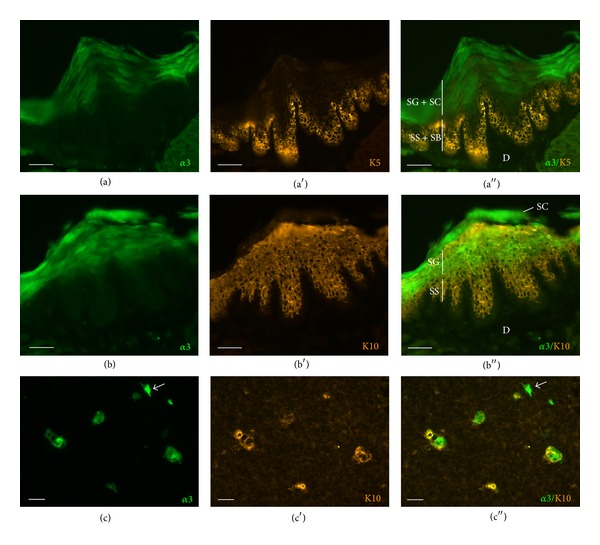
Alpha3-subunit-positive cells are terminally differentiated epithelial cells. (a)–(a′′) Chrna3-driven eGFP fluorescence in the stratum granulosum (SG) and stratum corneum (SC) of the epidermis, K5-immunoreactivity in the lowermost stratum spinosum (SS) and stratum basale (SB). D: dermis. Scale bar, 50 μm. (b)–(b′′) K10-immunoreactivity in stratum granulosum and stratum spinosum, overlapping with eGFP signal in stratum granulosum. Scale bar, 50 μm. (c)–(c′′) The majority of α3-subunit-expressing cells in the thymus are K10-immunoreactive; arrow points to an infrequently occurring K10-negative cell. Scale bar, 20 μm.
In addition, there was partial colocalization of eGFP with chromogranin A (CGA) in the thymus. CGA-immunoreactive cells were found mainly in the medulla, often attached to Hassall's corpuscle-like structures, as previously reported [37, 38] (Figure 6). CGA-immunoreactive granules were observed in a population of eGFP-positive cells, although CGA+/eGFP− and CGA−/eGFP+ cells were also present in about equal proportions (Figure 6).
Figure 6.

Immunolabeling for CGA. Both double- (eGFP+/CGA+, arrowhead) and single-labeled cells (eGFP+/CGA−, double-headed arrow; eGFP−/CGA+, arrow) can be observed. CGA-immunoreactive and eGFP-positive cells occur together in Hassall's corpuscle-like structures (higher magnification in inset). Scale bar, 20 μm.
4. Discussion
The present study demonstrates that the most abundantly expressed nAChR subunit in the murine thymus, the α3-subunit, is localized to a distinct mTEC type, that is, terminally differentiated mTEC expressing K10. In the skin, eGFP expression driven by the chrna3 promoter was more restricted than expected from previous immunohistochemical studies utilizing α3-subunit antibodies [39, 40], which possibly originates from the noted specificity problems of nAChR subunit antibodies documented by the use of respective gene-deficient mice [21]. In the epidermis, where keratinocytes undergo stepwise differentiation while migrating through the different layers towards the surface, we saw K10 prior to eGFP expression and eGFP shortly after keratinization when K10 was no longer detectable. This sequence of expression (K10 before nAChRα3) suggests that the known promoting effect of nicotine on K10 expression by keratinocytes [41] is not driven by α3-subunit containing nAChR but likely by another nAChR subtype, for example, α7 or α9α10 nAChR, also known to be expressed in keratinocytes [39, 41–43].
Assuming the same sequence of differentiation in Hassall's corpuscle-like structures, the few K10+/eGFP− and K10−/eGFP+ cells in the thymic medulla would not represent an entirely distinct cell population but mTEC at corresponding intermediate stages of differentiation. When K10 is ectopically expressed in K5-positive mTEC, the thymus presents premature involution with increased apoptosis and reduced proliferation of thymocytes [44] but this does not necessarily allow conclusions about physiologically K10-expressing terminal mTEC.
Terminally differentiated mTEC, also characterized by expression of involucrin, no longer express the autoimmune regulator (Aire) which plays a pivotal role in establishing self-tolerance by negative selection and FoxP3+ regulatory T cell (Treg) production [31]. Human Hassall's corpuscles express thymic stromal lymphopoietin which activates medullary dendritic cells to express high levels of CD28 ligands (CD80 and CD86) [45]. Such activated dendritic cells induce the generation of CD4+CD8−CD25+ T cells, leading to the suggestion that Hassall's corpuscles, via dendritic cells, trigger secondary positive selection of medium-to-high affinity self-reactive T cells resulting in Treg generation in the thymic medulla [45]. In mice, however, Treg develop normally in lymphotoxin β-receptor-deficient mice which lack terminally differentiated mTEC as judged from the absence of involucrin+ cells in the medulla [31, 46]. Thus, the function of terminally differentiated mTEC clusters is still unclear.
CGA, originally isolated from chromaffin cells of the adrenal medulla and later found to be produced by other endocrine cell types, is also expressed in epidermal keratinocytes in the stratum granulosum [47] which is again in parallel to CGA-immunoreactivity in Hassall's corpuscles [37, 38]. Its peptide fragment catestatin has multiple biological activities, including suppression of proinflammatory cytokine release by and increasing p-STAT3 levels in peritoneal and bone marrow-derived macrophages [48]. The present findings suggest that its thymic production and/or release are under cholinergic control involving nAChR containing the α3-subunit. On the other hand, catestatin is an inhibitor of α3β4 nAChR [49], so that it might act in an autoinhibitory feedback in α3-subunit-expressing terminally differentiated mTEC.
Except terminally differentiated mTEC and autonomic nerve endings, no further cellular elements of the thymus expressed chrna3-driven eGFP in the present study. In contrast, RT-PCR revealed mRNA coding for α3- and β4-subunits in isolated human CD4+CD8+ thymocytes [18]. This discrepancy might be due to species differences or incomplete expression of the transgene in the mouse strain used in this study.
In conclusion, the present study identifies nAChR α3-subunit expression by terminally differentiated mTEC. Their function is still unclear, which has been ascribed at least partly to the lack of adequate models allowing for isolating these cells [31], a problem that might be overcome utilizing the nAChRα3BAC-eGFP mouse strain analyzed in this study.
Acknowledgments
The authors thank Dr. Ines Ibanez-Tallon, MDC Berlin, Germany, for providing the Chrna3BAC-eGFP mouse strain, and M. Bodenbenner, A. Goldenberg, T. Papadakis, and L. Renno for skillful technical assistance. This study was supported by HMWK, LOEWE Research Focus Non-neuronal cholinergic systems.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Middlebrook AJ, Martina C, Chang Y, Lukas RJ, DeLuca D. Effects of nicotine exposure on T cell development in fetal thymus organ culture: arrest of T cell maturation. The Journal of Immunology. 2002;169(6):2915–2924. doi: 10.4049/jimmunol.169.6.2915. [DOI] [PubMed] [Google Scholar]
- 2.Antonica A, Magni F, Mearini L, Paolocci N. Vagal control of lymphocyte release from rat thymus. Journal of the Autonomic Nervous System. 1994;48(3):187–197. doi: 10.1016/0165-1838(94)90047-7. [DOI] [PubMed] [Google Scholar]
- 3.Maslinski W, Grabczewska E, Laskowska-Bozek H, Ryzewski J. Expression of muscarinic cholinergic receptors during T cell maturation in the thymus. European Journal of Immunology. 1987;17(7):1059–1063. doi: 10.1002/eji.1830170724. [DOI] [PubMed] [Google Scholar]
- 4.Nordman JC, Muldoon P, Clark S, Damaj MI, Kabbani N. The alpha-4 nicotinic receptor promotes CD4+ T-cell proliferation and a helper T-cell immune response. Molecular Pharmacology. 2014;85(1):50–61. doi: 10.1124/mol.113.088484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rinner I, Globerson A, Kawashima K, Korsatko W, Schauenstein K. A possible role for acetylcholine in the dialogue between thymocytes and thymic stroma. NeuroImmunoModulation. 1999;6(1-2):51–55. doi: 10.1159/000026364. [DOI] [PubMed] [Google Scholar]
- 6.Tria MA, Vantini G, Fiori MG, Rossi A. Choline acetyltransferase activity in murine thymus. Journal of Neuroscience Research. 1992;31(2):380–386. doi: 10.1002/jnr.490310221. [DOI] [PubMed] [Google Scholar]
- 7.Kawashima K, Fujii T. Expression of non-neuronal acetylcholine in lymphocytes and its contribution to the regulation of immune function. Frontiers in Bioscience. 2004;9:2063–2085. doi: 10.2741/1390. [DOI] [PubMed] [Google Scholar]
- 8.Schluep M, Willcox N, Vincent A, Dhoot GK, Newsom-Davis J. Acetylcholine receptors in human thymic myoid cells in situ: an immunohistological study. Annals of Neurology. 1987;22(2):212–222. doi: 10.1002/ana.410220205. [DOI] [PubMed] [Google Scholar]
- 9.Wakkach A, Poea S, Chastre E, et al. Establishment of a human thymic myoid cell line: phenotypic and functional characteristics. The American Journal of Pathology. 1999;155(4):1229–1240. doi: 10.1016/S0002-9440(10)65225-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Engel EK, Trotter JL, McFarlin DE, McIntosh CL. Thymic epithelial cell contains acetylcholine receptor. The Lancet. 1977;1(8025):1310–1311. doi: 10.1016/s0140-6736(77)91343-5. [DOI] [PubMed] [Google Scholar]
- 11.Wheatley LM, Urso D, Tumas K, Maltzman J, Loh E, Levinson AI. Molecular evidence for the expression of nicotinic acetylcholine receptor α-chain in mouse thymus. The Journal of Immunology. 1992;148(10):3105–3109. [PubMed] [Google Scholar]
- 12.Wakkach A, Guyon T, Bruand C, Tzartos S, Cohen-Kaminsky S, Berrih-Aknin S. Expression of acetylcholine receptor genes in human thymic epithelial cells: implications for myasthenia gravis. The Journal of Immunology. 1996;157(8):3752–3760. [PubMed] [Google Scholar]
- 13.Bruno R, Sabater L, Tolosa E, et al. Different patterns of nicotinic acetylcholine receptor subunit transcription in human thymus. Journal of Neuroimmunology. 2004;149(1-2):147–159. doi: 10.1016/j.jneuroim.2003.11.022. [DOI] [PubMed] [Google Scholar]
- 14.Poëa-Guyon S, Christadoss P, le Panse R, et al. Effects of cytokines on acetylcholine receptor expression: implications for myasthenia gravis. The Journal of Immunology. 2005;174(10):5941–5949. doi: 10.4049/jimmunol.174.10.5941. [DOI] [PubMed] [Google Scholar]
- 15.Lukas RJ, Changeux JP, Le Novère N, et al. International union of pharmacology. XX. Current status of the nomenclature for nicotinic acetylcholine receptors and their subunits. Pharmacological Reviews. 1999;51(2):397–401. [PubMed] [Google Scholar]
- 16.Elgoyhen AB, Vetter DE, Katz E, Rothlin CV, Heinemann SF, Boulter J. Alpha10: a determinant of nicotinic cholinergic receptor function in mammalian vestibular and cochlear mechanosensory hair cells. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(6):3501–3506. doi: 10.1073/pnas.051622798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lustig LR, Peng H, Hiel H, Yamamoto T, Fuchs PA. Molecular cloning and mapping of the human nicotinic acetylcholine receptor α10 (CHRNA10) Genomics. 2001;73(3):272–283. doi: 10.1006/geno.2000.6503. [DOI] [PubMed] [Google Scholar]
- 18.Mihovilovic M, Denning S, Mai Y, Whichard LP, Patel DD, Roses AD. Thymocytes and cultured thymic epithelial cells express transcripts encoding α-3, α-5 and β-4 subunits of neuronal nicotinic acetylcholine receptors: preferential transcription of the α-3 and β-4 genes by immature CD4+8+ thymocytes. Journal of Neuroimmunology. 1997;79(2):176–184. doi: 10.1016/s0165-5728(97)00120-3. [DOI] [PubMed] [Google Scholar]
- 19.Kuo Y, Lucero L, Michaels J, DeLuca D, Lukas RJ. Differential expression of nicotinic acetylcholine receptor subunits in fetal and neonatal mouse thymus. Journal of Neuroimmunology. 2002;130(1-2):140–154. doi: 10.1016/s0165-5728(02)00220-5. [DOI] [PubMed] [Google Scholar]
- 20.Mazzo F, Pistillo F, Grazioso G, et al. Nicotine-modulated subunit stoichiometry affects stability and trafficking of α3/β4 nicotinic receptor. The Journal of Neuroscience. 2013;33(30):12316–12328. doi: 10.1523/JNEUROSCI.2393-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Moser N, Mechawar N, Jones I, et al. Evaluating the suitability of nicotinic acetylcholine receptor antibodies for standard immunodetection procedures. Journal of Neurochemistry. 2007;102(2):479–492. doi: 10.1111/j.1471-4159.2007.04498.x. [DOI] [PubMed] [Google Scholar]
- 22.Frahm S, Ślimak MA, Ferrarese L, et al. Aversion to nicotine is regulated by the balanced activity of β4 and α5 nicotinic receptor subunits in the medial habenula. Neuron. 2011;70(3):522–535. doi: 10.1016/j.neuron.2011.04.013. [DOI] [PubMed] [Google Scholar]
- 23.Klug DB, Carter C, Crouch E, Roop D, Conti CJ, Richie ER. Interdependence of cortical thymic epithelial cell differentiation and T-lineage commitment. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(20):11822–11827. doi: 10.1073/pnas.95.20.11822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Danzl NM, Jeong S, Choi Y, Alexandropoulos K. Identification of novel thymic epithelial cell subsets whose differentiation is regulated by RANKL and Traf6. PLoS ONE. 2014;9(1) doi: 10.1371/journal.pone.0086129.e86129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.de Waal EJ, Rademakers LH. Heterogeneity of epithelial cells in the rat thymus. Microscopy Research and Technique. 1997;38(3):227–236. doi: 10.1002/(SICI)1097-0029(19970801)38:3<227::AID-JEMT4>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 26.Schuurman HJ, Kuper CF, Kendall MD. Thymic microenvironment at the light microscopic level. Microscopy Research and Technique. 1997;38(3):216–226. doi: 10.1002/(SICI)1097-0029(19970801)38:3<216::AID-JEMT3>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 27.von Gaudecker B, Kendall MD, Ritter MA. Immuno-electron microscopy of the thymic epithelial microenvironment. Microscopy Research and Technique. 1997;38(3):237–249. doi: 10.1002/(SICI)1097-0029(19970801)38:3<237::AID-JEMT5>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 28.Lee EN, Park JK, Lee JR, et al. Characterization of the expression of cytokeratins 5, 8, and 14 in mouse thymic epithelial cells during thymus regeneration following acute thymic involution. Anatomy & Cell Biology. 2011;44(1):14–24. doi: 10.5115/acb.2011.44.1.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Savino W, Dardenne M. Immunohistochemical studies on a human thymic epithelial cell subset defined by the anti-cytokeratin 18 monoclonal antibody. Cell and Tissue Research. 1988;254(1):225–231. doi: 10.1007/BF00220038. [DOI] [PubMed] [Google Scholar]
- 30.Klug DB, Carter C, Gimenez-Conti IB, Richie ER. Cutting edge: thymocyte-independent and thymocyte-dependent phases of epithelial patterning in the fetal thymus. The Journal of Immunology. 2002;169(6):2842–2845. doi: 10.4049/jimmunol.169.6.2842. [DOI] [PubMed] [Google Scholar]
- 31.White AJ, Nakamura K, Jenkinson WE, et al. Lymphotoxin signals from positively selected thymocytes regulate the terminal differentiation of medullary thymic epithelial cells. The Journal of Immunology. 2010;185(8):4769–4776. doi: 10.4049/jimmunol.1002151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Forssmann WG, Ito S, Weihe E, Aoki A, Dym M, Fawcett DW. An improved perfusion fixation method for the testis. Anatomical Record. 1977;188(3):307–314. doi: 10.1002/ar.1091880304. [DOI] [PubMed] [Google Scholar]
- 33.Rassadi S, Krishnaswamy A, Pié B, McConnell R, Jacob MH, Cooper E. A null mutation for the α3 nicotinic acetylcholine (ACh) receptor gene abolishes fast synaptic activity in sympathetic ganglia and reveals that ACh output from developing preeanglionic terminals is regulated in an activity-dependent retrograde manner. Journal of Neuroscience. 2005;25(37):8555–8566. doi: 10.1523/JNEUROSCI.1983-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shezen E, Okon E, Ben-Hur H, Abramsky O. Cytokeratin expression in human thymus: immunohistochemical mapping. Cell and Tissue Research. 1995;279(1):221–231. doi: 10.1007/BF00300707. [DOI] [PubMed] [Google Scholar]
- 35.Moll R, Franke WW, Schiller DL, Geiger B, Krepler R. The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells. Cell. 1982;31(1):11–24. doi: 10.1016/0092-8674(82)90400-7. [DOI] [PubMed] [Google Scholar]
- 36.Fuchs E, Green H. Changes in keratin gene expression during terminal differentiation of the keratinocyte. Cell. 1980;19(4):1033–1042. doi: 10.1016/0092-8674(80)90094-x. [DOI] [PubMed] [Google Scholar]
- 37.Ginda WJ, Gratzl M, Mayerhofer A, Warchol JB. Chromogranin A immunoreactivity in epithelial cells of the thymus. Folia Histochemica et Cytobiologica. 1996;34(2):91–93. [PubMed] [Google Scholar]
- 38.Raica M, Encicǎ S, Motoc A, Cîmpean AM, Scridon T, Bârsan M. Structural heterogeneity and immunohistochemical profile of Hassall corpuscles in normal human thymus. Annals of Anatomy. 2006;188(4):345–352. doi: 10.1016/j.aanat.2006.01.012. [DOI] [PubMed] [Google Scholar]
- 39.Kurzen H, Berger H, Jäger C, et al. Phenotypical and molecular profiling of the extraneuronal cholinergic system of the skin. Journal of Investigative Dermatology. 2004;123(5):937–949. doi: 10.1111/j.0022-202X.2004.23425.x. [DOI] [PubMed] [Google Scholar]
- 40.Kindt F, Wiegand S, Niemeier V, et al. Reduced expression of nicotinic α subunits 3, 7, 9 and 10 in lesional and nonlesional atopic dermatitis skin but enhanced expression of α subunits 3 and 5 in mast cells. British Journal of Dermatology. 2008;159(4):847–857. doi: 10.1111/j.1365-2133.2008.08774.x. [DOI] [PubMed] [Google Scholar]
- 41.Grando SA, Horton RM, Mauro TM, Kist DA, Lee TX, Dahl MV. Activation of keratinocyte nicotinic cholinergic receptors stimulates calcium influx and enhances cell differentiation. Journal of Investigative Dermatology. 1996;107(3):412–418. doi: 10.1111/1523-1747.ep12363399. [DOI] [PubMed] [Google Scholar]
- 42.Arredondo J, Nguyen VT, Chernyavsky AI, et al. Central role of α7 nicotinic receptor in differentiation of the stratified squamous epithelium. Journal of Cell Biology. 2002;159(2):325–336. doi: 10.1083/jcb.200206096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chernyavsky AI, Arredondo J, Vetter DE, Grando SA. Central role of α9 acetylcholine receptor in coordinating keratinocyte adhesion and motility at the initiation of epithelialization. Experimental Cell Research. 2007;313(16):3542–3555. doi: 10.1016/j.yexcr.2007.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Santos M, Río P, Ruiz S, et al. Altered T cell differentiation and notch signaling induced by the ectopic expression of keratin K10 in the epithelial cells of the thymus. Journal of Cellular Biochemistry. 2005;95(3):543–558. doi: 10.1002/jcb.20406. [DOI] [PubMed] [Google Scholar]
- 45.Watanabe N, Wang YH, Lee HK, Ito T, Cao W, Liu Y. Hassall's corpuscles instruct dendritic cells to induce CD4+CD25+ regulatory T cells in human thymus. Nature. 2005;436(7054):1181–1185. doi: 10.1038/nature03886. [DOI] [PubMed] [Google Scholar]
- 46.Martins VC, Boehm T, Bleul CC. Ltβr signaling does not regulate aire-dependent transcripts in medullary thymic epithelial cells. The Journal of Immunology. 2008;181(1):400–407. doi: 10.4049/jimmunol.181.1.400. [DOI] [PubMed] [Google Scholar]
- 47.Radek KA, Lopez-Garcia B, Hupe M, et al. The neuroendocrine peptide catestatin is a cutaneous antimicrobial and induced in the skin after injury. Journal of Investigative Dermatology. 2008;128(6):1525–1534. doi: 10.1038/sj.jid.5701225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rabbi MF, Labis B, Metz-Boutigue MH, Bernstein CN, Ghia JE. Catestatin decreases macrophage function in two mouse models of experimental colitis. Biochemical Pharmacology. 2014;89(3):386–398. doi: 10.1016/j.bcp.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 49.Sahu BS, Mohan J, Sahu G, et al. Molecular interactions of the physiological anti-hypertensive peptide catestatin with the neuronal nicotinic acetylcholine receptor. Journal of Cell Science. 2012;125(9):2323–2337. doi: 10.1242/jcs.103176. [DOI] [PubMed] [Google Scholar]