

Abstract
Expression of the IL-7 receptor α-chain (CD127) is decreased on CD8 T-cells in HIV infected patients and partially recovers in those receiving antiretroviral therapy with sustained viral suppression. We have shown that soluble HIV Tat protein down regulates CD127 expression on CD8 T-cells isolated from healthy HIV-negative individuals. Tat is taken up by CD8 T-cells via endocytosis, exits the endosome and then translocates to the inner leaflet of the cell membrane where it binds to the cytoplasmic tail of CD127 inducing receptor internalization and degradation by the proteasome. This down regulation of CD127 by Tat results in impaired CD8 T-cell function. Interestingly, suppression of CD127 by Tat is reversible and requires the continual presence of Tat in the culture media. We thus questioned whether the low IL-7 receptor expression evident on CD8 T-cells in HIV+ patients was similarly reversible and if suppression of the receptor could be maintained ex vivo by Tat protein alone. We show here that when CD8 T-cells isolated from HIV+ patients are incubated alone in fresh medium, low CD127 expression on the cell surface recovers to normal levels. This recovery of CD127, however, is completely inhibited by the addition of HIV Tat protein to the culture media. This study then provides evidence that soluble factor(s) are responsible for low CD127 expression on circulating CD8 T-cells in HIV+ individuals and further implicates Tat in suppressing this receptor essential to CD8 T-cell proliferation and function.
Introduction
Impaired cell mediated immunity is the clinical hallmark of HIV infection and is directly responsible for the appearance of many opportunistic infections in patients with progressive disease. In vitro studies have confirmed functional deficits in CD8 T-cells isolated from HIV+ individuals including reduced proliferation and impaired cytolytic activity. [1], [2], [3], [4] Indeed, both HIV− and EBV-specific CD8 T-cells can be found in the circulation at relatively normal frequencies in HIV-infected patients with advanced disease [5], [6], [7], [8], [9] yet these cells respond poorly to their cognate antigens and fail to express normal levels of perforin and interferon (IFN)-γ, or demonstrate effective cytolytic activity. [6], [9], [10], [11], [12], [13] This is of obvious advantage to HIV as by disarming cell mediated immunity the virus is able to avoid elimination and establish chronic infection.
Interleukin (IL)-7 is essential for normal T-cell development and function. In addition to playing a critical role in peripheral immune homeostasis [9], [14], [15], [16], [17], [18] and the development and maintenance of T-cell memory, [19], [20] IL-7 also plays an important role in the activation of CD8 T-cells in response to foreign antigen. IL-7 independently stimulates CD8 T-cell proliferation, [21], [22], [23], [24] and potentiates cytolytic activity [25], [26], [27], [28], [29], [30], [31] by enhancing production of IFN-γ following TCR stimulation [32], [33] and by inducing accumulation of intracellular perforin. [34], [35] Given the important role IL-7 plays in CD8 T-cell responses, decreased IL-7 signaling would be expected to result in impaired cell mediated immunity and inefficient control of viral pathogens including HIV.
IL-7 signaling occurs via its receptor, a heterodimer composed of a unique α-chain (CD127) [36] and the common γ-chain (CD132). [37] We and others have shown decreased expression of the IL-7R α-chain on CD8 T-cells in HIV-infected individuals with uncontrolled viral replication [38], [39], [40], [41], [42], [43], [44] and partial recovery in patients receiving highly active antiretroviral therapy with sustained viral suppression. [38], [41] Notably, the decrease in CD127 expression in HIV+ individuals correlates with impaired CD8 T-cell responses. Vingerhoets et al [45] found compared to controls CD8 T-cells from HIV+ individuals with low CD127 expression were less able to form blasts and up regulate CD25 in response to IL-7. Ferrari et al [46] also found anti-HIV CD8 T-cells isolated from patients with advanced disease could not be expanded in vitro following stimulation with HIV antigens and IL-7. Thus it appears decreased CD127 expression leads to impaired CD8 T-cell proliferation and function and thus may contribute to reduced cell mediated immunity in HIV+ patients.
The factors responsible for down regulating CD127 during HIV infection have yet to be definitively established. Notably, decreased CD127 expression has been observed on all CD8 T-cell subsets in HIV+ individuals including resting naïve cells with concomitant low CD38 expression suggesting suppression of this receptor may not be the result of chronic T-cell activation. [38], [39], [42], [47] Several soluble factors likely play a role and we have previously shown soluble HIV Tat protein specifically down regulates CD127 on the surface of CD8 T-cells isolated from healthy HIV-negative volunteers. [35], [48] Tat, a small 15 kdal viral polypeptide, is secreted by infected CD4+ cells [49], [50], [51], [52], [53], [54], [55] and is rapidly internalized by neighboring uninfected lymphocytes [51], [56], [57] through clathrin-coated pits. [58] Once inside the cell, Tat exits late endosomes upon the usual acidification of these vesicles [58], [59] and translocates to the inner leaflet of the plasma membrane where it binds to the cytoplasmic tail of CD127. [48] This interaction with Tat induces receptor aggregation and removal from the cell surface through a process dependent on microtubules and directs CD127 to the proteasome for degradation. [48] The effect of Tat on CD127 expression is both dose and time dependent, can be blocked with anti-Tat antibodies, [35] and occurs in the presence of cycloheximide indicating a direct effect and that new protein synthesis is not required [48]. Tat down regulates CD127 equally on both naïve and memory CD8 T-cells yet has no effect on a number of other cell surface proteins including CD25, CD28 and CD56 indicating a stable CD8 T-cell phenotype and lack of nonspecific activation in the presence of soluble Tat protein. Tat also has no effect on the expression of CD132, the common γ-chain that associates with CD127 to form the IL-7 receptor. Importantly, Tat-induced down regulation of CD127 results in deficits in CD8 T-cell activity. Pre-incubating CD8 T-cells with Tat inhibits proliferation and accumulation of intra-cellular perforin following stimulation with IL-7. [35] Thus, by down regulating CD127 expression on CD8 T-cells, soluble HIV Tat protein is able to decrease IL-7 signaling and impair both CD8 T-cell expansion and cytolytic capacity. Interestingly, Tat-induced down regulation of CD127 on the surface of CD8 T-cells is reversible and requires the continual presence of this viral protein. When Tat is removed from the culture media, CD127 returns to normal levels with 24 hours. [35].
In view of the fact that soluble Tat protein is able to down regulate CD127 on CD8 T-cells and that this down regulation is reversible after up to 72 hours in culture upon removal of Tat from the media, we asked whether CD127 expression could recover on CD8 T-cells isolated from HIV+ patients and if so if this recovery is inhibited by Tat.
Materials and Methods
Ethics Statement
This work was reviewed and approved by the Ottawa Hospital Research Ethics Board and informed written consent was obtained from all participants. No children were used in this study. Written consent was obtained from all participants after reading a 5 page document outlining the phlebotomy procedure, study outline and research goals.
Patients
Patients were recruited from The Ottawa Hospital Immunodeficiency Clinic according to the following inclusion criteria: age over 18 years; naïve to antiretroviral therapy or off therapy for at least three months; a blood plasma HIV viral load above detection (>50 copies/ml); CD4 T-cell count >200 cells/µl; no co-morbidities; and able to provide signed informed consent. Exclusion criteria included co-infection with hepatitis B and/or hepatitis C virus; and clinical or laboratory evidence of an active infection or malignancy. CD4 and CD8 T-cell counts, percentages and ratios as well as HIV viral loads were obtained from the patients’ medical records.
Reagents
Purified HIV-1 Tat protein (86 amino acids) was purchased from Advanced Bioscience Laboratories Inc. (Kensington, MD). Lyophilized protein was resuspended to 1 mg/ml in phosphate buffered saline (PBS) containing 1 mg/ml bovine serum albumin (BSA) and 0.1 mM dithiothreitol. Tat protein is reportedly >95% pure by heparin-affinity chromatography and reverse phase high performance liquid chromatography. Anti-CD8-phycoerythrin-Cy5 (PC5) (B9.11) and anti-CD127-phycoerythrin (PE) (R34.34) fluorochrome-labeled monoclonal antibodies were purchased from Immunotech Beckman Coulter (Marseille, France). All fluorochrome labeled antibodies were titrated and used at saturating concentrations.
Cell Purification and Culture
Patient blood was drawn into sodium heparin-containing tubes and processed within 2 hours. Peripheral blood mononuclear cells (PBMC) were isolated by Ficoll-Paque density centrifugation followed by CD8 T-cell purification using the MACS Microbead CD8+ Cell AutoMACS Isolation System (Miltenyi Biotec, Auburn, CA). By this method, cell purity was consistently >95% CD8+. Purified CD8 T-cells were incubated at 1×106 cells/ml in media comprised of RPMI 1640 (Hyclone, Logan, UT) supplemented with 20% fetal calf serum (FCS; Cansera, Rexdale, ON, Canada) plus 100 U/ml penicillin, 100 µg/ml streptomycin and 0.2 M L-glutamine (RPMI-20). All cultures were maintained in a humidified incubator at 37°C in the presence of 5% CO2.
Flow cytometry
At times indicated, cells were incubated with anti-CD8-PC5 and anti-CD127-PE fluorochrome-labeled antibodies for 30 minutes in the dark at room temperature, and then analyzed by flow cytometry using a Coulter Epics ALTRA flow cytometer (Fullerton, CA). Live cells were gated on the basis of side and forward scatter and at least 10,000 events were recorded for each sample. Isotype controls were performed for each fluorochrome-conjugated antibody. Resulting profiles were analyzed with De Novo FCS Express 2 software (Los Angeles, CA). As demonstrated previously, decreased detection of CD127 by flow cytometry using the R34.34 antibody correlates with reduced CD127 protein as determined by Western blot analysis [48], [60].
Statistical Analysis
Statistical comparisons were carried out using a two-tailed, paired student t-test with 95% confidence intervals and p values given throughout the text reflect this analysis. Given the sample sizes for the two patient groups, parallel nonparametric analyses were also carried out using the Wilcoxon matched pair test again with 95% confidence intervals. Statistical significance as defined by a p value greater or less than 0.05 remained the same for all comparisons irrespective of parametric or nonparametric analysis.
Results
Patients
Patient characteristics and demographics (N = 11) are shown in Table 1. The mean age was 39 years (range 21–56) and all were men. The mean CD4 T-cell count was 355 cells/µl or 23% (range 203–573; 13–41%) with a mean CD8 T-cell count of 1043 cells/µl or 60% (range 246–2411; 37–73%). The average CD4/CD8 T-cell ratio was 0.41. HIV viral loads ranged from 484 to 500,000 copies/ml with a mean of 94,023 copies/ml. Nine patients had never received antiretroviral therapy and 2 had been previously treated but had discontinued therapy five months prior to enrolment. The patients were otherwise in good health at the time of analysis.
Table 1. Study patients.
| Participant | Age | Sex | CD3CD4 count (%) | CD3CD8 count (%) | CD4/CD8 ratio | Viral load (copies/ml) | Prior Rx |
| 1 | 42 | M | 203 (16) | 817 (64) | 0.25 | 13,510 | No |
| 2 | 21 | M | 247 (17) | 995 (69) | 0.25 | 7,953 | No |
| 3 | 39 | M | 271 (18) | 967 (65) | 0.28 | 105,425 | Yes |
| 4 | 30 | M | 474 (28) | 932 (54) | 0.51 | 22,051 | No |
| 5 | 44 | M | 344 (22) | 923 (59) | 0.37 | 500,000 | No |
| 6 | 56 | M | 390 (24) | 868 (53) | 0.45 | 75,216 | No |
| 7 | 24 | M | 272 (41) | 246 (37) | 1.1 | 61,517 | No |
| 8 | 52 | M | 359 (21) | 1091 (64) | 0.3 | 484 | No |
| 9 | 43 | M | 530 (33) | 850 (53) | 0.6 | 27,158 | No |
| 10 | 43 | M | 573 (16) | 2411 (66) | 0.2 | 109,771 | Yes |
| 11 | 41 | M | 248 (13) | 1376 (73) | 0.2 | 111,174 | No |
| Average | 39 | 355 (23) | 1043 (60) | 0.41 | 94,023 |
Patients were analyzed in two separate groups, patients 1–7 (assayed at isolation and after 24 hours) and patients 6–11 (measured at isolation and every 24 hours over 72 hours). The groups were not statistically different in any characteristic analyzed including age, T-cell counts/percentages, or HIV viral loads (all p values ≥0.24).
CD127 recovers ex vivo on CD8 T-cells isolated from HIV+ individuals
CD127 is down regulated on CD8 T-cells in HIV+ patients and partially recovers following viral suppression with highly active antiretroviral therapy. [38], [39], [40], [41], [42], [43] We have previously shown soluble HIV Tat protein down regulates CD127 on healthy CD8 T-cells in culture [35] and that this down regulation is reversible with CD127 recovering back to normal levels once Tat is removed from the culture media. This prompted us to ask whether the suppression of CD127 on CD8 T-cells in HIV+ individuals is reversible and whether these cells could re-express CD127 when maintained in culture media ex vivo. To address this, CD8 T-cells were isolated from HIV-infected patients not on therapy and a portion of the cells were analyzed immediately for CD127 expression by flow cytometry. The remainder of the cells were washed in PBS and incubated for up to 72 hours in RPMI-20, and CD127 expression was monitored by flow cytometry at 24 hour intervals. Figure 1 shows typical flow histograms demonstrating CD127 expression on CD8 T-cells from an HIV-negative individual (panel A) and from an HIV+ individual (panel B) immediately following CD8 T-cell isolation (grey fill) and after 24 hours in culture media (black line). Consistent with previous reports, CD127 expression was significantly decreased on CD8 T-cells from the HIV+ individual immediately following isolation. However, when purified CD8 T-cells from this same individual were maintained ex vivo in RPMI-20, CD127 recovered over 24 hours to levels equivalent to that seen in the HIV-negative control. This was demonstrated in both percent positive staining cells and in mean channel fluorescence. In comparison, CD127 expression increased only marginally on CD8 T-cells isolated from the HIV-negative individual. Figure 2 shows similar data as both histograms for three representative HIV+ patients (panel A) and in composite form for seven patients (patients 1–7; panel D). In all cases, CD127 expression increased within 24 hours on CD8 T-cells isolated from HIV+ individuals when cultured in media alone (figure 2 panel A; grey fill, immediately following isolation; black line, after 24 hours in culture media). Indeed, the average mean channel fluorescence for CD127 staining on CD8 T-cells increased from 3.45+/−0.19 (mean +/− standard error of the mean) at the time of isolation to 5.59+/−0.61 after 24 hours in culture media (p = 0.005). We therefore conclude that suppression of CD127 on CD8 T-cells in HIV+ individuals is reversible and increases over 24 hours when the cells are maintained in fresh medium ex vivo. This suggests a soluble factor or factors are responsible for the down regulation of CD127 in vivo.
Figure 1. CD127 recovers ex vivo to normal levels on CD8 T-cells isolated from an HIV-infected individual.
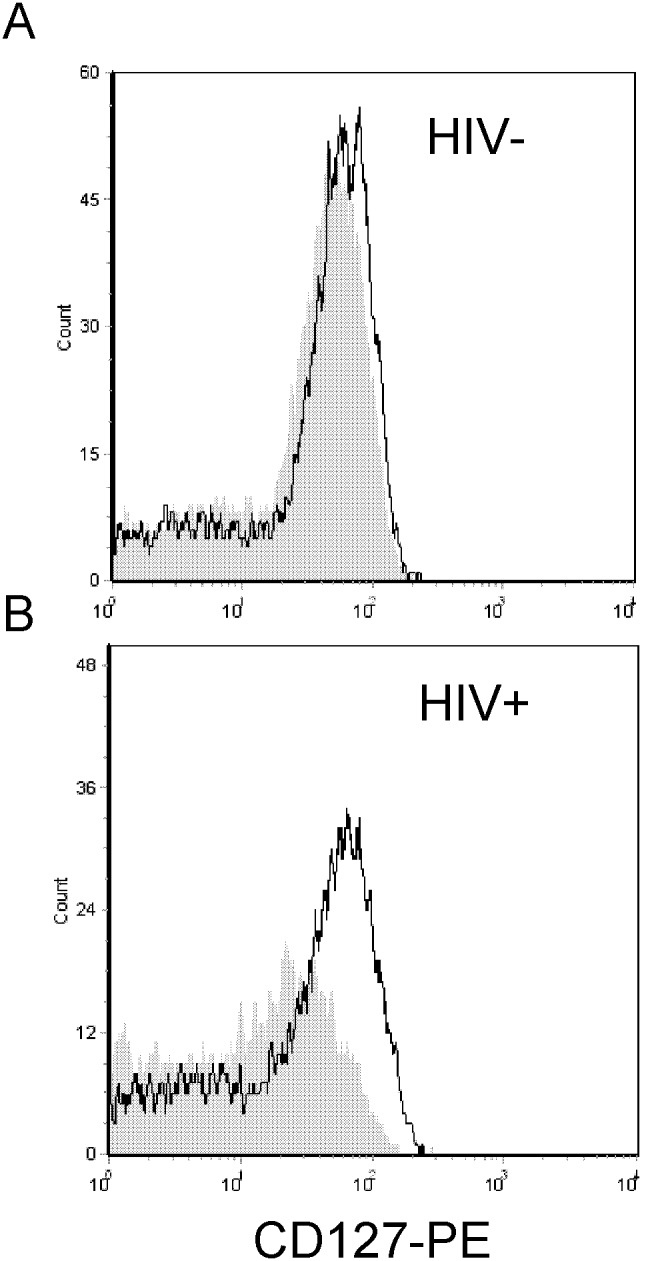
CD127 expression on purified CD8 T-cells is shown immediately following cell isolation (gray fill) and after 24 hours in culture medium (black line). Two representative histograms are shown, one from a healthy HIV-negative volunteer (panel A) and one from an HIV-infected patient with a CD4 count of 359 cells/µl and an HIV viral load (VL) of 484 copies/ml (panel B).
Figure 2. While CD127 expression recovers ex vivo on CD8 T-cells isolated from HIV+ patients, soluble HIV Tat protein maintains suppression of the receptor.

CD8 T-cells isolated from HIV-positive patients were cultured in media alone (panel A) or in media containing purified Tat protein at 2 µg/ml (panel B) or 10 µg/ml (panel C) for 24 hours. Three representative HIV-positive patients are shown (Patient 5: CD4 = 344 cells/µl and VL = 500,000 copies/ml; Patient 6: CD4 = 390 cells/µl and VL = 75,216 copies/ml; Patient 7: CD4 = 272 cells/µl and VL = 61,517 copies/ml). CD127 expression on purified CD8 T-cells is shown immediately following cell isolation (gray fill) and after 24 hours in culture (black line). Panel D: Composite data from 7 patients showing changes in CD127 mean channel fluorescence. CD8 T-cells isolated from HIV-positive patients were cultured in media alone, or in media containing purified Tat protein at 2 µg/ml or 10 µg/ml for 24 hours. CD127 mean channel fluorescence increased on cells maintained in media alone for 24 hours compared to the time of isolation (*p = 0.005), remained unchanged in the presence of 2 µg/ml soluble Tat protein (ns), and decreased slightly when cultured in the presence of 10 µg/ml Tat (**p = 0.007).
HIV Tat protein maintains suppression of CD127 on CD8 T-cells isolated from HIV+ patients
In view of our previous findings demonstrating soluble HIV Tat protein specifically down regulates CD127 on the surface of CD8 T-cells isolated from healthy volunteers, we questioned whether this viral protein alone could maintain suppression of the IL-7 receptor on CD8 T-cells isolated from HIV+ individuals. To address this, CD8 T-cells were purified from HIV+ patients as described and incubated in either RPMI-20 alone or in media containing purified Tat protein. Figure 2 shows flow histograms from three representative patients (panels B and C) and composite data from patients 1–7 (panel D). In contrast to cells incubated in medium alone, when the cells were cultured in the presence of soluble Tat protein, suppression of CD127 was maintained. Indeed, in the presence of 2 µg/ml Tat the expression of CD127 on CD8 T-cells remained stable (figure 2, panel B) with an average mean channel fluorescence of 3.20+/−0.21, essentially unchanged compared to the levels of CD127 at the time of isolation (p = 0.39; panel D). Interestingly, higher concentrations of Tat protein (10 µg/ml) were able to further suppress CD127 expression albeit slightly after 24 hours in culture (figure 2, panel C). Under these conditions, the average CD127 mean channel fluorescence decreased to 2.63+/−0.19 (p = 0.007 compared to time of isolation; panel D).
To more fully describe the dynamics of CD127 recovery and its suppression by Tat, CD8 T-cells isolated from six HIV+ individuals (participants 6–11) were followed over 72 hours. Figure 3 shows flow histograms from one representative patient (Patient 7) while composite data for all six patients are shown in panel D. While time to peak recovery varied somewhat from patient to patient, CD127 expression increased on purified CD8 T-cells isolated from HIV+ individuals when cultured in media alone and was maintained at levels comparable to HIV-negative controls for over 72 hours (figure 3, panel A). Consistent with the data above, the average CD127 mean channel fluorescence increased from 3.52+/−0.19 at the time of isolation to 6.35+/−0.54 after 24 hours in culture media (p = 0.0003), and then remained stable (panel D). When CD8 T-cells were cultured in media containing purified Tat protein, suppression of CD127 was fully maintained for up to at least 72 hours (figure 3, panels B, C and D). In the presence of 10 µg/ml Tat, CD127 mean channel fluorescence remained less than 3.2 over the 72 hours and well below recovered CD127 levels on cells maintained in parallel in medium alone (p≤0.003).
Figure 3. Soluble HIV Tat protein maintains suppression of CD127 expression on CD8 T-cells isolated from HIV-infected individuals over at least 72 hours.

Representative histograms show CD127 expression on CD8 T-cells isolated from an HIV-infected patient (Patient 7: CD4 = 272 cells/µl and VL = 61,517 copies/ml) and incubated in media alone (panel A) or in media containing 2 µg/ml (panel B) or 10 µg/ml (panel C) soluble Tat protein. CD127 expression on purified CD8 T-cells is shown immediately following cell isolation (gray fill) and after 24, 48 and 72 hours in culture (black lines). Panel D: Composite data from 6 patients showing changes in CD127 mean channel fluorescence. CD8 T-cells were isolated from HIV-infected patients and incubated in media alone, or in media containing 10 µg/ml Tat protein for 72 hours. For cells cultured in media alone, CD127 mean channel fluorescence increased within 24 hours and was maintained thereafter (p≤0.01 at 24, 48 and 72 hours compared to time of isolation). CD127 expression was not statistically different comparing cells cultured in media for 48 and 72 hours compared to 24 hours (p = 0.70 and 0.98 respectively). When cells were cultured in the presence of Tat protein, CD127 expression remained low with mean channel fluorescence readings significantly lower compared to cells cultured in media alone. (*indicates p≤0.003.).
Discussion
This work addresses two questions. First, we asked whether the low CD127 expression on CD8 T-cells in HIV+ patients is reversible. Several groups including our own have reported an increase in CD127 on the surface of CD8 T-cells in HIV+ patients following viral suppression with effective antiretroviral therapy. [38], [39], [40], [41], [42], [43] However, it is not clear whether this recovery is due to an increase in CD127 on existing T-cells or reflects a repopulation of new cells with higher CD127 expression during immune reconstitution. Although a contribution from the latter cannot be ruled out, we show here that suppression of CD127 on CD8 T-cells from HIV+ individuals is in fact reversible and that CD127 receptor density increases on existing cells ex vivo. The fact that CD8 T-cells from HIV+ patients are able to re-express CD127 once purified and placed in culture medium indicates reduced expression of the IL-7 receptor in vivo is the result of active suppression by soluble factors in the patient serum and not due to permanent alterations in the cells themselves. Similar recovery of CD127 on CD8 T-cells in mixed PBMC cultures from HIV+ patients has also been reported by Colle et al. [41] In their study, CD127 expression increased significantly after 3 days in culture on all CD8 T-cell subsets and most notably on naïve, central memory and effector memory cells. Rethi et al. [43] failed to detect CD127 recovery ex vivo but in their study they reported only a stable percentage of low/negative T-cells and therefore may have missed an increase in receptor expression since they did not measure changes in mean channel fluorescence on CD127+ cells.
We have previously shown soluble HIV Tat protein specifically down regulates CD127 on the surface of CD8 T-cells isolated from healthy HIV-negative volunteers. [35] Tat is secreted by HIV-infected cells [49], [51], [54] and can be detected in the media during in vitro infection [51], [52] as well as in the tissues and serum of HIV-infected patients. [53], [61] As noted in our previous study, Tat-induced suppression of CD127 is maintained only if Tat is continuously present in the media. When Tat is removed from the media, CD127 recovers on the cell surface to pre-treatment levels within 24 hours. [35] In view of this and the fact that soluble factors appear to actively suppress CD127 expression in vivo, we next asked whether soluble Tat protein alone could maintain suppression of CD127 ex vivo on CD8 T-cells isolated from HIV+ patients. We show here this is in fact the case. In all patients examined, CD127 uniformly remained low on CD8 T-cells isolated from HIV+ individuals when cultured in the presence of soluble Tat protein and essentially unchanged compared to the time of isolation. These data suggest soluble Tat protein may well play an active role in suppressing IL-7 receptor expression on CD8 T-cells in vivo.
Interestingly, lower concentrations of Tat are required to maintain suppression of CD127 on CD8 T-cells isolated from HIV+ individuals compared to concentrations needed to down regulate this receptor de novo on CD8 T-cells isolated from healthy HIV-negative volunteers. In a previous study we found 2 µg/ml of Tat induce a 4+/−1% decrease in CD127 expression on healthy CD8 T-cells while 10 µg/ml induced a 39+/−3% decline compared to cells maintained in media alone. [62] In the present study, Tat at 2 µg/ml was able to maintain full suppression of CD127 ex vivo on cells isolated from HIV+ patients. That lower concentrations are required to maintain receptor suppression is not surprising given the effects of Tat on CD127 are stoichiometric. Once taken up by CD8 T-cells, Tat interacts directly with the cytoplasmic tail of CD127 to induce receptor internalization and degradation. [48] The direct protein-protein interaction between Tat and CD127 necessary for receptor down regulation mandates higher Tat concentrations in healthy CD8 T-cells where the density of CD127 on the cell surface is much higher, whereas lower Tat concentrations could be sufficient to prevent re-accumulation of already low levels of CD127 on the surface of cells isolated from HIV+ patients.
Tat concentrations in the sera of HIV+ patients have been reported ranging 40–550 ng/ml, [53] some 4 to 250-fold lower than the concentrations used in our experiments. However, direct comparisons of in vivo protein concentrations to those used in in vitro assays should be approached with caution. For example, the Tat protein used in our assays is recombinant protein purified from E. coli. As such, recombinant proteins do not undergo post-translational modification and it is unlikely 100% of the purified protein retains biological activity. Thus in vivo concentrations of post-translationally modified Tat secreted from neighbouring HIV-infected CD4 cells and in vitro concentrations of recombinant Tat purified in the laboratory from E. coli may not be comparable. Perhaps more importantly the concentration of free Tat protein circulating in the sera of HIV infected patients likely does not reflect the levels of soluble Tat protein that cells are exposed to in vivo. Indeed, Tat concentrations in the peripheral circulation are likely quite low compared to concentrations in the lymph nodes and gut mucosa where the majority of HIV-infected T-cells reside.
Accumulating evidence indicates soluble Tat protein plays a significant role in the immune dysregulation evident in progressive HIV infection We have shown down regulation of CD127 by Tat results in impaired CD8 T-cell function including reduced proliferation and decreased accumulation of intracellular perforin following stimulation with IL-7. [35] Others have also shown lymphocytes exposed to extracellular Tat in vitro no longer proliferate to tetanus toxoid, [63], [64] Staphylococcal enterotoxin B, [65] or anti-CD3 monoclonal antibodies. [35], [66] In addition, neutralizing anti-Tat antibodies in the serum of HIV-infected individuals have been correlated with low viral loads and slower disease progression. [64], [67], [68], [69] [70] In view of this, Tat is currently being developed as both a prophylactic and therapeutic HIV vaccine in both animal models and in humans. [71], [72], [73], [74], [75], [76], [77] Ensoli and colleagues recently showed therapeutic immunization of HIV+ patients on antiretroviral therapy with Tat protein resulted in a durable anti-Tat humoral response. What is most striking is that individuals who developed anti-Tat antibodies demonstrated a normalization of T-cell phenotypes as well as increased CD4 and CD8 T-cell viability, proliferation and response to antigens. [75] Although CD127 was not analyzed in their study, the immune restoration evident in vaccinated individuals with anti-Tat antibodies could be explained at least in part by the recovery of IL-7 signaling. Anti-Tat antibodies generated through vaccination could neutralize soluble Tat protein in the serum and lymphoid tissues of patients and thereby allow recovery of CD127 on the surface of CD4 and CD8 T-cells. Indeed, we have also previously shown anti-Tat antibodies block Tat’s ability to down regulate CD127 on CD8 T-cells isolated from healthy individuals. [35] IL-7′s established roles in increasing cell viability through up regulation of Bcl-2, [78], [79] regulating peripheral T-cell number, [14], [15], [80], [81], [82] establishing T-cell memory, [18], [19], [20] and enhancing CD8 T-cell effector function [25], [26], [27], [31], [83], [84] could all explain the increase in cell viability, increase in CD4 T-cell number, increased percentage of central memory CD4 and CD8 T-cells as well as the increased expression of activation markers on CD8 T-cells and improved CD4 and CD8 T-cell responses to recall antigens demonstrated in Tat-vaccinated individuals. [75].
We show here that CD8 T-cells isolated from HIV+ individuals can re-express CD127 when purified and cultured ex vivo. This suggests existing CD8 T-cells present in HIV+ patients may not be irreversibly impaired and that a soluble factor or factors maintains suppression of CD127 in vivo. While a number of factors may be responsible for down regulating CD127 in HIV+ individuals, we demonstrate here that soluble Tat protein alone can maintain suppression of CD127 on the surface of CD8 T-cells isolated from HIV+ patients. This provides further evidence that Tat likely plays a major role in the suppression of CD127 in vivo. This in conjunction with our previous data showing Tat induced down regulation of CD127 results in impaired CD8 T-cell proliferation and cytolytic potential [35] and the apparent immune restoration documented by Ensoli and colleagues in patients undergoing therapeutic Tat vaccination [75] lends further support to the idea that soluble Tat protein likely contributes to poor lymphocyte responses and overall immune dysregulation in HIV+ patients.
Funding Statement
Elliott Faller was supported by a Doctoral research grant (200510lDB-162377-159103) from CIHR. P. MacPherson was supported by a New Investigator Award (200509CNI-161294-HPA-ADYP-39720) from CIHR (http://www.cihr-irsc.gc.ca). This Project was performed with grant funding from OHTN, grant number ROGA240 (www.ohtn.on.ca). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Sharma B, Gupta S (1985) Antigen-specific primary cytotoxic T lymphocyte (CTL) responses in acquired immune deficiency syndrome (AIDS) and AIDS-related complexes (ARC). Clin Exp Immunol 62: 296–303. [PMC free article] [PubMed] [Google Scholar]
- 2. Bettens F, Pichler CE, Herrmann B, de Weck AL, Pichler WJ (1991) Selective stimulation of CD4+ versus CD8+ T-cell subsets in symptomatic and asymptomatic HIV-1-infected individuals. AIDS Res Hum Retroviruses 7: 773–780. [DOI] [PubMed] [Google Scholar]
- 3. Gerstoft J, Dickmeiss E, Mathiesen L (1985) Cytotoxic capabilities of lymphocytes from patients with the acquired immunodeficiency syndrome. Scand J Immunol 22: 463–470. [DOI] [PubMed] [Google Scholar]
- 4. Miedema F, Petit AJ, Terpstra FG, Schattenkerk JK, de Wolf F, et al. (1988) Immunological abnormalities in human immunodeficiency virus (HIV)-infected asymptomatic homosexual men. HIV affects the immune system before CD4+ T helper cell depletion occurs. J Clin Invest 82: 1908–1914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Gea-Banacloche JC, Migueles SA, Martino L, Shupert WL, McNeil AC, et al. (2000) Maintenance of large numbers of virus-specific CD8+ T cells in HIV-infected progressors and long-term nonprogressors. J Immunol 165: 1082–1092. [DOI] [PubMed] [Google Scholar]
- 6. Migueles SA, Laborico AC, Shupert WL, Sabbaghian MS, Rabin R, et al. (2002) HIV-specific CD8+ T cell proliferation is coupled to perforin expression and is maintained in nonprogressors. Nat Immunol 3: 1061–1068. [DOI] [PubMed] [Google Scholar]
- 7. Migueles SA, Connors M (2001) Frequency and function of HIV-specific CD8(+) T cells. Immunol Lett 79: 141–150. [DOI] [PubMed] [Google Scholar]
- 8. Spiegel HM, Ogg GS, DeFalcon E, Sheehy ME, Monard S, et al. (2000) Human immunodeficiency virus type 1- and cytomegalovirus-specific cytotoxic T lymphocytes can persist at high frequency for prolonged periods in the absence of circulating peripheral CD4(+) T cells. J Virol 74: 1018–1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. van Baarle D, Hovenkamp E, Callan MF, Wolthers KC, Kostense S, et al. (2001) Dysfunctional Epstein-Barr virus (EBV)-specific CD8(+) T lymphocytes and increased EBV load in HIV-1 infected individuals progressing to AIDS-related non-Hodgkin lymphoma. Blood 98: 146–155. [DOI] [PubMed] [Google Scholar]
- 10. Heintel T, Sester M, Rodriguez MM, Krieg C, Sester U, et al. (2002) The fraction of perforin-expressing HIV-specific CD8 T cells is a marker for disease progression in HIV infection. Aids 16: 1497–1501. [DOI] [PubMed] [Google Scholar]
- 11. Appay V, Nixon DF, Donahoe SM, Gillespie GM, Dong T, et al. (2000) HIV-specific CD8(+) T cells produce antiviral cytokines but are impaired in cytolytic function. J Exp Med 192: 63–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Shankar P, Russo M, Harnisch B, Patterson M, Skolnik P, et al. (2000) Impaired function of circulating HIV-specific CD8(+) T cells in chronic human immunodeficiency virus infection. Blood 96: 3094–3101. [PubMed] [Google Scholar]
- 13. Yang OO, Lin H, Dagarag M, Ng HL, Effros RB, et al. (2005) Decreased perforin and granzyme B expression in senescent HIV-1-specific cytotoxic T lymphocytes. Virology 332: 16–19. [DOI] [PubMed] [Google Scholar]
- 14. Fry TJ, Connick E, Falloon J, Lederman MM, Liewehr DJ, et al. (2001) A potential role for interleukin-7 in T-cell homeostasis. Blood 97: 2983–2990. [DOI] [PubMed] [Google Scholar]
- 15. Fry TJ, Mackall CL (2001) Interleukin-7: master regulator of peripheral T-cell homeostasis? Trends Immunol 22: 564–571. [DOI] [PubMed] [Google Scholar]
- 16. Fry TJ, Mackall CL (2002) Interleukin-7 and immunorestoration in HIV: beyond the thymus. J Hematother Stem Cell Res 11: 803–807. [DOI] [PubMed] [Google Scholar]
- 17. Fry TJ, Moniuszko M, Creekmore S, Donohue SJ, Douek DC, et al. (2003) IL-7 therapy dramatically alters peripheral T-cell homeostasis in normal and SIV-infected nonhuman primates. Blood 101: 2294–2299. [DOI] [PubMed] [Google Scholar]
- 18. Schluns KS, Lefrancois L (2003) Cytokine control of memory T-cell development and survival. Nat Rev Immunol 3: 269–279. [DOI] [PubMed] [Google Scholar]
- 19. Huster KM, Busch V, Schiemann M, Linkemann K, Kerksiek KM, et al. (2004) Selective expression of IL-7 receptor on memory T cells identifies early CD40L-dependent generation of distinct CD8+ memory T cell subsets. Proc Natl Acad Sci U S A 101: 5610–5615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Kaech SM, Tan JT, Wherry EJ, Konieczny BT, Surh CD, et al. (2003) Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat Immunol 4: 1191–1198. [DOI] [PubMed] [Google Scholar]
- 21. Welch PA, Namen AE, Goodwin RG, Armitage R, Cooper MD (1989) Human IL-7: a novel T cell growth factor. J Immunol 143: 3562–3567. [PubMed] [Google Scholar]
- 22. Armitage RJ, Namen AE, Sassenfeld HM, Grabstein KH (1990) Regulation of human T cell proliferation by IL-7. J Immunol 144: 938–941. [PubMed] [Google Scholar]
- 23. Westermann J, Aicher A, Qin Z, Cayeux Z, Daemen K, et al. (1998) Retroviral interleukin-7 gene transfer into human dendritic cells enhances T cell activation. Gene Ther 5: 264–271. [DOI] [PubMed] [Google Scholar]
- 24. Soares MV, Borthwick NJ, Maini MK, Janossy G, Salmon M, et al. (1998) IL-7-dependent extrathymic expansion of CD45RA+ T cells enables preservation of a naive repertoire. J Immunol 161: 5909–5917. [PubMed] [Google Scholar]
- 25. Alderson MR, Sassenfeld HM, Widmer MB (1990) Interleukin 7 enhances cytolytic T lymphocyte generation and induces lymphokine-activated killer cells from human peripheral blood. J Exp Med 172: 577–587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Carini C, Essex M (1994) Interleukin 2-independent interleukin 7 activity enhances cytotoxic immune response of HIV-1-infected individuals. AIDS Res Hum Retroviruses 10: 121–130. [DOI] [PubMed] [Google Scholar]
- 27. Jicha DL, Mule JJ, Rosenberg SA (1991) Interleukin 7 generates antitumor cytotoxic T lymphocytes against murine sarcomas with efficacy in cellular adoptive immunotherapy. J Exp Med 174: 1511–1515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Finke S, Trojaneck B, Lefterova P, Csipai M, Wagner E, et al. (1998) Increase of proliferation rate and enhancement of antitumor cytotoxicity of expanded human CD3+ CD56+ immunologic effector cells by receptor-mediated transfection with the interleukin-7 gene. Gene Ther 5: 31–39. [DOI] [PubMed] [Google Scholar]
- 29. Lotze MT, Grimm EA, Mazumder A, Strausser JL, Rosenberg SA (1981) Lysis of fresh and cultured autologous tumor by human lymphocytes cultured in T-cell growth factor. Cancer Res 41: 4420–4425. [PubMed] [Google Scholar]
- 30. Rowshani AT, Uss A, Yong SL, van Lier RA, ten Berge IJ (2004) Effects of CD25 monoclonal antibody on proliferative and effector functions of alloactivated human T cells in vitro. Eur J Immunol 34: 882–899. [DOI] [PubMed] [Google Scholar]
- 31. Kos FJ, Mullbacher A (1992) Induction of primary anti-viral cytotoxic T cells by in vitro stimulation with short synthetic peptide and interleukin-7. Eur J Immunol 22: 3183–3185. [DOI] [PubMed] [Google Scholar]
- 32. Talayev VY, Zaichenko IY, Babaykina ON, Lomunova MA, Talayeva EB, et al. (2005) Ex vivo stimulation of cord blood mononuclear cells by dexamethasone and interleukin-7 results in the maturation of interferon-gamma-secreting effector memory T cells. Clin Exp Immunol 141: 440–448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Sawa Y, Arima Y, Ogura H, Kitabayashi C, Jiang JJ, et al. (2009) Hepatic interleukin-7 expression regulates T cell responses. Immunity 30: 447–457. [DOI] [PubMed] [Google Scholar]
- 34. Smyth MJ, Norihisa Y, Gerard JR, Young HA, Ortaldo JR (1991) IL-7 regulation of cytotoxic lymphocytes: pore-forming protein gene expression, interferon-gamma production, and cytotoxicity of human peripheral blood lymphocytes subsets. Cell Immunol 138: 390–403. [DOI] [PubMed] [Google Scholar]
- 35. Faller EM, McVey MJ, Kakal JA, MacPherson PA (2006) Interleukin-7 receptor expression on CD8 T-cells is downregulated by the HIV Tat protein. J Acquir Immune Defic Syndr 43: 257–269. [DOI] [PubMed] [Google Scholar]
- 36. Goodwin RG, Friend D, Ziegler SF, Jerzy R, Falk BA, et al. (1990) Cloning of the human and murine interleukin-7 receptors: demonstration of a soluble form and homology to a new receptor superfamily. Cell 60: 941–951. [DOI] [PubMed] [Google Scholar]
- 37. Noguchi M, Nakamura Y, Russell SM, Ziegler SF, Tsang M, et al. (1993) Interleukin-2 receptor gamma chain: a functional component of the interleukin-7 receptor. Science 262: 1877–1880. [DOI] [PubMed] [Google Scholar]
- 38. MacPherson PA, Fex C, Sanchez-Dardon J, Hawley-Foss N, Angel JB (2001) Interleukin-7 receptor expression on CD8(+) T cells is reduced in HIV infection and partially restored with effective antiretroviral therapy. J Acquir Immune Defic Syndr 28: 454–457. [DOI] [PubMed] [Google Scholar]
- 39. Boutboul F, Puthier D, Appay V, Pelle O, Ait-Mohand H, et al. (2005) Modulation of interleukin-7 receptor expression characterizes differentiation of CD8 T cells specific for HIV, EBV and CMV. Aids 19: 1981–1986. [DOI] [PubMed] [Google Scholar]
- 40. Koesters SA, Alimonti JB, Wachihi C, Matu L, Anzala O, et al. (2006) IL-7Ralpha expression on CD4(+) T lymphocytes decreases with HIV disease progression and inversely correlates with immune activation. Eur J Immunol 36: 336–344. [DOI] [PubMed] [Google Scholar]
- 41. Colle JH, Moreau JL, Fontanet A, Lambotte O, Joussemet M, et al. (2006) CD127 expression and regulation are altered in the memory CD8 T cells of HIV-infected patients–reversal by highly active anti-retroviral therapy (HAART). Clin Exp Immunol 143: 398–403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Paiardini M, Cervasi B, Albrecht H, Muthukumar A, Dunham R, et al. (2005) Loss of CD127 expression defines an expansion of effector CD8+ T cells in HIV-infected individuals. J Immunol 174: 2900–2909. [DOI] [PubMed] [Google Scholar]
- 43. Rethi B, Fluur C, Atlas A, Krzyzowska M, Mowafi F, et al. (2005) Loss of IL-7Ralpha is associated with CD4 T-cell depletion, high interleukin-7 levels and CD28 down-regulation in HIV infected patients. Aids 19: 2077–2086. [DOI] [PubMed] [Google Scholar]
- 44. Sasson SC, Zaunders JJ, Zanetti G, King EM, Merlin KM, et al. (2006) Increased plasma interleukin-7 level correlates with decreased CD127 and Increased CD132 extracellular expression on T cell subsets in patients with HIV-1 infection. J Infect Dis 193: 505–514. [DOI] [PubMed] [Google Scholar]
- 45. Vingerhoets J, Bisalinkumi E, Penne G, Colebunders R, Bosmans E, et al. (1998) Altered receptor expression and decreased sensitivity of T-cells to the stimulatory cytokines IL-2, IL-7 and IL-12 in HIV infection. Immunol Lett 61: 53–61. [DOI] [PubMed] [Google Scholar]
- 46. Ferrari G, King K, Rathbun K, Place CA, Packard MV, et al. (1995) IL-7 enhancement of antigen-driven activation/expansion of HIV-1-specific cytotoxic T lymphocyte precursors (CTLp). Clin Exp Immunol 101: 239–248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Benito JM, Lopez M, Lozano S, Gonzalez-Lahoz J, Soriano V (2008) Down-regulation of interleukin-7 receptor (CD127) in HIV infection is associated with T cell activation and is a main factor influencing restoration of CD4(+) cells after antiretroviral therapy. J Infect Dis 198: 1466–1473. [DOI] [PubMed] [Google Scholar]
- 48. Faller EM, Sugden SM, McVey MJ, Kakal JA, Macpherson PA (2010) Soluble HIV Tat Protein Removes the IL-7 Receptor α-Chain from the Surface of Resting CD8 T Cells and Targets It for Degradation. J Immunol 185: 2854–2866. [DOI] [PubMed] [Google Scholar]
- 49. Chang HC, Samaniego F, Nair BC, Buonaguro L, Ensoli B (1997) HIV-1 Tat protein exits from cells via a leaderless secretory pathway and binds to extracellular matrix-associated heparan sulfate proteoglycans through its basic region. Aids 11: 1421–1431. [DOI] [PubMed] [Google Scholar]
- 50. Ensoli B, Barillari G, Salahuddin SZ, Gallo RC, Wong-Staal F (1990) Tat protein of HIV-1 stimulates growth of cells derived from Kaposi’s sarcoma lesions of AIDS patients. Nature 345: 84–86. [DOI] [PubMed] [Google Scholar]
- 51. Ensoli B, Buonaguro L, Barillari G, Fiorelli V, Gendelman R, et al. (1993) Release, uptake, and effects of extracellular human immunodeficiency virus type 1 Tat protein on cell growth and viral transactivation. J Virol 67: 277–287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Frankel AD, Pabo CO (1988) Cellular uptake of the tat protein from human immunodeficiency virus. Cell 55: 1189–1193. [DOI] [PubMed] [Google Scholar]
- 53. Poggi A, Carosio R, Fenoglio D, Brenci S, Murdaca G, et al. (2004) Migration of V delta 1 and V delta 2 T cells in response to CXCR3 and CXCR4 ligands in healthy donors and HIV-1-infected patients: competition by HIV-1 Tat. Blood 103: 2205–2213. [DOI] [PubMed] [Google Scholar]
- 54. Rayne F, Debaisieux S, Yezid H, Lin YL, Mettling C, et al. (2010) Phosphatidylinositol-(4,5)-bisphosphate enables efficient secretion of HIV-1 Tat by infected T-cells. Embo J 29: 1348–1362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Xiao H, Neuveut C, Tiffany HL, Benkirane M, Rich EA, et al. (2000) Selective CXCR4 antagonism by Tat: implications for in vivo expansion of coreceptor use by HIV-1. Proc Natl Acad Sci U S A 97: 11466–11471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Helland DE, Welles JL, Caputo A, Haseltine WA (1991) Transcellular transactivation by the human immunodeficiency virus type 1 tat protein. J Virol 65: 4547–4549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Fittipaldi A, Giacca M (2005) Transcellular protein transduction using the Tat protein of HIV-1. Adv Drug Deliv Rev 57: 597–608. [DOI] [PubMed] [Google Scholar]
- 58. Vendeville A, Rayne F, Bonhoure A, Bettache N, Montcourrier P, et al. (2004) HIV-1 Tat enters T cells using coated pits before translocating from acidified endosomes and eliciting biological responses. Mol Biol Cell 15: 2347–2360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Yezid H, Konate K, Debaisieux S, Bonhoure A, Beaumelle B (2009) Mechanism for HIV-1 Tat insertion into the endosome membrane. J Biol Chem 284: 22736–22746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Ghazawi FM, Faller EM, Sugden SM, Kakal JA, MacPherson PA (2014) IL-7 downregulates IL-7Ralpha expression in human CD8 T cells by two independent mechanisms. Immunol Cell Biol 91: 149–158. [DOI] [PubMed] [Google Scholar]
- 61. Ensoli B, Gendelman R, Markham P, Fiorelli V, Colombini S, et al. (1994) Synergy between basic fibroblast growth factor and HIV-1 Tat protein in induction of Kaposi’s sarcoma. Nature 371: 674–680. [DOI] [PubMed] [Google Scholar]
- 62. Faller E, Kakal J, Kumar R, Macpherson P (2009) IL-7 and the HIV Tat protein act synergistically to down-regulate CD127 expression on CD8 T cells. Int Immunol 21: 203–216. [DOI] [PubMed] [Google Scholar]
- 63. Viscidi RP, Mayur K, Lederman HM, Frankel AD (1989) Inhibition of antigen-induced lymphocyte proliferation by Tat protein from HIV-1. Science 246: 1606–1608. [DOI] [PubMed] [Google Scholar]
- 64. Cohen SS, Li C, Ding L, Cao Y, Pardee AB, et al. (1999) Pronounced acute immunosuppression in vivo mediated by HIV Tat challenge. Proc Natl Acad Sci U S A 96: 10842–10847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Zagury D, Lachgar A, Chams V, Fall LS, Bernard J, et al. (1998) Interferon alpha and Tat involvement in the immunosuppression of uninfected T cells and C-C chemokine decline in AIDS. Proc Natl Acad Sci U S A 95: 3851–3856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Chirmule N, Than S, Khan SA, Pahwa S (1995) Human immunodeficiency virus Tat induces functional unresponsiveness in T cells. J Virol 69: 492–498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Re MC, Furlini G, Vignoli M, Ramazzotti E, Roderigo G, et al. (1995) Effect of antibody to HIV-1 Tat protein on viral replication in vitro and progression of HIV-1 disease in vivo. J Acquir Immune Defic Syndr Hum Retrovirol 10: 408–416. [DOI] [PubMed] [Google Scholar]
- 68. Re MC, Vignoli M, Furlini G, Gibellini D, Colangeli V, et al. (2001) Antibodies against full-length Tat protein and some low-molecular-weight Tat-peptides correlate with low or undetectable viral load in HIV-1 seropositive patients. J Clin Virol 21: 81–89. [DOI] [PubMed] [Google Scholar]
- 69. Zagury JF, Sill A, Blattner W, Lachgar A, Le Buanec H, et al. (1998) Antibodies to the HIV-1 Tat protein correlated with nonprogression to AIDS: a rationale for the use of Tat toxoid as an HIV-1 vaccine. J Hum Virol 1: 282–292. [PubMed] [Google Scholar]
- 70. Rezza G, Fiorelli V, Dorrucci M, Ciccozzi M, Tripiciano A, et al. (2005) The presence of anti-Tat antibodies is predictive of long-term nonprogression to AIDS or severe immunodeficiency: findings in a cohort of HIV-1 seroconverters. J Infect Dis 191: 1321–1324. [DOI] [PubMed] [Google Scholar]
- 71. Borsetti A, Baroncelli S, Maggiorella MT, Bellino S, Moretti S, et al. (2008) Viral outcome of simian-human immunodeficiency virus SHIV-89.6P adapted to cynomolgus monkeys. Arch Virol 153: 463–472. [DOI] [PubMed] [Google Scholar]
- 72. Borsetti A, Baroncelli S, Maggiorella MT, Moretti S, Fanales-Belasio E, et al. (2009) Containment of infection in tat vaccinated monkeys after rechallenge with a higher dose of SHIV89.6P(cy243). Viral Immunol 22: 117–124. [DOI] [PubMed] [Google Scholar]
- 73. Cafaro A, Caputo A, Fracasso C, Maggiorella MT, Goletti D, et al. (1999) Control of SHIV-89.6P-infection of cynomolgus monkeys by HIV-1 Tat protein vaccine. Nat Med 5: 643–650. [DOI] [PubMed] [Google Scholar]
- 74. Cafaro A, Caputo A, Maggiorella MT, Baroncelli S, Fracasso C, et al. (2000) SHIV89.6P pathogenicity in cynomolgus monkeys and control of viral replication and disease onset by human immunodeficiency virus type 1 Tat vaccine. J Med Primatol 29: 193–208. [DOI] [PubMed] [Google Scholar]
- 75. Ensoli B, Bellino S, Tripiciano A, Longo O, Francavilla V, et al. (2010) Therapeutic immunization with HIV-1 Tat reduces immune activation and loss of regulatory T-cells and improves immune function in subjects on HAART. PLoS One 5: e13540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Ensoli B, Fiorelli V, Ensoli F, Lazzarin A, Visintini R, et al. (2009) The preventive phase I trial with the HIV-1 Tat-based vaccine. Vaccine 28: 371–378. [DOI] [PubMed] [Google Scholar]
- 77. Ferrantelli F, Maggiorella MT, Schiavoni I, Sernicola L, Olivieri E, et al. (2011) A combination HIV vaccine based on Tat and Env proteins was immunogenic and protected macaques from mucosal SHIV challenge in a pilot study. Vaccine 29: 2918–2932. [DOI] [PubMed] [Google Scholar]
- 78. Jiang Q, Li WQ, Hofmeister RR, Young HA, Hodge DR, et al. (2004) Distinct regions of the interleukin-7 receptor regulate different Bcl2 family members. Mol Cell Biol 24: 6501–6513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Lee SH, Fujita N, Mashima T, Tsuruo T (1996) Interleukin-7 inhibits apoptosis of mouse malignant T-lymphoma cells by both suppressing the CPP32-like protease activation and inducing the Bcl-2 expression. Oncogene 13: 2131–2139. [PubMed] [Google Scholar]
- 80. Lenz DC, Kurz SK, Lemmens E, Schoenberger SP, Sprent J, et al. (2004) IL-7 regulates basal homeostatic proliferation of antiviral CD4+ T cell memory. Proc Natl Acad Sci U S A 101: 9357–9362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Sasson SC, Zaunders JJ, Kelleher AD (2006) The IL-7/IL-7 receptor axis: understanding its central role in T-cell homeostasis and the challenges facing its utilization as a novel therapy. Curr Drug Targets 7: 1571–1582. [DOI] [PubMed] [Google Scholar]
- 82. Sereti I, Dunham RM, Spritzler J, Aga E, Proschan MA, et al. (2009) IL-7 administration drives T cell-cycle entry and expansion in HIV-1 infection. Blood 113: 6304–6314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Liu S, Lizee G, Lou Y, Liu C, Overwijk WW, et al. (2007) IL-21 synergizes with IL-7 to augment expansion and anti-tumor function of cytotoxic T cells. Int Immunol 19: 1213–1221. [DOI] [PubMed] [Google Scholar]
- 84. Swainson L, Verhoeyen E, Cosset FL, Taylor N (2006) IL-7R alpha gene expression is inversely correlated with cell cycle progression in IL-7-stimulated T lymphocytes. J Immunol 176: 6702–6708. [DOI] [PubMed] [Google Scholar]