

Abstract
The blood-testis barrier (BTB) is found between adjacent Sertoli cells in the testis where it creates a unique microenvironment for the development and maturation of meiotic and postmeiotic germ cells in seminiferous tubes. It is a compound proteinous structure, composed of several types of cell junctions including tight junctions (TJs), adhesion junctions and gap junctions (GJs). Some of the junctional proteins function as structural proteins of BTB and some have regulatory roles. The deletion or functional silencing of genes encoding these proteins may disrupt the BTB, which may cause immunological or other damages to meiotic and postmeiotic cells and ultimately lead to spermatogenic arrest and infertility. In this review, we will summarize the findings on the BTB structure and function from genetically-modified mouse models and discuss the future perspectives.
Keywords: blood-testis barrier, genetically-modified mouse, seminiferous tubule, sertoli cells, spermatogenesis
INTRODUCTION
Blood-testis barrier (BTB) is found between adjacent Sertoli cells within the seminiferous tubules.1,2,3,4,5,6,7,8,9 The BTB divides the seminiferous tubules into the basal and apical (adluminal) compartments. Meiosis, spermiogenesis and spermiation take place in the apical compartment; whereas, spermatogonial cell division and differentiation to preleptotene spermatocytes occur in the basal compartment.10,11 Thus, the BTB creates a unique microenvironment for meiotic and postmeiotic cells by forming an immunological barrier that separates meiotic and postmeiotic germ cells from blood circulation (reviewed in12).
The BTB consists of several types of cellular junctions including tight junctions (TJs), gap junctions (GJs) and adhesion junctions, and many junctional proteins are involved in the establishment of BTB (reviewed in12,13,14). Defects in these proteins can cause BTB disfunction which may elicit immune responses against meiotic and postmeiotic cells, ultimately leading to spermatogenetic failure and male infertility. Furthermore, functions of BTB may also be compromised due to the defects of genes that regulate the formation and function of cell junctions. In this article, we will review recent findings in BTB functional genes obtained from genetically-modified mice.
TECHNOLOGIES USED FOR BTB FUNCTION STUDY
In vitro method
Since Sertoli cells cultured at high density in vitro show the ability to form junctions that mimic BTB to some extent,15,16 an in vitro system based on the culture of primary Sertoli cells has been established and used as a model for BTB study.17,18,19,20,21 Because it is relatively easy, quick and cheap, many studies have utilized this method to investigate the structure and function of the cell junctions.17,18,19,20,21,22,23,24 However, since the main function of BTB is to provide microenvironment for meiotic and postmeiotic cell development, this in vitro system is not suitable for the study of major aspects of BTB function because coculture of germ cells with Sertoli cells cannot achieve meiosis.25 Moreover, the BTB structure and/or function may also be affected by germ cells. Therefore, this primary Sertoli cell culture system is insufficicent for in-depth study of the structure, function and regulation of BTB in the testis.
In vivo method
Genetically-modified mice have been widely used to understand the functional roles of specific gene in development. There are two basic technical approaches used to produce genetically-modified mice, namely, transgenic and knockout (KO) mice.26,27,28 The transgenic mouse approach involves pronuclear injection into a zygote, where the gene of interest will randomly integrate into the mouse genome.29 The second approach, pioneered by Oliver Smithies and Mario Capecchi, involves modification of embryonic stem cells with a DNA construct containing DNA sequences homologous to the target gene.28 Embryonic stem cells with deletion of the target gene are selected and then injected into the mouse blastocysts. This manipulation causes absence of the gene (null) from all the cells of mouse. This approach, usually called conventional KO technology, is approporiate for investigating the physiological function of tissue or cell type-specific genes.30 A refined version of the KO technology, conditional KO (cKO), which is based on tissue and cell type-specific deletion of a gene of interest, shows significant advantages over conventional KO, especially for those genes whose conventional KO causes embryonic lethality.31 The most widely used approach at present for cKO is the Cre-LoxP system, which involves a ‘floxed’ mouse line bearing alleles of the gene to be deleted with recombinase-specific sites (i.e. two LoxP repeats flanking critical exons) and a transgenic mouse line expressing the Cre-recombinase driven by a promoter with a desired temporal and/or spatial expression pattern.32,33,34,35 The gene of interest flanked by two LoxP sites will be deleted or disrupted when Cre-recombinase is expressed in specified tissues or cell types at a specific development time point.32,33,34,35 Most resulting cKO mice have no evident developmental abnormalities in tissues except the one of interest and thus can be used for studies of gene function in a specified tissue or cell type at specific time point.
By using these genetic approaches, especially conventional and conditional gene KOs, about 400 genes involved in murine spermatogenesis have been inactivated,36,37,38 but only a few have been associated with the structure and function of BTB. In the following sections, we will summarize BTB-associated genes identified from genetically-modified mice.
DEFECTS IN SPERMATOGENESIS IN KO MICE OF BTB-ASSOCIATED PROTEINS
The BTB-associated genes are classified into two major groups based on their roles in BTB structure and function. The first group (Table 1) includes the known structural components of the BTB, and the other (Table 2) consists of those that regulate BTB formation, integrity and function. Since many of the mechanisms involved in this process are unknown, these may also include structural components.
Table 1.
Defects of spermatogenesis in the BTB structural genetically-modified mice
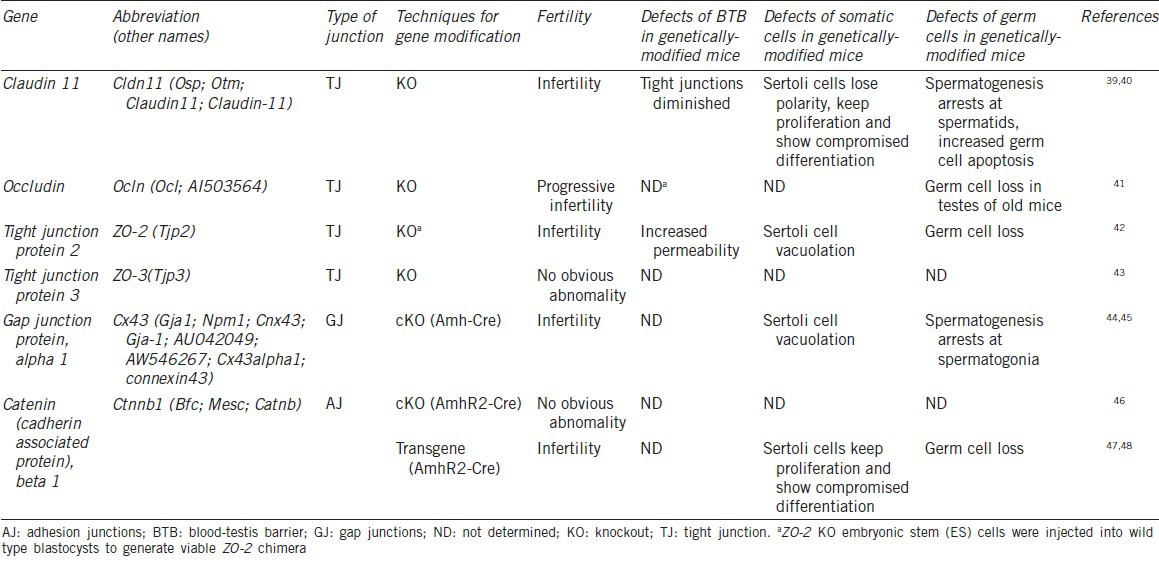
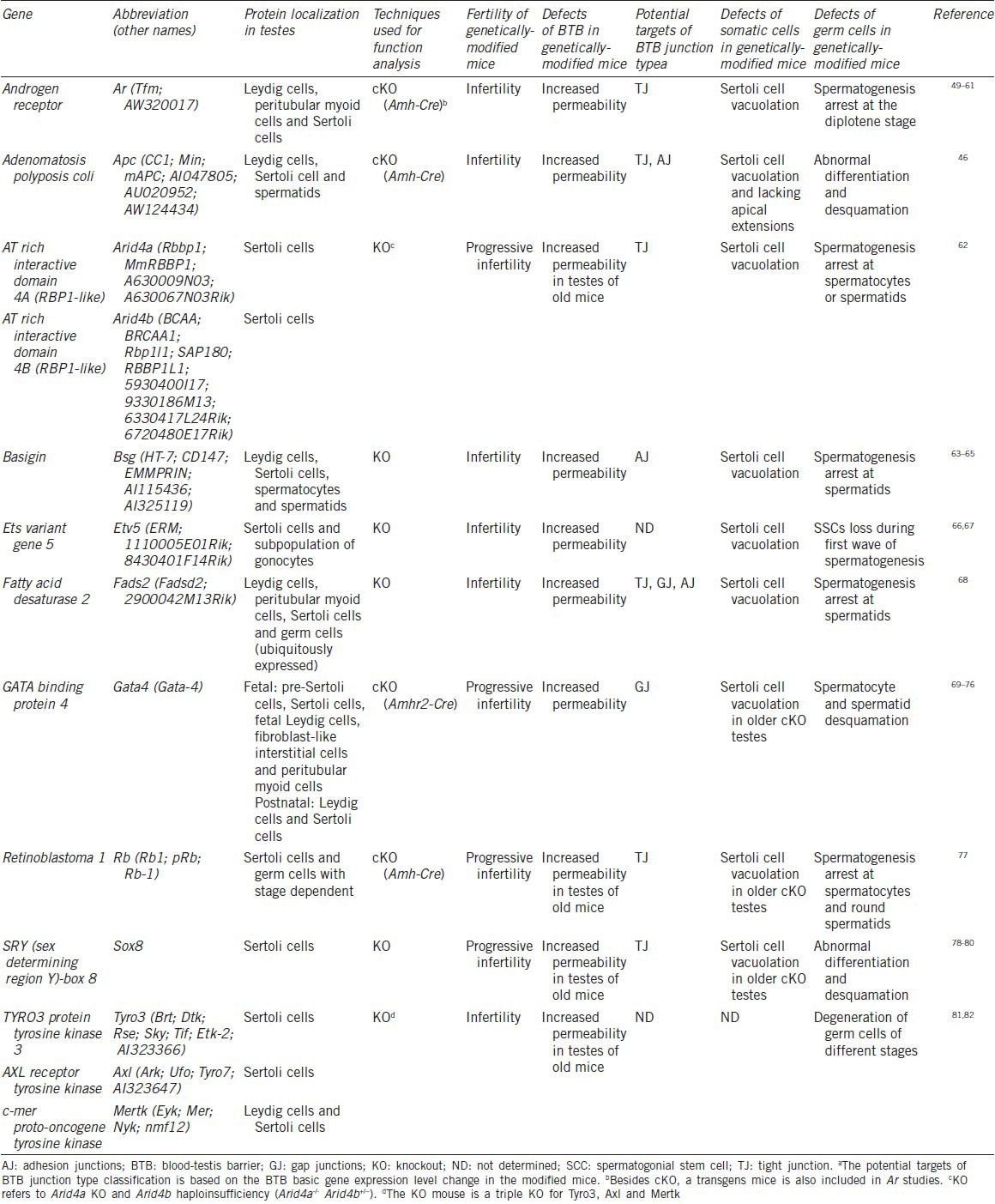
Table 2.
Defects of spermatogenesis in the BTB regulatory genetically-modified mice
Function of BTB structural components
Several cellular junctions function together to establish BTB with each type of cellular junction composed of multiple structural proteins. Deficiency in these proteins would cause significant damage to BTB and consequently spermatogenic failure.
Claudin-11 (Cldn-11)
Claudins, 20–27 kDa phosphoproteins, are the main constituents of the TJs in mammalian body.83,84,85,86 They are intercellular adhesion molecules with variable pore-like properties.87,88 To date, about 24 different claudins have been identified and many of them show a distinct organ-specific distribution.85,89,90 In mice and rat testes, Cldn-11 is specifically expressed in the Sertoli cells and responsible for the formation of the typically parallel tight junctional strands between Sertoli cells.85 In mouse testis, Cldn-11 expression peaks between postnatal day 6–16, coinciding with the BTB formation.91,92
Cldn-11 KO mice were the first mouse model used for the study of BTB.39 In prepubertal and adult Cldn-11 KO mice, the lumens of the seminiferous tubules are narrow and often filled with Sertoli cells.39,40 Adult mouse testes lacking Cldn-11 in Sertoli cells are devoid of a mature BTB and show increased apoptotic germ cells.40 Cldn-11 KO Sertoli cells lose polarity and detach from the basement membrane of seminiferous tubules. They experience an epithelial to fibroblastic cell transformation and proliferate actively while still maintaining the expression of Stertoli cell specific differentiation markers. As expected, Cldn-11 KO mice are sterile.39
Occludin (Ocln)
Occludin, a 65 kDa protein, was the first component of the TJ strand identified.93,94,95,96 It expresses in Sertoli cells, together with claudins, serving as a key component of TJs in BTB.96 In mouse, Ocln is detected by immunofluorescence in testis cords as early as embryonic day 13.5.97 By postnatal day 14, it is detected as focal wavy bands toward the base of seminiferous tubules that contain a number of germ cells.97 By postnatal day 23 and in adult mice, these bands are present in all tubules at all stages of seminiferous epithelial cycle.97 As in mice, Ocln is also detected at all stages of the seminiferous epithelial cycle in dogs and Korean wild rabbits Lepus coreanus.98,99 However, in rats, Ocln protein expression is stage-specific, expressing heavily in Sertoli cells in seminiferous tubes of all stages except stage VIII, where it is not detectable by immunostaining.96,99 Interestingly, Ocln is not expressed in seminiferous tubules of guinea pigs (Cavia porcellus) and humans.95,99
Compared to Cldn-11 KO mice, the abnormalities of spermatogenesis in Ocln KO mice increase slowly and gradually with ageing.39,40,41 In testis of 6-week-old Ocln KO mice, the seminiferous tubules and spermatogenesis are histo-cytologically indistinguishable from those in wide-type mice. Around 40–60 weeks of age, the seminiferous tubules of KO mice display atrophy. The atrophic tubules are devoid of germ cells, but retain Sertoli cells along the basement membrane.41 The exact mechanisms underlying this age-dependent effect in testis of Ocln KO mice remain unknown.
Zonula occludens (ZO) proteins
TJ integral membrane proteins such as claudins and Ocln are tethered to the actin cytoskeleton by adaptor proteins, notably the closely related ZO proteins ZO-1, ZO-2 and ZO-3.101,102 These three closely related and widely expressed ZO proteins belong to the membrane-associated guanylate kinase-like protein superfamily.101,102 ZO-1 protein is also known as TJ protein 1 (TJP1). It is detected at the inter-Sertoli cell junctions in testis of guinea pig and mouse.97,100,103,104 In normal human testis, ZO-1 and ZO-2 are observed at the adherent site of adjacent Sertoli cells.100,105 The nuclear localization of some ZO proteins is also reported in particular conditions.106,107,108,109 Recently, these three ZO proteins have been deleted in mice.110,111 Although ZO-3 KO mice lack an obvious phenotype, mice deficient in ZO-1 or ZO-2 shows early embryonic lethality.110,111 By microinjecting ZO-2 KO embryonic stem cells into wild-type mouse blastocysts, Xu et al. (2009)42 generated viable ZO-2 chimera.42 The adult chimera presented a set of phenotypes in different organs. Male ZO-2 chimera show reduced fertility and pathological changes in the testis. Lanthanum tracer experiments showed a compromised BTB function in these mice.42,112 Based on the gene expression and localization analyses, the authors found that the expression level of ZO-1, ZO-3, Cldn-11 and Ocln is not apparently affected when compared to the controls. ZO-1 and Ocln still localize to the BTB region, but Cldn-11 and Connexin43 are misslocalized from BTB. These results indicate there is limited redundance between ZO-2 and other ZO proteins in adult mice.
Connexin-43 (Cx 43)
Cx43, also known as GJ protein alpha 1 (Gja1), is the predominant testicular GJ protein located between adjacent Sertoli cells and between Sertoli cells and germ cells.43 It is colocalized with Ocln, ZO-1 and N-cadherin at the base of the epithelium, and also observed at the focal sites in the epithelium.97 To study the function of Cx43 in spermatogenesis, mice with Cx43 specifically deleted in Sertoli cells have been generated.44,45 Studies on these mice revealed that the expression of Cx43 in Sertoli cells is required for normal testicular development and initiation of spermatogenesis.44,45 Adult Sertoli cell-specific Cx43 cKO mice are sterile with a dramatic reduction in size and weight of testes.44,45 Their spermatogenesis is arrested at spermatogonia in 95% of seminiferous tubules with the number of spermatogonia dramatically decreased and Sertoli cells increased. Sertoli cell-only syndrome and Sertoli cell clusters are also noted in these mice.44,45
Cadherin associated protein beta 1 (β-catenin)
Cadherin associated protein beta 1 (Cttnb1, β-catenin) is a multifunctional molecule that functions in intercellular adhesion and signal transduction.113,114 It is colocalized with N-cadherin between adjacent Sertoli cells in the seminiferous tubules near the basal and the lower one-third of the adluminal compartments, and also at cell-cell contacts sites between Sertoli cells and spermatocytes in testes of Sprague-Dawley rats.115,116 N-cadherin is considered as a structural component of BTB, so the colocalization of β-catenin with N-cadherin at the inter-Sertoli cells contact point suggests that it is also an integral component of BTB.12,117,118 β-catenin is also an essential component of the WNT/β-catenin signaling pathway, which plays important roles in multiple developmental processes including testis development.119,120,121
By crossing with mice expressing Cre recombinase driven by the anti-Mullerian hormone (AMH) type II receptor promoter (Amhr2) in Sertoli cells, Ctnnb1 is specifically deleted in Sertoli cells.45 Histological examination of testes of adult (>12 weeks) Ctnnb1 cKO mice does not show any abnormities in testicular morphology.45 Constitutively activated β-catenin in Sertoli cells leads to continuous proliferation and compromised differentiation of Sertoli cells.46,47 Compared with the controls, Sertoli cells in the adult mutant mice still express AMH and glial cell-derived neurotrophic factor (GDNF) at high levels, which are normally expressed only in immature Sertoli cells. Defective differentiation of germ cells and increased apoptosis were observed in these mutant mice. As expected, the epididymis of the adult mutant mice are devoid of sperm.47 Besides, as a structural component of BTB, it also plays an essential role in the regulation of Sertoli cell proliferation and differentiation. Actually, it has been reported to regulate cell proliferation and differentiation through the WNT/β-catenin signaling pathway.118,121,122 As for its role in BTB, based on the observation that its deficiency does not cause detectable reproductive defects, we speculate that β-catenin may just serve as an adaptor for N-cadherin. To confirm or refute this speculation, more studies are required.
Function of BTB regulatory elements
Androgen receptor
Androgen receptor (Ar), a member of the steroid hormone receptor superfamily, mediates androgen action and plays an important role in male reproduction (reviewed in124,125,126,127). In testes, Ar can be detected in Sertoli cells, peritubular myoid cells and cells in the interstitial spaces including Leydig cells and perivascular smooth muscle cells.49,50,55,56,57,58 It has been reported that the Sertoli cell-specific Ar cKO mice are infertile, due to spermatogenic arrest predominately at the diplotene stage with almost no sperm observed in the epididymis.51,52,59,61 The defects in BTB structure of these cKO mice are associated with the reduced expression of BTB proteins like Cldn-11, ZO-1, Ocln and gelsolin and with a significantly enhanced expression of vimentin.52,54,60 It is noteworthy that the Arflox (ex1-neo)/Y mice had a partial defect in androgen sensitivity when carry this floxed allele, and a marked reduction in AR protein levels in different tissues including the testis and show defects in spermiogenesis.51 The BTB in Arflox (ex1-neo)/Y/Amh-Cre mice is disrupted, possibly due to the reduced expression of Cldn-3.53 These results from Ar mouse models indicate that the function of AR in Sertoli cells is essential for the maintenance of fully competent Sertoli cell function in BTB integrity as well as the sustenance of appropriate hormone levels to support the completion of spermatogenesis.51,52,59,61,128,129,130
Adenomatous polyposis coli (Apc)
Mutations in Apc, a multifunction tumor suppressor protein, are associated with the development of various human cancers, including colon, liver, ovarian, endometrial and testicular cancers.131,132,133,134 In a mouse model that expresses a truncated form of Apc in Sertoli cells, despite having normal embryonic and early postnatal testicular development, premature germ cell loss and Sertoli cell only (SCO) seminiferous tubules were observed.46 The cKO of Apc does not affect the Sertoli cell quiescence, apoptosis or differentiation, as evidenced by the absence of proliferating cell nuclear antigens and DNA damages in Sertoli cells, as well as AMH expression.46 However, these Sertoli cells lose their apical extensions, which normally enclose germ cells at late stages of spermatogenesis.46 As for the BTB structure, ZO-1 and N-cadherin proteins are seen as diffused and away from the BTB site in Apc cKO testes.46 As a result, deficiency of the Apc in Sertoli cells disrupts the BTB and causes spermatogenic failure most probably by affecting localization of junctional proteins.46
AT rich interactive domain 4A and AT rich interactive domain 4B
AT rich interactive domain 4A (Arid4a) and AT rich interactive domain 4B (Arid4b) are members of the ARID (AT-rich interaction domain) gene family. ARID4A and ARID4B proteins, also known as RB-binding protein 1 (RBBP1, RBP1) and RBBP1-like protein 1 (RBBP1L1), are the members of chromatin-remodeling complex and function as transcriptional repressors upon recruitment by RB.135,135,136,137,138 In situ hybridization analysis reveal that Arid4a and Arid4b are expressed mainly in Sertoli cells of testes.62 Mice with complete deficiency of Arid4a and haploinsufficiency of Arid4b showed progressive loss of male fertility, accompanied by impaired BTB, hypogonadism and seminal vesicle agenesis/hypodysplasia.62 These mice show spermatogenic arrest at meiotic spermatocytes or postmeiotic spermatids.62 These observations recapitulate the defects found in the Sertoli cell-specific Ar KO mice and the Sertoli cell-specific Rb KO mice.62 Gene expression evaluation revealed that ARID4A and ARID4B contribute to the optimal expression of Cldn-3 by functioning as positive coregulators in the context of the AR and RB complex.62 Furthermore, increased permeability of the BTB in the testes of Arid4a KO and Arid4b haploinsufficiency mice are observed based on a biotin tracer injection experiment. Together, Arid4a and Arid4b are critical for physiological function of Sertoli cells.
Basigin (Bsg)
Bsg is a transmembrane glycoprotein enriched with N-glycans.139,140 It is highly expressed in gonads and plays a crucial role in both male and female reproduction.64,65 In the mouse testis, BSG is expressed in Sertoli cells, Leydig cells, spermatocytes and spermatids.64,65 Bsg KO males are sterile.65 The Bsg KO testes are devoid of elongated spermatids and mature spermatozoa but have numerous round spermatids.63 Significantly increased apoptotic germ cells and compromised integrity of the BTB are observed in Bsg KO testes. Immunolocalization analysis of BTB component proteins indicates that no obvious difference in the localization of Cxadr, Cx43 or Cldn-11 are seen between wild type and Bsg KO testes, however, the expression of N-cadherin was greatly reduced at the basal compartment of the seminiferous tubules (the site of the BTB) in Bsg KO mice.63 These results imply that Bsg deficiency can compromise BTB integrity.
Ets-variant gene 5
The Ets-variant gene 5 (Etv5), also known as Ets-related molecule or ERM, is a member of the PEA3 subfamily of the ETS family of transcription factors. It is mainly expressed in adult Sertoli cells.66,67 Mice with a targeted deletion of Etv5 can undergo first wave of spermatogenesis, but lose their spermatogonial stem cells during this time, and subsequently show SCO phenotype.66 The disappearance of spermatogonial stem cells in the mutants is attributed to the failure of spermatogonial stem cell proliferation without affecting their differentiation by lack of Etv5. The integrity of BTB in Etv5 KO mice is disturbed which was shown by biotin tracer experiment.67 Whether the BTB is regulated by Etv5 directly or indirectly is still unknown, which deserves the further analysis.
Fatty acid desaturase 2
Fatty acid desaturase 2 (Fads2) is responsible for the initial step in the enzymatic cascade of ω3- and ω6-polyunsaturated fatty acid synthesis from essential fatty acids.142,143,144 Fads2 KO mice are sterile, their testis weight is reduced to two-thirds of that of age-matched wild type littermates.68 The lumen of the seminiferous tubules and epididymis of the adult mutants lacks spermatozoa.68 The epididymal ductuli fill with detritus and immature spermatids. Immunohistochemical studies revealed that Ocln, Cldn-11, JAM-A, ZO-1, Cx43 and β-catenin are dislocated throughout the basolateral and apical compartments of the Fads2 KO Sertoli cell membrane.68 Furthermore, transmission electron microscopic analysis highlighted that the well-structured TJ structures between Sertoli cells are missing in Fads2 KO testes. Finally, compromised selective permeation of BTB in KO testes has been revealed by the lanthanum nitrate and fluorescence dyes perfusion experiments.68
GATA binding protein 4
Transcription factor GATA binding protein 4 (Gata4) has been implicated in the development and function of the mammalian testis.145 During fetal testicular development, Gata4 is expressed in pre-Sertoli cells, Sertoli cells, Leydig cells, fibroblast-like interstitial cells and peritubularmyoid cells.74,76 After birth, Gata4 is found mainly in the Sertoli cells and adult Leydig cells.69,70,71,72,75 Mice, whose Gata4 conditionally is deleted in Sertoli cells, develop age-dependent testicular atrophy and are infertile.73 Histological analysis demonstrated that the older cKO testes displayed Sertoli cell vacuolation, germ cell depletion, multinucleated giant cells and syncytia of degenerating spermatids.73 Biotinylated tracer injection experiments indicate that the BTB appeared intact in young cKO mice (2.5 months), but it had a compromised integrity in the 6-month-old cKO mice.73 Furthermore, biotinylated germ cells, including multinuclear giant cells were evident in seminiferous tubules of 6-month-old cKO mice.73 Thus, the older Gata4 cKO mice develop increased permeability of the BTB with the advancing of age.
Retinoblastoma 1 (RB)
RB protein, encoded by Rb gene, is a negative regulator of the cell cycle and the first tumor suppressor found.146,147 The Sertoli cell-specific Rb KO mice displayed progressive infertility in males.77 Initially, loss of Rb in Sertoli cells has no gross effect on Sertoli cell function and the mice are fertile at 6 week of age.77 However, by the age of 10–14 weeks, the cKO mice demonstrated severe Sertoli cell dysfunction and infertility.77 The most striking defects in mature Sertoli cell function are increased permeability of the BTB by biotin tracer experiment.77 Detailed analysis found that TJ components, Cldn-3 and Ocln, are downregulated in Rb cKO Sertoli cells.77 The progressive loss of integrity of BTB in the Rb cKO testes suggested that Rb was initially dispensable for the formation of the BTB but might be indispensable for its remodeling as maturing germ cells crossed from the basal to adluminal compartment and this function might be directly related to the regulation of TJ genes.
Sex determining region Y-box8
Sex determining region Y-box 8 (Sox8) is a member of the Sox family of developmental transcription factor genes and is closely related to Sox9, a key gene in testis determination pathway in mammals.148,149,150,151 In testis, it is expressed in the developing mouse testis around the time of sex determination and continues beyond 16 days post coitum in Sertoli cells.79 Sox8 KO mice exhibit a progressive male infertility phenotype.80 These KO males sporadically produced litters of reduced size at young ages and showed an age-dependent deregulation of spermatogenesis, characterized by sloughing of spermatocytes and round spermatids, spermiation failure and a progressive disorganization of the spermatogenic cycle, which resulted in the inappropriate placement and juxtaposition of germ cell types within the epithelium.80 Cldn-3 was significantly decreased in the Sox8 KO testes.78 Furthermore, the use of biotin tracers showed increased BTB permeability in the Sox8 KO adult testes.78 Thus, Sox8 is essential in Sertoli cells for germ cell differentiation, partly by controlling the microenvironment of the seminiferous epithelium.
TYRO3 protein tyrosine kinase 3, AXL receptor tyrosine kinase and c-mer proto-oncogene tyrosine kinase
TYRO3 protein tyrosine kinase 3 (Tyro3), AXL receptor tyrosine kinase (Axl) and c-mer proto-oncogene tyrosine kinase (Mer) constitute the TAM family of receptor tyrosine kinases, characterized by a conserved sequence within the kinase domain and adhesion molecule-like extracellular domains.152,153 This small family of receptor tyrosine kinases regulates an intriguing mix of processes, including cell proliferation, survival, cell adhesion and migration and regulation of inflammatory cytokine release.152,153,154 Tyro3, Axl and Mer (TAM) receptor tyrosine kinases triple KO (TAM KO) male mice are infertile due to impaired spermatogenesis.81,82 These triple KO testes showed a progressive loss of germ cells from elongated spermatids to spermatogonia.82 Young adult TAM KO mice exhibited oligo-astheno-teratozoospermia and various morphological malformations of the sperm.82 With the progress of mice age, germ cells were eventually depleted from the seminiferous tubules. Furthermore, biotin can be detected in the seminiferous tubules of 20- to 30-week-old testes indicating that BTB was initially built in TAM KO mice, but subsequently compromised as the mice aged.82 Moreover, major inflammatory cytokines, including tumor necrosis factor-α, interleukin-6 and monocyte chemotactic protein 1 were upregulated in the testis of TAM KO mice, and predominantly located in Sertoli cells.82 It is therefore suggested that the TAM receptors are important in the maintenance of the immune homeostasis in the testis through the BTB.
CONCLUSIONS
Based on the literature reviewed above, we conclude that:
BTB gene KO mice, once their BTB integrity is compromised, always show some common abnormalities, e.g. germ cell apoptosis, development arrest, aggregated Sertoli cells in apical compartment, SCO phenotype and infertility. This indicates that the BTB integrity is essential for normal spermatogenesis and male fertility
Deletion of genes encoding proteins involved in different types of junctions often causes different phenotypes in seminiferous tubules, suggesting that different junction types in the BTB may play distinct role in maintaining the integrity of BTB in structure and function
Deletion of different genes of the same cell junction composed of the BTB, e.g. Cldn, Ocln and ZO-2, causes slightly different abnormalities in testicular tubules and fertility of animals, which indicates that these proteins function in non-redundant manner
Although the interactions between germ cells and Sertoli cells are believed to play a role in BTB function and integrity, a direct convincing evidence to support this hypothesis, where BTB is compromised after specific deletion of a gene in germ cells, is still lacking.
FUTURE PERSPECTIVES
BTB and male infertility
Unexplained male infertility accounts for 30%–40% of men with abnormal semen parameters.155 The causes of spermatogenic defects in infertile patients are multifactorial. Endocrine disruption of testicular development during neonatal period, due to environmental pollution, genetic and epigenetic factors, is the most frequent explanation invoked for unexplained male infertility.155,156,157,158 These factors have been associated with testicular dysgenesis, male infertility and recently testicular malignancy.155 It is predicted that these multifactors are associated with the BTB and could participate in the etiopathology of human male infertility by dysregulating BTB. For example, cKO of Cx43 in mouse Sertoli cells results in a very similar spermatogenic failure seen in infertile men.44,45,159,160 Azoospermic patients with severe spermatogenic failure have been reported to show altered expression of Cx43 mRNA.161 Furthermore, significantly positive correlation is reported between the histological score and intensity of the testicular Cx43 expression in oligozoospermic men.162 Similar staining pattern of Cx43 are found in testes of healthy men and patients with hypospermatogenesis or spermatogenic arrest at meiotic and postmeiotic stages, while no staining is observed in the seminiferous tubules of patients with spermatogenic arrest at spermatogonia or SCO syndrome.163 It is thus suggested that, to understand the etipathology of human infertility, the expression and localization of BTB proteins should be studied in men with spermatogenic defects and compared to those observed in BTB gene KO mice.
BTB and spermatogenic microenvironment or biomarkers
In testis, blood vessels, lymphatic vessels and nerves are only present in the interstitium between seminiferous tubules, but not inside the seminiferous tubules. The entry of nutrients (e.g. sugars, amino acids) and regulatory molecules (e.g. hormones, electrolytes), but not toxicants (e.g. environmental toxicants, drugs, chemicals) into the apical compartment where meiotic and postmeiotic germ cells reside is tightly regulated by BTB.12 The selectivity of BTB, thus, provides a unique microenvironment for the development of meiotic and postmeiotic germ cells in the apical compartment.12 The BTB may also function to prevent some molecules from emission from the apical compartment. It is, therefore, reasonable to think that if the integrity of BTB is compromised, some molecules that are only present within the seminiferous tubules normally may diffuse into the blood. These molecules can be used as circulation blood biomarkers of the integrity of BTB or the damage to the microenvironment of spermatogenesis.
BTB and cell specific conditional KO strategy
Normal BTB formation and function require numerous genes, many of which are ubiquitously expressed and function in other organs as well. Conventional KO of these genes may cause embryonic or perinatal lethality in homozygotes, e.g. ZO-1 or ZO-2 KO mice show early embryonic lethality.106,107 Even if the KO of a ubiquitously expressed gene is not lethal, it may cause alterations in the physiology of many organs, which could complicate the studies especially for reproduction, the process also regulated tightly by hypothalamic and pituitary. Therefore, the cKO approach shows obvious advantages over conventional KO. However, till now, only a few genes have been investigated by specific deletion in Sertoli cells for their role in BTB and spermatogenesis. Therefore, to delineate the function of BTB in spermatogenesis, much work is needed by using the conditional gene KO approach.
AUTHOR CONTRIBUTIONS
QHS, XHJ and IB conceived the ideas for preparing this review article, wrote the first draft and prepared the final version of the manuscript. WZ, SY, ZW and HJC modified the manuscript.
COMPETING INTERESTS
All authors declare no competing interests.
ACKNOWLEDGMENTS
This work was supported by the National Basic Research Program (Nos. 2013CB947900, 2013CB945502 and 2014CB943101) of China (973), by grants from National Natural Science Foundation of China (No. 31371519) and the Knowledge Innovation Program of the Chinese Academy of Sciences (No. KSCX2-EW-R-07).
REFERENCES
- 1.Chiquoine AD. Observations on the early events of cadmium necrosis of the testis. Anat Rec. 1964;149:23–35. doi: 10.1002/ar.1091490104. [DOI] [PubMed] [Google Scholar]
- 2.Kormano M. Distribution of injected L-3,4-dihydroxyphenylalanine (L-dopa) in the adult rat testis and epididymis. Acta Physiol Scand. 1967;71:125–6. doi: 10.1111/j.1748-1716.1967.tb03717.x. [DOI] [PubMed] [Google Scholar]
- 3.Dym M, Fawcett DW. The blood-testis barrier in the rat and the physiological compartmentation of the seminiferous epithelium. Biol Reprod. 1970;3:308–26. doi: 10.1093/biolreprod/3.3.308. [DOI] [PubMed] [Google Scholar]
- 4.Kormano M. Dye permeability and alkaline phosphatase activity of testicular capillaries in the postnatal rat. Histochimie. 1967;9:327–38. doi: 10.1007/BF00305816. [DOI] [PubMed] [Google Scholar]
- 5.Kormano M. Penetration of intravenous trypan blue into the rat testis and epididymis. Acta Histochem. 1968;30:133–6. [PubMed] [Google Scholar]
- 6.Fawcett DW, Leak LV, Heidger PM., Jr Electron microscopic observations on the structural components of the blood-testis barrier. J Reprod Fertil Suppl. 1970;10:105–22. [PubMed] [Google Scholar]
- 7.Dym M, Cavicchia JC. Further observations on the blood-testis barrier in monkeys. Biol Reprod. 1977;17:390–403. doi: 10.1095/biolreprod17.3.390. [DOI] [PubMed] [Google Scholar]
- 8.Dym M. Basement membrane regulation of Sertoli cells. Endocr Rev. 1994;15:102–15. doi: 10.1210/edrv-15-1-102. [DOI] [PubMed] [Google Scholar]
- 9.Siu MK, Cheng CY. Dynamic cross-talk between cells and the extracellular matrix in the testis. Bioessays. 2004;26:978–92. doi: 10.1002/bies.20099. [DOI] [PubMed] [Google Scholar]
- 10.Russell L. Movement of spermatocytes from the basal to the adluminal compartment of the rat testis. Am J Anat. 1977;148:313–28. doi: 10.1002/aja.1001480303. [DOI] [PubMed] [Google Scholar]
- 11.Fawcett DW. Intercellular bridges. Exp Cell Res. 1961;(Suppl 8):174–87. doi: 10.1016/0014-4827(61)90347-0. [DOI] [PubMed] [Google Scholar]
- 12.Cheng CY, Mruk DD. The blood-testis barrier and its implications for male contraception. Pharmacol Rev. 2012;64:16–64. doi: 10.1124/pr.110.002790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cheng CY, Mruk DD. A local autocrine axis in the testes that regulates spermatogenesis. Nat Rev Endocrinol. 2010;6:380–95. doi: 10.1038/nrendo.2010.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pelletier RM, Byers SW. The blood-testis barrier and Sertoli cell junctions: structural considerations. Microsc Res Tech. 1992;20:3–33. doi: 10.1002/jemt.1070200104. [DOI] [PubMed] [Google Scholar]
- 15.Byers SW, Hadley MA, Djakiew D, Dym M. Growth and characterization of polarized monolayers of epididymal epithelial cells and Sertoli cells in dual environment culture chambers. J Androl. 1986;7:59–68. doi: 10.1002/j.1939-4640.1986.tb00871.x. [DOI] [PubMed] [Google Scholar]
- 16.Janecki A, Steinberger A. Polarized Sertoli cell functions in a new two-compartment culture system. J Androl. 1986;7:69–71. doi: 10.1002/j.1939-4640.1986.tb00873.x. [DOI] [PubMed] [Google Scholar]
- 17.Hadley MA, Byers SW, Suarez-Quian CA, Kleinman HK, Dym M. Extracellular matrix regulates Sertoli cell differentiation, testicular cord formation, and germ cell development in vitro. J Cell Biol. 1985;101:1511–22. doi: 10.1083/jcb.101.4.1511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Mruk DD, Cheng CY. An in vitro system to study Sertoli cell blood-testis barrier dynamics. Methods Mol Biol. 2011;763:237–52. doi: 10.1007/978-1-61779-191-8_16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Janecki A, Jakubowiak A, Steinberger A. Regulation of transepithelial electrical resistance in two-compartment Sertoli cell cultures: in vitro model of the blood-testis barrier. Endocrinology. 1991;129:1489–96. doi: 10.1210/endo-129-3-1489. [DOI] [PubMed] [Google Scholar]
- 20.Janecki A, Jakubowiak A, Steinberger A. Effects of cyclic AMP and phorbol ester on transepithelial electrical resistance of Sertoli cell monolayers in two-compartment culture. Mol Cell Endocrinol. 1991;82:61–9. doi: 10.1016/0303-7207(91)90009-h. [DOI] [PubMed] [Google Scholar]
- 21.Okanlawon A, Dym M. Effect of chloroquine on the formation of tight junctions in cultured immature rat Sertoli cells. J J Androl. 1996;17:249–55. [PubMed] [Google Scholar]
- 22.Siu MK, Wong CH, Lee WM, Cheng CY. Sertoli-germ cell anchoring junction dynamics in the testis are regulated by an interplay of lipid and protein kinases. J Biol Chem. 2005;280:25029–47. doi: 10.1074/jbc.M501049200. [DOI] [PubMed] [Google Scholar]
- 23.Wong CH, Mruk DD, Siu MK, Cheng CY. Blood-testis barrier dynamics are regulated by {alpha} 2-macroglobulin via the c-Jun N-terminal protein kinase pathway. Endocrinology. 2005;146:1893–908. doi: 10.1210/en.2004-1464. [DOI] [PubMed] [Google Scholar]
- 24.Siu ER, Wong EW, Mruk DD, Porto CS, Cheng CY. Focal adhesion kinase is a blood-testis barrier regulator. Proc Natl Acad Sci U S A. 2009;106:9298–303. doi: 10.1073/pnas.0813113106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Loir M. Trout Sertoli cells and germ cells in primary culture: I. Morphological and ultrastructural study. Gamete Res. 1989;24:151–69. doi: 10.1002/mrd.1120240204. [DOI] [PubMed] [Google Scholar]
- 26.Jaenisch R, Mintz B. Simian virus 40 DNA sequences in DNA of healthy adult mice derived from preimplantation blastocysts injected with viral DNA. Proc Natl Acad Sci U S A. 1974;71:1250–4. doi: 10.1073/pnas.71.4.1250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gordon JW, Ruddle FH. Integration and stable germ line transmission of genes injected into mouse pronuclei. Science. 1981;214:1244–6. doi: 10.1126/science.6272397. [DOI] [PubMed] [Google Scholar]
- 28.Thomas KR, Capecchi MR. Site-directed mutagenesis by gene targeting in mouse embryo-derived stem cells. Cell. 1987;51:503–12. doi: 10.1016/0092-8674(87)90646-5. [DOI] [PubMed] [Google Scholar]
- 29.Gordon JW, Scangos GA, Plotkin DJ, Barbosa JA, Ruddle FH. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc Natl Acad Sci USA. 1980;77:7380–4. doi: 10.1073/pnas.77.12.7380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sun QY, Liu K, Kikuchi K. Oocyte-specific knockout: a novel in vivo approach for studying gene functions during folliculogenesis, oocyte maturation, fertilization, and embryogenesis. Biol Reprod. 2008;79:1014–20. doi: 10.1095/biolreprod.108.070409. [DOI] [PubMed] [Google Scholar]
- 31.Xu X, Fu XD. Conditional knockout mice to study alternative splicing in vivo. Methods. 2005;37:387–92. doi: 10.1016/j.ymeth.2005.07.019. [DOI] [PubMed] [Google Scholar]
- 32.Araki K, Imaizumi T, Okuyama K, Oike Y, Yamamura K. Efficiency of recombination by Cre transient expression in embryonic stem cells: comparison of various promoters. J Biochem. 1997;122:977–82. doi: 10.1093/oxfordjournals.jbchem.a021860. [DOI] [PubMed] [Google Scholar]
- 33.Kellendonk C, Tronche F, Reichardt HM, Schutz G. Mutagenesis of the glucocorticoid receptor in mice. J Steroid Biochem Mol Biol. 1999;69:253–9. doi: 10.1016/s0960-0760(99)00041-2. [DOI] [PubMed] [Google Scholar]
- 34.Nagy A. Cre recombinase: the universal reagent for genome tailoring. Genesis. 2000;26:99–109. [PubMed] [Google Scholar]
- 35.Sauer B, Henderson N. Site-specific DNA recombination in mammalian cells by the Cre recombinase of bacteriophage P1. Proc Natl Acad Sci U S A. 1988;85:5166–70. doi: 10.1073/pnas.85.14.5166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jamsai D, O’Bryan MK. Mouse models as tools in fertility research and male-based contraceptive development. Handb Exp Pharmacol. 2010:179–94. doi: 10.1007/978-3-642-02062-9_10. [DOI] [PubMed] [Google Scholar]
- 37.Massart A, Lissens W, Tournaye H, Stouffs K. Genetic causes of spermatogenic failure. Asian J Androl. 2012;14:40–8. doi: 10.1038/aja.2011.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yatsenko AN, Iwamori N, Iwamori T, Matzuk MM. The power of mouse genetics to study spermatogenesis. J Androl. 2010;31:34–44. doi: 10.2164/jandrol.109.008227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gow A, Southwood CM, Li JS, Pariali M, Riordan GP, et al. CNS myelin and sertoli cell tight junction strands are absent in Osp/claudin-11 null mice. Cell. 1999;99:649–59. doi: 10.1016/s0092-8674(00)81553-6. [DOI] [PubMed] [Google Scholar]
- 40.Mazaud-Guittot S, Meugnier E, Pesenti S, Wu X, Vidal H, et al. Claudin 11 deficiency in mice results in loss of the Sertoli cell epithelial phenotype in the testis. Biol Reprod. 2010;82:202–13. doi: 10.1095/biolreprod.109.078907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Saitou M, Furuse M, Sasaki H, Schulzke JD, Fromm M, et al. Complex phenotype of mice lacking occludin, a component of tight junction strands. Mol Biol Cell. 2000;11:4131–42. doi: 10.1091/mbc.11.12.4131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Xu J, Anuar F, Ali SM, Ng MY, Phua DC, et al. Zona occludens-2 is critical for blood-testis barrier integrity and male fertility. Mol Biol Cell. 2009;20:4268–77. doi: 10.1091/mbc.E08-12-1236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Brehm R, Steger K. Regulation of Sertoli cell and germ cell differentation. Adv Anat Embryol Cell Biol. 2005;181:1–93. [PubMed] [Google Scholar]
- 44.Sridharan S, Simon L, Meling DD, Cyr DG, Gutstein DE, et al. Proliferation of adult sertoli cells following conditional knockout of the Gap junctional protein GJA1 (connexin 43) in mice. Biol Reprod. 2007;76:804–12. doi: 10.1095/biolreprod.106.059212. [DOI] [PubMed] [Google Scholar]
- 45.Brehm R, Zeiler M, Ruttinger C, Herde K, Kibschull M, et al. A sertoli cell-specific knockout of connexin43 prevents initiation of spermatogenesis. Am J Pathol. 2007;171:19–31. doi: 10.2353/ajpath.2007.061171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tanwar PS, Zhang L, Teixeira JM. Adenomatous polyposis coli (APC) is essential for maintaining the integrity of the seminiferous epithelium. Mol Endocrinol. 2011;25:1725–39. doi: 10.1210/me.2011-0057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Boyer A, Hermo L, Paquet M, Robaire B, Boerboom D. Seminiferous tubule degeneration and infertility in mice with sustained activation of WNT/CTNNB1 signaling in sertoli cells. Biol Reprod. 2008;79:475–85. doi: 10.1095/biolreprod.108.068627. [DOI] [PubMed] [Google Scholar]
- 48.Tanwar PS, Kaneko-Tarui T, Zhang L, Rani P, Taketo MM, et al. Constitutive WNT/beta-catenin signaling in murine Sertoli cells disrupts their differentiation and ability to support spermatogenesis. Biol Reprod. 2010;82:422–32. doi: 10.1095/biolreprod.109.079335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Anthony CT, Kovacs WJ, Skinner MK. Analysis of the androgen receptor in isolated testicular cell types with a microassay that uses an affinity ligand. Endocrinology. 1989;125:2628–35. doi: 10.1210/endo-125-5-2628. [DOI] [PubMed] [Google Scholar]
- 50.Vornberger W, Prins G, Musto NA, Suarez-Quian CA. Androgen receptor distribution in rat testis: new implications for androgen regulation of spermatogenesis. Endocrinology. 1994;134:2307–16. doi: 10.1210/endo.134.5.8156934. [DOI] [PubMed] [Google Scholar]
- 51.Holdcraft RW, Braun RE. Androgen receptor function is required in Sertoli cells for the terminal differentiation of haploid spermatids. Development. 2004;131:459–67. doi: 10.1242/dev.00957. [DOI] [PubMed] [Google Scholar]
- 52.Wang RS, Yeh S, Chen LM, Lin HY, Zhang C, et al. Androgen receptor in sertoli cell is essential for germ cell nursery and junctional complex formation in mouse testes. Endocrinology. 2006;147:5624–33. doi: 10.1210/en.2006-0138. [DOI] [PubMed] [Google Scholar]
- 53.Meng J, Holdcraft RW, Shima JE, Griswold MD, Braun RE. Androgens regulate the permeability of the blood-testis barrier. Proc Natl Acad Sci U S A. 2005;102:16696–700. doi: 10.1073/pnas.0506084102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Willems A, De Gendt K, Allemeersch J, Smith LB, Welsh M, et al. Early effects of Sertoli cell-selective androgen receptor ablation on testicular gene expression. Int J Androl. 2010;33:507–17. doi: 10.1111/j.1365-2605.2009.00964.x. [DOI] [PubMed] [Google Scholar]
- 55.Kimura N, Mizokami A, Oonuma T, Sasano H, Nagura H. Immunocytochemical localization of androgen receptor with polyclonal antibody in paraffin-embedded human tissues. J Histochem Cytochem. 1993;41:671–8. doi: 10.1177/41.5.8468448. [DOI] [PubMed] [Google Scholar]
- 56.Zhou X, Kudo A, Kawakami H, Hirano H. Immunohistochemical localization of androgen receptor in mouse testicular germ cells during fetal and postnatal development. Anat Rec. 1996;245:509–18. doi: 10.1002/(SICI)1097-0185(199607)245:3<509::AID-AR7>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 57.Bremner WJ, Millar MR, Sharpe RM, Saunders PT. Immunohistochemical localization of androgen receptors in the rat testis: evidence for stage-dependent expression and regulation by androgens. Endocrinology. 1994;135:1227–34. doi: 10.1210/endo.135.3.8070367. [DOI] [PubMed] [Google Scholar]
- 58.Sar M, Lubahn DB, French FS, Wilson EM. Immunohistochemical localization of the androgen receptor in rat and human tissues. Endocrinology. 1990;127:3180–6. doi: 10.1210/endo-127-6-3180. [DOI] [PubMed] [Google Scholar]
- 59.Chang C, Chen YT, Yeh SD, Xu Q, Wang RS, et al. Infertility with defective spermatogenesis and hypotestosteronemia in male mice lacking the androgen receptor in Sertoli cells. Proc Natl Acad Sci U S A. 2004;101:6876–81. doi: 10.1073/pnas.0307306101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Willems A, Batlouni SR, Esnal A, Swinnen JV, Saunders PT, et al. Selective ablation of the androgen receptor in mouse sertoli cells affects sertoli cell maturation, barrier formation and cytoskeletal development. PloS One. 2010;5:e14168. doi: 10.1371/journal.pone.0014168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.De Gendt K, Swinnen JV, Saunders PT, Schoonjans L, Dewerchin M, et al. A Sertoli cell-selective knockout of the androgen receptor causes spermatogenic arrest in meiosis. Proc Natl Acad Sci USA. 2004;101:1327–32. doi: 10.1073/pnas.0308114100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wu RC, Jiang M, Beaudet AL, Wu MY. ARID4A and ARID4B regulate male fertility, a functional link to the AR and RB pathways. Proc Natl Acad Sci USA. 2013;110:4616–21. doi: 10.1073/pnas.1218318110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Bi J, Li Y, Sun F, Saalbach A, Klein C, et al. Basigin null mutant male mice are sterile and exhibit impaired interactions between germ cells and Sertoli cells. Dev Biol. 2013;380:145–56. doi: 10.1016/j.ydbio.2013.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Chen L, Bi J, Nakai M, Bunick D, Couse JF, et al. Expression of basigin in reproductive tissues of estrogen receptor-{alpha} or -{beta} null mice. Reproduction. 2010;139:1057–66. doi: 10.1530/REP-10-0069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Igakura T, Kadomatsu K, Kaname T, Muramatsu H, Fan QW, et al. A null mutation in basigin, an immunoglobulin superfamily member, indicates its important roles in peri-implantation development and spermatogenesis. Dev Biol. 1998;194:152–65. doi: 10.1006/dbio.1997.8819. [DOI] [PubMed] [Google Scholar]
- 66.Chen C, Ouyang W, Grigura V, Zhou Q, Carnes K, et al. ERM is required for transcriptional control of the spermatogonial stem cell niche. Nature. 2005;436:1030–4. doi: 10.1038/nature03894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Morrow CM, Hostetler CE, Griswold MD, Hofmann MC, Murphy KM, et al. ETV5 is required for continuous spermatogenesis in adult mice and may mediate blood testes barrier function and testicular immune privilege. Ann N Y Acad Sci. 2007;1120:144–51. doi: 10.1196/annals.1411.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Stoffel W, Holz B, Jenke B, Binczek E, Gunter RH, et al. Delta6-desaturase (FADS2) deficiency unveils the role of omega3- and omega6-polyunsaturated fatty acids. EMBO J. 2008;27:2281–92. doi: 10.1038/emboj.2008.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ketola I, Anttonen M, Vaskivuo T, Tapanainen JS, Toppari J, et al. Developmental expression and spermatogenic stage specificity of transcription factors GATA-1 and GATA-4 and their cofactors FOG-1 and FOG-2 in the mouse testis. Euro J Endocrinol. 2002;147:397–406. doi: 10.1530/eje.0.1470397. [DOI] [PubMed] [Google Scholar]
- 70.Oreal E, Mazaud S, Picard JY, Magre S, Carre-Eusebe D. Different patterns of anti-Mullerian hormone expression, as related to DMRT1, SF-1, WT1, GATA-4, Wnt-4, and Lhx9 expression, in the chick differentiating gonads. Dev Dyn. 2002;225:221–32. doi: 10.1002/dvdy.10153. [DOI] [PubMed] [Google Scholar]
- 71.Ketola I, Rahman N, Toppari J, Bielinska M, Porter-Tinge SB, et al. Expression and regulation of transcription factors GATA-4 and GATA-6 in developing mouse testis. Endocrinology. 1999;140:1470–80. doi: 10.1210/endo.140.3.6587. [DOI] [PubMed] [Google Scholar]
- 72.Lavoie HA, McCoy GL, Blake CA. Expression of the GATA-4 and GATA-6 transcription factors in the fetal rat gonad and in the ovary during postnatal development and pregnancy. Mol Cell Endocrinol. 2004;227:31–40. doi: 10.1016/j.mce.2004.07.016. [DOI] [PubMed] [Google Scholar]
- 73.Kyronlahti A, Euler R, Bielinska M, Schoeller EL, Moley KH, et al. GATA4 regulates Sertoli cell function and fertility in adult male mice. Mol Cell Endocrinol. 2011;333:85–95. doi: 10.1016/j.mce.2010.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Bielinska M, Seehra A, Toppari J, Heikinheimo M, Wilson DB. GATA-4 is required for sex steroidogenic cell development in the fetal mouse. Dev Dyn. 2007;236:203–13. doi: 10.1002/dvdy.21004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.McCoard SA, Wise TH, Fahrenkrug SC, Ford JJ. Temporal and spatial localization patterns of Gata4 during porcine gonadogenesis. Biol Reprod. 2001;65:366–74. doi: 10.1095/biolreprod65.2.366. [DOI] [PubMed] [Google Scholar]
- 76.Viger RS, Mertineit C, Trasler JM, Nemer M. Transcription factor GATA-4 is expressed in a sexually dimorphic pattern during mouse gonadal development and is a potent activator of the Mullerian inhibiting substance promoter. Development. 1998;125:2665–75. doi: 10.1242/dev.125.14.2665. [DOI] [PubMed] [Google Scholar]
- 77.Nalam RL, Andreu-Vieyra C, Braun RE, Akiyama H, Matzuk MM. Retinoblastoma protein plays multiple essential roles in the terminal differentiation of Sertoli cells. Mol Endocrinol. 2009;23:1900–13. doi: 10.1210/me.2009-0184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Singh AP, Cummings CA, Mishina Y, Archer TK. SOX8 regulates permeability of the blood-testes barrier that affects adult male fertility in the mouse. Biol Reprod. 2013;88:133. doi: 10.1095/biolreprod.112.107284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Schepers G, Wilson M, Wilhelm D, Koopman P. SOX8 is expressed during testis differentiation in mice and synergizes with SF1 to activate the Amh promoter in vitro. J Biol Chem. 2003;278:28101–8. doi: 10.1074/jbc.M304067200. [DOI] [PubMed] [Google Scholar]
- 80.O’Bryan MK, Takada S, Kennedy CL, Scott G, Harada S, et al. Sox8 is a critical regulator of adult Sertoli cell function and male fertility. Dev Biol. 2008;316:359–70. doi: 10.1016/j.ydbio.2008.01.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lu Q, Gore M, Zhang Q, Camenisch T, Boast S, et al. Tyro-3 family receptors are essential regulators of mammalian spermatogenesis. Nature. 1999;398:723–8. doi: 10.1038/19554. [DOI] [PubMed] [Google Scholar]
- 82.Zhang Y, Li N, Chen Q, Yan K, Liu Z, et al. Breakdown of immune homeostasis in the testis of mice lacking Tyro3, Axl and Mer receptor tyrosine kinases. Immunol Cell Biol. 2013;91:416–26. doi: 10.1038/icb.2013.22. [DOI] [PubMed] [Google Scholar]
- 83.Furuse M, Fujita K, Hiiragi T, Fujimoto K, Tsukita S. Claudin-1 and -2: novel integral membrane proteins localizing at tight junctions with no sequence similarity to occludin. J Cell Biol. 1998;141:1539–50. doi: 10.1083/jcb.141.7.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sonoda N, Furuse M, Sasaki H, Yonemura S, Katahira J, et al. Clostridium perfringens enterotoxin fragment removes specific claudins from tight junction strands: Evidence for direct involvement of claudins in tight junction barrier. J Cell Biol. 1999;147:195–204. doi: 10.1083/jcb.147.1.195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Morita K, Sasaki H, Fujimoto K, Furuse M, Tsukita S. Claudin-11/OSP-based tight junctions of myelin sheaths in brain and Sertoli cells in testis. J Cell Biol. 1999;145:579–88. doi: 10.1083/jcb.145.3.579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Fink C, Weigel R, Fink L, Wilhelm J, Kliesch S, et al. Claudin-11 is over-expressed and dislocated from the blood-testis barrier in Sertoli cells associated with testicular intraepithelial neoplasia in men. Histochem Cell Biol. 2009;131:755–64. doi: 10.1007/s00418-009-0576-2. [DOI] [PubMed] [Google Scholar]
- 87.Tsukita S, Furuse M. Pores in the wall: claudins constitute tight junction strands containing aqueous pores. J Cell Biol. 2000;149:13–6. doi: 10.1083/jcb.149.1.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Van Itallie CM, Anderson JM. Claudins and epithelial paracellular transport. Annu Rev Physiol. 2006;68:403–29. doi: 10.1146/annurev.physiol.68.040104.131404. [DOI] [PubMed] [Google Scholar]
- 89.Tsukita S, Furuse M, Itoh M. Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol. 2001;2:285–93. doi: 10.1038/35067088. [DOI] [PubMed] [Google Scholar]
- 90.Hewitt KJ, Agarwal R, Morin PJ. The claudin gene family: expression in normal and neoplastic tissues. BMC cancer. 2006;6:186. doi: 10.1186/1471-2407-6-186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Hellani A, Ji J, Mauduit C, Deschildre C, Tabone E, et al. Developmental and hormonal regulation of the expression of oligodendrocyte-specific protein/claudin 11 in mouse testis. Endocrinology. 2000;141:3012–9. doi: 10.1210/endo.141.8.7625. [DOI] [PubMed] [Google Scholar]
- 92.Shima JE, McLean DJ, McCarrey JR, Griswold MD. The murine testicular transcriptome: characterizing gene expression in the testis during the progression of spermatogenesis. Biol Reprod. 2004;71:319–30. doi: 10.1095/biolreprod.103.026880. [DOI] [PubMed] [Google Scholar]
- 93.Furuse M, Hirase T, Itoh M, Nagafuchi A, Yonemura S, et al. Occludin: a novel integral membrane protein localizing at tight junctions. J Cell Biol. 1993;123:1777–88. doi: 10.1083/jcb.123.6.1777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ando-Akatsuka Y, Saitou M, Hirase T, Kishi M, Sakakibara A, et al. Interspecies diversity of the occludin sequence: cDNA cloning of human, mouse, dog, and rat-kangaroo homologues. J Cell Biol. 1996;133:43–7. doi: 10.1083/jcb.133.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Saitou M, Ando-Akatsuka Y, Itoh M, Furuse M, Inazawa J, et al. Mammalian occludin in epithelial cells: its expression and subcellular distribution. Eur J Cell Biol. 1997;73:222–31. [PubMed] [Google Scholar]
- 96.Morrow CM, Mruk D, Cheng CY, Hess RA. Claudin and occludin expression and function in the seminiferous epithelium. Philos Trans R Soc Lond B Biol Sci. 2010;365:1679–96. doi: 10.1098/rstb.2010.0025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Cyr DG, Hermo L, Egenberger N, Mertineit C, Trasler JM, et al. Cellular immunolocalization of occludin during embryonic and postnatal development of the mouse testis and epididymis. Endocrinology. 1999;140:3815–25. doi: 10.1210/endo.140.8.6903. [DOI] [PubMed] [Google Scholar]
- 98.Gye MC. Expression of occludin in canine testis and epididymis. Reprod Domest Anim. 2004;39:43–7. doi: 10.1046/j.1439-0531.2003.00474.x. [DOI] [PubMed] [Google Scholar]
- 99.Yoon SI, Park CJ, Nah WH, Gye MC. Expression of occludin in testis and epididymis of wild rabbits, Lepus sinensis coreanus. Reprod Domest Anim. 2009;44:745–50. doi: 10.1111/j.1439-0531.2008.01064.x. [DOI] [PubMed] [Google Scholar]
- 100.Moroi S, Saitou M, Fujimoto K, Sakakibara A, Furuse M, et al. Occludin is concentrated at tight junctions of mouse/rat but not human/guinea pig Sertoli cells in testes. Am J Physiol. 1998;274:C1708–17. doi: 10.1152/ajpcell.1998.274.6.C1708. [DOI] [PubMed] [Google Scholar]
- 101.Balda MS, Matter K. Transmembrane proteins of tight junctions. Semin Cell Dev Biol. 2000;11:281–9. doi: 10.1006/scdb.2000.0177. [DOI] [PubMed] [Google Scholar]
- 102.Gonzalez-Mariscal L, Betanzos A, Avila-Flores A. MAGUK proteins: structure and role in the tight junction. Semin Cell Dev Biol. 2000;11:315–24. doi: 10.1006/scdb.2000.0178. [DOI] [PubMed] [Google Scholar]
- 103.Pelletier RM, Okawara Y, Vitale ML, Anderson JM. Differential distribution of the tight-junction-associated protein ZO-1 isoforms alpha+and alpha- in guinea pig Sertoli cells: a possible association with F-actin and G-actin. Biol Reprod. 1997;57:367–76. doi: 10.1095/biolreprod57.2.367. [DOI] [PubMed] [Google Scholar]
- 104.Byers S, Graham R, Dai HN, Hoxter B. Development of Sertoli cell junctional specializations and the distribution of the tight-junction-associated protein ZO-1 in the mouse testis. Am J Anat. 1991;191:35–47. doi: 10.1002/aja.1001910104. [DOI] [PubMed] [Google Scholar]
- 105.Fink C, Weigel R, Hembes T, Lauke-Wettwer H, Kliesch S, et al. Altered expression of ZO-1 and ZO-2 in Sertoli cells and loss of blood-testis barrier integrity in testicular carcinoma in situ. Neoplasia. 2006;8:1019–27. doi: 10.1593/neo.06559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Islas S, Vega J, Ponce L, Gonzalez-Mariscal L. Nuclear localization of the tight junction protein ZO-2 in epithelial cells. Exp Cell Res. 2002;274:138–48. doi: 10.1006/excr.2001.5457. [DOI] [PubMed] [Google Scholar]
- 107.Traweger A, Fuchs R, Krizbai IA, Weiger TM, Bauer HC, et al. The tight junction protein ZO-2 localizes to the nucleus and interacts with the heterogeneous nuclear ribonucleoprotein scaffold attachment factor-B. J Biol Chem. 2003;278:2692–700. doi: 10.1074/jbc.M206821200. [DOI] [PubMed] [Google Scholar]
- 108.Jaramillo BE, Ponce A, Moreno J, Betanzos A, Huerta M, et al. Characterization of the tight junction protein ZO-2 localized at the nucleus of epithelial cells. Exp Cell Res. 2004;297:247–58. doi: 10.1016/j.yexcr.2004.03.021. [DOI] [PubMed] [Google Scholar]
- 109.Kausalya PJ, Phua DC, Hunziker W. Association of ARVCF with zonula occludens (ZO)-1 and ZO-2: binding to PDZ-domain proteins and cell-cell adhesion regulate plasma membrane and nuclear localization of ARVCF. Mol Cell Biol. 2004;15:5503–15. doi: 10.1091/mbc.E04-04-0350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Katsuno T, Umeda K, Matsui T, Hata M, Tamura A, et al. Deficiency of zonula occludens-1 causes embryonic lethal phenotype associated with defected yolk sac angiogenesis and apoptosis of embryonic cells. Mol Cell Biol. 2008;19:2465–75. doi: 10.1091/mbc.E07-12-1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Adachi M, Inoko A, Hata M, Furuse K, Umeda K, et al. Normal establishment of epithelial tight junctions in mice and cultured cells lacking expression of ZO-3, a tight-junction MAGUK protein. Mol Cell Biol. 2006;26:9003–15. doi: 10.1128/MCB.01811-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Cambrosio Mann M, Friess AE, Stoffel MH. Blood-tissue barriers in the male reproductive tract of the dog: a morphological study using lanthanum nitrate as an electron-opaque tracer. Cell Tissues Organs. 2003;174:162–9. doi: 10.1159/000072719. [DOI] [PubMed] [Google Scholar]
- 113.Webster MR, Weeraratna AT. A Wnt-er migration: the confusing role of beta-catenin in melanoma metastasis. Sci Signal. 2013;6:pe11. doi: 10.1126/scisignal.2004114. [DOI] [PubMed] [Google Scholar]
- 114.Burgers TA, Williams BO. Regulation of Wnt/beta-catenin signaling within and from osteocytes. Bone. 2013;54:244–9. doi: 10.1016/j.bone.2013.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Lie PP, Xia W, Wang CQ, Mruk DD, Yan HH, et al. Dynamin II interacts with the cadherin- and occludin-based protein complexes at the blood-testis barrier in adult rat testes. J Endocrinol. 2006;191:571–86. doi: 10.1677/joe.1.06996. [DOI] [PubMed] [Google Scholar]
- 116.Xia W, Wong CH, Lee NP, Lee WM, Cheng CY. Disruption of Sertoli-germ cell adhesion function in the seminiferous epithelium of the rat testis can be limited to adherens junctions without affecting the blood-testis barrier integrity: an in vivo study using an androgen suppression model. J Cell Physiol. 2005;205:141–57. doi: 10.1002/jcp.20377. [DOI] [PubMed] [Google Scholar]
- 117.Lee NP, Mruk D, Lee WM, Cheng CY. Is the cadherin/catenin complex a functional unit of cell-cell actin-based adherens junctions in the rat testis? Biol Reprod. 2003;68:489–508. doi: 10.1095/biolreprod.102.005793. [DOI] [PubMed] [Google Scholar]
- 118.Lombardi AP, Royer C, Pisolato R, Cavalcanti FN, Lucas TF, et al. Physiopathological aspects of the Wnt/beta-catenin signaling pathway in the male reproductive system. Spermatogenesis. 2013;3:e23181. doi: 10.4161/spmg.23181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Liu CF, Bingham N, Parker K, Yao HH. Sex-specific roles of beta-catenin in mouse gonadal development. Hum Mol Genet. 2009;18:405–17. doi: 10.1093/hmg/ddn362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Chang H, Guillou F, Taketo MM, Behringer RR. Overactive beta-catenin signaling causes testicular sertoli cell tumor development in the mouse. Biol Reprod. 2009;81:842–9. doi: 10.1095/biolreprod.109.077446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Maatouk DM, DiNapoli L, Alvers A, Parker KL, Taketo MM, et al. Stabilization of beta-catenin in XY gonads causes male-to-female sex-reversal. Hum Mol Genet. 2008;17:2949–55. doi: 10.1093/hmg/ddn193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Wang J, Liu B, Gu S, Liang J. Effects of Wnt/beta-catenin signalling on proliferation and differentiation of apical papilla stem cells. Cell Prolif. 2012;45:121–31. doi: 10.1111/j.1365-2184.2012.00806.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Shimizu T, Kagawa T, Inoue T, Nonaka A, Takada S, et al. Stabilized beta-catenin functions through TCF/LEF proteins and the Notch/RBP-Jkappa complex to promote proliferation and suppress differentiation of neural precursor cells. Mol Cell Biol. 2008;28:7427–41. doi: 10.1128/MCB.01962-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Verhoeven G, Willems A, Denolet E, Swinnen JV, De Gendt K. Androgens and spermatogenesis: lessons from transgenic mouse models. Philos Trans R Soc Lond B Biol Sci. 2010;365:1537–56. doi: 10.1098/rstb.2009.0117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Patrao MT, Silva EJ, Avellar MC. Androgens and the male reproductive tract: an overview of classical roles and current perspectives. Arq Bras Endocrinol Metabol. 2009;53:934–45. doi: 10.1590/s0004-27302009000800006. [DOI] [PubMed] [Google Scholar]
- 126.Zhou X. Roles of androgen receptor in male and female reproduction: lessons from global and cell-specific androgen receptor knockout (ARKO) mice. J Androl. 2010;31:235–43. doi: 10.2164/jandrol.109.009266. [DOI] [PubMed] [Google Scholar]
- 127.Wang RS, Yeh S, Tzeng CR, Chang C. Androgen receptor roles in spermatogenesis and fertility: lessons from testicular cell-specific androgen receptor knockout mice. Endocr Rev. 2009;30:119–32. doi: 10.1210/er.2008-0025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Tan KA, De Gendt K, Atanassova N, Walker M, Sharpe RM, et al. The role of androgens in sertoli cell proliferation and functional maturation: studies in mice with total or Sertoli cell-selective ablation of the androgen receptor. Endocrinology. 2005;146:2674–83. doi: 10.1210/en.2004-1630. [DOI] [PubMed] [Google Scholar]
- 129.Denolet E, De Gendt K, Allemeersch J, Engelen K, Marchal K, et al. The effect of a sertoli cell-selective knockout of the androgen receptor on testicular gene expression in prepubertal mice. Mol Endocrinol. 2006;20:321–34. doi: 10.1210/me.2005-0113. [DOI] [PubMed] [Google Scholar]
- 130.Eacker SM, Shima JE, Connolly CM, Sharma M, Holdcraft RW, et al. Transcriptional profiling of androgen receptor (AR) mutants suggests instructive and permissive roles of AR signaling in germ cell development. Mol Endocrinol. 2007;21:895–907. doi: 10.1210/me.2006-0113. [DOI] [PubMed] [Google Scholar]
- 131.Aoki K, Taketo MM. Adenomatous polyposis coli (APC): a multi-functional tumor suppressor gene. J Cell Sci. 2007;120:3327–35. doi: 10.1242/jcs.03485. [DOI] [PubMed] [Google Scholar]
- 132.Kato N, Shibuya H, Fukase M, Tamura G, Motoyama T. Involvement of adenomatous polyposis coli (APC) gene in testicular yolk sac tumor of infants. Hum Pathol. 2006;37:48–53. doi: 10.1016/j.humpath.2005.09.008. [DOI] [PubMed] [Google Scholar]
- 133.Wu R, Hendrix-Lucas N, Kuick R, Zhai Y, Schwartz DR, et al. Mouse model of human ovarian endometrioid adenocarcinoma based on somatic defects in the Wnt/beta-catenin and PI3K/Pten signaling pathways. Cancer Cell. 2007;11:321–33. doi: 10.1016/j.ccr.2007.02.016. [DOI] [PubMed] [Google Scholar]
- 134.Moreno-Bueno G, Hardisson D, Sanchez C, Sarrio D, Cassia R, et al. Abnormalities of the APC/beta-catenin pathway in endometrial cancer. Oncogene. 2002;21:7981–90. doi: 10.1038/sj.onc.1205924. [DOI] [PubMed] [Google Scholar]
- 135.Defeo-Jones D, Huang PS, Jones RE, Haskell KM, Vuocolo GA, et al. Cloning of cDNAs for cellular proteins that bind to the retinoblastoma gene product. Nature. 1991;352:251–4. doi: 10.1038/352251a0. [DOI] [PubMed] [Google Scholar]
- 136.Cao J, Gao T, Stanbridge EJ, Irie R. RBP1L1, a retinoblastoma-binding protein-related gene encoding an antigenic epitope abundantly expressed in human carcinomas and normal testis. J Natl Cancer Inst. 2001;93:1159–65. doi: 10.1093/jnci/93.15.1159. [DOI] [PubMed] [Google Scholar]
- 137.Lai A, Kennedy BK, Barbie DA, Bertos NR, Yang XJ, et al. RBP1 recruits the mSIN3-histone deacetylase complex to the pocket of retinoblastoma tumor suppressor family proteins found in limited discrete regions of the nucleus at growth arrest. Mol Cell Biol. 2001;21:2918–32. doi: 10.1128/MCB.21.8.2918-2932.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Fleischer TC, Yun UJ, Ayer DE. Identification and characterization of three new components of the mSin3A corepressor complex. Mol Cell Biol. 2003;23:3456–67. doi: 10.1128/MCB.23.10.3456-3467.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Miyauchi T, Kanekura T, Yamaoka A, Ozawa M, Miyazawa S, et al. Basigin, a new, broadly distributed member of the immunoglobulin superfamily, has strong homology with both the immunoglobulin V domain and the beta-chain of major histocompatibility complex class II antigen. J Biochem. 1990;107:316–23. doi: 10.1093/oxfordjournals.jbchem.a123045. [DOI] [PubMed] [Google Scholar]
- 140.Tang W, Chang SB, Hemler ME. Links between CD147 function, glycosylation, and caveolin-1. Mol Biol Cell. 2004;15:4043–50. doi: 10.1091/mbc.E04-05-0402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Kuno N, Kadomatsu K, Fan QW, Hagihara M, Senda T, et al. Female sterility in mice lacking the basigin gene, which encodes a transmembrane glycoprotein belonging to the immunoglobulin superfamily. FEBS Lett. 1998;425:191–4. doi: 10.1016/s0014-5793(98)00213-0. [DOI] [PubMed] [Google Scholar]
- 142.Cunnane SC. Problems with essential fatty acids: time for a new paradigm? Prog Lipid Res. 2003;42:544–68. doi: 10.1016/s0163-7827(03)00038-9. [DOI] [PubMed] [Google Scholar]
- 143.Sprecher H, Luthria DL, Mohammed BS, Baykousheva SP. Reevaluation of the pathways for the biosynthesis of polyunsaturated fatty acids. J Lipid Res. 1995;36:2471–7. [PubMed] [Google Scholar]
- 144.Cho HP, Nakamura M, Clarke SD. Cloning, expression, and fatty acid regulation of the human delta-5 desaturase. J Biol Chem. 1999;274:37335–9. doi: 10.1074/jbc.274.52.37335. [DOI] [PubMed] [Google Scholar]
- 145.Viger RS, Guittot SM, Anttonen M, Wilson DB, Heikinheimo M. Role of the GATA family of transcription factors in endocrine development, function, and disease. Mol Endocrinol. 2008;22:781–98. doi: 10.1210/me.2007-0513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Nguyen DX, McCance DJ. Role of the retinoblastoma tumor suppressor protein in cellular differentiation. J Cell Biochem. 2005;94:870–9. doi: 10.1002/jcb.20375. [DOI] [PubMed] [Google Scholar]
- 147.Lipinski MM, Jacks T. The retinoblastoma gene family in differentiation and development. Oncogene. 1999;18:7873–82. doi: 10.1038/sj.onc.1203244. [DOI] [PubMed] [Google Scholar]
- 148.Foster JW, Dominguez-Steglich MA, Guioli S, Kwok C, Weller PA, et al. Campomelic dysplasia and autosomal sex reversal caused by mutations in an SRY-related gene. Nature. 1994;372:525–30. doi: 10.1038/372525a0. [DOI] [PubMed] [Google Scholar]
- 149.Wagner T, Wirth J, Meyer J, Zabel B, Held M, et al. Autosomal sex reversal and campomelic dysplasia are caused by mutations in and around the SRY-related gene SOX9. Cell. 1994;79:1111–20. doi: 10.1016/0092-8674(94)90041-8. [DOI] [PubMed] [Google Scholar]
- 150.Vidal VP, Chaboissier MC, de Rooij DG, Schedl A. Sox9 induces testis development in XX transgenic mice. Nat Genet. 2001;28:216–7. doi: 10.1038/90046. [DOI] [PubMed] [Google Scholar]
- 151.Schepers GE, Bullejos M, Hosking BM, Koopman P. Cloning and characterisation of the Sry-related transcription factor gene Sox8. Nucleic Acids Res. 2000;28:1473–80. doi: 10.1093/nar/28.6.1473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Linger RM, Keating AK, Earp HS, Graham DK. TAM receptor tyrosine kinases: biologic functions, signaling, and potential therapeutic targeting in human cancer. Adv Cancer Res. 2008;100:35–83. doi: 10.1016/S0065-230X(08)00002-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Tang H, Chen S, Wang H, Wu H, Lu Q, et al. TAM receptors and the regulation of erythropoiesis in mice. Haematologica. 2009;94:326–34. doi: 10.3324/haematol.13635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Verma A, Warner SL, Vankayalapati H, Bearss DJ, Sharma S. Targeting Axl and Mer kinases in cancer. Mol Cancer Ther. 2011;10:1763–73. doi: 10.1158/1535-7163.MCT-11-0116. [DOI] [PubMed] [Google Scholar]
- 155.Nieschlag E, Behre H, Nieschlag S, Van Ahlen H. Andrology. Male reproductive health and dysfunction. Internistische Praxis. 2011;51:751. [Google Scholar]
- 156.Dada R, Kumar M, Jesudasan R, Fernandez JL, Gosalvez J, et al. Epigenetics and its role in male infertility. J Assist Reprod Genet. 2012;29:213–23. doi: 10.1007/s10815-012-9715-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Singh K, Jaiswal D. Human male infertility: a complex multifactorial phenotype. Reprod Sci. 2011;18:418–25. doi: 10.1177/1933719111398148. [DOI] [PubMed] [Google Scholar]
- 158.Skakkebaek NE, Rajpert-De Meyts E, Main KM. Testicular dysgenesis syndrome: an increasingly common developmental disorder with environmental aspects. Hum Reprod. 2001;16:972–8. doi: 10.1093/humrep/16.5.972. [DOI] [PubMed] [Google Scholar]
- 159.Pointis G, Gilleron J, Carette D, Segretain D. Testicular connexin 43, a precocious molecular target for the effect of environmental toxicants on male fertility. Spermatogenesis. 2011;1:303–17. doi: 10.4161/spmg.1.4.18392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Gilleron J, Malassine A, Carette D, Segretain D, Pointis G. Chemical connexin impairment in the developing gonad associated with offspring infertility. Curr Med Chem. 2011;18:5145–58. doi: 10.2174/092986711797636117. [DOI] [PubMed] [Google Scholar]
- 161.Defamie N, Berthaut I, Mograbi B, Chevallier D, Dadoune JP, et al. Impaired gap junction connexin43 in Sertoli cells of patients with secretory azoospermia: a marker of undifferentiated Sertoli cells. Lab Invest. 2003;83:449–56. doi: 10.1097/01.lab.0000059928.82702.6d. [DOI] [PubMed] [Google Scholar]
- 162.Matsuo Y, Nomata K, Eguchi J, Aoki D, Hayashi T, et al. Immunohistochemical analysis of connexin43 expression in infertile human testes. Acta Histochem Cytochem. 2007;40:69–75. doi: 10.1267/ahc.07001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Steger K, Tetens F, Bergmann M. Expression of connexin 43 in human testis. Histochem Cell Biol. 1999;112:215–20. doi: 10.1007/s004180050409. [DOI] [PubMed] [Google Scholar]