

Abstract
Objective
Seizures are currently defined by their electrographic features. However, neuronal networks are intrinsically dependent upon neurotransmitters of which little is known regarding their peri-ictal dynamics. Evidence supports adenosine as having a prominent role in seizure termination, as its administration can terminate and reduce seizures in animal models. Further, microdialysis studies in humans suggest adenosine is elevated peri-ictally, but the relationship to the seizure is obscured by its temporal measurement limitations. Because electrochemical techniques can provide vastly superior temporal resolution, we test the hypothesis that extracellular adenosine concentrations rise during seizure termination in an animal model and humans using electrochemistry.
Methods
White farm swine (n=45) were used in an acute cortical model of epilepsy and 10 human epilepsy patients were studied during intraoperative electrocorticography (Ecog). Wireless Instantaneous Neurotransmitter Concentration Sensor (WINCS) based fast scan cyclic voltametry (FSCV) and fixed potential amperometry were obtained utilizing an adenosine specific triangular waveform or biosensors respectively.
Results
Simultaneous Ecog and electrochemistry demonstrated an average adenosine rise of 260% compared to baseline at 7.5 ± 16.9 seconds with amperometry (n=75 events) and 2.6 ± 11.2 seconds with FSCV (n=15 events) prior to electrographic seizure termination. In agreement with these animal data, adenosine elevation prior to seizure termination in a human patient utilizing FSCV was also seen.
Significance
Simultaneous Ecog and electrochemical recording supports the hypothesis that adenosine rises prior to seizure termination, suggesting that adenosine itself may be responsible for seizure termination. Future work using intraoperative WINCS based FSCV recording may help to elucidate the precise relationship between adenosine and seizure termination.
Keywords: Electrochemistry, Fast Scan Cyclic Voltammetry, Amperometry, Purine, Adenosine, Seizure, Epilepsy, Large Animal Model, Human Trial
Introduction
Adenosine has a long history in epilepsy research.1 It is an endogenous inhibitor of excitatory synaptic transmission with potent anticonvulsant properties in the mammalian brain1. It appears to facilitate neuroprotection against ischemia and seizures and to be essential for natural seizure termination.1–4 When applied exogenously, it has been reported to terminate seizures in virtually all animal models in which it has been tested.1, 4–15 In humans, it likely plays a prominent role, along with GABA, in seizure regulation, as measured by electroencephalography (EEG).10 Due to its fast metabolism, adenosine is quickly and tightly regulated, which makes it a promising therapeutic intervention for epilepsy.1–4 However, despite evidence that adenosine suppresses epileptic activity, its peri-ictal dynamics are not known. This knowledge gap may be attributed in large part to difficulty in measuring adenosine, which has a very short, 8–15 second half-life in humans.16
Microdialysis, a method in which aliquots of effluent are collected from tissue, has been used to measure adenosine in relation to seizures. During and Spencer, for example, used it to measure peri-ictal adenosine changes in humans and found increases of 6–31 fold three minutes after seizures.10 However, microdialysis is slow and constrained by a need for at least 2 minutes to collect an individual aliquot. Whereas epileptiform events occur over tens of seconds, microdialysis simply can not explore the peri-ictal dynamics of adenosine.10 Winn et al. used a freeze-blow technique in which animal brains are quickly frozen prior to analyte measurement, and found evidence of peri-ictal adenosine increase.17 However this technique requires immediate sacrifice of the subject and permits assessment only of the total tissue content of adenosine, not of active extracellular concentrations alone.
Two alternative techniques, fast scan cyclic voltammetry (FSCV) and fixed potential amperometry (FPA), offer a viable and technically superior means of measuring changes in extracellular adenosine in vivo during peri-ictal events. Both are electrochemical techniques that have the sub-second time resolution needed to record adenosine peri-ictally. An additional benefit of these techniques is their sub-millimeter spatial resolution. They typically sample radially from 250 to 50 μm, a much more narrowly defined area than that sampled by microdialysis, which perfuses a relatively large area. Given the short half-life (<15 seconds) of adenosine, and because it has been shown to be electrochemically active, both FPA and FSCV appear to be ideal means of measuring adenosine release during seizure activity in animal models.18 Indeed, previous research in small animal models has shown that both techniques provide complimentary information.19–21
Although both techniques record electrochemical activity, there are important differences between them. FSCV relies on Faradic principles and measures analytic substrates at an implanted carbon fiber microelectrode (CFM) by detecting the oxidation and reduction currents of molecules. FPA uses carbon-based microelectrodes to detect the current associated with electroactive compounds. They also differ in their safety for human use. The components of the working and reference electrodes in FSCV are considered safe, whereas those for FPA are not. The enzymatic preparations in the adenosine FPA biosensors contain Nafion, which is not approved for humans by the U.S. Food and Drug Administration. FPA biosensors also contain bovine proteins which are unable to be sterilized for surgical use in humans.
We chose to use both methods to test the hypothesis that adenosine release is elevated in a large animal model during epileptiform events with the goal of testing that same hypothesis in humans. For our FSCV recordings we used an in-house designed wireless instantaneous neurochemical sensing system (WINCS). Here, we demonstrate the effective measurement by FSCV and FPA of cortical extracellular adenosine release during electrographic events induced by acute chemoconvulsant injection in swine.
Having found that both techniques were effective in measuring alterations in adenosine in a large animal model, we chose FSCV to evaluate human subjects undergoing surgery for partial epilepsy. Applied in a small group of human subjects, we demonstrate the safety and potential clinical utility of FSCV for real-time neurochemical monitoring. The findings suggest that this technique could advance understanding of the mechanisms of seizure onset and help define the boundaries of the seizure onset zone in humans with epilepsy.
Methods
Animal Recordings
All basic research studies had prior approval of the Mayo Clinic Institutional Animal Care and Use Committee. Studies were performed on 45 castrated male domestic swine (Large-white/Landrace/Duroc cross), each weighing between 30–35kg.
Surgery
Pig surgery was performed in a manner that conformed as closely as possible to the conditions in the human operative theater, as previously described.22 Epileptiform activity was induced by subcortical injection 4–5 mm below the cortical surface of 5 μl Benzyl-penicillin (PCN).22 This simple protocol reliably produces initial effective spikes as previously described, followed by runs of spikes greater than 3.5 Hz (Figure 1).
Figure 1.
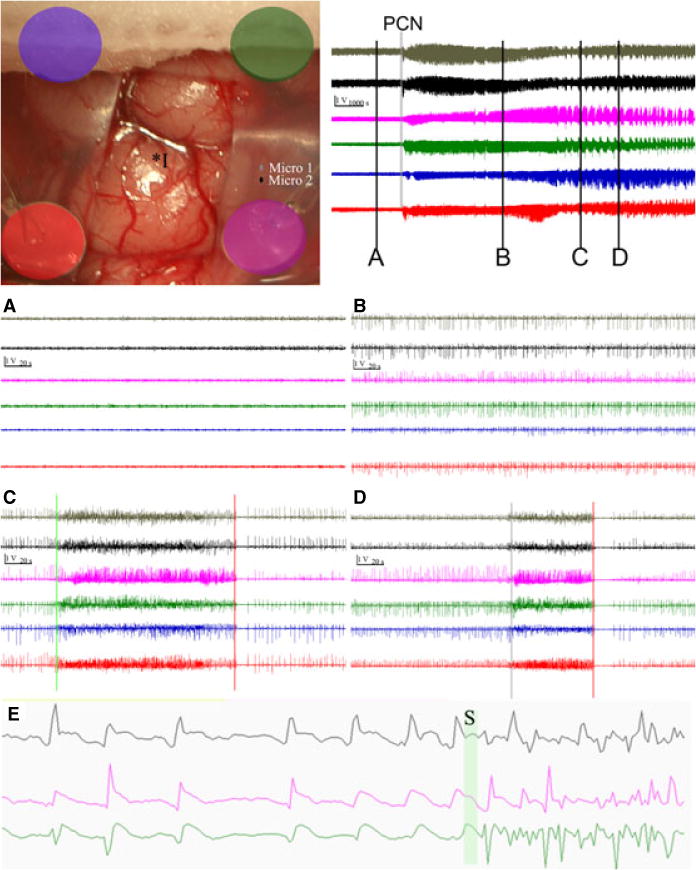
upper left: Intraoperative picture of grid implantation and electrode setup. Color coded electrodes correspond to channels in attached EEG data. Position of PCN injection (I*) and microwire EEG channels (micro 1 and 2) are also shown. Upper right top: overall time course after injection (vertical bar is 1 volt, horizontal bar is 1000 seconds), bars with letter labels correspond to time for panels with appropriate letter below (vertical bar is 1 volt, horizontal is 20 seconds): grey bar labeled by PCN is the PCN injection. (A) grade 1 activity, EEG prior to injection of 5500 U of PCN (B) grade 2 activity, spiking seen for first 1–2 hours after PCN injection (C, D) examples of electrographic events (grade 3–4 activity) used to evaluate electrochemical data. Note the clear transition to synchronized activity greater than 3.5 Hz (green bars) and the most reliable time point for each event, the clear seizure stoppage with prolonged electrographic silence (red bars). (E) 10 seconds of EEG data from 1 microwire electrode (top) and two macrocontacts (bottom two) at increased magnification demonstrates the spike waveform. The green bar (S) represents a transition into an event.
Histology
All animals were sacrificed at the end of the procedure. Brains were removed, fixed in 10% formalin and examined after sectioning to determine the exact location of the working electrode position (e.g., the FSCV carbon fiber microelectrode or amperometry biosensor). All recording electrodes were implanted at a depth of 3–7 mm with the implantation goal of 4–5 mm. Only animals with accurate electrode or biosensor positioning were considered for data analysis.
Amperometry
Amperometry for ADO, Hx, ATP, and control sensor (without enzymes) was performed with commercial Pinnacle Biosensors (biosensor 7001, integrated with reference, Pinnacle Technology INC. Lawrence, KA). These sensors are fabricated in collaboration with Sarissa (Sarissa-Biomedical Ltd, Coventry, UK). The sensor coatings are provided by Sarissa on the Pinnacle hardware, and handling and use of these sensors was performed as suggested by Sarissa-Biomedical, Ltd. The signal was digitized on an 8 channel recording system (e-corder 821,eDAQ Plt Ltd, Denistone East, Australia) and recorded on a Dell Latitude E6400 with software provided with the device (eDAQ Chart, v5.5.8). Multichannel amperometry was driven by the EP352 Biosensor Isopod (eDAQ) for each sensor channel (up to 3). Prior to their use, amperometry biosensors were stored at 4° C. Rehydration was performed as described by Sarrisa-Biomedical, Ltd, with the proximal 3 mm submersed in Buffer A (2 mM Sodium Phosphate buffer, pH 7.4, 100 mM NaCl, 1 mM MgCl2, and 2 mM glycerol) for at least 10 minutes. From that point on, biosensors were used only once, handled in congruence with manufacturer instructions, and were not allowed to re-dry. Calibration protocol can be reviewed in the supplemental material.
In vivo implantation was performed immediately after calibration. Implantations were performed by microdriver measurement to a 4–5 mm depth in the cortex. The sensors were polarized at +500 mV. The biosensors were then allowed to asymptote for 15 minutes. In all cases, an analyte specific biosensor was placed with a null sensor in order to perform differential measurements. One hour of recording was taken after stabilization. Data analysis were performed by importing Edaq Chart data in .txt format to Spike2 6.11. The following time points were chosen for analysis: the onset of the epileptiform event was defined as confluent spiking greater than 3.5 Hz. The termination of the epileptiform event was defined as a pause without spiking of greater than 3 second at the end of the event. Adenosine onset time was defined as the low point (lowest current (nA) signal) of the adenosine signal during seizure, and the peak of adenosine response (high point of the current (nA) signal) was also recorded. These time points are shown and marked as seen in Figure 2.
Figure 2. Fixed potential amperometry (FPA) analysis of adenosine release during epileptiform events.
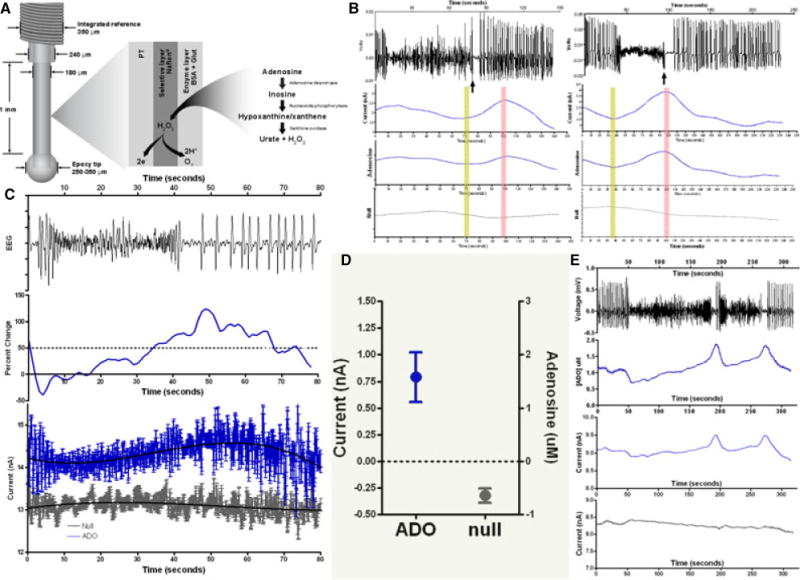
Amperometric (FPA) analysis of acute seizure model in swine: (A) schematic drawing of the dimensions and enzymatic setup of the biosensor used to detect adenosine; (B) two separate examples of seizures (EEG top) (note there is little variation in the raw null signal (bottom grey), but the raw (adenosine) and differential (adenosine-conrtrol (null)) signal demonstrate an increase in relative adenosine). The rising adenosine signal (yellow bar) and peak (pink bar) are noted; (C) averaged adenosine response for 5 seizures: top represents EEG signal, middle graph represents the normalized response of differential adenosine recordings (intervening pink bars represent mean + SEM (error bars) of the onset of adenosine increase and peak signal relative to seizure pause); bottom represents mean ± SD of the raw signal with blue the adenosine, grey control (null), and black lines the mean polynomial fit for this data; (D) 95% confidence interval for average peak values of adenosine (left y-axis nA, right is uM) and null. As expected there is a decrease in signal across time in the control (null) (grey), but also a marked increase in the adenosine concentration; (E) example of one event in which there were two pauses, each of which is associated with a unique adenosine increase.
In Vitro FSCV
FSCV was performed acquiring raw cyclic voltammograms using with WINCS (Wireless Instantaneous Neurochemical Sensor) as previously described, and analyzed by WincsWare (Mayo Clinic, Rochester, MN, USA).18, 23–27 For FSCV measurements of ADO, a triangular waveform was used with a voltage ramp from −0.4 to 1.5 V and back to −0.4 V at a scan rate of 400 V s−1 repeated at 10 Hz with a resting potential of −0.4 V between scans as described by Venton.19, 20 In all experiments the background current was allowed to stabilize for at least 15 min. All potentials were reported against an Ag/AgCl reference electrode in a grounded Faraday cage. Experiments utilized a carbon-fiber microelctrode (CFM) which was constructed as previously described.26
FSCV and EEG time-locked data
In order to establish the exact temporal relationship of the EEG data and FSCV signal, we altered our original software (internal programming) to send a synchchronized pulse of 6 V for 1 ms during every 5 seconds of acquisition and a double pulse for every 10 pulses from the WINCS base unit to the neuralynx EEG recorder. FSCV data were then converted to current-by-time output at 1.45V and 1.0V. These data were then synchchronized to the EEG data within an estimated accuracy of 1/100 of a second. Three consecutive swine were used to evaluate the relationship between the EEG data and WINCS-based FSCV at seizure onset and seizure termination, defined as an isoelectric signal with greater than 3 second pause. Time points for rise, peak, and return to baseline (offset) of the FSCV signal were identified. We used 5 consecutive epileptiform events per animal for a total of 15 events.
Computer analysis
Offline EEG data processing was performed utilizing matlab r2009b with mef2 formatting.28 Processing of FSCV and amperometry data was conducted with conversion to txt files and analysis in spike 2 version 6.11 (Cambridge Electronic Design, Cambridge, UK). Statistical procedures in the software package Graph Pad Prism (Graph Pad Software, Inc., San Diego, USA) as well as JMP 8.0 (2008, SAS Institute Inc., Cary, NC, USA) were used.
Statistical Tests
For ADO amperometry 75 events (evenly collected amoungst animal recordings) were recorded, meeting the central limit theorem, and t-test analysis was used. For FSCV ADO and Hx/ADO Amperometry (n=15 and n=12 respectively), Kruskal-Wallis tests were used for 3 group analysis (i.e. Null, ADO, Hx) and Wilcoxon matched pairs test was utilized as well. Statistical procedures in the software package Graph Pad Prism (Graph Pad Software, Inc., San Diego, USA) as well as JMP 8.0 (2008, SAS Institute Inc., Cary, NC, USA) were used.
Human Recordings
Prior to enrollment, this study was approved by our Mayo Clinic (Rochester) Institutional Review Board and neurosurgical/neurology departmental review boards. Participants included ten patients with medically intractable temporal lobe epilepsy who had been scheduled for an anterior temporal lobectomy. These 10 patients underwent electrocorticography (Ecog), which is the standard procedure to determine the extent of neocortical resection. In addition to the standard neurosurgical elements, these included a sterilized WINCS unit, two separate sets of three working electrodes, and a base station computer. After appropriate positioning, a standard craniotomy for anterior temporal lobectomy was performed. Electrocorticography was performed as previously described.29 A separate FSCV electrode was placed 5 μm into the cortical surface. Recordings where then performed for 10 minute periods. Additional details of the consent and implantation process can be found in supplemental materials.
Results
Profile of epileptiform activity induced by penicillin injection in the swine model
Penicillin (PCN) injection reliably induced epileptiform events and spiking in the pig cortex as previously described.22 Figure 1 demonstrates a typical macro and micro-cortical electrode implantation. PCN injection-induced spiking occurs within three minutes and within one to two hours progresses to runs of spiking events focal to the PCN injection site (Figure 1)22. These epileptiform events conform to induced clinical convulsions in other animal models. Although different in important ways from the chronic seizures that arise from complex epileptic foci in humans, these induced convulsions are referred to as seizures in the animal literature. The first such epileptiform events among 23 pigs (for a total of ninety epileptiform events) were selected for correlation analysis to assess their relationship to the electrochemical findings. The mean ± SD seizure duration was 55.2 ± 31.9 seconds (95% CI 62.0 to 48.6).
Amperometric (FPA) measurement of Adenosine during epileptiform events
Pig epileptiform events were analyzed using adenosine differential FPA (co-monitored with EEG). A total of 20 pigs and 75 random chosen seizure events were analyzed with this technique (Figure 2). Notably, there was minimal variation in the recorded adenosine signals when FPA was performed without PCN injection. However, during grade IV seizure events, there was a one-to-one statistical correlation between the electrographic event and rise in adenosine peri-ictally (Figure 2). For this reason, the rises and peaks of adenosine were recorded relative to the most reliable time period in each epileptiform event, which was the post-ictal pause in spiking.
The rise in the adenosine signal was perfectly correlated with epileptiform event termination and had a mean onset at 7.5 ± 16.9 seconds (95% CI −11.4 to −3.6 seconds) prior to seizure termination (Figure 2), with a median rise point at −1 second and a range of −57 seconds to 31 seconds over 75 events. The peak adenosine signal occurred after the seizure termination with a mean of 19.5 ± 10.6 seconds (95% CI 17.1 to 21.9 seconds). The mean increase in adenosine concentration was 649 ± 513 nM (95% CI 505 to 794 nM) (Figure 2). The range of peri-ictal increase was 100 [nM] to 2200 [nM] of Adenosine. There were no significant correlations between the rise of adenosine and the duration of epileptiform event (r=0.06), time to adenosine peak (r=−0.11), or increase in the concentration of adenosine (r=0.3).
FSCV characterization of purines
To determine the background current at a given time, adenosine biosensors used in conjunction with FPA must be referenced to a null electrode. Contemporary adenosine biosensors use a sequence of reactions as demonstrated in Figure 2a. Reference biosensors typically consist of an electrode coated with Nafion and bonding agents, but inosine biosensors (lacking only the adenosine deaminase) have been also been used.30, 31 While this is a clever solution, a critical problem remains. Adenosine is a substrate for nucleoside phosphorylase, which has a high affinity for adenosine.32, 33 Although this high affinity does not affect the adenosine signal from the adenosine biosensor, it does make referential recordings uncertain until proper kinetic studies are performed. For this reason, to characterize purines, we used FSCV, an electrochemical technique capable of identifying electroactive adenosine without this potential confound.19–21 Supplemental Figure 1 demonstrates the plethora of potential molecules that may interfere in the signaling cascade present on the FPA adenosine biosensor. Notably, only adenosine, guanosine, and hypoxanthine (Hx) have electroactivity at 5 μM (SF1). Fortunately, using FSCV guanosine was reliably distinguished from adenosine by the subtracted unfolded voltammogram (SF 1B). Hx could interfere, but only if present in logarithmically higher concentrations than adenosine, which it was not.
FSCV measurement of adenosine during epileptiform events
Fifteen epileptiform events in 3 pigs were analyzed to assess their concordance with the FPA data. Figure 3 demonstrates an example of a seizure in relation to the synchronized FSCV signal. Here, there is a clear increase in adenosine associated with seizure termination (Figure 3). Analysis of these events showed Peak 1 of ADO at 1.45V. In those events with significant increases in Peak 1, Peak 2 can be seen at 1.0 V (Figure 3b), these two peaks are considered the neurochemical signature of ADO.18 Consistent with our previous FPA data, the rise in the adenosine signal had a mean increase at 2.6 ± 11.2 seconds (95% CI −8.8 to 3.5 seconds) prior to seizure termination. The peak adenosine signal typically occurred after seizure termination at a mean of 26.8 ± 16.5 seconds (95% CI 16.6 to 34.9 seconds) which is concordant with our FPA data.
Figure 3. Fast scan cyclic voltammetry (FSCV) analysis of epileptiform events.
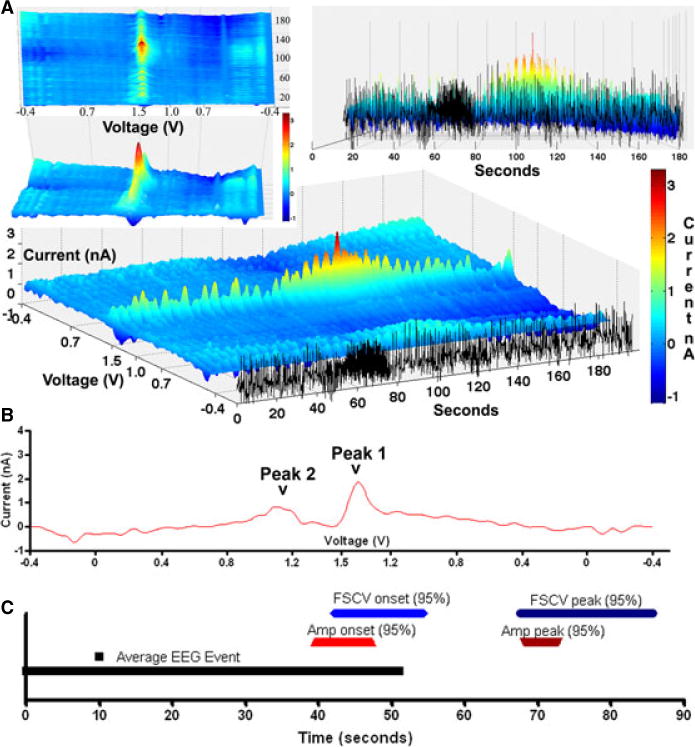
Time-locked FSCV and EEG: (A) Example of a epileptiform event evaluated by FSCV synchronized to EEG data with 3D mesh plots of the continuous data with the x axis representing seconds, the black tracing set apart, representing ne channel of EEG data, the y axis representing the voltage (V) of voltammogram scans starting at −0.4 to 1.5 (mid-y axis) to −0.4; and the z-axis representing the current (nA). Note after termination of the terminates, there is an elevation in the current at 1.5 V; (B) unfolded voltammogram demonstrating the unique Peak 1 (1.5V) and Peak 2 (1.0V) increases consistent with adenosine; (C) timeline showing average length of EEG event (black line), 95% CI of the onset and peaks of FSCV (blue) (n=15) and a comparative technique, amperometry (red) (n=75).
Adenosine and hypoxanthine resolved through amperometry
Although FSCV was capable of distinguishing most of the adenosine metabolites from adenosine (SF1), there appeared to be some overlap between the oxidation peaks of hypoxanthine (Hx) and adenosine. To determine whether Hx contributed significantly to the peri-ictal rise in adenosine, we performed concomitant FPA recordings of adenosine, Hx, and a null prepared electrode in 3 pigs. We analyzed 4 consecutive epileptiform events in each animal, for a total o f 12 events (Figure 4).
Figure 4. Biosensor amperometric separation of adenosine and hypoxanthene (Hx) during epileptiform events.
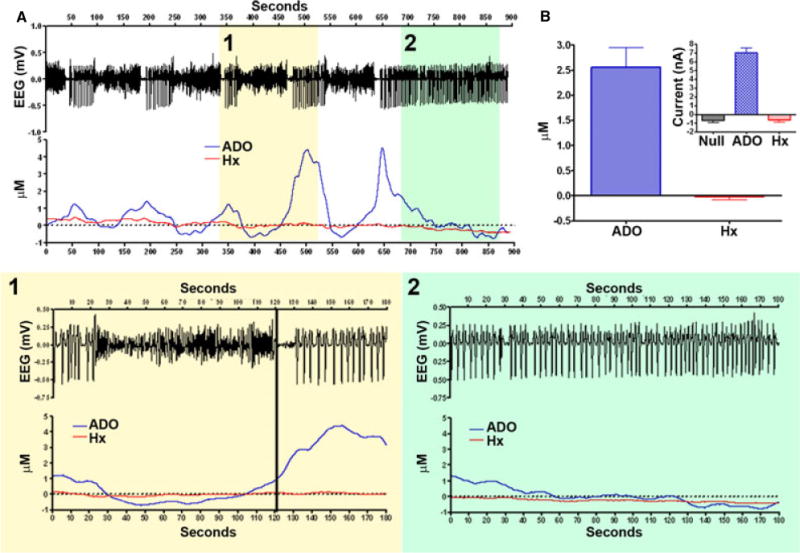
(A) Continuous fixed potential amperometry (FPA) recording over 900 seconds demonstrating 5 epileptiform events in which (1) represents a higher magnification of such an event, and (2) shows continuous spiking at the end. Note there is little variation in Hx, but adenosine consistently varies with the epileptiform activity; (B) adenosine and Hx micromolar change with event termination, 95% CI for adenosine is 1.7 to 3.4 μM and for Hx is −0.2 to 0.1. Inset shows variation in raw current (nA) of the FPA recordings. Note that there is a general decrement with null and Hx as is typical with amperometry.
The raw FPA average of control, adenosine, and Hx responses at the termination of seizure events were as follows: null −0.7±0.5 nA; adenosine 7.0±1.2 nA; and Hx −0.6±0.7 nA (95% CI) (null: −1.4 to 0 nA, adenosine: 5.6 to 8.5 nA, Hx: −1.4 to 0.2nA). There was a significant difference in signal strength between adenosine and Hx, and adenosine and control, but not between Hx and control (p=0.0089, Kruskal-Wallis) (Figure 4).
A comparison of the averaged normalized adenosine and Hx responses at the termination of seizure events revealed the following: adenosine: 2.6±1.4, Hx: 0±0.2 nA (95% CI); adenosine: 1.7 to 3.4; and Hx: −0.2 to 0.1 nA. There was a significant difference between adenosine and Hx (p=0.0005, Wilcoxon matched pairs). Thus, although microdialysis studies have found a more prolonged increase of Hx relative to a single epileptiform event, our results show no significant change in Hx above baseline and that increased Hx reaches the level of adenosine elevation. These results suggest that adenosine alone may be important.5
Human FSCV
Having successfully recorded adenosine in our swine model, we sought to assess this technology in human patients undergoing surgery for epilepsy. At the time of this study, FSCV had not yet been conducted in human epilepsy subjects so our initial trials focused on safety.34 Ten epilepsy patients consented to participate in the study (Table 1). A CFM was implanted into the temporal lobe of each patient for 15 minutes during intraoperative Ecog prior to resection. It is estimated that 10% of temporal lobe epilepsy patients will have lateral temporal lobe seizure activity and that in the other 90% seizure activity will be generated from mesial temporal structures.29 Thus, we anticipated that of the 10 patients in the safety study, at least one would have lateral neocortical seizure activity. Our expectation was met. Patient #2 had lateral neocortical seizure activity, which occurred with FSCV recording and showed a Peak 1 indicative of adenosine (Figure 5). The remaining nine patients had stable neocortical recordings at Peak 1 and did not have lateral neocortical events.
Table 1.
Clinical FSCV intraoperative implantations during intraoperative ECoG
| Patient no. | Side | Age | Sex | EEG activity
|
FSCV change | Complications | |
|---|---|---|---|---|---|---|---|
| Mesial | Lateral | ||||||
| 1 | Left | 53 | M | + | − | − | None |
| 2 | Right | 55 | F | + | + | + | None |
| 3 | Right | 33 | M | + | − | − | None |
| 4 | Right | 39 | M | + | − | − | None |
| 5 | Left | 21 | M | + | − | − | None |
| 6 | Left | 53 | F | + | − | − | None |
| 7 | Right | 22 | F | + | − | − | None |
| 8 | Right | 40 | F | + | − | − | None |
| 9 | Left | 54 | F | + | − | − | None |
| 10 | Left | 55 | M | + | − | − | None |
Figure 5. Human Intraoperative Fast Scan Cyclic Voltammetry (FSCV) Recording.
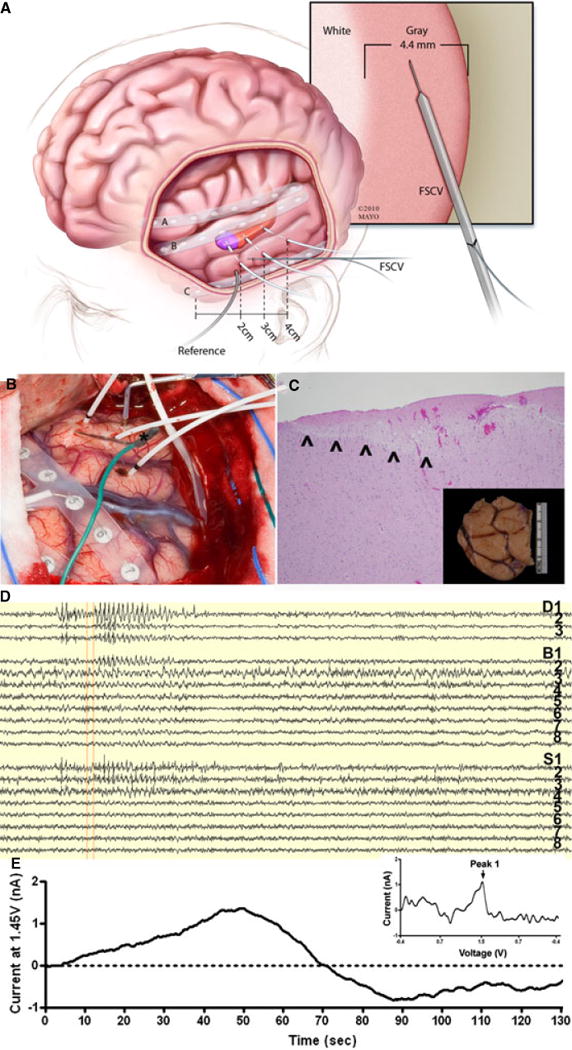
Representative intraoperative human FSCV recorded by the wireless instantaneous neurochemical concentration sensing system (WINCS): (A) a schematic demonstrating the human intraoperative implantation, experimental setup, illustrating the implantation depth electrophysiology electrodes and glass-insulated CFM in the gray matter of temporal cortex targeted for resection for seizure control; (B) intraoperative photo of human surgery utilizing FSCV. A glass capillary carbon fiber microelectrode (*green wire) is delivered into the lateral neocortex near the electrocorticography electrode S3 during pre-lobectomy electrocorticography. FSCV is performed relative to a stainless steel electrode. There is an eight contact strip over the inferior temporal gyrus (right of exposure), an eight contact strip over the superior temporal gyrus (most visible in center of exposure) and a separate strip is over the interior frontal lobe. There are 3 mesial temporal depths in the exposure (white leads) penetrating the middle temporal gyrus; (C) 10x hematoxylin and eosin stain of electrode insertion site on pathology. Note the entire FSCV electrode tract is within grey matter (inset shows the gross anterior temporal lobe specimen with methylene blue at the insertion site); (D) 140 seconds of electrocorticography demonstrating spontaneous epileptiform activity with that from 7 to 30 seconds most prominent in the mesial temporal lobe (D1–3) with associated lateral neocortical activity (S1–3). (E) Current (at 1.45 V or Peak 1 of adenosine) vs. time plot corresponding in time to the above (D) EEG. Note there appears to be an increase in Peak 1 of adenosine with the peak near the termination of this epileptiform event in patient 2. Inset: unfolded voltammogram of the signal at 53 seconds, when scanning from −0.4 to 1.5, showing a unique oxidation peak at 1.45 Volts.
Discussion
This study demonstrates an integrated effort to establish safety and importance of electrochemical techniques for detecting neurochemical changes during human epilepsy surgery. Firstly, utilizing a model we developed for translational development of intraoperative EEG monitoring, we demonstrate that it is possible to conduct neurochemical measurement during such events to determine the relationship of adenosine to seizure onset and termination in both a swine model and during intraoperative epilepsy surgery in humans. Our findings, utilizing a novel form of electrochemical detection, demonstrate the peri-ictal dynamics in alive and intact animals of adenosine which support its role in seizure termination.
To establish equivalency between FSCV and FPA in the cortical measurement of extracellular adenosine release during epileptiform events, we conducted FSCV with a free-floating CFM implanted in the pig cortex. We found that extracellular ADO increases as measured by both FSCV and FPA just prior to the termination of epileptiform events. Further enzyme-based FPA studies were conducted to confirm that FSCV was recording ADO alone and not Hx.
Our FPA and FSCV findings indicate that extracellular levels of ADO increase just prior to the termination of acute epileptiform events and attain peak values following seizure termination. In this acute chemoconvulsant swine model, the second-to-second rise in ADO levels during seizure activity and the peak values immediately following seizure events were predictive of the termination of epileptiform events. Considering that PCN is a GABAa receptor inhibitor, these studies provide evidence that when the GABAa is inhibited, that ADO participates in epileptiform event termination.
As noted in the introduction, because of its inadequate time resolution, microdialysis can not measure ADO relative to seizure activity.35 Therefore, it has become increasingly more common for researchers to utilize electrochemistry to measure ADO in in vitro preparations.36 For example, Etherington et al., used a slice model of epileptiform activity induced by high frequency stimulation in magnesium-free conditions and found that ADO increases at or after seizure termination.30, 31 The order of ADO increase they observed was 1 to 5 μM, which agrees with our data (Figure 4). While their study corroborates the validity of electrochemistry for analyzing epileptiform activity, our in vivo experiment takes this technique a step closer to clinical utility.37 Like Etherington et al., we used FPA to assess seizure levels of purines and FSCV to further characterize the signal.30, 31
Having established the utility of FSCV coupled with a free-floating CFM to measure ADO release during seizure events in the swine model, we tested the procedure in ten human patients with intractable partial epilepsy. We expected one of these 10 patients to have a lateral neocortical seizure onset zone.29 In the single patient with lateral neocortical seizures in our series, an ADO signal was observed, which exhibited a time course and pattern similar to that seen during seizure events in our pig model. In the remaining 9 patients there was no such change, which was not surprising, given the expectation again that only 10% of these consecutively selected patients would have neocortical epilepsy.29
Our findings also provide support for ADO as a natural anti-epileptic and describe its peri-ictal dynamics. Adenosine antagonists have been capable of inducing epileptiform activity in animals,38, 39 and in a variety of animal models, adenosine treatment has resulted seizure reduction.1, 4–12, 14, 15 Unfortunately, its therapeutic usefulness as an antiepileptic is limited by the fact that systemic administration of adenosine induces cardiac asytole and by the fact that at present there is no currently available effective local delivery system.2, 3
Our human study was designed to demonstrate the safety of FSCV recording during epilepsy surgery. This aim is of clinical importance, given that introducing enzymatic elements and foreign proteins, such as bovine proteins into the human brain through an FPA biosensor makes amperometry unsafe for human use. The fact that there were no complications or post-surgical infection in these initial ten patients supports the possibility of expanding the protocol to include mesial temporal or extratemporal FSCV monitoring. Clearly, a larger cohort is needed to reach conclusions about the clinical utility and further safety of intraoperative FSCV monitoring. We are now working on the design of a human biosensor probe, which could be implanted into mesial (versus lateral) temporal structures, and are designing more reliable electrodes to perform multiple array measurements.
Our overall goal in utilizing neurochemical monitoring in epilepsy surgery is to provide an additional means of identifying the seizure onset zone prior to resective surgery for intractable epilepsy. Current methods rely on EEG and neuroimaging, which are helpful but imperfect techniques.29, 38, 39 As an adjunctive tool for defining the borders of epileptogenic tissue, electrochemical characterization via FSCV could potentially improve surgical outcomes. In the future, using FSCV to monitor the neurochemical changes during epileptiform events may potentially be utilized in devices designed to stop seizures as they occur, such as local delivery of ADO itself.
Conclusions
Our findings support the role of ADO in seizure termination during seizures in a large animal model, which is confirmed in two separate techniques of electrochemisty which corroborate each other. Our human study demonstrates the clinical safety of intraoperative FSCV in a small group of patients, suggesting that FSCV is a potentially useful tool for investigating neurochemical changes during human epilepsy surgery. Our clinical goal is to move toward larger scale human trials with the ultimate aim of chronic neurochemical monitoring. Continuous neurochemical feedback could not only aid in diagnosis and the demarcation of the epileptogenic zone, but eventually provide the necessary input to neuromodulation devices designed to abort seizures before they begin.
Supplementary Material
Supplementary Figure 1: (a) In-vitro flow cell analysis of 5 μM of relative analyte (ie. Adenosine (ADO), ATP, Hypoxanthene (HX), etc. represented by folded voltammograms. All analytes were analyzed with a triangular waveform FSCV from −0.4 to 1.5. Note most purinomic compounds are unique with this electrochemical analysis. However HX and ADO are very similar with the exception of the lack of the 1.0 Volt peak. (b) Although purine nucleic acid components appear similar (Adenosine and Guanosine) in unfolded voltammograms, they are dissimilar when unfolded and are easily resolved.
Acknowledgments
We would like to thank Dr. Penelope Duffy for her extensive editorial work associated with this manuscript. We would also like to thank Drs. Paul Garris, Susannah Tye, Dong-Pyo Jang, Ben Brinkman, and John Bledsoe and the physician extenders supporting this research: Kathy Mortland, Wanda Windschitl, and Debra Gorman. Our thanks also to critical members of the engineering team: Christopher Kimble, April Horne, Dave Johnson, Ken Kressin, Justin Robinson, Andrew Wenger, Bruce Winter, Sid Whitlock, Michael P. Marsh, and Shaun Herring.
Financial and Material Support: This work was supported by: NIH (K08 NS052232, R01 NS 70872 and R01 NS075013 awards to KHL), Mayo Foundation (2008–2010 Research Early Career Development Award for Clinician Scientists award to KHL), NIH (R01 NS063039-01 award to GAW) and Bernard and Irene Waterman Award in Individualized Medicine (award to JJVG). This project was further funded by the Epilepsy Foundation (award to JJVG).
Footnotes
Disclosures and Conflict of Interest: We confirm that we have read the Journal’s position on issues involved in ethical publication and affirm that this report is consistent with those guidelines. Author Kendall H. Lee has received support from: NIH (K08 NS052232, R01 NS 70872 and R01 NS075013), and Mayo Foundation (2008–2010 Research Early Career Development Award for Clinician Scientists award). Author Gregory A. Worrell has received support from NIH (R01 NS063039-01). Author Jamie J. Van Gompel received support from the Bernard and Irene Waterman Award in Individualized Medicine as well as the Epilepsy Foundation. The remaining authors Mark R. Bower, Matt Stead, Su-Youne Chang, Stephen Goerss, Inyong Kim, Kevin Bennet, Fredric B. Meyer, W. Richard Marsh, and Charles D. Blaha have no conflicts of interest or support to disclose. This publication was further supported by NIH/NCRR CTSA Grant Number UL1 RR024150. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIH.
References
- 1.Boison D. The adenosine kinase hypothesis of epileptogenesis. Progress in Neurobiology. 2008;84:249–62. doi: 10.1016/j.pneurobio.2007.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Boison D. Adenosine as a neuromodulator in neurological diseases. Current Opinion in Pharmacology. 2008;8:2–7. doi: 10.1016/j.coph.2007.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Boison D. Adenosine as a modulator of brain activity. Drug News & Perspectives. 2007;20:607–11. doi: 10.1358/dnp.2007.20.10.1181353. [DOI] [PubMed] [Google Scholar]
- 4.Boison D. Adenosine kinase, epilepsy and stroke: mechanisms and therapies. Trends in Pharmacological Sciences. 2006;27:652–8. doi: 10.1016/j.tips.2006.10.008. [DOI] [PubMed] [Google Scholar]
- 5.Aden U, O’Connor WT, Berman RF. Changes in purine levels and adenosine receptors in kindled seizures in the rat. Neuroreport. 2004;15:1585–9. doi: 10.1097/01.wnr.0000133227.94662.c9. [DOI] [PubMed] [Google Scholar]
- 6.Alasvand Zarasvand M, Mirnajafi-Zadeh J, Fathollahi Y, et al. Anticonvulsant effect of bilateral injection of N6-cyclohexyladenosine into the CA1 region of the hippocampus in amygdala-kindled rats. Epilepsy Research. 2001;47:141–9. doi: 10.1016/s0920-1211(01)00300-x. [DOI] [PubMed] [Google Scholar]
- 7.Anschel DJ, Ortega EL, Kraus AC, et al. Focally injected adenosine prevents seizures in the rat. Experimental Neurology. 2004;190:544–7. doi: 10.1016/j.expneurol.2004.07.017. [DOI] [PubMed] [Google Scholar]
- 8.Boison D. Adenosine and epilepsy: from therapeutic rationale to new therapeutic strategies. Neuroscientist. 2005;11:25–36. doi: 10.1177/1073858404269112. [DOI] [PubMed] [Google Scholar]
- 9.Boison D, Huber A, Padrun V, et al. Seizure suppression by adenosine-releasing cells is independent of seizure frequency. Epilepsia. 2002;43:788–96. doi: 10.1046/j.1528-1157.2002.33001.x. [DOI] [PubMed] [Google Scholar]
- 10.During MJ, Spencer DD. Adenosine: a potential mediator of seizure arrest and postictal refractoriness. Annals of Neurology. 1992;32:618–24. doi: 10.1002/ana.410320504. [DOI] [PubMed] [Google Scholar]
- 11.Gouder N, Fritschy J-M, Boison D. Seizure suppression by adenosine A1 receptor activation in a mouse model of pharmacoresistant epilepsy. Epilepsia. 2003;44:877–85. doi: 10.1046/j.1528-1157.2003.03603.x. [DOI] [PubMed] [Google Scholar]
- 12.Huber A, Padrun V, Deglon N, et al. Grafts of adenosine-releasing cells suppress seizures in kindling epilepsy. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:7611–6. doi: 10.1073/pnas.131102898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Vianna EPM, Ferreira AT, Dona F, et al. Modulation of seizures and synaptic plasticity by adenosinergic receptors in an experimental model of temporal lobe epilepsy induced by pilocarpine in rats. Epilepsia. 2005;46(Suppl 5):166–73. doi: 10.1111/j.1528-1167.2005.01027.x. [DOI] [PubMed] [Google Scholar]
- 14.Wiesner JB, Ugarkar BG, Castellino AJ, et al. Adenosine kinase inhibitors as a novel approach to anticonvulsant therapy. Journal of Pharmacology & Experimental Therapeutics. 1999;289:1669–77. [PubMed] [Google Scholar]
- 15.Yildirim M, Marangoz C. Anticonvulsant effects of focal and intracerebroventricular adenosine on penicillin-induced epileptiform activity in rats. Brain Research. 2007;1127:193–200. doi: 10.1016/j.brainres.2006.10.024. [DOI] [PubMed] [Google Scholar]
- 16.Fredholm BB. Adenosine receptors as targets for drug development. Drug News & Perspectives. 2003;16:283–9. doi: 10.1358/dnp.2003.16.5.829316. [DOI] [PubMed] [Google Scholar]
- 17.Winn HR, Welsh JE, Rubio R, et al. Changes in brain adenosine during bicuculline-induced seizures in rats. Effects of hypoxia and altered systemic blood pressure. Circulation Research. 1980;47:568–77. doi: 10.1161/01.res.47.4.568. [DOI] [PubMed] [Google Scholar]
- 18.Van Gompel JJ, Chang S-Y, Goerss SJ, et al. Development of intraoperative electrochemical detection: wireless instantaneous neurochemical concentration sensor for deep brain stimulation feedback. Neurosurgical Focus. 2010;29:E6. doi: 10.3171/2010.5.FOCUS10110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cechova S, Venton BJ. Transient adenosine efflux in the rat caudate-putamen. Journal of Neurochemistry. 2008;105:1253–63. doi: 10.1111/j.1471-4159.2008.05223.x. [DOI] [PubMed] [Google Scholar]
- 20.Swamy BEK, Venton BJ. Subsecond detection of physiological adenosine concentrations using fast-scan cyclic voltammetry. Analytical Chemistry. 2007;79:744–50. doi: 10.1021/ac061820i. [DOI] [PubMed] [Google Scholar]
- 21.Xu Y, Venton BJ. Rapid determination of adenosine deaminase kinetics using fast-scan cyclic voltammetry. Physical Chemistry Chemical Physics. 2010;12:10027–32. doi: 10.1039/c0cp00294a. [DOI] [PubMed] [Google Scholar]
- 22.Van Gompel JJ, Bower MR, Worrell GA, et al. Swine model for translational research of invasive intracranial monitoring. Epilepsia. 2011;52:e49–53. doi: 10.1111/j.1528-1167.2011.03096.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Agnesi F, Tye SJ, Bledsoe JM, et al. Wireless Instantaneous Neurotransmitter Concentration System-based amperometric detection of dopamine, adenosine, and glutamate for intraoperative neurochemical monitoring. Journal of Neurosurgery. 2009;111:701–11. doi: 10.3171/2009.3.JNS0990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bledsoe JM, Kimble CJ, Covey DP, et al. Development of the Wireless Instantaneous Neurotransmitter Concentration System for intraoperative neurochemical monitoring using fast-scan cyclic voltammetry. Journal of Neurosurgery. 2009;111:712–23. doi: 10.3171/2009.3.JNS081348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kimble CJ, Johnson DM, Winter BA, et al. Wireless Instantaneous Neurotransmitter Concentration Sensing System (WINCS) for intraoperative neurochemical monitoring. Conference Proceedings: Annual International Conference of the IEEE Engineering in Medicine & Biology Society. 2009;2009:4856–9. doi: 10.1109/IEMBS.2009.5332773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Shon Y-M, Chang S-Y, Tye SJ, et al. Comonitoring of adenosine and dopamine using the Wireless Instantaneous Neurotransmitter Concentration System: proof of principle. Journal of Neurosurgery. 2010;112:539–48. doi: 10.3171/2009.7.JNS09787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shon Y-M, Lee KH, Goerss SJ, et al. High frequency stimulation of the subthalamic nucleus evokes striatal dopamine release in a large animal model of human DBS neurosurgery. Neuroscience Letters. 2010;475:136–40. doi: 10.1016/j.neulet.2010.03.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brinkmann BH, Bower MR, Stengel KA, et al. Large-scale electrophysiology: acquisition, compression, encryption, and storage of big data. Journal of Neuroscience Methods. 2009;180:185–92. doi: 10.1016/j.jneumeth.2009.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Van Gompel JJ, Meyer FB, Marsh WR, et al. Stereotactic electroencephalography with temporal grid and mesial temporal depth electrode coverage: does technique of depth electrode placement affect outcome? Journal of Neurosurgery. 2010;113:32–8. doi: 10.3171/2009.12.JNS091073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Etherington L-AV, Frenguelli BG. Endogenous adenosine modulates epileptiform activity in rat hippocampus in a receptor subtype-dependent manner. European Journal of Neuroscience. 2004;19:2539–50. doi: 10.1111/j.0953-816X.2004.03355.x. [DOI] [PubMed] [Google Scholar]
- 31.Etherington L-AV, Patterson GE, Meechan L, et al. Astrocytic adenosine kinase regulates basal synaptic adenosine levels and seizure activity but not activity-dependent adenosine release in the hippocampus. Neuropharmacology. 2009;56:429–37. doi: 10.1016/j.neuropharm.2008.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Stoeckler JD, Poirot AF, Smith RM, et al. Purine nucleoside phosphorylase. 3. Reversal of purine base specificity by site-directed mutagenesis. Biochemistry. 1997;36:11749–56. doi: 10.1021/bi961971n. [DOI] [PubMed] [Google Scholar]
- 33.Yegutkin GG. Nucleotide- and nucleoside-converting ectoenzymes: Important modulators of purinergic signalling cascade. Biochimica et Biophysica Acta. 2008;1783:673–94. doi: 10.1016/j.bbamcr.2008.01.024. [DOI] [PubMed] [Google Scholar]
- 34.Phillips PEM, Stuber GD, Heien MLAV, et al. Subsecond dopamine release promotes cocaine seeking. Nature. 2003;422:614–8. doi: 10.1038/nature01476. [DOI] [PubMed] [Google Scholar]
- 35.Morales V, x00E, nA, et al. Simultaneous glutamate and EEG activity measurements during seizures in rat hippocampal region with the use of an electrochemical biosensor. Journal of Neuroscience Methods. 2008;168:48–53. doi: 10.1016/j.jneumeth.2007.09.005. [DOI] [PubMed] [Google Scholar]
- 36.Schrader J, Wahl M, Kuschinsky W, et al. Increase of adenosine content in cerebral cortex of the cat during bicuculline-induced seizure. Pflugers Archiv – European Journal of Physiology. 1980;387:245–51. doi: 10.1007/BF00580977. [DOI] [PubMed] [Google Scholar]
- 37.Chang S-Y, Kim I, Marsh MP, et al. Wireless fast-scan cyclic voltammetry to monitor adenosine in patients with essential tremor during deep brain stimulation. Mayo Clinic Proceedings. 2012;87:760–5. doi: 10.1016/j.mayocp.2012.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Avsar E, Empson RM. Adenosine acting via A1 receptors, controls the transition to status epilepticus-like behaviour in an in vitro model of epilepsy. Neuropharmacology. 2004;47:427–37. doi: 10.1016/j.neuropharm.2004.04.015. [DOI] [PubMed] [Google Scholar]
- 39.Roseti C, Martinello K, Fucile S, et al. Adenosine receptor antagonists alter the stability of human epileptic GABAA receptors. Proceedings of the National Academy of Sciences of the United States of America. 2008;105:15118–23. doi: 10.1073/pnas.0807277105. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure 1: (a) In-vitro flow cell analysis of 5 μM of relative analyte (ie. Adenosine (ADO), ATP, Hypoxanthene (HX), etc. represented by folded voltammograms. All analytes were analyzed with a triangular waveform FSCV from −0.4 to 1.5. Note most purinomic compounds are unique with this electrochemical analysis. However HX and ADO are very similar with the exception of the lack of the 1.0 Volt peak. (b) Although purine nucleic acid components appear similar (Adenosine and Guanosine) in unfolded voltammograms, they are dissimilar when unfolded and are easily resolved.