

SUMMARY
High-risk human papillomavirus (hr-HPV) infection is necessary but not sufficient for cervical cancer development. Recently, P16INK4A gene silencing through hypermethylation has been proposed as an important cofactor in cervical carcinogenesis due to its tumor suppressor function. We aimed to investigate P16INK4A methylation status in normal and neoplastic epithelia and evaluate an association with HPV infection and genotype. This cross-sectional study was performed with 141 cervical samples from patients attending Hospital Moncorvo Filho, Rio de Janeiro. HPV detection and genotyping were performed through PCR and P16INK4A methylation by nested-methylation specific PCR (MSP). HPV frequency was 62.4% (88/141). The most common HPV were HPV16 (37%), HPV18 (16.3%) and HPV33/45(15.2%). An upward trend was observed concerning P16INK4A methylation and lesion degree: normal epithelia (10.7%), low grade lesions (22.9%), high grade (57.1%) and carcinoma (93.1%) (p < 0.0001). A multivariate analysis was performed to evaluate an association between methylation, age, tobacco exposure, HPV infection and genotyping. A correlation was found concerning methylation with HPV infection (p < 0.0001), hr-HPV (p = 0.01), HSIL (p < 0.0007) and malignant lesions (p < 0.0001). Since viral infection and epigenetic alterations are related to cervical carcinoma, we suggest that P16INK4A methylation profile maybe thoroughly investigated as a biomarker to identify patients at risk of cancer.
Keywords: HPV; Cervical cancer; Methylation, P16INK4A
RESUMO
É reconhecido que infecções por papilomavírus humanos de alto risco (HPV) são causa necessária, mas não suficiente para o desenvolvimento do câncer cervical. Recentemente, estudos de silenciamento gênico apontaram que a hipermetilação do gene p16INK4A é importante co-fator para a carcinogênese cervical, eliminando a função supressora de tumor da proteína p16 em lesões malignas. Entretanto poucos estudos avaliaram a relação da metilação com a progressão da doença. Nosso objetivo foi investigar o padrão de metilação do gene P16INK4A em diferentes graus de lesão cervical e sua associação com a infecção por diferentes tipos de HPV. Nosso estudo de corte transversal avaliou 141 amostras cervicais de pacientes atendidas no Hospital Moncorvo Filho, Rio de Janeiro. A detecção e tipagem do HPV foi realizada pela técnica de reação em cadeia da polimerase (PCR), e a metilação do gene P16INK4A pela PCR-metilação específica em formato nested (MSP). A frequência de HPV foi de 62,4% (88/141). O tipo mais prevalente foi o HPV16 (37%), seguido pelo HPV18 (16,3%) e HPV33/45 (15,2%). Curva ascendente foi observada quanto ao padrão de metilação do gene P16INK4A e o grau da lesão: a metilação foi identificada em somente 10,7% das amostras de epitélio normal, em 22,9% das lesões de baixo grau, em 57,1% das lesões de alto grau e em 93,1% dos carcinomas (p < 0,0001). Foram feitas análises univariada e multivariada a fim de correlacionar metilação, idade, exposição ao tabaco, infecção e genótipo de HPV. Foi encontrada correlação da metilação com a infecção pelo HPV (p < 0,0001), genótipos de alto risco (p = 0,01), lesões de alto grau (p < 0,0007) e câncer (p < 0,0001). Uma vez que infecções pelo HPV e alterações epigenéticas mostraram forte associação estatística com o carcinoma cervical, sugerimos que estes padrões de metilação possam ser avaliados como potenciais biomarcadores, combinados à detecção dos HPV oncogênicos para identificação de pacientes em risco de câncer.
INTRODUCTION
Cervical cancer is one of the most common malignancies affecting women worldwide, and its occurrence has steadily increased in those of young ages. This disease is more prevalent among low socioeconomic status populations and is a major health problem in developing countries. It is estimated that cervical carcinoma is responsible for 274,000 deaths worldwide annually24.
It is well established that infection of high-risk human papillomavirus (hr-HPV) is necessary for cervical cancer development; yet it has been observed that infection alone is not a sufficient cause. Rather, genetic and epigenetic factors have been suggested as contributing mechanisms to cervical carcinogenesis20.
Methylation is an epigenetic mechanism of gene silencing that has been recognized to play an important role in several human neoplasias. In mammalian somatic cells, a methyl group is covalently added to cytosine in the context of CpG islands of gene promoters. DNA methylation of these CpG-rich promoters silences gene expression by changing the accessibility of DNA to transcription factors or by recruiting additional silencing-associated proteins2.
A growing number of studies evaluating methylation of host genes in cervical tissue have been published8 19. Methylation has already been detected in precancerous stages, suggesting that in addition to the functional implications of gene inactivation in tumor development, these aberrant methylation patterns represent excellent targets for novel diagnostic approaches based on sensitive PCR techniques2.
Despite the diverse studies available in the literature, there is still a high degree of heterogeneity in methylation frequencies for some genes related to cervical cancer development and progression8 9. P16INK4A is an important target due to its role in cell cycle19. This tumor suppressor gene encodes the p16 protein that is directly involved in inhibiting the G1–S transition of the cell cycle. Specifically, p16 inhibits the ability of CDK4 and CDK6 to phosphorylate the retinoblastoma (Rb) protein and loss of its normal function might lead to uncontrolled cell growth, escape from senescence and induction of proliferation20.
It has been speculated that aberrant methylation in women with or without HPV infection may help identify subgroups at increased risk for histological progression or cancer development. Nevertheless, there have been few studies evaluating tumor suppressor methylation in different stages of cervical disease22 23. Hence, the aim of this study was to investigate the methylation status ofP16INK4A in the full spectrum of cervical lesions as well as in normal epithelia and evaluate a possible association of this epigenetic event with HPV infection and genotype, in order to verify a possible role for methylation in cancer establishment.
MATERIALS AND METHODS
Study population and study design
This cross-sectional study was performed with 141 cervical samples (exfoliated cell and biopsy samples) representing the full spectrum of cervical pathology (i.e., samples with and without various degrees of Cervical Intraepithelial Neoplasia - CIN and Invasive cervical cancer - ICC) as well as in normal healthy epithelia. Samples were collected from patients attending Moncorvo Filho Hospital, Rio de Janeiro, Brazil between December 2004 and November 2006. This research was approved by ethical review boards from the mentioned institution. All the subjects filled out a questionnaire based interview regarding their age and tobacco exposure. Samples from women who were pregnant or infected with HIV were excluded from the study. Samples from the endocervix were collected and bisected for standard cytopathologic diagnosis (and histopathologic, when available), while the other portion was immersed in TRIS-EDTA buffered solution and frozen at -20 °C for subsequent molecular analyses. Cervical specimens were evaluated using the Bethesda System and classified as normal epithelia, low-grade squamous intraepithelial cells (LSIL), high grade squamous intraepithelial cells (HSIL), squamous cells carcinoma (SCC) and adenocarcinoma (AC)18.
HPV DNA detection and typing
HPV DNA detection was performed with consensus primers MY09/11, which amplify a 450bpDNA sequence within the L1 region of HPV, were used to detect generic HPV DNA. Briefly, amplification was carried out in 50 µL reaction mixture (1X PCR buffer, 200 µM dNTPs, 1.5mM MgCl2, 50 pmol of each primer, 0.25 U unit of Taq polymerase, and 5 µL of sample) with 40 cycles of amplification. Each cycle included a denaturation step at 94 °C for one minute, an annealing step at 55 °C for one minute, and a chain elongation step at 72 °C for one minute using DNA Thermal Cycler (Life Technologies). A β-globin primer pair (0.1 pmol each), which amplify a 330bp region of the human DNA, was used as internal control. Polymerase chain reaction (PCR) products were analyzed on 1.5% agarose gel with ethidium bromide staining. HPV typing was executed by PCR amplification with primers from the E6/E7 gene DNA sequences of HPV 6, 11 (low risk for cervical cancer) and 16, 18, 31, 33, 35, 45 and 58 (high risk for cervical cancer)17. PCR conditions were as follow: 94 °C for five min, followed by 35 cycles of 94 °C for 30 seconds, 60 °C for one minute, and 72 °C for one minute. No Template Controls (NTCs) for background contamination were also included. The PCR run was completed after an extension step for ten minutes at 72 °C.
Bisulfite modification
Bisulfite modification was performed according to HERMAN et al. 7 with slight protocol modifications. One µg of genomic DNA was denatured with NaOH. The bisulfite treatment (during which methylated DNA is protected, and unmethylated cytosine is converted to uracil) was carried out for 16h at 50 °C on denatured genomic DNA. Samples were then purified using the Wizard DNA Clean-Up System (Promega®, Madison, WI, USA), treated again with NaOH, ethanol-precipitated and resuspended in sterile water.
Fig. 1. Gel A shows a methylated sample (band on p16M), Gel B shows a methylated sample with mixed forms (P16U + and p16M +). In both gels, Line 1 presents the ladder 50bp Track-It (Lifetech), Line 2 shows positive control for p16 unmethylated, Line 3 shows positive control for p16 methylated, Line 4 and 5 show negative controls for both primer pairs. Line 6 and 7 shows a cervical cancer sample (gel A) and a HSIL sample (Gel B).
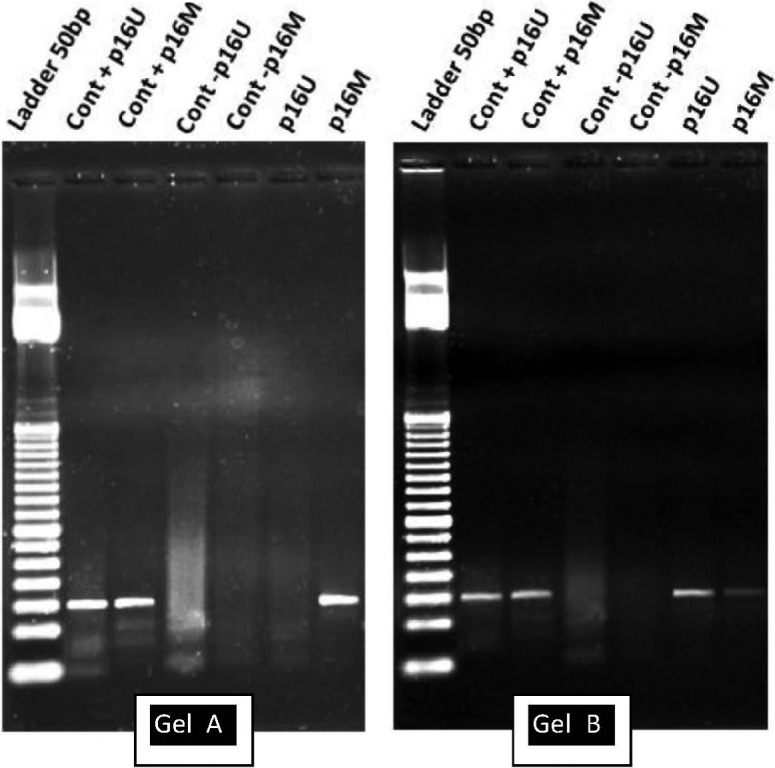
Nested-methylation specific PCR- MSP-PCR was done according to PALMISANO et al. 15 with slight modifications. Briefly, first reaction was carried out with 5 µL of bisulfite treated DNA using the primers did not discriminate between methylated and unmethylated nucleotides. Hence, amplicons were subjected to a second PCR step that were designed to recognize bisulfite-induced modifications of unmethylated cytosines, converted to uracils through PCR, from unmodified cytosines of methylated samples, by using two pairs of primers (methylated M and unmethylated U). NTC's were included. Methylation of a specific gene was considered to be present if both the specimen and the positive control DNA were amplified by M primers after sodium bisulfite modification. PCR results were interpreted by two independent reviewers that had no access to histology or cytology results.
Statistical analysis
Statistical analysis was performed using S-PLUS 8.0 (TIBCO Software, USA). Differences were considered statistically significant when p < 0.05 (CI: 95%). The frequency of categorical variables was described by their absolute values and percentages, whereas numerical variables as mean and standard deviation. Chi-square or Fisher's exact tests were used for the comparison of categorical variables. For numerical variables, as far as P16INK4A methylation status is concerned, Student's t or Willcoxon rank-sum tests were used. A multivariate analysis using logistic regression was performed to evaluate the association between the presence of methylation of P16INK4A and other variables such as lesion of the uterine cervix, smoking habits, age and HPV infection.
RESULTS
After cytological and histological analysis, studied samples were classified as normal, corresponding to 19.8% (28/141) of the cases, LSIL (24.8%, 35/141), HSIL (34.8%, 49/141) and invasive cancer (20.6%, 29/141). Among cancer cases, 25 were classified as squamous cell carcinoma (SCC) and four as adenocarcinoma (AC), according to histopathological diagnosis.
The average age of the studied women was 39.1 ± 15.9 (19-85 years old). In samples presenting cervical lesions, there was an upward trend in the patient's age according to the severity of the lesions (LSIL: 35.9 ± 13.7; HSIL: 38.6 ± 17.9; Ca: 42.8 ± 12.4, respectively). Nearly 40% of the patients were current smokers and the average tobacco exposure was 15.3 ± 3.1 cigarettes per day. Women without lesions presented the smaller tobacco exposure (11.7 ± 8.0 cigarettes/day) and those with LSIL, the highest (19.2 ± 8.2). Patients showing AC were not smokers, while those presenting SCC consumed 14.8 ± 6.8 cigarettes/day, average.
Regarding the molecular detection of HPV DNA, 62% (88/141) of the studied cases were HPV positive, by using consensual primers MY09/11. Patients presenting no cytological alterations had an HPV prevalence of 7.1% (2/28), while in altered cases with cytologic identifiable lesions it was detected in 76.1% (86/113) of the samples. Among them, LSIL showed prevalence of 60% (21/35) and according to the severity of the lesion, prevalence rates increased to 75.5% (37/49) in HSIL and 96.5% (28/29) in carcinoma. A strong statistical association was revealed concerning HPV DNA detection and cervical lesion (OR: 41.6(8.6-70.6); p < 0.0001, CI: 95%) (Table 1).
Table 1. Univariate analysis of independent variables and methylation status of P16INK4A among the studied patients.
| Variable | P16INK4A methylation status | ||
|---|---|---|---|
| Methylated | Unmethylated | p value | |
| Age (years) | p = 0.248 | ||
| < 40 years | 17 | 27 | |
| ≥ 40 years | 49 | 48 | |
| Age (years) | 43.2 ± 15.3 | 36.9 ± 13.4 | p = 0.011 |
| Active smoking | p = 0.107 | ||
| YES | 38 | 31 | |
| NO | 28 | 44 | |
| Cigarettes/day | 15.9 ± 8.8 | 14.4 ± 9.4 | p = 0.622 |
| Cigarettes/day | p = 0.394 | ||
| < 20 | 14 | 2 | |
| ≥ 20 | 11 | 5 | |
| HPV positive | p < 0.0001 | ||
| YES | 58 | 30 | |
| NO | 8 | 45 | |
| hr-HPV | p < 0.0001 | ||
| YES | 55 | 22 | |
| NO | 3 | 8 | |
| Cytology results | p < 0.0001 | ||
| Normal | 3 | 25 | |
| Altered | 63 | 50 | |
HPV genotyping was successful in 96.6% (85/88) of the samples. Undetermined HPV were designed as HPV X. High frequency of hr-HPV was detected, representing 78.4% (69/88) of the studied samples, prevailing in samples with altered cytology results (p = 0.03). Infections with low risk types were identified in 7.9% (7/88) of the cases, all in cytological negative cases. We detected 10.2% (9/88) of the samples presenting co-infection with two genotypes (Table 2). Among hr-HPV detected, HPV16 was the most prevalent type (37%), followed by HPV18 (16.3%), HPV33 and HPV45 (15.2% both). Other high risk genotypes were found in lower rates such as HPV31 (2.4%) and HPV58 (1.2%). HPV35 was not detected in any of the samples included in the study. Among low risk types, HPV6 was the most common, being detected in 13% of the cases, followed by HPV11 that was shown in one case of co-infection along with HPV6.
Table 2. Prevalence of low and high-risk human papillomavirus (HPV) genotypes according to histological grading of the lesions.
| Lesion | Low risk HPV | High risk HPV* | Co-infection | HPV X | TOTAL |
|---|---|---|---|---|---|
| Normal Cells | 1 | 1 | - | - | 2 |
| LSIL | 3 | 16 | 1a | 1 | 21 |
| HSIL | 3 | 26 | 6b | 2 | 37 |
| Ca | - | 26 | 2c | - | 28 |
| TOTAL | 7 (7.9%) | 69 (78.4%) | 9 (10.2%) | 3 (3.4%) | 88 |
High risk HPV versus cervical lesions (χ2: p = 0.03);
HPV 6/16,
HPV 6/11, 6/33, 6/16, 6/18, 16/45, 33/45,
Two cases of 16/18.
The methylation profile of the P16INK4A gene was determined: sixty-six (44.6%) of the cases displayed hypermethylated of P16INK4A promoter. Among them, 15.2% (10/66) presented an exclusively methylated pattern, while the rest 84.4% (56/66) showed a mixed pattern composed by methylated and unmethylated DNA (amplification observed for both primers M and U).
The frequency of methylation in samples showing cytological/histological normal results was 10.7% (3/28). Low and high grade lesions displayed methylation 22.9% (8/35) and 57.1% (28/49), respectively. The highest frequency was observed in malignant lesions, 93.1% (27/29). An upward trend was observed according to the severity of the cervical lesions, with a high statistical significance (p < 0.001).
After determining P16INK4A hypermethylation status, an univariate analysis were conducted in order to verify a possible association between variables such as age, tobacco smoking, smoking dosage (cigarettes per day), cytological diagnosis, HPV infection and genotype, summarized in Table 1. There was no significant differences regarding the average number of cigarettes smoked per day between women who presented methylated and unmethylated P16INK4A profile (p = 0.622). Furthermore, no significant association was found between active tobacco exposure (p = 0.107) or smoke quantitation (p = 0.394) and P16INK4A methylation status. Despite a significant statistical difference in the average age of women with P16INK4A methylated or unmethylated (p = 0.011), no association between age and P16INK4A methylation status was found (p = 0.248). Interestingly, a strong statistical significance was found between presence of HPV genome, presence of any hr-HPV genotype and with altered cytology results and lesions (p < 0.0001) (Table 1).
To confirm univariate analysis results and avoid confounding, a multivariate analysis was performed using logistic regression (Table 3). According to our results, no association was found between age, active tobacco exposure or smoking quantitation and the presence of P16INK4A methylation. On the other hand, a strong statistical significance was found between HPV DNA detection/hr-HPV genotypes and methylated P16INK4A profile (p < 0.0001 and p = 0.01). Among hr-HPV genotypes, the chance of methylation among women with HPV18 infection (p = 0.02, OR: 10.75 (1.35-25.3)) and HPV16 (p = 0.03, OR: 5.81 (1.14-29.4)) was higher when compared to other genotypes detected in our study (data not shown in the table). Interestingly, no association was found between methylation status and low grade lesions (p = 0.05) but we found a correlation between high grade and cancer cytology results and the presence of methylation (p = 0.005 for HSIL and p < 0.0001 for Ca).
Table 3. Multivariate analysis of independent variables and methylation status of P16INK4A among the studied patients using logistic regression model.
| Variable | P16INK4A methylation status | ||
|---|---|---|---|
| Methylated (n = 66) | Unmethylated (n = 75) | p value * [OR (IC)] | |
| Age (years) | 43.2 | 36.9 | p = 0.1 |
| OR: 1.02 (0.99-1.05) | |||
| Tobacco exposure | 38 | 31 | p = 0.11 |
| OR: 1.95 (0.84-4.50) | |||
| Cigarettes/day | 15.9 | 14.4 | p = 0.1 |
| OR: 1.04 (0.76-2.74) | |||
| Presence of HPV | 66 | 30 | p < 0.0001 |
| OR: 6.04 (2.25-16.4) | |||
| hr-HPV | 55 | 22 | p = 0.01 |
| OR: 5.82 (1.95-19.3) | |||
| LSIL | 8 | 27 | p = 0.05 |
| OR: 3.82 (0.95-15.4) | |||
| HSIL | 28 | 21 | p = 0.005 |
| OR: 10.23 (2.73-38.3) | |||
| Carcinoma | 27 | 2 | p < 0.0001 |
| OR: 52.08 (10.57-72.69) | |||
DISCUSSION
In the last decade, it has been shown that DNA methylation patterns are profoundly altered in several human neoplasias. In malignant cells, while the most part of the genome undergoes global genomic hypomethylation, CpG islands within gene promoter regions suffer dense hypermethylation. This is the best characterized epigenetic change associated with gene transcriptional silencing, thus providing a DNA-based surrogate marker for expression status10. It has been increasingly recognized over the past decade that the CpG islands of a large number of genes which are unmethylated in normal tissue are methylated to varying degrees in multiple types of human neoplasm, including cervical cancer9 21 22.
The most promising hypermethylated genes that could be used as biomarkers for cervical cancer have been recently analyzed in a systematic review23. A large heterogeneity among results from different studies was found regarding the hypermethylation frequency of several genes. In addition to these discordances, few published studies have evaluated the correlation between P16INK4A hypermethylation in cervical lesions along with HPV detection and genotyping14. We investigated the frequency of P16INK4A hypermethylation in samples presenting all cervical carcinogenesis lesion stages and attempted to correlate this phenomenon with HPV infection, genotyping and cytological results.
Our results showed a higher frequency of methylated alleles when compared to previous studies, which points out frequencies varying from 5% to 60% in cervical cancer samples and 3% to 35% in premalignant lesions3 8 12 22. It is noteworthy that the majority of studies used cervical biopsies and conventional MSP technique to evaluate P16INK4A methylation status. In contrast, in the current study we used exfoliated cell samples, which are the primary non invasive screening method for detection of cervical cancer and precursor lesions, combined with Nested-MSP. The feasibility of this combination has been demonstrated previously6.
In our study the methylation mixed pattern (amplification with both primers: p16U and p16M) predominated in contrast with NEHLS et al. 13 study, which performed MSP technique with micro-dissected biopsies and showed a predominance of simple methylated forms. The percentage of mixed methylation pattern decreased after micro-dissection probably due to removal of contaminating cells such as normal, stromal cells and inflammatory cells.
We believe that these methodological differences might partly explain our results, once PALMISANO et al. 15 showed that the sensitivity of Nested-MSP is five-fold higher than the conventional method. Furthermore, these discrepancies may be due to the different characteristics of study populations since it is well established that factors such as age and smoking habits are also associated with changes in the epigenetic profile of an individual25.
Univariate analysis results showed a strong statistical association between the presence of cervical lesion and P16INK4A methylation (p < 0.0001) and multivariate analysis confirmed this association. However, it was statistically significant only for HSIL and Ca cases (p = 0.005 and p < 0.0001, respectively) (Table 3). Our results suggest that P16INK4A hypermethylation may be a cofactor in cervical carcinogenesis stages, contributing to the emergence of premalignant lesions and neoplastic transformation of cervical epithelium, which is in agreement with JEONG et al. 8 and VIRMANI et al. 22. We believe that this phenomenon contributes to progression of low-grade lesions towards high-grade lesions, which are considered the true precursor of cervical cancer. Nevertheless, other studies are necessary to confirm our data.
We also had the opportunity to assess the association between several variables (age, tobacco exposure, cervical lesion degree, HPV infection and genotype) and P16INK4A hypermethylation. No statistical association was found regarding smoking nor age with the presence of methylated forms of P16INK4A (p = 0.1; p = 0.11, respectively) (Table 1). LEA et al. 11 found that aberrant methylation of p16 gene is strongly correlated with active tobacco exposure in cervical high-grade dysplasia and SCC. However, the authors did not evaluate the number of cigarettes consumed per day to eliminate possible bias and did not perform a multivariate analysis. In our study we were able to evaluate this association but no statistical association was found (Table 3).
In our study, we report a highly significant statistical association between the presence of hr-HPV infection and detection of methylated P16INK4A forms (p < 0.001) as showed by SPATHIS et al. 19. In fact, diverse studies have already correlated viral exposure to epigenetic alterations and human cancer such as Hepatitis B Virus and Hepatitis C Virus with hepatocellular carcinoma16 26.
WU et al. 25 described a relevant HPV16 and 18 infection association within activation of p16INK4A gene through promoter hypermethylation in lung cancer samples obtained from non-smoking patients. The authors proposed that HPV infection might promote hypermethylation. It is well known that E7 gene of HPV16 and 18 is involved in Rb inactivation and release of histone deacetylase (HDAC) from E2F-Rb-HDAC complex, enhancing P16INK4A hypermethylation through chromatin remodeling by HDAC5. Such DNA methylomes were now confirmed by in vivo studies (FERNANDEZ et al. 4).
Besides, a study performed by BURGERS et al. 1 found that HPV oncoproteins have the ability to modulate the enzymatic machinery involved in gene methylation in vitro and in vivo. The authors described that HPV16 E7 protein binds to and stimulates the activity of the DNMT1 enzyme, which are directly involved in DNA methylation. These discoveries suggested that the virus could indirectly promote the silencing of its own genes, evading host immune defenses to favor the establishment of a persistent infection. However, uncontrolled activation of these enzymes could eventually lead to important cellular genes silencing, such as gene P16INK4A.
New approaches for the recognition and treatment of high-risk patients are needed to improve survival and avoid over-treatment of low-risk patients. Indeed, identification of viral and host factors that contribute to the delineation of women at the highest risk of HPV infection persistence and progression will lead to selected candidate markers that can potentially be used for diagnostic or prognostic purposes.
Despite their being interesting, our results should be interpreted with caution since we cannot state whether hr-HPV infection triggers P16INK4A methylation and larger studies are needed to confirm our findings. Such studies are of great importance since hypermethylation counteracts p16 expression disturbing cell cycle control and thus may represent a risk factor for cervical premalignant and malignant lesion development.
Methylome studies, along with our results suggest these epigenetic events to be biomarkers that can be used as complementary tools to define risk of cancer establishment along with hr-HPV detection. The clinical validation and utility of candidate markers must, therefore, be thoroughly investigated.
Acknowledgments
This work was supported by grants from FAPERJ E-26111.511/2010, PROPPI/UFF and CNPq PhD grants.
Funding Statement
Financial support: Grant from Faperj (APQ1 E26/111.511/2010) and Proppi/UFF.
Footnotes
Financial Support: Grant from Faperj (APQ1 E26/111.511/2010) and Proppi/UFF.
REFERENCES
- 1.Burgers WA, Blanchon L, Pradhan S, Launoit YD, Kouzarides T, Fuks K. Viral oncoproteins target the DNA methyltransferases. Oncogene. 2007;26:1650–5. doi: 10.1038/sj.onc.1209950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Esteller M. Epigenetics in cancer. N Engl J Med. 2008;358:1148–59. doi: 10.1056/NEJMra072067. [DOI] [PubMed] [Google Scholar]
- 3.Feng Q, Balasubramanian A, Hawes SE, Toure P, Sow PS, Dem A, et al. Detection of hypermethylated genes in women with and without cervical neoplasia. J Natl Cancer Inst. 2005;97:273–82. doi: 10.1093/jnci/dji041. [DOI] [PubMed] [Google Scholar]
- 4.Fernandez AF, Rosales C, Lopez-Nieva P, Graña O, Ballestar E, Ropero S, et al. The dynamic DNA methylomes of double-stranded DNA viruses associated with human cancer. Genome Res. 2009;19:438–51. doi: 10.1101/gr.083550.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Finzer P, Kuntzen C, Soto U, ZurHausen H, Rosl F. Inhibitors of histone deacetylase arrest cell cycle and induce apoptosis in cervical carcinoma cells circumventing human papillomavirus oncogene expression. Oncogene. 2001;20:4768–76. doi: 10.1038/sj.onc.1204652. [DOI] [PubMed] [Google Scholar]
- 6.Gustafson KS, Furth EE, Heitjan DF, Fansler ZB, Clark DP. DNA methylation profiling of cervical squamous intraepithelial lesions using liquid-based cytology specimens: an approach that utilizes receiver-operating characteristic analysis. Cancer. 2004;102:259–68. doi: 10.1002/cncr.20425. [DOI] [PubMed] [Google Scholar]
- 7.Herman JG, Graff JR, Myohanen S, Nelkin BD, Baylin SB. Methylation-specific PCR: a novel PCR assay for methylation status of CpG islands. Proc Natl Acad Sci. 1996;93:9821–6. doi: 10.1073/pnas.93.18.9821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jeong DH, Youm MY, Kim YN, Lee KB, Sung MS, Yoon HK, et al. Promoter methylation of p16, DAPK, CDH1, and TIMP-3 genes in cervical cancer: correlation with clinicopathologic characteristics. Int J Gynecol Cancer. 2006;16:1234–40. doi: 10.1111/j.1525-1438.2006.00522.x. [DOI] [PubMed] [Google Scholar]
- 9.Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet. 2002;3:415–28. doi: 10.1038/nrg816. [DOI] [PubMed] [Google Scholar]
- 10.Jones PA, Liang G. Rethinking how DNA methylation patterns are maintained. Nat Rev Genet. 2009;10:805–811. doi: 10.1038/nrg2651. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lea JS, Coleman R, Kurien A, Schorge JO, Miller DS, Minna JD. Aberrant p16 methylation is a biomarker for tobacco exposure in cervical squamous cell carcinogenesis. Am J Obstet Gynecol. 2004;190:674–9. doi: 10.1016/j.ajog.2003.09.036. [DOI] [PubMed] [Google Scholar]
- 12.Narayan G, Arias-Pulido H, Koul S, Vargas H, Zhang FH, Villella J, et al. Frequent promoter methylation of CDH1, DAPK, RARB, and HIC1 genes in carcinoma of cervix uteri: its relationship to clinical outcome. Mol Cancer. 2003;2:1–12. doi: 10.1186/1476-4598-2-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Nehls K, Vinokurova S, Schmidt D, Kommoss F, Reuschenbach M, Kisseljov F, et al. p16 methylation does not affect protein expression in cervical carcinogenesis. Eur J Cancer. 2008;44:2496–505. doi: 10.1016/j.ejca.2008.07.014. [DOI] [PubMed] [Google Scholar]
- 14.Nuovo GJ, Plaia TW, Belinsky SA, Baylin SB, Herman JG. In situ detection of the hypermethylation induced inactivation of the p16 gene as an early event in oncogenesis. Proc Natl Acad Sci. 1999;96:12754–9. doi: 10.1073/pnas.96.22.12754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Palmisano WA, Divine KK, Saccomanno G, Gilliland FD, Baylin SB, Herman JG, et al. Predicting lung cancer by detecting aberrant promoter methylation in sputum. Cancer Res. 2000;60:5954–8. [PubMed] [Google Scholar]
- 16.Sanchez-Cespedes M, Decker PA, Doffek KM, Esteller M, Westra WH, Alawi EA, et al. Increased loss of chromosome 9p21 but not p16 inactivation in primary non-small cell lung cancer from smokers. Cancer Res. 2001;61:2092–6. [PubMed] [Google Scholar]
- 17.Silva KC, Rosa MLG, Moyses N, Afonso LA, Oliveira LHS, Cavalcanti SMB. Risk factors associated with human papillomavirus infection in two populations from Rio de Janeiro, Brazil. Mem Inst Oswaldo Cruz. 2009;104:885–91. doi: 10.1590/s0074-02762009000600011. [DOI] [PubMed] [Google Scholar]
- 18.Solomon D, Davey D, Kuman R, Moriarty A, O'Connor D, Prey M, et al. The 2001 Bethesda System: terminology for reporting results of cervical cytology. JAMA. 2002;287:2114–9. doi: 10.1001/jama.287.16.2114. [DOI] [PubMed] [Google Scholar]
- 19.Spathis A, Aga E, Alepaki M, Chranioti A, Meristoudis C, Panayiotides I, et al. Promoter methylation of p16(INK4A), hMLH1, and MGMT in liquid-based cervical cytology samples compared with clinicopathological findings and HPV presence. Infect Dis Obstet Gynecol. 2011;2011:1–5. doi: 10.1155/2011/927861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Szalmás A, Kónya J. Epigenetic alterations in cervical carcinogenesis. Semin Cancer Biol. 2009;19:144–52. doi: 10.1016/j.semcancer.2009.02.011. [DOI] [PubMed] [Google Scholar]
- 21.Teng IW, Hou PC, Lee KD, Chu PY, Yeh KT, Jin VX, et al. Targeted methylation of two tumor suppressor genes is sufficient to transform mesenchymal stem cells into cancer stem/initiating cells. Cancer Res. 2011;71:4653–63. doi: 10.1158/0008-5472.CAN-10-3418. [DOI] [PubMed] [Google Scholar]
- 22.Virmani AK, Muller C, Rathi A, Zoechbauer-Mueller S, Mathis M, Gazdar SF. Aberrant methylation during cervical carcinogenesis. Clin Cancer Res. 2001;7:584–9. [PubMed] [Google Scholar]
- 23.Wentzensen N, Sherman ME, Schiffman M, Wang SS. Utility of methylation markers in cervical cancer early detection: appraisal of the state-of-the-science. Gynecol Oncol. 2009;112:293–9. doi: 10.1016/j.ygyno.2008.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.WHO/ICO Information Centre on HPV and Cervical Cancer Human papillomavirus and related cancers in Brazil. [[cited 2010 May 12)];Summary Report. 2010 Available from: www.who.int/hpvcentre. [Google Scholar]
- 25.Wu MF, Cheng YW, Lai JC, Hsu MC, Chen JT, Liu WS, et al. Frequent p16INK4a promoter hypermethylation in human papillomavirus-infected female lung cancer in Taiwan. Int J Cancer. 2005;113:440–5. doi: 10.1002/ijc.20597. [DOI] [PubMed] [Google Scholar]
- 26.Yang B, Guo M, Herman JG, Clark DP. Aberrant promoter methylation profiles of tumor suppressor genes in hepatocellular carcinoma. Am J Pathol. 2003;163:1101–7. doi: 10.1016/S0002-9440(10)63469-4. [DOI] [PMC free article] [PubMed] [Google Scholar]