

Abstract
A major hindrance for the development of psychiatric drugs is the prediction of how treatments can alter complex behaviors in assays which have good throughput and physiological complexity. Here we report the development of a medium-throughput screen for drugs which alter anxiety-like behavior in adult zebrafish. The observed phenotypes were clustered according to shared behavioral effects. This barcoding procedure revealed conserved functions of anxiolytic, anxiogenic and psychomotor stimulating drugs and predicted effects of poorly characterized compounds on anxiety. Moreover, anxiolytic drugs all decreased, while anxiogenic drugs increased, serotonin turnover. These results underscore the power of behavioral profiling in adult zebrafish as an approach which combines throughput and physiological complexity in the pharmacological dissection of complex behaviors.
Introduction
Along with benzodiazepines, drugs targeting the serotonergic system represent a major class of anxiolytic drugs. Among available serotonergic drugs, selective serotonin reuptake inhibitors still represent the most prescribed treatment for anxiety disorders, even though they are associated with low efficacy in a considerable proportion of patients, a delayed onset of therapeutic action, and diverse collateral effects which reduce tolerance (e.g., sexual dysfunction, weight changes). Benzodiazepines, on the other hand, are associated with decreased responsiveness over time, withdrawal-related symptoms, and sedation [1]. The need for novel, more efficient anti-anxiety drugs is paramount [2], but the proper level for consistent results with relatively high throughput is difficult to determine [3], [4].
While in other fields of pharmaceutical discovery in vitro target-based assays are sufficient to accelerate discovery and increase throughput, these approaches (while certainly initially useful [5]) are unfeasible in most areas of psychopharmacology, where the appropriate targets remain unknown [4], [6]. Thus, psychopharmacological research relies on phenotype-based approaches, in which behavior is the principal endpoint [7]–[12].
A major obstacle in the discovery of psychopharmacological agents is the difficulty in predicting how candidate drugs can alter complex behaviors. While the usual approach of complete identification of the mechanisms of disease pathology is certainly useful, this strategy can preclude the discovery of novel psychoactive drugs that target unexpected processes. It has been proposed that phenotype-based approaches in the context of a whole organism are a suitable alternative to overcome these limitations [4], [7], [10]–[12], but the throughput of these assays is usually low [3], [4], [6]. Behavioral assays in mammals represent a high degree of physiological complexity in relation to in vitro target-based assays, but the throughput is low; conversely, in vitro assays are high-throughput but low-content [3], [4], [6]. Behavioral assays in larval and adult zebrafish have the potential to combine the high content of phenotype-based approaches with the medium-to-high-throughput of in vitro chemical screening methods [13]–[22].
Zebrafish became a widely used model organism due to its fecundity, physiological complexity, and the existence of many genetic and genomic tools [6], [16], [23]. While larval zebrafish has been proposed as an ideal model for phenotype-based behavioral assays in psychopharmacological drug discovery [24], the behavioral repertoire of developing zebrafish is considerably restricted [25], [26] and considerable neurochemical and behavioral differences exist between larvae and adults [27]–[29]. In contrast, adult zebrafish display a complete repertoire of behaviors which have been characterized physiologically and pharmacologically [17]. Among these, drug-sensitive phenotypes of anxiety, such as geotaxis [30]–[33] and scototaxis [34]–[38], have been described and pharmacologically and behaviorally validated.
Here, we describe the results of phenotyping in the scototaxis test, using adult zebrafish, as a medium-throughput, high-content assay for anxiolytic and anxiogenic drugs. Compounds analyzed included drugs with known anxiolytic effect (benzodiazepines, buspirone), drugs with known anxiogenic effect (caffeine), as well as drugs with known motor stimulating effects (diethylpropion, bupropion). In addition, drugs acting on adenosinergic (DPCPX, PACPX, ZM 241,285, and DMPX), glutamatergic (NMDA, MK-801), serotonergic (serotonin, WAY 100,635, SB 224,289, moclobemide, Hypericum perforatum extract) and nitrergic (L-NOARG, SNP) systems were tested. Multiple behavioral parameters were measured, including time spent in the white compartment, locomotion into and on the white compartment, and ethologically-defined endpoints such as thigmotaxis, erratic swimming, risk assessment and freezing. These endpoints were then analyzed using a clustering paradigm, used before for profiling rest/wake promoting-drugs in larval zebrafish [34], [36], [39], [40] and to analyze anxiety-like behavior and habituation in the novel tank test [41], [42].
Results and Discussion
Given the diversity of potential drug effects in the different behavioral parameters, a “behavioral fingerprint” was assigned to each compound and dose by determining the Maximum Predictive Value [15], [43] for each effect and applying clustering algorithms to organize behavioral parameters and molecules [44]. This analysis allowed the organization of the data set broadly into anxiolytic, anxiogenic, and motor stimulating, identifying four clusters which correspond to “avoidance” (Time on white and Thigmotaxis), “locomotor” (Entries in white and Midline crossings), “risk assessment” (Latency to white, Risk assessment and Erratic swimming) and “fear” (Freezing) measures (Figure 1). For example, buspirone and diazepam produced a marked anxiolytic-like effect – increasing time spent in the white compartment and decreasing risk assessment, thigmotaxis and freezing in the white compartment (Figures 2A and 2B) –, while caffeine had an opposite profile (Figure 2C).
Figure 1. Behavioral fingerprint of selected drugs on the scototaxis test.
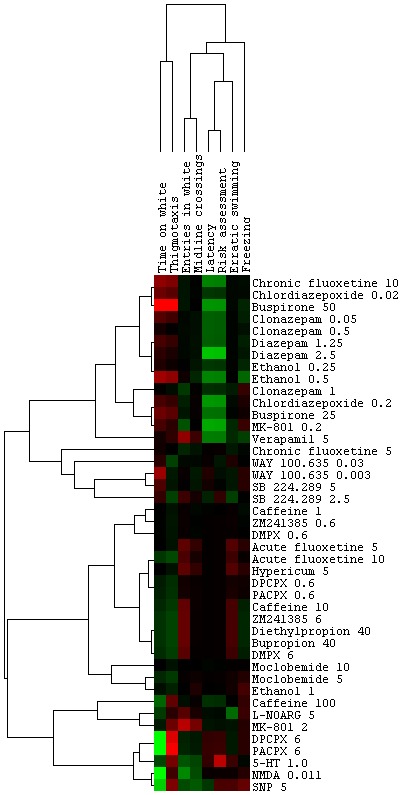
Pharmacological manipulations were hierarchically clustered to link compounds to behaviors. In the clustergram, each cell represents the Maximum Predictive Value (red – higher than controls; green – lower than controls).
Figure 2. Effects of (A) buspirone, (B) diazepam and (C) caffeine on time on white (upper left), risk assessment (upper right), thigmotaxis (lower left), and freezing (lower right).

Bars represent standard error of the mean, and whiskers represent the 2.5 and 97.5 percentile. *, p<0.05; **, p<0.01; ***, p<0.001.
Cluster analysis revealed a high degree of predictive validity in the proposed assay (Figures 1–3). First, anxiolytic drugs with clinical efficacy (benzodiazepines, chronic [14 days] fluoxetine) cluster together with buspirone, and anxiogenic drugs (NMDA, 5-HTP) cluster with caffeine (Figure 1). Second, motor-stimulating drugs (which represent potential false positives in locomotor-based assays) form their own cluster (Figure 1). For example, a low dose of caffeine (Figure 3A), a low dose of ethanol (Figure 3B), and bupropion (Figure 3C) increased locomotion, without effects on ethological measures or time in the white compartment. Second, drugs with multiple targets (e.g., ethanol, caffeine) correlated with drugs in different cluster in a dose-dependent way (Figure 1); caffeine, for example, clustered with anxiogenic drugs at a higher dose, and with stimulant drugs at a lower dose (Figure 1). Third, anxiolytic/anxiogenic and locotomor stimulating effects closely followed those observed in mammals. Fourth, compounds which clustered on the “anxiolytic” effect (Figure 1) all reduced serotonin turnover, which was correlated with time spent on the white compartment in these groups (r2 = 0.5688, p = 0.007) (Figure 4). These analyses indicate that compounds with shared systems effects produce similar phenotypes which are conserved across vertebrates. While from a neuroanatomical and genomic point of view the serotonergic system diverges from that of mammals [45], these and other data strongly suggest that the function of the serotonergic system is conserved across vertebrates. It should also be observed that some behavioral components (time on white, thigmotaxis, latency to white and risk assessment) are more strongly affected by drug treatments (Figure 1), suggesting that those parameters have a stronger predictive value to pharmacological treatments. Interestingly, time on white and thigmotaxis cluster together, while latency to white and risk assessment fall together on another cluster. Erratic swimming and freezing, while affected by anxiogenic and anxiolytic drug treatments, show a weaker liability. These results are in accordance with those observed in the novel tank test [15], in which erratic swimming and freezing had weaker predictive power in relation to time in the upper half of the tank and latency to upper half.
Figure 3. Effects of (A) caffeine, (B) ethanol, (C) bupropion and (D) verapamil on time on white (top) and entries on white (bottom).

Bars represent standard error of the mean, and whiskers represent the 2.5 and 97.5 percentile. *, p<0.05; **, p<0.01; ***, p<0.001.
Figure 4. Drugs which cluster on the 'anxiolytic' group decrease 5-HT turnover in the brain.

(A) Turnover rates, as measured by 5-HIAA:5-HT ratios, normalized to the values of vehicle-treated animals, for the following drugs: fluoxetine (FLX; chronic treatment with 10 mg/kg); chlordiazepoxide (CDZ; 0.02 mg/kg); clonazepam (CLZ; 0.05 mg/kg); diazepam (DZP; 1.25 mg/kg); buspirone (BUS; 50 mg/kg); ethanol (EtOH, 2.5%); dizocilpine (MK; 0.005 mg/kg); verapamil (VER; 5 mg/kg); WAY 100,635 (WAY; 0.03 mg/kg); and SB 224,289 (SB; 2.5 mg/kg). Asterisks mark statistically significant differences in relation to vehicle-treated animals (F10, 43 = 45.99, p<0.0001, one-way ANOVA followed by Dunnett's Multiple Comparison test). Bars represent mean (B) Correlation between turnover rates (Y-axis) and time spent in the white compartment (X-axis) for vehicle- and drug-treated animals (n = 4 for each point). Points represent means and error bars represent standard errors. A negative correlation is found between the decrease in serotonin turnover and the increase in time on white produced by a drug (r2 = 0.5688, p = 0.0073).
Furthermore, cluster analysis revealed novel behavioral effects of poorly characterized substances. For example, the calcium channel blocker verapamil, an anti-arrhythmic and anti-anginal agent, produced a small anxiolytic effect, clustering with sedative doses of ethanol and clonazepam (Figure 1, Figure 3D). Interestingly, verapamil has been shown to be sedative in larval zebrafish [15], [41]. This effect is unlikely to be a consequence of antihypertensive effects, because sodium nitroprusside (SNP) had an opposite effect and clustered with NMDA (Figure 1).
These results reveal a conserved neuropharmacology in vertebrates and identify novel regulators of anxiety, such as the glutamatergic/nitrergic system. Previously validated targets in zebrafish anxiety assays include the cholinergic system [30], [46], [47], histamine [48]–[50], central benzodiazepine receptors [34], [51]–[54], endogenous opioids [32], [55], endocannabinoids [55]–[57], serotonin [58]–[60], and adenosine [34], [53], [61], [62]. The behavioral profiling observed in this paper is also predictive of decreased serotonin turnover, suggesting a common neurobiological mechanism of anxiolysis. This is a surprising result given that, while the effects of serotonergic drugs on zebrafish behavior seem to be rather conserved, from a genomic and neuroanatomical point of view the serotonergic system from mammals is different from that of teleosts (e.g., presence of hypothalamic and tectal serotonergic nuclei and duplicated htr1a and sert genes in zebrafish) [45]. Nonetheless, these results support a role for the serotonergic system in controlling zebrafish anxiety, suggesting conserved function, if not conserved structure.
The medium throughput of this method in relation to, e.g., larval profiling [41] is offset by the increased information content produced by analyzing multiple parameters (anxiety-like responses, dark preference, motor parameters) and using developed, adult animals. We underscore that the outstanding predictive validity of the proposed assay is also accompanied by construct validity [6], [41], [42], which enriches and directs the predictive validity of the model. Therefore, light/dark preference in adult animals can complement traditional target-based discovery methodologies, combining the physiological complexity of in vivo assays with medium-to-high-throughput, low-cost screening [63]. This has been done previously – albeit with a limited amount of drug treatments – with the novel tank test, with results similar to those presented here: caffeine, for example, clustered among anxiogenic manipulations, while chronic fluoxetine clustered among anxiolytic manipulations [15]. Similarly, anxiogenic treatments increase erratic swimming and freezing duration in the novel tank test [15] as well as in the present experiments. Caution should be taken, however, in generalizing results from both assays, since drug effects in the light/dark and in the novel tank tests are not always the same – and, in fact, some drugs, such as pCPA and acute fluoxetine, produce opposite effects in each test [58]. Moreover, there is substantial evidence for different stimulus control in these tests [63], reinforcing the hypothesis that they model different aspects of anxiety-like behavior. While it is not fully understood whether exposure to the light/dark test could impact latter testing with the novel tank test, in principle both tests could be used in a 'test battery' of behavioral assays. This approach could greatly increase the information content and circumvent the limitation of analyzing a small amount of variables.
In conclusion, the present work, combined with other attempts at clustering behavioral variables and treatments in adult and larval zebrafish [15], [41], [42], suggest that behavioral screening is able to characterize relatively large classes of chemical compounds, revealing differences in efficacy and side effects (e.g., sedation) that cannot be detected in vitro.
Materials and Methods
Ethical statement
Animals were housed and manipulated in ways that minimized their potential suffering, as per the recommendations of the Canadian Council on Animal Care [6]. All procedures complied with the Brazilian Society for Neuroscience and Behavior’s (SBNeC) guidelines for the care and use of animals in research, and experiments were approved by the Comitê de Ética no Uso de Animais (CEUA) from UEPA.
Subjects and housing
430 adult zebrafish from the shortfin wild type phenotype were bought in a local ornamental fish shop and brought to the laboratory facilities, where the animals were left to acclimate for at least two weeks before experiments begun. Animals were group-housed in 40 L tanks, with a maximum density of 25 fish per tank. Tanks were filled with deionized and reconstituted water at room temperature (28°C) and a pH of 7.0–8.0. Lighting was provided by fluorescent lamps in a cycle of 14–10 hours (LD), according to the standards of zebrafish care [64].
Drugs
Anhydrous caffeine was bought from Quimis (Diadema/SP, Brazil). PACPX, DPCPX, ZM 241385 and DMPX were bought from Research Biochemicals International (Natick/MA, USA). Clonazepam, diazepam and moclobemide were bought from Roche (Brazil). Fluoxetine hydrochloride was bought from Eli Lily (Brazil). Buspirone hydrochloride, bupropion and diethylpropion were bought from Bristol-Myers Squibb (Brazil). Ethanol was bought from Cromoline (Brazil). Chlordiazepodixe was bought from Farmasa (Brazil). NMDA and serotonin were bought from Tocris (Bristol, UK). MK-801, WAY 100635, SB 224289, sodium nitroprusside and L-NAME were bought from Sigma (Saint Louis/MO, USA). Hypericum perforatum hydroalcoholic extract was a kind gift from Dr. Marcelo Pereira, and prepared as follows: above-ground parts of the plant were dried for 10 days at room temperature, after which they were ground by an atomic blender; 100 g of the plant powder was soaked in 96% ethanol for 72 h and then filtered and concentrated by a vacuum distiller. The concentrated solution was decanted chloroform in three consecutive steps, and the resulting solution was vaporized and desiccated at 50°C under sterile conditions. Drugs were dissolved on Cortland’s salt solution or 1% DMSO prior to experiments, and injected intraperitoneally at a volume of 0.1 µl per mg body weight with a 10 µl microsyringe equipped with a 33G needle (Hamilton, USA). Chronic fluoxetine treatment was made by daily injections for 14 days.
Light/dark preference
Determination of drug effects on scototaxis were carried as described elsewhere [65]. Briefly, after drug injection and effect onset animals were transferred to the central compartment of a black and white tank (15 cm×10 cm×45 cm h×d×l) for a 3-min. acclimation period, after which the doors which delimit this compartment were removed and the animal was allowed to freely explore the apparatus for 15 min. The following variables were recorded, along with the reference to their extended definition in the Zebrafish Behavior Catalog [17]:
time on the white compartment: the time spent in the top third of the tank (percentage of the trial) (ZBC 1.137);
squares crossed: the number of 10 cm2 squares crossed by the animal in the white compartment (ZBC 1.54);
latency to white: the amount of time the animal spends in the black compartment before its first entry in the white compartment (s);
entries in white compartment: the number of entries the animal makes in the white compartment in the whole session (ZBC 1.54);
erratic swimming: the number of “erratic swimming” events, defined as a zig-zag, fast, unpredictable course of swimming of short duration (ZBC 1.51);
freezing: the proportional duration of freezing events (in % of time in the white compartment), defined as complete cessation of movements with the exception of eye and operculae movements (ZBC 1.68).
thigmotaxis: the proportional duration of thigmotaxis events (in % of time in the white compartment), defined as swimming in a distance of 2 cm or less from the white compartment’s walls (ZBC 1.173).
risk assessment: the number of “risk assessment” events, defined as a fast (<1 s) entry in the white compartment followed by re-entry in the black compartment, or as a partial entry in the white compartment (i.e., the pectoral fin does not cross the midline).
Maximum Predictive Value calculations
For each variable analyzed, Maximum Predictive Values (MPVs) were calculated as the ratio of the mean difference between control and treatment groups and their pooled standard deviations [40]:
 |
Where pooled standard deviations are defined as
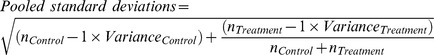 |
Clustering algorithm
After calculation of MPVs, these values were input into Cluster 3.0 (University of Tokyo, Japan), where hierarchical clustering was performed across behavioral endpoints and drug treatments and doses. Data were centered around the median, and clustering was then made using Spearman Rank Correlation with Average linkage as similarity metric. Clustering results were then visualized as dendograms and colored arrays in Java TreeView (University of Glasgow, UK).
HPLC analysis of indoleamines
Serotonin and 5-HIAA (5 mg) were dissolved in 100 mL of eluting solution (50 ml MilliQ water, 0.43 ml HClO4 70% [0.2 N], 10 mg EDTA, 9.5 mg sodium metabissulfite) and frozen at −20°C, to later be used as a standard.
The HPLC system consisted of a delivery pump (LC20-AT, Shimadzu), a 20 µL sample injector (Rheodyne), a degasser (DGA-20A5), and an analytical column (Shimadzu Shim-Pack VP-ODS, 250×4.6 mm internal diameter). The integrating recorder was a Shimadzu CBM-20A (Shimadzu, Kyoto, Japan). An electrochemical detector (Model L-ECD-6A) with glassy carbon was be used at a voltage setting of +0.83 V, with a sensitivity set at 8 nA full deflection. The mobile phase consisted of a solution of 70 mM phosphate buffer (pH 2.9), 0.2 mM EDTA, 5% methanol and 20% sodium metabissulfite as a conservative. The column temperature was set at 17°C, and the isocratic flow rate was 1.8 ml/min. 0.5 mL of extracellular fluid (ECF) were extracted by quickly removing one brain from the skull and incubating it in 2 mL of 50 mM TBS, pH 7.4, containing 90 mM NaCl, 2.5 mM CaCl2, 1 mM glutathione for 30 min at 4°C (7). This fluid was then mixed with 0.5 mL of eluting solution, filtered through a 0.22 µm syringe filter, and then injected into the HPLC system.
Funding Statement
This research was financed by Conselho Nacional de Pesquisa (CNPq/Brazil, grants 483336/2009-2 and 400039/2009-5). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Baldwin DS, Waldman S, Allgulander C (2011) Evidence-based pharmacological treatment of generalized anxiety disorder. Int J Neuropsychopharmacol 14: 697–710 10.1017/S1461145710001434 [DOI] [PubMed] [Google Scholar]
- 2. Griebel G, Holmes A (2013) 50 years of hurdles and hope in anxiolytic drug discovery. Nat Rev Drug Discov 12: 667–687 10.1038/nrd4075 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Tecott LH, Nestler EJ (2004) Neurobehavioral assessment in the information age. Nat Neurosci 7: 462–466 10.1038/nn1225 [DOI] [PubMed] [Google Scholar]
- 4. Crabbe JC, Morris RGM (2004) Festina lente: Late-night thoughts on high-throughput screening of mouse behavior. Nat Neurosci 7: 1175–1179 10.1038/nn1343 [DOI] [PubMed] [Google Scholar]
- 5. Brennand KJ, Simone A, Tran N, Gage FH (2012) Modeling psychiatric disorders at the cellular and network levels. Mol Psychiatry 17: 1239–1253 Available: 10.1038/mp.2012.20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Kokel D, Peterson RT (2008) Chemobehavioural phenomics and behaviour-based psychiatric drug discovery in the zebrafish. Briefings Funct Genomics Proteomics 7: 483–490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Gottesman II, Gould TD (2003) The endophenotype concept in psychiatry: Etymology and strategic intentions. Am J Psychiatry 160: 636–645. [DOI] [PubMed] [Google Scholar]
- 8. Van der Staay FJ (2006) Animal models of behavioral dysfunctions: Basic concepts and classifications, and an evaluation strategy. Brain Res Rev 52: 131–159 Available: http://www.ncbi.nlm.nih.gov/pubmed/16529820. [DOI] [PubMed] [Google Scholar]
- 9. Arguello PA, Gogos JA (2006) Modeling madness in mice: One piece at a time. Neuron 52: 179–196 10.1016/j.neuron.2006.09.023 [DOI] [PubMed] [Google Scholar]
- 10. Gould TD, Gottesman II (2006) Psychiatric endophenotypes and the development of valid animal models. Genes, Brain Behav 5: 113–119 10.1111/j.1601183X.2005.00186.x [DOI] [PubMed] [Google Scholar]
- 11.Stewart AM, Kalueff A V (2014) Developing better and more valid animal models of brain disorders. Behav Brain Res In press. [DOI] [PubMed]
- 12. Kalueff AV, Ren-Patterson RF, LaPorte JL, Murphy DL (2008) Domain interplay concept in animal models of neuropsychiatric disorders: A new strategy for high-throughput neurophenotyping research. Behav Brain Res 188: 243–249 10.1016/j.bbr.2007.11.011 [DOI] [PubMed] [Google Scholar]
- 13. Kari G, Rodeck U, Dicker AP (2007) Zebrafish: An emerging model system for human disease and drug discovery. Clin Pharmacol Ther 82: 70–80 10.1038/sj.clpt.6100223 [DOI] [PubMed] [Google Scholar]
- 14. Ahmad F, Noldus LPJJ, Tegelenbosch RAJ, Richardson MK (2012) Zebrafish embryos and larvae in behavioural assays. Behaviour 149: 1241–1281 10.1163/1568539X-00003020 [DOI] [Google Scholar]
- 15. Cachat J, Stewart A, Utterback E, Hart P, Gaikwad S, et al. (2011) Three-dimensional neurophenotyping of adult zebrafish behavior. PLoS One 6: e17597 10.1371/journal.pone.0017597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Zon LI, Peterson RT (2005) In vivo drug discovery in the zebrafish. Nat Rev Drug Discov 4: 35–44 10.1038/nrd1606 [DOI] [PubMed] [Google Scholar]
- 17. Kalueff AV, Gebhardt M, Stewart AM, Cachat JM, Brimmer M, et al. (2013) Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 10: 70–86 10.1089/zeb.2012.0861 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Stewart AM, Kalueff AV (2012) The developing utility of zebrafish models for cognitive enhancers research. Curr Neuropharmacol In press. [DOI] [PMC free article] [PubMed]
- 19. Nguyen M, Poudel MK, Stewart AM, Kalueff AV (2013) Skin too thin? The developing utility of zebrafish skin (neuro)pharmacology for CNS drug discovery research. Brain Res Bull 98: 145–154 10.1016/j.brainresbull.2013.08.004 [DOI] [PubMed] [Google Scholar]
- 20. Cachat J, Canavello P, Elegante M, Bartels B, Hart P, et al. (2010) Modeling withdrawal syndrome in zebrafish. Behav Brain Res 208: 371–376. [DOI] [PubMed] [Google Scholar]
- 21. Shimada Y, Hirano M, Nishimura Y, Tanaka T (2012) A high-throughput fluorescence-based assay system for appetite-regulating gene and drug screening. PLoS One 7: e52549 10.1371/journal.pone.0052549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Hill AJ, Teraoka H, Heideman W, Peterson RE (2005) Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol Sci 86: 6–19 10.1093/toxsci/k [DOI] [PubMed] [Google Scholar]
- 23. Gerlai R (2010) High-throughput behavioral screens: The first step towards finding genes involved in vertebrate brain function using zebrafish. Molecules 15: 2609–2622 10.3390/molecules15042609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Key B, Devine CA (2003) Zebrafish as an experimental model: Strategies for developmental and molecular neurobiology studies. Methods Cell Sci 25: 1–6 10.1023/B:MICS.0000006849.98007.03 [DOI] [PubMed] [Google Scholar]
- 25. Redfern WS, Waldron G, Winter MJ, Butler P, Holbrook M, et al. (2008) Zebrafish assays as early safety pharmacology screens: Paradigm shift or red herring? J Pharmacol Toxicol Methods 58: 110–117 10.1016/j.vascn.2008.05.006 [DOI] [PubMed] [Google Scholar]
- 26. Ahmad F, Noldus LPJJ, Tegelenbosch RAJ, Richardson MK (2012) Zebrafish embryos and larvae in behavioural assays. Behaviour 149: 1241–1281 10.1163/1568539X-00003020 [DOI] [Google Scholar]
- 27. Maximino C, Herculano AM (2010) A review of monoaminergic neuropsychopharmacology in zebrafish. Zebrafish 7: 359–378 10.1089/zeb.2010.0669 [DOI] [PubMed] [Google Scholar]
- 28. Norton W, Bally-Cuif L (2010) Adult zebrafish as a model organism for behavioural genetics. BMC Neurosci 11: 90 Available: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2919542&tool=pmcentrez&rendertype=abstract. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Schilling TF (2002) Morphology of larval and adult zebrafish. In: Nüsslein-Volhard C, Dahm R, editors. Zebrafish: A Practical Approach. Oxford: Oxford University Press. 59–94. [Google Scholar]
- 30. Levin ED, Bencan Z, Cerutti DT (2007) Anxiolytic effects of nicotine in zebrafish. Physiol Behav 90: 54–58 10.1016/j.physbeh.2006.08.026 [DOI] [PubMed] [Google Scholar]
- 31. Egan RJ, Bergner CL, Hart PC, Cachat JM, Canavello PR, et al. (2009) Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav Brain Res 205: 38–44 10.1016/j.bbr.2009.06.022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Stewart A, Wu N, Cachat J, Hart P, Gaikwad S, et al. (2011) Pharmacological modulation of anxiety-like phenotypes in adult zebrafish behavioral models. Prog Neuropsychopharmacol Biol Psychiatry 35: 1421–1431 10.1016/j.pnpbp.2010.11.035 [DOI] [PubMed] [Google Scholar]
- 33. Blaser RE, Goldsteinholm K (2012) Depth preference in zebrafish, Danio rerio: Control by surface and substrate cues. Anim Behav 83: 953–959 10.1016/j.anbehav.2012.01.014 [DOI] [Google Scholar]
- 34. Maximino C, Silva AWB da, Gouveia A Jr, Herculano AM (2011) Pharmacological analysis of zebrafish (Danio rerio) scototaxis. Prog Neuropsychopharmacol Biol Psychiatry 35: 624–631 10.1016/j.pnpbp.2011.01.006 [DOI] [PubMed] [Google Scholar]
- 35.Araujo J, Maximino C, Brito TM de, Silva AWB da, Batista E de JO, et al. (2012) Behavioral and pharmacological aspects of anxiety in the light/dark preference test. In: Kalueff A V, Stewart AM, editors. Zebrafish Protocols for Neurobehavioral Research. New York: Humana Press. doi:10.1007/978-1-61779-597-8_14. [Google Scholar]
- 36.Stewart A, Maximino C, de Brito TM, Herculano AM, Gouveia A Jr, et al. (2011) Neurophenotyping of adult zebrafish using the light/dark box paradigm. In: Kalueff A V, Cachat JM, editors. Zebrafish Neurobehavioral Protocols. New York: Humana Press. 157–167. doi:10.1007/978-1-60761-953-6_13. [Google Scholar]
- 37. Maximino C, Brito TM, Colmanetti R, Pontes AAA, Castro HM de, et al. (2010) Parametric analyses of anxiety in zebrafish scototaxis. Behav Brain Res 210: 1–7 10.1016/j.bbr.2010.01.031 [DOI] [PubMed] [Google Scholar]
- 38.Maximino C (2011) Parâmetros da escototaxia como modelo comportamental de ansiedade no paulistinha (Danio rerio, Cyprinidae, Pisces) Universidade Federal do Pará. doi:10.6084/m9.figshare.928644.
- 39. Serra EL, Medalha CC, Mattioli R (1999) Natural preference of zebrafish (Danio rerio) for a dark environment. Brazilian J Med Biol Res 32: 1551–1553. [DOI] [PubMed] [Google Scholar]
- 40. Maximino C, Brito TM De, Dias CA de M, Gouveia A Jr, Morato S (2010) Scototaxis as anxiety-like behavior in fish. Nat Protoc 5: 209–216 10.1038/nprot.2009.225 [DOI] [PubMed] [Google Scholar]
- 41. Rihel J, Prober DA, Arvanites A, Lam K, Zimmerman S, et al. (2010) Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation. Science (80-) 347: 348–351 10.1126/science.1183090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Kokel D, Bryan J, Laggner C, White R, Cheung CYJ, et al. (2010) Rapid behavior-based identification of neuroactive small molecules in the zebrafish. Nat Chem Biol 6: 231–237 10.1038/nchembio.307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Stewart AM, Cachat J, Green J, Gaikwad S, Kyzar E, et al. (2013) Constructing the habituome for phenotype-driven zebrafish research. Behav Brain Res 236: 110–117 10.1016/j.bbr.2012.08.026 [DOI] [PubMed] [Google Scholar]
- 44.Linker A, Stewart A, Gaikwad S, Cachat JM, Elegante MF, et al. (2011) Assessing the Maximum Predictive Validity for neuropharmacological anxiety screening assays using zebrafish. In: Kalueff A V, Cachat JM, editors. Zebrafish Neurobehavioral Protocols. New York: Humana Press. 181–190. [Google Scholar]
- 45.Herculano AM, Maximino C (2014) Serotonergic modulation of zebrafish behavior: Towards a paradox. Prog Neuropsychopharmacol Biol Psychiatry In press. doi:10.1016/j.pnpbp.2014.03.008. [DOI] [PubMed]
- 46. Papke RL, Ono F, Stokes C, Urban JM, Boyd RT (2012) The nicotinic acetylcholine receptors of zebrafish and an evaluation of pharmacological tools used for their study. Biochem Pharmacol 84: 352–365 Available: 10.1016/j.bcp.2012.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Sackerman J, Donegan JJ, Cunningham CS, Nguyen NN, Lawless K, et al. (2010) Zebrafish behavior in novel environments: Effects of acute exposure to anxiolytic compounds and choice of Danio rerio line. Int J Comp Psychol 23: 43–61. [PMC free article] [PubMed] [Google Scholar]
- 48. Norton WHJ, Stumpenhorst K, Faus-Kessler T, Folchert A, Rohner N, et al. (2011) Modulation of fgfr1a signaling in zebrafish reveals a genetic basis for the aggression–boldness syndrome. J Neurosci 31: 13796–13807 10.1523/JNEUROSCI.2892-11.2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Peitsaro N, Kaslin J, Anichtchik O V, Panula P (2003) Modulation of the histaminergic system and behaviour by α-fluoromethylhistidine in zebrafish. J Neurochem 86: 432–441 10.1046/j.1471-4159.2003.01850.x [DOI] [PubMed] [Google Scholar]
- 50. Cofiel LP V, Mattioli R (2009) L-histidine enhances learning in stressed zebrafish. Brazilian J Med Biol Res 42: 128–134. [DOI] [PubMed] [Google Scholar]
- 51. Bencan Z, Sledge D, Levin ED (2009) Buspirone, chlordiazepoxide and diazepam effects in a zebrafish model of anxiety. Pharmacol Biochem Behav 94: 75–80 10.1016/j.pbb.2009.07.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Richendrfer H, Pelkowski SD, Colwill RM, Creton R (2011) On the edge: Pharmacological evidence for anxiety-related behavior in zebrafish larvae. Behav Brain Res. 10.1016/j.bbr.2011.11.041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Steenbergen PJ, Richardson MK, Champagne DL (2011) Patterns of avoidance behaviours in the light/dark preference test in young juvenile zebrafish: A pharmacological study. Behav Brain Res 222: 15–25 10.1016/j.bbr.2011.03.025 [DOI] [PubMed] [Google Scholar]
- 54. Lau BYB, Mathur P, Gould GG, Guo S (2011) Identification of a brain center whose activity discriminates a choice behavior in zebrafish. Proc Natl Acad Sci U S A 108: 2581–2586 10.1073/pnas.1018275108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Stewart AM, Kalueff AV (2014) The behavioral effects of acute Δ9-tetrahydrocannabinol and heroin (diacetylmorphine) exposure in adult zebrafish. Brain Res 1543: 109–119 Available: 10.1016/j.brainres.2013.11.002. [DOI] [PubMed] [Google Scholar]
- 56. Connors KA, Valenti TW, Lawless K, Sackerman J, Onaivi ES, et al. (2013) Similar anxiolytic effects of agonists targeting serotonin 5-HT1A or cannabinoid CB1 receptors on zebrafish behavior in novel environments. Aquat Toxicol. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Investigation O (2014) Acute administration of THC impairs spatial but not associative memory function in zebrafish. Psychopharmacology (Berl) In press. 10.1007/s00213-014-3522-5 [DOI] [PubMed] [Google Scholar]
- 58. Maximino C, Puty B, Benzecry R, Araújo J, Lima MG, et al. (2013) Role of serotonin in zebrafish (Danio rerio) anxiety: Relationship with serotonin levels and effect of buspirone, WAY 100635, SB 224289, fluoxetine and para-chlorophenylalanine in two behavioral models. Neuropharmacology 71: 83–97 10.1016/j.neuropharm.2013.03.006 [DOI] [PubMed] [Google Scholar]
- 59. Herculano AM, Maximino C (2014) Serotonergic modulation of zebrafish behavior: Towards a paradox. Prog Neuropsychopharmacol Biol Psychiatry In press. 10.1016/j.pnpbp.2014.03.008 [DOI] [PubMed] [Google Scholar]
- 60. Stewart AM, Cachat J, Gaikwad S, Robinson K, Gebhardt M, et al. (2013) Perspectives on experimental models of serotonin syndrome in zebrafish. Neurochem Int 62: 893–902 10.1016/j.neuint.2013.02.018 [DOI] [PubMed] [Google Scholar]
- 61. Wong K, Elegante M, Bartels B, Elkhayat S, Tien D, et al. (2010) Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav Brain Res 208: 450–457 10.1016/j.bbr.2009.12.023 [DOI] [PubMed] [Google Scholar]
- 62. Maximino C, Lima MG, Oliveira KRM, Picanço-diniz DLW, Herculano AM (2011) Adenosine A1, but not A2, receptor blockade increases anxiety and arousal in zebrafish. Basic Clin Pharmacol Toxicol 109: 203–207 10.1111/j.1742-7843.2011.00710.x [DOI] [PubMed] [Google Scholar]
- 63. Maximino C, Benzecry R, Matos KRO, Batista E de JO, Herculano AM, et al. (2012) A comparison of the light/dark and novel tank tests in zebrafish. Behaviour 149: 1099–1123 10.1163/1568539X-00003029 [DOI] [Google Scholar]
- 64.Canadian Council on Animal Care (2005) CCAC guidelines on: The care and use of fish in research, teaching and testing. Available: http://www.ccac.ca/en/CCAC_Programs/Guidelines_Policies/GDLINES/Fish/Fish_Guidelines_English.pdf.
- 65. Lawrence C (2007) The husbandry of zebrafish (Danio rerio): A review. Aquaculture 269: 1–20 10.1016/j.aquaculture.2007.04.077 [DOI] [Google Scholar]