

Abstract
Purpose
Formation of epiretinal membranes (ERMs) in the posterior fundus results in visual impairment. ERMs have been associated with numerous clinical conditions, including proliferative diabetic retinopathy (PDR), a neovascular disease. Apelin has been identified as a novel angiogenesis contributor. The aim of this study was to investigate the correlation between apelin and ERMs after PDR.
Methods
ERM samples were obtained by vitrectomy from 12 subjects with PDR (aged 57±6 years; duration of diabetes 16±7 years), and 12 subjects with idiopathic ERM (aged 68±5 years). The samples were processed for immunohistochemistry and reverse transcription–PCR (RT–PCR). We also analyzed samples from patients with PDR who received an intravitreal injection of bevacizumab (IVB) before vitrectomy.
Results
The mRNA expression of apelin was significantly higher in the PDR ERMs than in the idiopathic ERMs. Accordingly, immunohistochemical analysis revealed strong expression of apelin in all eight PDR ERMs without IVB, and was double-labeled with glial fibrillary acidic protein antibody (GFAP), platelet endothelial cell adhesion molecule-1 (CD31), cytokeratin (CK) and vascular endothelial growth factor (VEGF) but not with fibronectin. They were mainly located in the adventitia. In contrast, the expression of apelin was lower in the PDR ERMs after IVB and the idiopathic ERMs.
Conclusions
The results showed that apelin was involved in the formation of ERMs and promoted the formation of adventitia, including glial, endothelial, and RPE cells. Bevacizumab blocked the expression of apelin and regressed gliosis and angiogenesis.
Introduction
Epiretinal membranes (ERMs) involving the macular or perimacular regions can cause a reduction in vision, metamorphopsia, micropsia, or occasionally monocular diplopia. The incidence of ERMs increases with age and may approach 20% of the total population by age 70 [1]. The presence of ERMs has been associated with various clinical conditions, including proliferative diabetic retinopathy (PDR) [2]. The prevalence of ERMs in PDR was reported to be 20% in type 1 diabetes and 5% in type 2 diabetes [3,4]. Histopathological studies showed that ERMs were composed of various cell types such as glial cells, fibroblasts, and endothelial cells [1,5,6,7], but the pathogenic mechanisms are still unknown. However, different peptide factors, including cytokines and growth factors, have been detected in ERMs and vitreous fluid, coincident with ocular diseases such as PDR [5-9]. Because some peptide factors are soluble mediators of angiogenesis, diabetic neovascularization might be caused or aggravated by these factors [8].
In recent years, apelin signaling has been identified as an important contributor to angiogenesis [10-20]. Apelin is crucial for embryonic vascular development [10-12] and for the postnatal formation of retinal vessels [13-16]. Apelin promotes angiogenesis in vitro and vivo [16-20] and stimulates endothelial cell proliferation, migration, and tube formation in vitro [15,17,18]. The expression of apelin was upregulated during tumor neovascularization, and overexpression was reported to increase in vivo tumor growth [11,19,20]. Consequently, apelin signaling could represent an interesting new therapeutic target during pathological neovascularization [21-24]. Since apelin has been recognized in previous findings as a factor contributing to angiogenesis, and its role has not been examined in PDR, in the present study we examined the expression of apelin and related factors in ERMs obtained from patients with PDR.
Methods
This study adhered to the tenets of the Declaration of Helsinki (human subjects) and to the ARVO Statement on human subject research. We received institutional approval from the review committee of the People’s Hospital affiliated with Peking University. Informed consent was obtained from each patient to collect samples. Inclusion criteria included (1) younger than 70 years old, (2) absence of renal or hematological diseases or uremia, (3) no administration of chemotherapy or life-support measures, and (4) the fewest possible chronic pathologies other than diabetes. Exclusion criteria included (1) systematic or ocular historical treatment with anti-VEGF therapy, (2) previous ocular surgery, (3) a history of tumor, and (4) a history of other neovascular ocular diseases. The ERMs were surgically removed from consecutive eyes with secondary PDR and idiopathic ERMs as the control subjects, who underwent pars plana vitrectomy and membrane peeling. Samples derived from 12 patients with PDR (aged 57±6 years, duration of diabetes 16±7 years) and 12 patients with idiopathic ERM (aged 68±5 years) were processed for reverse transcription (RT)–PCR and immunofluorescence staining.
In addition, according to the previous treatment in the reported articles [25-29] and our experience [30,31], 1.25 mg/0.05 ml of bevacizumab (Avastin; Genentech, South San Francisco, CA) was injected into the vitreous cavity in eight samples (aged 51±8 years, duration of diabetes 14±7 years) from patients with PDR as preoperative adjunctive therapy 7 days before vitrectomy. A topical antibiotic was prescribed 3 days before the bevacizumab injection. Anesthetic drops were instilled followed by standard surgical preparation with 0.5% povidone-iodine solution and insertion of a sterile lid speculum. Bevacizumab was injected with a sharp 30-gauge needle through pars plana 4.0 mm from the limbus supertemporally or supernasally. All eyes were examined 1 day after injection particularly for the anterior chamber and posterior segment reaction. Topical antibiotic was prescribed to use for 7 days after IVB.
RNA extraction and amplification with reverse transcription–PCR
Total cellular RNA was prepared using an extraction reagent (TRIzol; Invitrogen), and 2 μg of retina RNA was converted into cDNA in a total reaction volume of 25 µl, containing 1 µg Oligo (dT), 5µl M-MLV 5× reaction buffer, 1.25 µl dNTP, 25 units Recombinant RNasin Ribonuclease Inhibitor, and 200 units M-MLV reverse transcriptase. The mixture was incubated for 60 min at 42 °C and terminated by heating at 95 °C for 5 min. RT–PCR analysis was performed, as previously described [32]. A volume of 1 μl of each cDNA product from the reverse transcription procedure was used as the template for PCR amplification in a reaction mixture containing PCR buffer (10 mmol/l Tris-HCl, pH 8.3, 50 mmol/l KCl, 1.5 mmol/l MgCl), 0.2 mmol/l dNTPs, a 0.2 mmol/l set of oligonucleotide primers, and 2.5 units Taq DNA polymerase in a final volume of 50μl. cDNA reverse-transcribed from total RNA was amplified by using primers specific for human apelin (sense, 5′-CAC CTC GCA CCT GCT GTA-3′; anti-sense, 5′-GAA CGG GAA TCA TCC AAA C-3′; 119 bp) and human GAPDH (sense, 5′-TTG ACG CTG GGG CTG GCA TT-3′; anti-sense, 5′-TGG AGG CCA TGT GGG CCA TGA-3′; 117 bp). PCR was performed after initial denaturation at 95 °C for 3 min. Each cycle consisted of a heat-denaturation step at 95 °C for 20 s, annealing of primers at either 63 °C (apelin and GAPDH) for 15s, and then polymerization at 72 °C for 15 s. Negative controls for PCR were performed using “templates” derived from RT reactions lacking either reverse transcriptase or total RNA. After 35 cycles, 15 μl of each reaction mixture were electrophoresed on a 10% Tris-borate-EDTA agarose gel and stained with ethidium bromide. Results of mRNA were quantified indirectly using Glyko BandScan (ProZyme, version 5.0, San Leandro, CA) to analyze the grayscale image.
Hematoxylin and eosin staining and immunofluorescence staining
These samples were embedded in optimum cutting temperature compound (Miles Laboratories, Naperville, IL), flash-frozen in liquid nitrogen, and then stored at −80 °C. Routine hematoxylin and eosin (H&E) staining was performed and examined with light microscopy. Frozen sections (6 µm thick) were cut with a cryostat, mounted on 3-aminopropyltriethoxysilane-coated glass slides, and air-dried overnight at room temperature. The sections were fixed sequentially with 4% paraformaldehyde /4% sucrose in PBS (1X; 137 mM NaCl, 2.7 mM KCl, 4.3 mM NaH2PO4, 1.47 mM KH2PO4, pH 7.4; 20 min), washed (3×, PBS), 100% methanol (10 min), and 0.2% Triton-X100 (10 min). Blocking was performed with 10% goat serum/PBS (1 h, 23 °C) to block nonspecific staining. Primary antibodies were diluted into 10% goat serum/PBS and incubated overnight at 4 °C. The following antibody was used: rabbit polyclonal anti-apelin antibody (1:200, No. ab59469; Abcam, Cambridge, MA). For double-labeling immunofluorescence studies, the antibodies were then incubated with a monoclonal mouse antiglial fibrillary acidic protein antibody (GFAP; 1:150 dilution; Zhongshan Goldenbridge Biotechnology, Beijing, China), a monoclonal mouse anti-PECAM-1 (platelet endothelial cell adhesion molecule-1, CD31) antibody (1:150 dilution; Zhongshan Goldenbridge Biotechnology), a monoclonal mouse anti-cytokeratin (CK) antibody (1:150 dilution; Zhongshan Goldenbridge Biotechnology), a monoclonal mouse anti-fibronectin (FN) antibody (extracellular matrix, ECM), (1:150 dilution; Zhongshan Goldenbridge Biotechnology), and a monoclonal mouse anti-VEGF antibody (1:100; No. sc-7269; Santa Cruz, CA). After blocking, the sections were washed (3×, PBS) and then incubated with secondary antibodies diluted in 20% FBS, 10% goat serum, and PBS, respectively (1 h, 37 °C). Secondary antibodies used fluorescein isothiocyanate–conjugated goat anti-mouse-tetramethyl rhodamine isothiocyanate (1:200; No. ZF-0312; Zhongshan Goldenbridge Biotechnology) and cyanogen (CY) 3-conjugated goat anti-rabbit-fluorescein isothiocyanate (1:200 dilution; No. BA1032, Sigma, Carlsbad, CA). The samples were counterstained with 4', 6-diamidino-2-phenylindole (DAPI; 1:1,000; No. D9542, Sigma) and then covered with a non-fluorescent sealant. Immunofluorescence was viewed using a fluorescence microscope (DS-Ri1-U2, Nikon, Japan) and images acquired using a DS-U2u camera with NIS-Elements Imaging Software.
Statistical analysis
The results were expressed as the means±standard error of the mean (SEM), except as noted. The χ2 test was used to test for significance of the difference between genders in the PDR group and the idiopathic group. Differences between the PDR group and the idiopathic group were estimated with the nonparametric Mann–Whitney rank-sum test and the Student t test when appropriate. The statistical analysis was performed using a commercially available statistical software package (SPSS for Windows, version 17.0, SPSS, Chicago, IL). p<0.05 was considered statistically significant. Experiments were performed at least three times.
Results
Samples derived from 12 patients with PDR ERMs (four women, aged 57±6 years, duration of diabetes 16±7 years) and 12 patients with idiopathic ERM (six women, aged 68±5 years) were processed for RT–PCR and immunofluorescence staining. Age and gender did not vary significantly between the PDR group and the idiopathic group (p>0.05, p>0.05). In addition, 1.25 mg/0.05 ml of bevacizumab was injected into the vitreous cavity as preoperative adjunctive therapy 7 days before vitrectomy in eight samples (aged 51±8 years, duration of diabetes 14±7 years) of the PDR group.
Expression of apelin in ERMs was examined with RT–PCR analysis (Figure 1). mRNA encodings were highly expressed as apelin in patients with PDR. The expression of apelin was detected in 12 of 12 (100%) patients with PDR, but in only four of 12 (33%) control subjects (p<0.001; Figure 1). Semi-quantitative analysis was performed based on the gray scale ratio, which revealed that apelin in the PDR group was 7.81±0.54 versus 0.42±0.30 in the idiopathic group, and showed statistically difference between the two groups (t=4.338, p<0.001).
Figure 1.
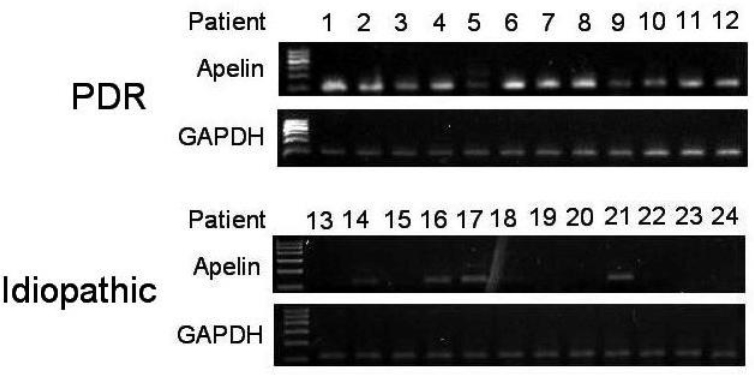
RT–PCR analysis of apelin in proliferative diabetic retinopathy (PDR) epiretinal membranes (ERMs) and idiopathic epiretinal membranes. Lanes 1–12 are samples from the PDR group, and lanes 13–24 are samples from the idiopathic group. Results were quantified indirectly using BandScan to analyze the grayscale image. Semi-quantitative analysis was performed based on the gray scale ratio, which revealed that the apelin in the PDR ERMs group was 7.81±0.54 versus 0.42±0.30 in idiopathic ERMs group, and showed statistically difference between the two groups (t=4.338, P<0.001).
Histopathological examinations
The ERMs from patients with PDR were composed of densely cellular tissue (Figure 2A) or highly vascularized tissue (Figure 2B) and consisted of cellular components, such as retinal pigment epithelial cells, glial cells, fibroblasts, myofibroblasts, endothelial cells, and other cells. The specimens from control subjects showed the crimped nature of the collagen fibers and the sparse cellular components (Figure 2C).
Figure 2.
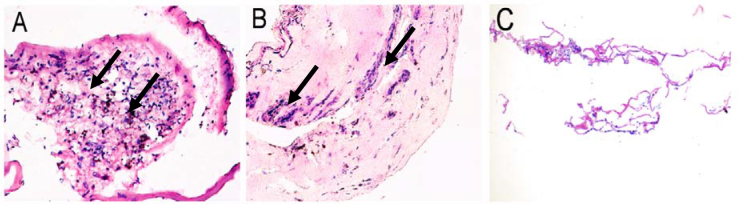
Histopathologic findings in fibrovascular membranes of proliferative diabetic retinopathy (PDR; A, B) and in idiopathic epiretinal membranes (ERMs; C). A: H&E staining shows densely cellular tissue in ERMs from PDR patients (arrow). B: H&E staining shows highly vascularized tissue and large-calibre vessels and gliosis in ERMs from patients with PDR (arrow). C: H&E staining shows sparse cellular tissue in idiopathic ERMs derived from the control subjects.
Immunofluorescence staining in epiretinal membranes
Immunohistochemical analysis was performed to identify the apelin protein expression in the PDR ERMs and idiopathic ERMs (Figure 3, Figure 4). Strong staining of apelin was detected in the specimens of all fibrovascular membranes from patients with PDR (Figure 3A,D,G, and Figure 4A). No apelin was detected in the idiopathic ERMs (Figure 4D) and weak staining of apelin in the membranes from patients with PDR after intravitreal injection of bevacizumab (Figure 4G); meanwhile, we also found large-caliber vessels and fibroglial tissue in ERMs regressed after intravitreal injection of bevacizumab. In addition, we examined whether apelin is coexpressed with the glial cell-specific marker GFAP. The ERMs after PDR contained a large area composed of glial cells (Figure 4B), and many cells in that area were labeled with anti-GFAP and antiapelin (Figure 4C). Similar results were obtained in experiments with vascular endothelial marker CD31 (Figure 3F), RPE cell maker cytokeratin (CK; Figure 3C), and VEGF (Figure 3I), but anti-fibronectin did not label sites reactive with antiapelin. The antibodies were not expressed in these tissues. No immunostaining was observed in the negative controls, which were performed by substituting the primary antiserum with PBS (Figure 5).
Figure 3.

Immunofluorescence staining for apelin, cytokeratin (CK), platelet endothelial cell adhesion molecule-1 (CD31), and vascular endothelial growth factor (VEGF) in proliferative diabetic retinopathy (PDR) epiretinal membranes. Cryosections were double-probed with antibodies against apelin (A, D, G) and (B) CK, (E) CD31, and (H) VEGF and detected using fluorochrome-conjugated secondary antibodies. Nuclei were labeled using 4', 6-diamidino-2-phenylindole (DAPI). Merged images (C, F, I) contain three colour channels representing apelin (red), CK, CD31, and VEGF (green), and DAPI (blue). The arrow in C, F, I showed apelin was co-expressed with CK, CD31, and VEGF in in ERMs from patients with PDR, respectively. Scale bar represents 50 μm.
Figure 4.

Indirect immunofluorescence evaluation of apelin and glial fibrillary acidic protein distribution in human epiretinal membranes (ERMs) derived from patients with proliferative diabetic retinopathy (PDR; A, B, C). Apelin and glial fibrillary acidic protein (GFAP) distribution in idiopathic epiretinal membranes (ERMs) from the control subjects (D, E, F). ERMs from patients with proliferative diabetic retinopathy (PDR) after intravitreal injection of bevacizumab (G, H, I). Cryosections were double-probed with antibodies against apelin (A, D, G), GFAP (B), and GFAP (E, H), and were detected with fluorochrome-conjugated secondary antibodies. Nuclei were detected using 4', 6-diamidino-2-phenylindole (DAPI) (C, F, I). The arrow in C showed apelin was co-expressed with GFAP. The arrow in F and I showed apelin was not expressed in idiopathic ERMs and ERMs from PDR patients after intravitreal injection of bevacizumab, respectively. Scale bar represents 100 μm.
Figure 5.

Fluorescence immunostaining of human epiretinal membranes (ERMs) in negative controls. No immunostaining was observed in negative controls (arrow), which were performed by substituting the primary antiserum with phosphate buffered saline. Secondary antibodies were used cyanogen (CY) 3-conjugated goat anti-rabbit-fluorescein isothiocyanate (A) and FITC-conjugated goat anti-mouse-tetramethyl rhodamine isothiocyanate (B). Nuclei were detected using DAPI. Merged images (C), DAPI (blue). Scale bar represents 100 μm.
We also found adventitia in the PDR ERMs, which were consistent with other experiments previously reported. Many cell components that highly expressed apelin closely surrounded the newly formed vessels in the ERMs from patients with PDR (Figure 3F). Furthermore, we detected several related factors with dual-color immunofluorescence staining in these specimens. Our results showed that many cell types surround the new vessels, such as retinal pigment epithelial cells (Figure 3C), glial cells (Figure 4C), fibroblasts, and myofibroblasts. Furthermore, ECM with the cell types participated in constituting adventitia (Figure 6). All cells and ECM protein closely surround new vessels to form a microenvironment in which cells secrete growth factors or cytokines that promoted vascular formation, such as VEGF in ERMs from patients with PDR.
Figure 6.
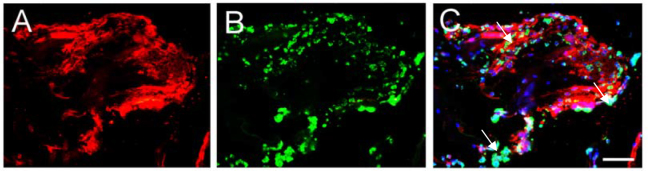
Indirect immunofluorescence evaluation of apelin and fibronectin distribution in human epiretinal membranes (ERMs) derived from patients with proliferative diabetic retinopathy (PDR). Cryosections were double-probed with antibodies against (A) apelin and (B) fibronectin. Nuclei were detected using 4', 6-diamidino-2-phenylindole (DAPI). C: Merged images contain three color channels representing apelin (red), fibronectin (green), and DAPI (blue). The arrow showed apelin was not co-expressed with fibronectin in ERMs from PDR patients. Scale bar represents 100 μm.
Discussion
The results of the present study showed that the expression of apelin mRNA was significantly higher in the PDR ERMs than in the idiopathic ERMs. In addition, the expression of apelin was strongly positive in ERMs from PDR and coexpressed with glial cell-specific markers, vascular endothelial cells markers, and RPE cell markers but not with FN. Recent findings showed that apelin was implicated in glial and vessel differentiation [14-20] and the expression of apelin was higher in the vascular system, particularly in vascular endothelial cells [18,21], and upregulated at the leading edge of vessel formation [13]. Furthermore, a recent report showed the angiogenic activity of apelin in Matrigel experiments, which indicated apelin was a novel angiogenic factor in retinal endothelial cells [15]. Moreover, in our study, the coexpression of apelin and VEGF in ERMs from PDR suggested that two factors may work together synergistically in angiogenesis and gliosis. From the positive staining of apelin in the endothelial cells, glial cells, and RPE cells, we might infer that the increased apelin was due to local production of apelin, presumably as an autocrine function of the retinal cells.
Recent evidence showed that diabetic retinopathy also affects the glial and neural cells of the retina [33,34]. Retinal glial cells might be associated with retinal dysfunctions such as PDR and DR [35-37]. Reactive changes in glial cells such as an upregulation of GFAP occur early during the course of the disease and precede the onset of overt vascular changes [38,39]. Müller cells are an important constituent of the fibroproliferative tissue formed during PDR [40] and produce growth factors, which activate vascular endothelial cells [41-43]. The occurrence of ERMs in PDR may contribute to the upregulation of growth factors secondary to the changes in Müller cell function [44,45]. Our study showed that apelin was colocalized with GFAP in ERMs from patients with PDR other than the control subjects. We believe our results indicate that the formation of a mixed cellular microenvironment around the new vessels by glial cell proliferation is a consequence of elevated apelin expression.
In our study, we also confirmed adventitia in the ERMs of PDR. Adventitia plays an important role in the neural network, endocrine system, metabolism, immune regulation, damage repair, and regeneration of tissue. Adventitia participates not only in vascular oxidative stress, inflammation, vascular remodeling, and homeostasis, but also as “initiating factors” in various vascular diseases [46-48]. Adventitia plays an important role in vascular biology, and can differentiate into endothelial cells, smooth muscle cells, and mesangial cells, participate in repairing vascular injury, and cause neointimal lesions [49,50]. Our studies showed that many cell types closely surrounded the new vessels, such as retinal pigment epithelial cells, glial cells, fibroblasts, myofibroblasts, and ECM, and these components highly expressed apelin. These phenomena implied a local microenvironment around new vessels, in which cells secrete angiogenic factors or cytokines, such as apelin, VEGF, promoting vascular formation and stimulating the vascular components released by angiogenic factors. Kidoya et al. [20] found that apelin upregulated the expression of adhesion molecules and promoted cell aggregation, and this function was not dependent on cell expansion. Thus, we presumed that the formation of a microenvironment around a new vessel might be caused by the ability of apelin.
The mechanism of ERM formation in PDR is still unknown, but the first step is thought to be neovascularization. In this process, many angiogenic factors play an important role [5,7-9,51]. Previous studies have reported that apelin and VEGF have positive synergistic effects, where increased expression of one can contribute to the expression of the other [52]. In previous research, we found that apelin, GFAP, and VEGF mRNA and protein levels were significantly increased in the retinas of diabetic rats [53]. Moreover, apelin induced GFAP and VEGF expression. F13A, a specific antagonist of apelin, suppressed GFAP and VEGF expression in vivo. It is consistent with studies that showed knockout apelin or the APJ gene can inhibit hypoxia-induced cell proliferation; this inhibition is not dependent on the VEGF signaling pathway [54]. We also detected plasma and vitreous concentrations of apelin with and without IVB in patients with PDR [30]. No significant differences were found in vitreous or plasma apelin concentrations between the two groups. The results suggest that apelin may not be directly regulated by VEGF. In this study, we found that bevacizumab decreased the expression of apelin and induced vascular and gliosis regression. Our team also recently reported that apelin is suppressed by bevacizumab in central retinal vein occlusion [55]. Therefore, based on the present findings, it is hard to say by which specific mechanism apelin and VEGF interacted. It seems reasonable to assume that apelin and VEGF may be mutually promoted during the development of DR.
In our experiment, the mRNA expression of apelin was detected in the 12 samples (100%) of the PDR group and in four samples (33%) of the idiopathic ERM group. Although a statistically significant difference (p<0.001) was observed between these two groups, the PCR results were not consistent with the immunofluorescence staining results, which showed negative staining of apelin in the idiopathic ERM group derived from the control subjects and no staining in the negative controls. Negative immunocytochemistry controls used PBS to replace the primary antibody. It could exclude the non-specific staining of the tissues by the second stage reagents, but it could not eliminate the possibility that the primary antibody was bound by non-specific means. Since the tissues from the PDR patient samples varied in their affinity for non-specific antibodies, a proper control would be to replace the primary antibody with a preimmune rabbit or mouse immunoglobulin. The non-specific staining in the first-stage antibodies could influence the immunofluorescence staining results.
It has been reported that apelin plays a role in central and peripheral cardiovascular regulation in conscious rats [56]. Apelin lowers blood pressure via a nitric oxide (NO)-dependent mechanism, and the effect of apelin on blood pressure was abolished in the presence of a NO synthase inhibitor [57]. Many researchers indicated that NO, increased by NO synthase (NOS), played an important role in angiogenesis, which mediated endothelial cell survival, proliferation, migration, and interaction with the extracellular matrix [58,59]. Endothelial nitric oxide synthase (eNOS) is a key enzyme that induces endothelial cells to produce NO, which is regulated by the phosphatidylinositol 3-kinase/protein kinase B (PI3K/Akt) signal pathway, which stimulates angiogenesis [60,61]. Recently, our research team found that apelin promoted proliferation, migration, and collagen I expression through the PI3K/Akt signaling pathways in RPE cells [62]. Therefore, NO may be downstream of apelin, and regulated via the PI3K/Akt signaling pathways.
In summary, we found high mRNA expression of apelin in ERMs after PDR. Moreover, immunofluorescence revealed the presence of apelin in the vascular and glial component of ERMs. In addition, intravitreal bevacizumab injections significantly reduced the expression of apelin and regressed vessels and fibroglial tissue in ERMs after PDR. Our results showed that apelin was involved in the formation of adventitia and promoted cell proliferation and angiogenesis of ERMs after PDR, and bevacizumab might be useful in preventing the development of ERMs after PDR.
Acknowledgments
We appreciate the technical help and advice from Chu LQ at Beijing Shijitan Hospital. This study was supported by National Natural Science Foundation of China (81271027 and 81260152) and an EFSD/CDS/Lilly grant (2127000043).
References
- 1.Roth AM, Foos RY. Surface wrinkling retinopathy in eyes enucleated at autopsy. Trans Am Acad Ophthalmol Otolaryngol. 1971;75:1047–58. [PubMed] [Google Scholar]
- 2.Margherio RR, Margherio AR. Epiretinal macular membranes. In Principles and Practice of Ophthalmology. 2nd ed. Albert DM, Jakobiec FA, Azar DT, Gragoudas ES. Philadelphia, W. B. Saunders 2000; 2103. [Google Scholar]
- 3.Klein R, Klein BE, Moss SE, Davis MD, DeMets DL. The Wisconsin epidemiologic study of diabetic retinopathy. II. Prevalence and risk of diabetic retinopathy when age at diagnosis is less than 30 years. Arch Ophthalmol. 1984;102:520–6. doi: 10.1001/archopht.1984.01040030398010. [DOI] [PubMed] [Google Scholar]
- 4.Klein R, Klein BE, Moss SE, Davis MD, DeMets DL. The Wisconsin epidemiologic study of diabetic retinopathy. III. Prevalence and risk of diabetic retinopathy when age at diagnosis is 30 or more years. Arch Ophthalmol. 1984;102:527–32. doi: 10.1001/archopht.1984.01040030405011. [DOI] [PubMed] [Google Scholar]
- 5.Simó R, Carrasco E, García-Ramírez M, Hernández C. Angiogenic and antiangiogenic factors in proliferative diabetic retinopathy. Curr Diabetes Rev. 2006;2:71–98. doi: 10.2174/157339906775473671. [DOI] [PubMed] [Google Scholar]
- 6.Smiddy WE, Maguire AM, Green WR, Michels RG, de la Cruz Z, Enger C, Jaeger M, Rice TA. Idiopathic epiretinal membranes: ultrastructural characteristics and clinicopathologic correlation. Ophthalmology. 1989;96:811–20. doi: 10.1016/s0161-6420(89)32811-9. [DOI] [PubMed] [Google Scholar]
- 7.Wiedemann P. Growth factors in retinal diseases: proliferative vitreoretinopathy, proliferative diabetic retinopathy, and retinal degeneration. Surv Ophthalmol. 1992;36:373–84. doi: 10.1016/0039-6257(92)90115-a. [DOI] [PubMed] [Google Scholar]
- 8.Elner SG, Elner VM, Jaffe GJ, Stuart A, Kunkel SL, Strieter RM. Cytokines in proliferative diabetic retinopathy and proliferative vitreoretinopathy. Curr Eye Res. 1995;14:1045–53. doi: 10.3109/02713689508998529. [DOI] [PubMed] [Google Scholar]
- 9.El-Ghrably IA, Dua HS, Orr GM, Fischer D, Tighe PJ. Detection of cytokine mRNA production in infiltrating cells in proliferative vitreoretinopathy using reverse transcription polymerase chain reaction. Br J Ophthalmol. 1999;83:1296–9. doi: 10.1136/bjo.83.11.1296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cox CM, D’Agostino SL, Miller MK, Heimark RL, Krieg PA. Apelin, the ligand for the endothelial G-protein-coupled receptor, APJ, is a potent angiogenic factor required for normal vascular development of the frog embryo. Dev Biol. 2006;296:177–89. doi: 10.1016/j.ydbio.2006.04.452. [DOI] [PubMed] [Google Scholar]
- 11.Kälin RE, Kretz MP, Meyer AM, Kispert A, Heppner FL, Brändli AW. Paracrine and autocrine mechanisms of apelin signaling govern embryonic and tumor angiogenesis. Dev Biol. 2007;305:599–614. doi: 10.1016/j.ydbio.2007.03.004. [DOI] [PubMed] [Google Scholar]
- 12.Devic E, Rizzoti K, Bodin S, Knibiehler B, Audigier Y. Amino acid sequence and embryonic expression of msr/apj, the mouse homolog of Xenopus X-msr and human APJ. Mech Dev. 1999;84:199–203. doi: 10.1016/s0925-4773(99)00081-7. [DOI] [PubMed] [Google Scholar]
- 13.Saint-Geniez M, Masri B, Malecaze F, Knibiehler B, Audigier Y. Expression of the murine msr/apj receptor and its ligand apelin is upregulated during formation of the retinal vessels. Mech Dev. 2002;110:183–6. doi: 10.1016/s0925-4773(01)00558-5. [DOI] [PubMed] [Google Scholar]
- 14.Saint-Geniez M, Argence CB, Knibiehler B, Audigier Y. The msr/apj gene encoding the apelin receptor is an early and specific marker of the venous phenotype in the retinal vasculature. Gene Expr Patterns. 2003;3:467–72. doi: 10.1016/s1567-133x(03)00062-0. [DOI] [PubMed] [Google Scholar]
- 15.Kasai A, Shintani N, Oda M, Kakuda M, Hashimoto H, Matsuda T, Hinuma S, Baba A. Apelin is a novel angiogenic factor in retinal endothelial cells. Biochem Biophys Res Commun. 2004;325:395–400. doi: 10.1016/j.bbrc.2004.10.042. [DOI] [PubMed] [Google Scholar]
- 16.Kasai A, Shintani N, Kato H, Matsuda S, Gomi F, Haba R, Hashimoto H, Kakuda M, Tano Y, Baba A. Retardation of retinal vascular development in apelin-deficient mice. Arterioscler Thromb Vasc Biol. 2008;28:1717–22. doi: 10.1161/ATVBAHA.108.163402. [DOI] [PubMed] [Google Scholar]
- 17.Masri B, Morin N, Cornu M, Knibiehler B, Audigier Y. Apelin (65–77) activates p70 S6 kinase and is mitogenic for umbilical endothelial cells. FASEB J. 2004;18:1909–11. doi: 10.1096/fj.04-1930fje. [DOI] [PubMed] [Google Scholar]
- 18.Kleinz MJ, Davenport AP. Immunocytochemical localization of the endogenous vasoactive peptide apelin to human vascular and endocardial endothelial cells. Regul Pept. 2004;118:119–25. doi: 10.1016/j.regpep.2003.11.002. [DOI] [PubMed] [Google Scholar]
- 19.Sorli SC, Le Gonidec S, Knibiehler B, Audigier Y. Apelin is a potent activator of tumour neoangiogenesis. Oncogene. 2007;26:7692–9. doi: 10.1038/sj.onc.1210573. [DOI] [PubMed] [Google Scholar]
- 20.Kidoya H, Ueno M, Yamada Y, Mochizuki N, Nakata M, Yano T, Fujii R, Takakura N. Spatial and temporal role of the apelin/APJ system in the caliber size regulation of blood vessels during angiogenesis. EMBO J. 2008;27:522–34. doi: 10.1038/sj.emboj.7601982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Masri B, van den Berghe L, Sorli C, Knibiehler B, Audigier Y. Apelin signalisation and vascular physiopathology. J Soc Biol. 2009;203:171–9. doi: 10.1051/jbio/2009021. [DOI] [PubMed] [Google Scholar]
- 22.Masri B, Knibiehler B, Audigier Y. Apelin signalling: a promising pathway from cloning to pharmacology. Cell Signal. 2005;17:415–26. doi: 10.1016/j.cellsig.2004.09.018. [DOI] [PubMed] [Google Scholar]
- 23.Principe A, Melgar-Lesmes P, Fernández-Varo G, del Arbol LR, Ros J, Morales-Ruiz M, Bernardi M, Arroyo V, Jiménez W. The hepatic apelin system: a new therapeutic target for liver disease. Hepatology. 2008;48:1193–201. doi: 10.1002/hep.22467. [DOI] [PubMed] [Google Scholar]
- 24.Sorli SC, van den Berghe L, Masri B, Knibiehler B, Audigier Y. Therapeutic potential of interfering with apelin signalling. Drug Discov Today. 2006;11:1100–6. doi: 10.1016/j.drudis.2006.10.011. [DOI] [PubMed] [Google Scholar]
- 25.Avery RL, Pearlman J, Pieramici DJ, Rabena MD, Castellarin AA, Nasir MA, Giust MJ, Wendel R, Patel A. Intravitreal bevacizumab (Avastin) in the treatment of proliferative diabetic retinopathy. Ophthalmology. 2006;113:e1. doi: 10.1016/j.ophtha.2006.05.064. [DOI] [PubMed] [Google Scholar]
- 26.Spaide RF, Fisher YL. Intravitreal bevacizumab (Avastin) treatment of proliferative diabetic retinopathy complicated by vitreous hemorrhage. Retina. 2006;26:275–8. doi: 10.1097/00006982-200603000-00004. [DOI] [PubMed] [Google Scholar]
- 27.Jorge R, Costa RA, Calucci D, Cintra LP. Scott IUl. Intravitreal bevacizumab (Avastin) for persistent new vessels in diabetic retinopathy (IBEPE study). Retina. 2006;26:1006–13. doi: 10.1097/01.iae.0000246884.76018.63. [DOI] [PubMed] [Google Scholar]
- 28.Moradian S, Ahmadieh H, Malihi M, Soheilian M, Dehghan MH, Azarmina M. Intravitreal bevacizumab in active progressive proliferative diabetic retinopathy. Graefes Arch Clin Exp Ophthalmol. 2008;246:1699–705. doi: 10.1007/s00417-008-0914-4. [DOI] [PubMed] [Google Scholar]
- 29.Rizzo S, Genovesi-Ebert F, Di Bartolo E, Vento A, Miniaci S, Williams G. Injection of intravitreal bevacizumab (Avastin) as a preoperative adjunct before vitrectomy surgery in the treatment of severe proliferative diabetic retinopathy (PDR). Graefes Arch Clin Exp Ophthalmol. 2008;246:837–42. doi: 10.1007/s00417-008-0774-y. [DOI] [PubMed] [Google Scholar]
- 30.Qian J, Lu Q, Tao Y, Jiang YR. Vitrous and plasma concentration of apelin and vascular endothelial growth factor after intravitreal Bevaciaumab in eyes with proliferative diabetic retinopathy. Retina. 2011;31:161–8. doi: 10.1097/IAE.0b013e3181e46ad8. [DOI] [PubMed] [Google Scholar]
- 31.Ma Y, Zhang Y, Zhao T, Jiang YR. Vascular endothelial growth factor in plasma and vitreous fluid of patients with proliferative diabetic retinopathy patients after intravitreal injection of bevacizumab. Am J Ophthalmol. 2012;153:307–13. doi: 10.1016/j.ajo.2011.08.006. [DOI] [PubMed] [Google Scholar]
- 32.Harada T, Harada C, Sekiguchi M, Wada K. Light-induced retinal degeneration suppresses developmental progression of flip-to-flop alternative splicing in GluR1. J Neurosci. 1998;18:3336–43. doi: 10.1523/JNEUROSCI.18-09-03336.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mizutani M, Gerhardinger C, Lorenzi M. Müller cell changes in human diabetic retinopathy. Diabetes. 1998;47:445–9. doi: 10.2337/diabetes.47.3.445. [DOI] [PubMed] [Google Scholar]
- 34.Barber AJ, Lieth E, Khin SA, Antonetti DA, Buchanan AG, Gardner TW, Penn State Retina Research Group Neural apoptosis in the retina during experimental and human diabetes. J Clin Invest. 1998;102:783–91. doi: 10.1172/JCI2425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gupta N, Brown KE, Milam AH. Activated microglia in human retinitis pigmentosa, late-onset retinal degeneration, and age-related macular degeneration. Exp Eye Res. 2003;76:463–71. doi: 10.1016/s0014-4835(02)00332-9. [DOI] [PubMed] [Google Scholar]
- 36.McGillem GS, Dacheux RF. Migration of retinal microglia in experimental proliferative vitreoretinopathy. Exp Eye Res. 1998;67:371–5. doi: 10.1006/exer.1998.0526. [DOI] [PubMed] [Google Scholar]
- 37.Cantó Soler MV, Gallo JE, Dodds RA, Hokfelt T, Villar MJ, Suburo AM. Y1 receptor of neuropeptide Y as a glial marker in proliferative vitreoretinopathy and diseased human retina. Glia. 2002;39:320–4. doi: 10.1002/glia.10107. [DOI] [PubMed] [Google Scholar]
- 38.Barber AJ, Antonetti DA, Gardner TW. Altered expression of retinal occludin and glial fibrillary acidic protein in experimental diabetes. Invest Ophthalmol Vis Sci. 2000;41:3561–8. [PubMed] [Google Scholar]
- 39.Rungger-Brändle E, Dosso AA, Leuenberger PM. Glial reactivity, an early feature of diabetic retinopathy. Invest Ophthalmol Vis Sci. 2000;41:1971–80. [PubMed] [Google Scholar]
- 40.Nork TM, Wallow IHL, Sramek SJ, Anderson G. Müller’s cell involvement in proliferative diabetic retinopathy. Arch Ophthalmol. 1987;105:1424–9. doi: 10.1001/archopht.1987.01060100126042. [DOI] [PubMed] [Google Scholar]
- 41.Ohira A, de Juan E. Characterisation of glial involvement in proliferative diabetic retinopathy. Ophthalmologica. 1990;201:187–95. doi: 10.1159/000310150. [DOI] [PubMed] [Google Scholar]
- 42.Behzadian MA, Wang XL, Shabrawey M, Caldwell RB. Effects of hypoxia on glial cell expression of angiogenesis-regulating factors VEGF and TGF-beta. Glia. 1998;24:216–25. [PubMed] [Google Scholar]
- 43.Amin RH, Frank RN, Kennedy A, Eliott D, Puklin JE, Abrams GW. Vascular endothelial growth factor is present in glial cells of the retina and optic nerve of human subjects with nonproliferative diabetic retinopathy. Invest Ophthalmol Vis Sci. 1997;38:36–47. [PubMed] [Google Scholar]
- 44.Tout S, Chan-Ling T, Hollander H, Stone J. The role of Müller cells in the formation of the blood-retinal barrier. Neuroscience. 1993;55:291–301. doi: 10.1016/0306-4522(93)90473-s. [DOI] [PubMed] [Google Scholar]
- 45.Abu El-Asrar AM, Desmet S, Meersschaert A, Dralands L, Missotten L, Geboes K. Expression of the inducible isoform of nitric oxide synthase in the retinas of human subjects with diabetes mellitus. Am J Ophthalmol. 2001;132:551–6. doi: 10.1016/s0002-9394(01)01127-8. [DOI] [PubMed] [Google Scholar]
- 46.Scott NA, Cipolla GD, Ross CE, Dunn B, Martin FH, Simonet L, Wilcox JN. Identification of a Potential Role for the Adventitia in Vascular Lesion Formation After Balloon Overstretch Injury of Porcine Coronary Arteries. Circulation. 1996;93:2178–87. doi: 10.1161/01.cir.93.12.2178. [DOI] [PubMed] [Google Scholar]
- 47.Torsney E, Hu Y, Xu Q. Adventitial Progenitor Cells Contribute to Arteriosclerosis. Trends Cardiovasc Med. 2005;15:64–8. doi: 10.1016/j.tcm.2005.02.003. [DOI] [PubMed] [Google Scholar]
- 48.Edelman ER, Adams DH, Karnovsky MJ. Effect of controlled adventitial heparin delivery on smooth-muscle cell proliferation following endothelial injury. Proc Natl Acad Sci USA. 1990;87:3773–7. doi: 10.1073/pnas.87.10.3773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Barker SG, Tilling LC, Miller GC, Beesley JE, Fleetwood G, Stavri GT, Baskerville PA, Martin JF. The adventitia and atherogenesis: removal initiates intimal proliferation in the rabbit which regresses on generation of a 'neoadventitia'. Atherosclerosis. 1994;105:131–44. doi: 10.1016/0021-9150(94)90043-4. [DOI] [PubMed] [Google Scholar]
- 50.Plekhanova OS, Stepanova VV, Ratner EI, Bobik A, Tkachuk VA, Parfyonova YV. Urokinase plasminogen activator in injured adventitia increases the number of myofibroblasts and augments early proliferation. J Vasc Res. 2006;43:437–46. doi: 10.1159/000094906. [DOI] [PubMed] [Google Scholar]
- 51.Murata T, Isibashi T, Khalil M, Hata Y, Yoshikawa H, Inomata H. Vascular endothelial growth factor plays a role in hyperpermeability of diabetic retinal vessels. Ophthalmic Res. 1995;27:48–52. doi: 10.1159/000267567. [DOI] [PubMed] [Google Scholar]
- 52.Tiani C, Garcia-Pras E, Mejias M, de Gottardi A, Berzigotti A, Bosch J, Fernandez M. Apelin signaling modulates splanchnic angiogenesis and portosystemic collateral vessel formation in rats with portal hypertension. J Hepatol. 2009;50:296–305. doi: 10.1016/j.jhep.2008.09.019. [DOI] [PubMed] [Google Scholar]
- 53.Lu Q, Feng J, Jiang YR. The role of apelin in the retina of diabetic rats. PLoS ONE. 2013;8:e69703. doi: 10.1371/journal.pone.0069703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Eyries M, Siegfried G, Ciumas M, Montagne K, Agrapart M, Lebrin F, Soubrier F. Hypoxia-induced apelin expression regulates endothelial cell proliferation and regenerative angiogenesis. Circ Res. 2008;103:432–40. doi: 10.1161/CIRCRESAHA.108.179333. [DOI] [PubMed] [Google Scholar]
- 55.Zhao T, Lu Q, Tao Y, Liang XY, Wang K, Jiang YR. Effects of apelin and vascular endothelial growth factor on central retinal vein occlusion in monkey eyes intravitreally injected with bevacizumab: a preliminary study. Mol Vis. 2011;17:1044–55. [PMC free article] [PubMed] [Google Scholar]
- 56.Kagiyama S, Fukuhara M, Matsumura K, Lin Y, Fujii K, Iida M. Central and peripheral cardiovascular actions of apelin in conscious rats. Regul Pept. 2005;125:55–9. doi: 10.1016/j.regpep.2004.07.033. [DOI] [PubMed] [Google Scholar]
- 57.Tatemoto K, Takayama K, Zou MX, Kumaki I, Zhang W, Kumano K, Fujimiya M. The novel peptide apelin lowers blood pressure via a nitric oxide-dependent mechanism. Regul Pept. 2001;99:87–92. doi: 10.1016/s0167-0115(01)00236-1. [DOI] [PubMed] [Google Scholar]
- 58.Bulotta S, Perrotta C, Cerullo A, De Palma C, Clementi E, Borgese N. A cellular system to study the role of nitric oxide in cell death, survival, and migration. Neurotoxicology. 2005;26:841–5. doi: 10.1016/j.neuro.2005.01.011. [DOI] [PubMed] [Google Scholar]
- 59.Cooke JP. NO and angiogenesis. Atheroscler Suppl. 2003;4:53–60. doi: 10.1016/s1567-5688(03)00034-5. [DOI] [PubMed] [Google Scholar]
- 60.Chung BH, Kim JD, Kim CK, Kim JH, Won MH, Lee HS, Dong MS, Ha KS, Kwon YG, Kim YM. Icariin stimulates angiogenesis by activating the MEK/ERK-and PI3K/Akt/eNOS-dependent signal pathways in human endothelial cells. Biochem Biophys Res Commun. 2008;376:404–8. doi: 10.1016/j.bbrc.2008.09.001. [DOI] [PubMed] [Google Scholar]
- 61.Lee SJ, Namkoong S, Kim YM, Kim CK, Lee H, Ha KS, Chung HT, Kwon YG, Kim YM. Fractalkine stimulates angiogenesis by activating the Raf-1/MEK/ERK-and PI3K/Akt/ eNOS-dependent signal pathways. Am J Physiol Heart Circ Physiol. 2006;291:H2836–46. doi: 10.1152/ajpheart.00113.2006. [DOI] [PubMed] [Google Scholar]
- 62.Qin D, Zheng XX, Jiang YR. apelin-13 induces proliferation, migration, and collagen I mRNA expression in human RPE cells via PI3K-Akt and MEK-Erk signaling pathways. Mol Vis. 2013;19:2227–36. [PMC free article] [PubMed] [Google Scholar]