

Abstract
The objectives of the present study were to investigate the role of K–Cl cotransporter 3 (KCC3) in the regulation of cellular invasion and the clinicopathological significance of its expression in esophageal squamous cell carcinoma (ESCC). Immunohistochemical analysis performed on 70 primary tumor samples obtained from ESCC patients showed that KCC3 was primarily found in the cytoplasm of carcinoma cells. Although the expression of KCC3 in the main tumor (MT) was related to several clinicopathological features, such as the pT and pN categories, it had no prognostic impact. KCC3 expression scores were compared between the MT and cancer nest (CN), and the survival rate of patients with a CN > MT score was lower than that of patients with a CN ≤ MT score. In addition, the survival rate of patients in whom KCC3 was expressed in the invasive front of tumor was lower than that of the patients without it. Furthermore, multivariate analysis demonstrated that the expression of KCC3 in the invasive front was one of the most important independent prognostic factors. The depletion of KCC3 using siRNAs inhibited cell migration and invasion in human ESCC cell lines. These results suggest that the expression of KCC3 in ESCC may affect cellular invasion and be related to a worse prognosis in patients with ESCC.
1. Introduction
The K–Cl cotransporter (KCC) mediates the coupled movement of K+ and Cl− ions across the plasma membrane and is involved in the regulation of cell volume, transepithelial ion transport, and maintenance of intracellular Cl− concentrations ([Cl−]i) [1, 2]. Four isoforms of the KCC have been identified and are termed KCC1, KCC2, KCC3, and KCC4 [3]. The four KCC isoforms share a common protein structure with 12 transmembrane regions in a central hydrophobic domain, together with hydrophilic N- and C-termini that may be cytoplasmic [4]. Although the expression of KCC1 is reportedly ubiquitous [5], that of KCC2 is restricted to neurons in the central nervous system [6]. KCC3 is expressed in the muscle, brain, lung, heart, and kidney [7], and KCC4 transcripts are the most abundant in the heart and kidney [4].
Several recent studies described the important roles of KCC in cancer development, tumor invasion, and possibly metastasis [8–11]. KCC3 was found to be important for cell-cycle progression, migration, and invasion in cervical carcinoma, ovarian cancer, breast cancer, and glioma [8, 9, 12, 13]. In addition, the overexpression of KCC3 downregulated the formation of the E-cadherin/β-catenin complex, and the subsequent upregulation of KCl cotransport activity was shown to benefit cancer cells in the epithelial-mesenchymal transition (EMT) [8]. However, the roles of KCC3 in the invasion of esophageal squamous cell carcinoma (ESCC) cells remain uncertain. Furthermore, the clinicopathological meaning of the expression of KCC3 in human ESCCs has not yet been evaluated.
The objectives of the present study were to investigate the roles of KCC3 in the cell migration and invasion of ESCC. Furthermore, we analyzed the expression of KCC3 in human ESCC samples and determined its relationships with the clinicopathological features and prognosis of ESCC patients. Our results revealed the important role of KCC3 in the tumor progression of ESCC.
2. Materials and Methods
2.1. Patients and Primary Tissue Samples
ESCC tumor samples were obtained from 70 patients with histologically proven primary ESCC who underwent esophagectomy (potentially curative R0 resection) at Kyoto Prefectural University of Medicine (Kyoto, Japan) between 1998 and 2007 and were analyzed retrospectively. These samples were embedded in paraffin 24 h after being fixed in formalin. Patient eligibility criteria included not having developed synchronous tumors or multiple metachronous tumors and not having received preoperative chemotherapy or radiation therapy. We excluded patients with noncuratively resected tumors or nonconsecutive data. All patients gave their written informed consent for inclusion in this study. Relevant clinicopathological and survival data were obtained from the hospital database. Staging was principally based on the seventh TNM staging system [14].
2.2. Immunohistochemistry
Paraffin sections (3 μm thick) of tumor tissue were subjected to immunohistochemical staining for KCC3 using the avidin-biotin-peroxidase method. Briefly, paraffin sections were dewaxed in xylene and hydrated through a graded series of alcohols. Antigen retrieval was performed by heating the samples in Dako REAL Target Retrieval Solution (Glostrup, Denmark) for 40 min at 95°C. Endogenous peroxidase activity was quenched by incubating the sections for 30 min in 0.3% H2O2. Sections were incubated for one hour at room temperature with the following antibody: the KCC3 antibody (HPA034563; Atlas Antibodies AB, Stockholm, Sweden). The avidin-biotin-peroxidase complex system (Vectastain ABC Elite kit; Vector Laboratories, Burlingame, CA, USA) was used for color development with diaminobenzidine tetrahydrochloride. Sections were counterstained with hematoxylin. These sections were then dehydrated through a graded series of alcohols, cleared in xylene, and mounted. Control sections of known positive ESCC were included in each antibody run, and negative control sections were produced by omitting the primary antibody.
Immunohistochemical samples stained with KCC3 were graded semiquantitatively by considering both the staining intensity and percentage of positive tumor cells using an immunoreactive score (IRS) [15]. Staining intensity was scored as 0 (no staining), 1 (weak staining), 2 (moderate staining), or 3 (strong staining). The proportion of positive tumor cells was scored from 0 to 1.0. The score of each sample was calculated as the maximum multiplied product of the intensity and proportion scores (0 to 3.0).
2.3. Cell Culture
The human ESCC cell lines TE5 and TE9 were obtained from the Cell Resource Center for Biomedical Research at the Institute of Development, Aging, and Cancer (Tohoku University, Sendai, Japan) [16]. These cells were grown in RPMI-1640 medium (Nacalai Tesque, Kyoto, Japan) supplemented with 100 U/mL of penicillin, 100 μg/mL of streptomycin, and 10% fetal bovine serum (FBS). Cells were cultured in flasks or dishes in a humidified incubator at 37°C under 5% CO2 in air.
2.4. Small Interfering RNA (siRNA) Transfection
Cells were transfected with 10 nmol/L KCC3 siRNA (Stealth RNAi siRNA #HSS HSS115159, HSS190762, HSS190763; Invitrogen, Carlsbad, CA) using the Lipofectamine RNAiMAX reagent (Invitrogen), according to the manufacturer's instructions. The medium containing siRNA was replaced with fresh medium after 24 h. We used three independent KCC3 siRNAs to exclude off target effects. The control siRNA provided (Stealth RNAi siRNA Negative Control; Invitrogen) was used as a negative control.
2.5. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Total RNA was extracted using an RNeasy kit (Qiagen, Valencia, CA). Messenger RNA (mRNA) expression was measured by quantitative real-time PCR (7300 Real-Time PCR System; Applied Biosystems, Foster City, CA) with TaqMan Gene Expression Assays (Applied Biosystems), according to the manufacturer's instructions. Expression levels were measured for the following gene: KCC3 (Hs00994548_m1) (Applied Biosystems). Expression was normalized for the KCC3 gene to the housekeeping gene beta-actin (ACTB, Hs01060665_g1; Applied Biosystems). Assays were performed in duplicate.
2.6. Analysis of Cell Migration and Invasion
The migration assay was conducted using a Cell Culture Insert with a pore size of 8 μm (BD Biosciences, Bedford, MA). Biocoat Matrigel (BD Biosciences) was used to evaluate cell invasion potential. Briefly, cells (1.5 × 105 cells per well) were seeded in the upper chamber in serum-free medium 24 h after siRNA transfection. The lower chamber contained medium with 10% FBS. The chambers were incubated for 48 h at 37°C in 5% CO2, and nonmigrated or noninvaded cells were then removed from the upper side of the membrane by scrubbing with cotton swabs. Migrated or invaded cells were fixed on the membrane and stained with Diff-Quick staining reagents (Sysmex, Kobe, Japan). The migrated or invaded cells on the lower side of the membrane were counted in four independent fields of view at 100x magnification of each insert. Each assay was performed in triplicate.
2.7. Statistical Analysis
Statistical analysis was carried out using Fisher's exact test to investigate correlations between clinicopathological parameters and KCC3 expression. Survival curves were constructed using the Kaplan-Meier method, and differences in survival were examined using the log-rank test. Multivariate analysis of the factors influencing survival was performed using the Cox proportional hazard model. Multiple comparisons were carried out using Dunnett's test after one-way ANOVA. Differences were considered significant when the associated P value was less than 0.05. All analyses were performed using statistical software (JMP, version 10; SAS Institute Inc., Cary, NC, USA). Correlation analyses were performed by creating Fit Y by X plots using JMP.
3. Results
3.1. KCC3 Protein Expression in Human ESCCs
An immunohistochemical investigation with the KCC3 antibody revealed the expression of KCC3 in the parabasal cell layer of normal esophageal mucosa (Figure 1(a)). We examined the expression of KCC3 in 70 primary tumor samples of human ESCC based on their immunohistochemical reactivity. The KCC3 protein was mostly expressed in the cytoplasm of carcinoma cells (Figure 1(b)). The KCC3 score in the main tumor (MT) varied widely between the tumors. The minimum KCC3 score was 0 while the maximum KCC3 score was 2.4 in MT (median = 0.725; mean ± standard error of the mean (SEM) = 0.780 ± 0.072). Regarding the expression of KCC3 in MT, we divided ESCC patients into 2 groups using the median staining score: a low grade KCC3 expression group with staining scores ≤0.725, n = 35, and a high grade KCC3 expression group with staining scores >0.725, n = 35. Figures 1(c) and 1(d) show the representative histopathological findings of low or high KCC3 expression samples. Correlations between the expression of KCC3 in MT and various clinicopathological parameters were analyzed (Table 1). We found correlations between the expression of KCC3 in MT and location of the primary tumor, the pT or pN category (Table 1).
Figure 1.
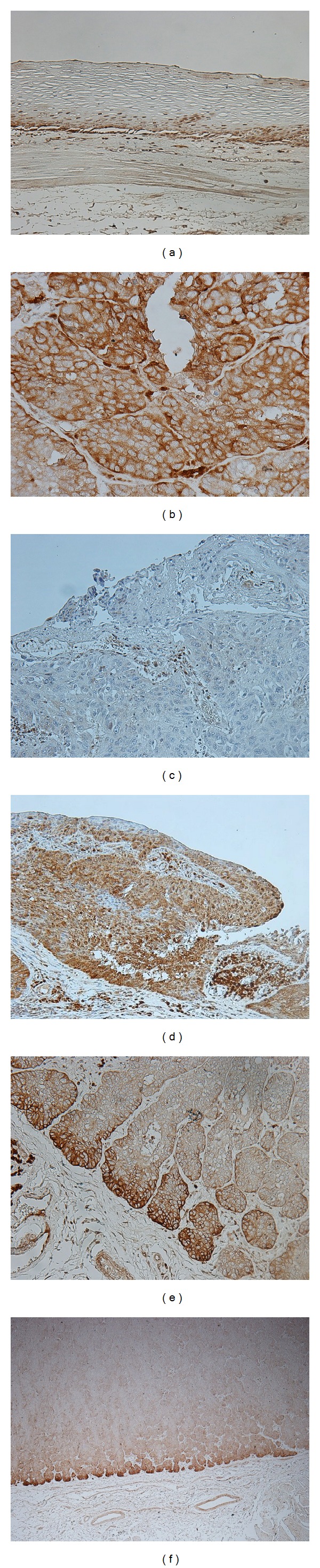
KCC3 protein expression in human esophageal squamous cell carcinoma (ESCC). (a) Immunohistochemical staining of noncancerous esophageal epithelia with the KCC3 antibody. Magnification: ×200. (b) Immunohistochemical staining of primary human ESCC samples with the KCC3 antibody. Magnification: ×400. (c) Immunohistochemical staining of primary human ESCC samples with the low grade expression of KCC3 in the main tumor (MT). Magnification: ×200. (d) Immunohistochemical staining of primary human ESCC samples with the high grade expression of KCC3 in the main tumor (MT). Magnification: ×200. (e) Immunohistochemical staining of primary human ESCC samples with the high grade expression of KCC3 in the cancer nest (CN). Magnification: ×200. (f) Immunohistochemical staining of primary human ESCC samples that expressed KCC3 in the invasive front of the tumor. Magnification: ×40.
Table 1.
Relationships between the clinicopathological features of esophageal cancer and expression of KCC3 in the main tumor or cancer nest.
| Variable | MT | P value | CN | P value | CN/MT | P value | |||
|---|---|---|---|---|---|---|---|---|---|
| Low | High | Low | High | CN ≤ MT | CN > MT | ||||
| (n = 35) | (n = 35) | (n = 36) | (n = 34) | (n = 31) | (n = 39) | ||||
| Age | |||||||||
| <60 years | 11 | 11 | 1.000 | 11 | 11 | 1.000 | 6 | 16 | 0.071 |
| ≥60 years | 24 | 24 | 25 | 23 | 25 | 23 | |||
| Gender | |||||||||
| Male | 30 | 29 | 1.000 | 30 | 29 | 1.000 | 25 | 34 | 0.520 |
| Female | 5 | 6 | 6 | 5 | 6 | 5 | |||
| Location of the primary tumor | |||||||||
| Ut-Mt | 18 | 29 | 0.0101∗ | 19 | 28 | 0.0112∗ | 21 | 26 | 1.000 |
| Lt-Ae | 17 | 6 | 17 | 6 | 10 | 13 | |||
| Histological type | |||||||||
| Well/moderately differentiated SCC | 22 | 27 | 0.297 | 26 | 23 | 0.796 | 22 | 27 | 1.000 |
| Poorly differentiated SCC | 13 | 8 | 10 | 11 | 9 | 12 | |||
| Tumor size | |||||||||
| <50 mm | 22 | 27 | 0.2968 | 24 | 25 | 0.6069 | 24 | 25 | 0.2967 |
| ≥50 mm | 13 | 8 | 12 | 9 | 7 | 14 | |||
| Lymphatic invasion | |||||||||
| Negative | 15 | 18 | 0.6324 | 20 | 13 | 0.1606 | 18 | 15 | 0.1482 |
| Positive | 20 | 17 | 16 | 21 | 13 | 24 | |||
| Venous invasion | |||||||||
| Negative | 19 | 21 | 0.8094 | 20 | 20 | 0.8133 | 18 | 22 | 1.000 |
| Positive | 16 | 14 | 16 | 14 | 13 | 17 | |||
| pT | |||||||||
| pT1 | 10 | 23 | 0.0037∗ | 14 | 19 | 0.2309 | 18 | 15 | 0.1482 |
| pT2-3 | 25 | 12 | 22 | 15 | 13 | 24 | |||
| pN | |||||||||
| pN0 | 11 | 22 | 0.0160∗ | 15 | 18 | 0.473 | 16 | 17 | 0.631 |
| pN1-3 | 24 | 13 | 21 | 16 | 15 | 22 | |||
MT: main tumor; CN: cancer nest; Ut: upper thoracic esophagus; Mt: middle thoracic esophagus; Lt: lower thoracic esophagus; Ae: abdominal esophagus; SCC: squamous cell carcinoma; pT: pathological T stage; pN: pathological N stage.
∗ P < 0.05: Fisher's exact test.
We then focused on the expression of KCC3 in the cancer nest (CN) of ESCC (Figure 1(e)) and analyzed the KCC3 score in CN. The minimum KCC3 score was 0, and the maximum KCC3 score was 2.6 in CN (median = 1.000; mean ± SEM = 1.087 ± 0.096). The KCC3 score in CN was positively correlated with the KCC3 score in MT (R 2 = 0.3388, P < 0.0001) (Figure 2). Regarding the expression of KCC3 in CN, we divided ESCC patients into 2 groups using the median staining score, a low grade KCC3 expression group with staining scores ≤1.000, n = 36, and a high grade KCC3 expression group with staining scores >1.000, n = 34, and compared their clinicopathological features (Table 1). A correlation was found between the expression of KCC3 in CN and location of the primary tumor (Table 2). Regarding the comparison of KCC3 scores between MT and CN in each sample, we divided ESCC patients into 2 groups, CN > MT, n = 39, and CN ≤ MT, n = 31, and compared their clinicopathological features (Table 1). A correlation was not found between the comparison of KCC3 scores and clinicopathological features (Table 1).
Figure 2.
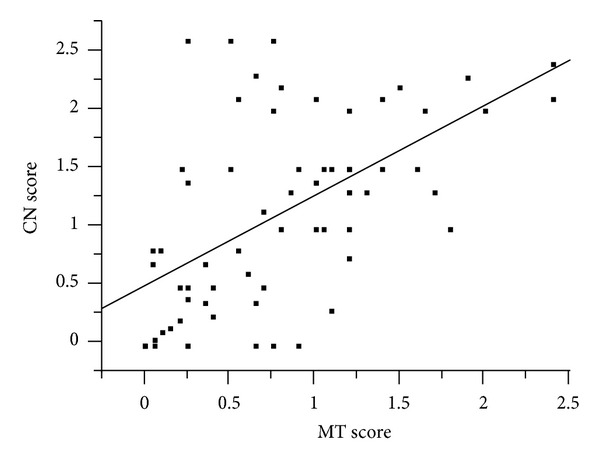
A correlation analysis of the relationship between the KCC3 score in main tumor (MT) and KCC3 score in the cancer nest (CN) was performed by producing Fit Y by X plots. The KCC3 score in CN was positively correlated with the KCC3 score in MT (R 2 = 0.3388, P < 0.0001).
Table 2.
Relationships between the clinicopathological features of esophageal cancer and expression of KCC3 in the invasive front of the tumor.
| Variable | Invasive front | P value | |
|---|---|---|---|
| Negative | Positive | ||
| (n = 22) | (n = 48) | ||
| Age | |||
| <60 years | 6 | 16 | 0.783 |
| ≥60 years | 16 | 32 | |
| Gender | |||
| Male | 21 | 38 | 0.154 |
| Female | 1 | 10 | |
| Location of the primary tumor | |||
| Ut-Mt | 12 | 35 | 0.172 |
| Lt-Ae | 10 | 13 | |
| Histological type | |||
| Well/moderately differentiated SCC | 15 | 34 | 1.000 |
| Poorly differentiated SCC | 7 | 14 | |
| Tumor size | |||
| <50 mm | 14 | 35 | 0.575 |
| ≥50 mm | 8 | 13 | |
| Lymphatic invasion | |||
| Negative | 10 | 23 | 1.000 |
| Positive | 12 | 25 | |
| Venous invasion | |||
| Negative | 13 | 27 | 1.000 |
| Positive | 9 | 21 | |
| pT | |||
| pT1 | 8 | 25 | 0.3035 |
| pT2-3 | 14 | 23 | |
| pN | |||
| pN0 | 8 | 25 | 0.3035 |
| pN1-3 | 14 | 23 | |
| MT | |||
| Low | 20 | 15 | <0.0001∗ |
| High | 2 | 33 | |
| CN | |||
| Low | 19 | 17 | <0.0001∗ |
| High | 3 | 31 | |
| CN/MT | |||
| CN ≤ MT | 14 | 17 | 0.0385∗ |
| CN > MT | 8 | 31 | |
Ut: upper thoracic esophagus; Mt: middle thoracic esophagus; Lt: lower thoracic esophagus; Ae: abdominal esophagus; SCC: squamous cell carcinoma; pT: pathological T stage; pN: pathological N stage; MT: main tumor; CN: cancer nest.
∗ P < 0.05: Fisher's exact test.
Furthermore, we analyzed the localization of KCC3 expression in tumors. In 48 cases, the expression of KCC3 was found in the invasive front of the tumor (Figure 1(f)). Regarding the expression of KCC3 in the invasive front of the tumor, we divided ESCC patients into 2 groups, negative (n = 22) and positive (n = 48), and compared their clinicopathological features (Table 2). A correlation was found between the expression of KCC3 in the invasive front and the MT score, CN score, or their comparison (CN/MT) (Table 2). No correlation was found between the expression of KCC3 in the invasive front and any other clinicopathological parameter (Table 2).
3.2. Prognostic Impact of KCC3 Protein Expression for Patients with ESCC
We determined the prognostic impact of the expression of KCC3 for patients with ESCC. Regarding the expression of KCC3 in MT, no significant difference was observed in the 5-year survival rate between patients with the high grade expression of KCC3 and those with the low grade expression of KCC3 in MT (Figure 3(a)). Similarly, no significant differences were observed in the 5-year survival rate between patients with the high grade expression of KCC3 and those with the low grade expression of KCC3 in CN (Figure 3(b)). Regarding comparisons of KCC3 score, the 5-year survival rate of the patients with CN > MT (55.5%) was lower than that of patients with CN ≤ MT (76.7%) (P = 0.133) (Figure 3(c)). The 5-year survival rate of the patients with KCC3 expression in the invasive front (57.1%) was lower than that of the patients without it (81.8%), although there was no statistical difference (P = 0.089) (Figure 3(d)). Interestingly, when patients were divided into 2 groups, CN > MT and invasive front positive, n = 31, and others, n = 39, the 5-year survival rate of patients with CN > MT and invasive front positive (46.1%) was significantly lower than that of other patients (79.0%) (P = 0.022) (Figure 3(e)).
Figure 3.
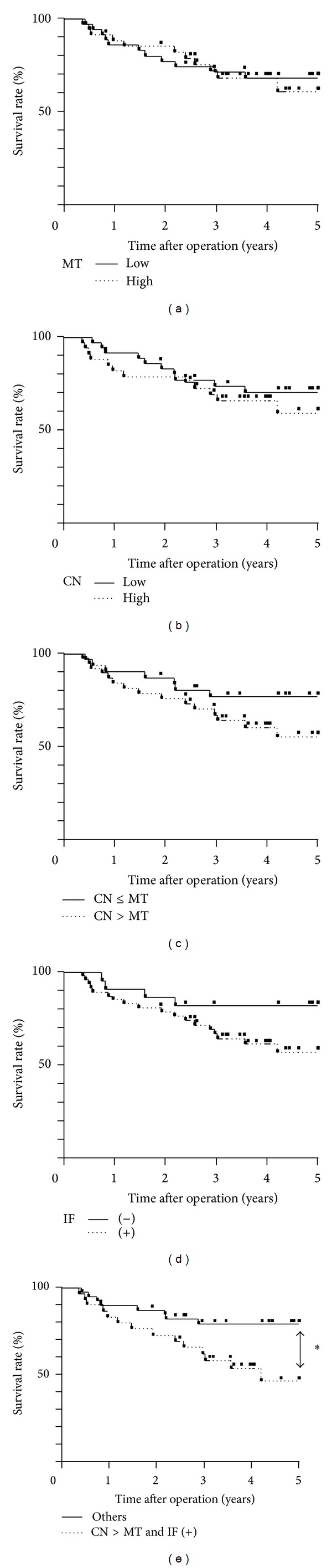
Survival curve of patients after curative resection for esophageal squamous cell carcinoma (ESCC) according to the expression of KCC3. (a) Patients were classified into two groups: low grade expression of KCC3 (n = 35) and high grade expression of KCC3 (n = 35) in the main tumor (MT). (b) Patients were classified into two groups: the low grade expression of KCC3 (n = 36) and high grade expression of KCC3 (n = 34) in the cancer nest (CN). (c) Patients were classified into two groups based on comparisons of the KCC3 score: CC ≤ MT (n = 31) and CN > MT (n = 39) in the cancer nest (CN). (d) Patients were classified into two groups based on the expression of KCC3 in the invasive front of the tumor: negative (n = 22) and positive (n = 48). (e) Patients were classified into two groups: patients with CN > MT and invasive front positive (n = 31) and others (n = 39). ∗ P < 0.05: log-rank test.
Furthermore, we assessed which of the 13 variables studied (age, gender, location of the primary tumor, histological type, tumor size, lymphatic invasion, venous invasion, pT and pN category, KCC3 score in MT, KCC3 score in CN and CN/MT, and KCC3 expression in the invasive front) influenced survival following curative resection of esophageal cancer. Univariate analysis of survival after esophagectomy revealed that lymphatic invasion, venous invasion, and the pT and pN categories were found to be significant prognostic factors (P = 0.017, 0.017, 0.002, and 0.003, resp.) (Table 3). Multivariate analysis with variables whose P values were less than 0.100 in univariate analysis demonstrated that lymphatic invasion, pT and pN category, and KCC3 expression in invasive front were independent prognostic factors (P = 0.044, 0.015, 0.011, and 0.001, resp.) (Table 4). KCC3 expression in invasive front was the strongest prognostic factor of all clinicopathological features. These findings suggest that the expression of KCC3 might be a valuable prognostic factor for patients with ESCC.
Table 3.
Five-year survival rate of patients with esophageal cancer according to various clinicopathological parameters.
| Variables | 5-year survival rate (%) |
P value |
|---|---|---|
| Age | ||
| <60 years | 53.13 | 0.1179 |
| ≥60 years | 72.2 | |
| Gender | ||
| Male | 65.35 | 0.8597 |
| Female | 68.57 | |
| Location of the primary tumor | ||
| Ut-Mt | 73.63 | 0.0564 |
| Lt-Ae | 50.82 | |
| Histological type | ||
| Well/moderately differentiated SCC | 71.94 | 0.2301 |
| Poorly differentiated SCC | 53.43 | |
| Tumor size | ||
| <50 mm | 68.68 | 0.2809 |
| ≥50 mm | 59.52 | |
| Lymphatic invasion | ||
| Negative | 78.4 | 0.0168∗ |
| Positive | 54.78 | |
| Venous invasion | ||
| Negative | 78.18 | 0.0169∗ |
| Positive | 48.7 | |
| pT | ||
| pT1 | 82.22 | 0.0024∗ |
| pT2-3 | 50.68 | |
| pN | ||
| pN0 | 82.72 | 0.0029∗ |
| pN1-3 | 49.68 | |
| MT | ||
| Low | 68.21 | 0.7838 |
| High | 60.56 | |
| CN | ||
| Low | 70.48 | 0.4151 |
| High | 59.25 | |
| CN/MT | ||
| CN ≤ MT | 76.74 | 0.1329 |
| CN > MT | 55.48 | |
| Invasive front | ||
| Negative | 81.82 | 0.0887 |
| Positive | 57.07 |
Ut: upper thoracic esophagus; Mt: middle thoracic esophagus; Lt: lower thoracic esophagus; Ae: abdominal esophagus; SCC: squamous cell carcinoma; pT: pathological T stage; pN: pathological N stage; MT: main tumor; CN: cancer nest.
∗ P < 0.05: log-rank test.
Table 4.
Prognostic factors of esophageal cancer according to multivariate analysis.
| Variables | Risk ratio | 95% CI | P value |
|---|---|---|---|
| Location of the primary tumor | |||
| Ut-Mt | Ref. | 0.861452–2.20629 | 0.1813 |
| Lt-Ae | 1.37267 | ||
| Lymphatic invasion | |||
| Negative | Ref. | 1.012886–2.725895 | 0.0437∗ |
| Positive | 1.605417 | ||
| Venous invasion | |||
| Negative | Ref. | 0.954510–2.395284 | 0.08 |
| Positive | 1.483897 | ||
| pT | |||
| pT1 | Ref. | 1.122531–3.250204 | 0.0146∗ |
| pT2-3 | 1.834223 | ||
| pN | |||
| pN0 | Ref. | 1.152392–3.450263 | 0.0110∗ |
| pN1-3 | 1.911249 | ||
| Invasive front | |||
| Negative | Ref. | 1.357757–4.524146 | 0.0014∗ |
| Positive | 2.332559 |
Ut: upper thoracic esophagus; Mt: middle thoracic esophagus; Lt: lower thoracic esophagus; Ae: abdominal esophagus; pT: pathological T stage; pN: pathological N stage; Ref.: referent.
∗ P < 0.05: Cox's proportional hazards model; 95% CI: 95% confidence interval.
3.3. KCC3 Controlled Cell Migration and Invasion in ESCC Cells
We conducted knockdown experiments with KCC3 siRNA in ESCC cells and analyzed the effects of KCC3 knockdown on cell migration and invasion. We used three independent KCC3 siRNAs to exclude off target effects. All three KCC3 siRNAs effectively reduced KCC3 mRNA levels in both TE5 and TE9 cells (Figure 4(a)). In TE5 cells, all three KCC3 siRNAs significantly inhibited cell migration and invasion (Figure 4(b)). In TE9 cell, downregulation of KCC3 inhibited cell migration and invasion, too (Figure 4(c)). These results suggest that KCC3 plays an important role in regulating cell migration and invasion in ESCC cells.
Figure 4.
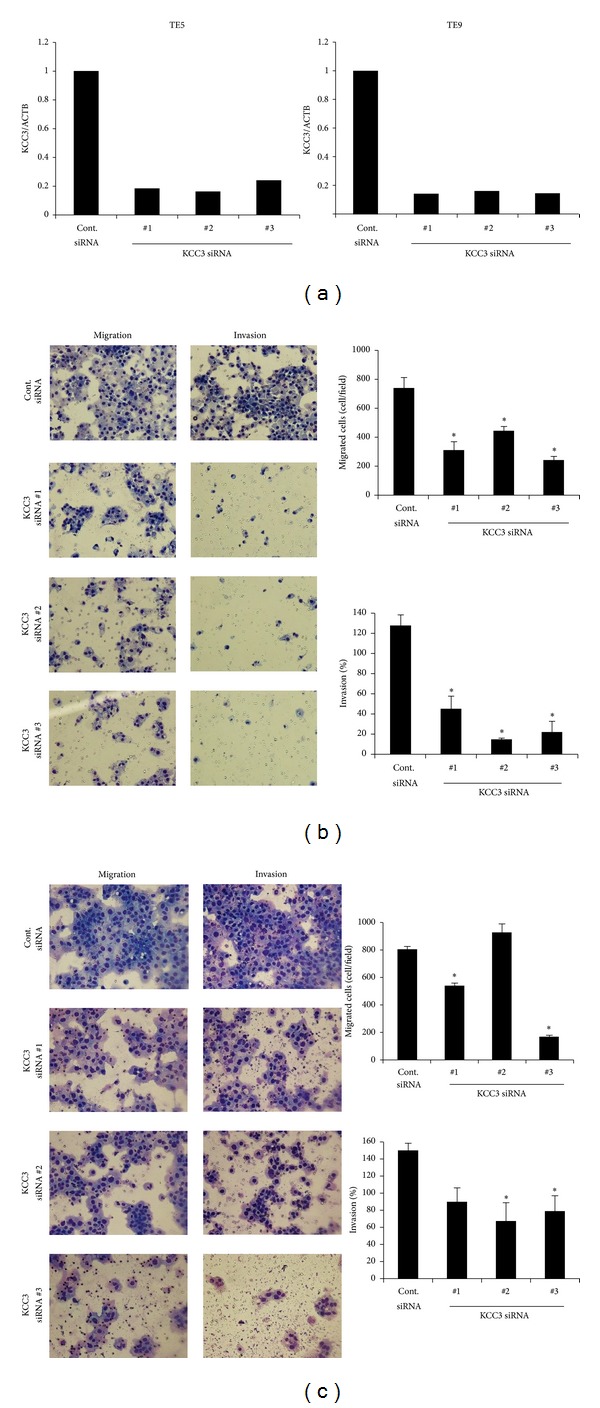
KCC3 controlled the cell migration and invasion of esophageal squamous cell carcinoma (ESCC) cells. (a) KCC3 siRNA effectively reduced KCC3 mRNA levels in both TE5 and TE9 cells. Three independent KCC3 siRNAs were investigated to exclude off target effects. (b) The downregulation of KCC3 significantly inhibited cell migration and invasion in TE5 cells. Cell migration and invasion were determined by the Boyden chamber assay. Mean ± SEM; n = 3. ∗ P < 0.05: Dunnett's test (ANOVA: migration; P < 0.0001, invasion; P < 0.0001). (c) The downregulation of KCC3 inhibited cell migration and invasion in TE9 cells. Cell migration and invasion were determined by the Boyden chamber assay. Mean ± SEM; n = 3. ∗ P < 0.05: Dunnett's test (ANOVA: migration; P < 0.0001, invasion; P = 0.0210).
4. Discussion
Recent studies have shown that ion channels and transporters play crucial roles in cellular functions, and their roles have been studied in cancer cells [17, 18]. Various types of ion channels, such as voltage-gated K+ channels, voltage-gated HERG channels, Ca2 + channels, and transient receptor potential channels, have been found to be expressed in gastrointestinal cancer cells and tissues and to regulate tumor behavior [19–22]. Regulators of intracellular pH such as anion exchanger, sodium-hydrogen exchanger, and carbonic anhydrases also related to tumor development of gastrointestinal cancer cells [23–25]. Furthermore, several reports have indicated that Cl− channels/transporters, such as Cl− channels, chloride intracellular channel (CLIC), KCC, and NKCC, play crucial roles in the tumorigenesis of colorectal, gastric, cervical, breast, lung, and prostate cancer cells [17, 18, 26]. We have also focused on and investigated transepithelial Cl− transport in various types of cancer cells [11, 27–29].
In the present study, we investigated the KCC3 expression in ESCC and determined its relationships with clinicopathological features and prognosis. To the best of our knowledge, this is the first report examining KCC3 expression in human ESCC tissue. Our results showed that KCC3 expression in MT related to several clinicopathological features, such as the pT and pN categories. However, the expression of KCC3 in MT itself did not have a prognostic impact. Although these results may not be persuasive because of the limitation of a small sample size, they showed that KCC3 was expressed in MT of ESCC from an early stage. Regarding the expression of KCC in cancer tissue, previous studies demonstrated that KCC3 was abundant in cervical carcinoma and CN invaded deeply into stromal tissues whereas KCC4 was abundant in metastatic cervical and ovarian cancer tissues [8, 10]. Furthermore, both the progression-free and overall survival rates of patients with the high grade expression of KCC4 were significantly poorer than those of patients with the low grade expression of KCC4 in cervical cancer [10], which suggested a relationship between the expression pattern of KCC and clinical outcome. Therefore, we focused on the distribution of KCC3 in tumors and analyzed its expression in CN or the invasive front of the tumor. Although the expression of KCC3 in CN itself had no prognostic impact, the 5-year survival rate of patients with a CN > MT score was slightly lower than that of patients with a CN ≤ MT score. Furthermore, the 5-year survival rate of patients in whom KCC3 was expressed in the invasive front was lower than that of patients without it, and multivariate analysis revealed that the expression of KCC3 in the invasive front was the strongest prognostic factor of all clinicopathological features. These results suggest the role of KCC3 in cancer invasion as well as the importance of its distribution in tumors as a prognostic predictor. We have previously identified several prognostic biomarkers in human ESCC, such as Ki-67, antiphosphohistone H3, p21, and E2F5 [30–33]. The expressions of these cell-cycle related proteins were mainly analyzed in MT. On the other hand, we focused on the distribution of KCC3 in the present study and showed its prognostic impact via cellular invasion.
Recent studies have indicated the importance of KCC in the cell migration and invasion of glioma, cervical, ovarian, and breast cancer cells [8–10, 12, 13]. Regarding the mechanism by which KCC regulates tumor invasion, KCC3 was previously shown to downregulate the formation of the E-cadherin/β-catenin complex in order to promote EMT, which is important for cervical cancer cell invasiveness [8]. In addition, a previous study reported that the motor protein-dependent membrane trafficking of KCC4 was important for cancer cell invasion [10]. Our in vitro study also demonstrated the important roles of KCC3 in cell migration and invasion in ESCC cells. One possible mechanism by which KCC regulates the malignant behavior of cancer cells may be through the regulation of [Cl−]i [11, 13]. Recent studies have shown that [Cl−]i is a critical signal mediator for the regulation of various cellular functions [34–36]. For instance, we showed that [Cl−]i could act as an important signal to control the gene expression of the epithelial Na+ channel via a tyrosine kinase in renal epithelial A6 cells [36]. We also previously reported that [Cl−]i controlled cell-cycle progression in gastric and prostate cancer cells [27–29, 37, 38]. Shen et al. showed that an alteration in the [Cl−]i concentration affected the activity of the retinoblastoma protein and cdc2 kinase, two key cell-cycle regulators that control progression from the G1 into the S phase and from the G2 into the M phase, respectively [13]. We considered KCC to be one of the important transporters that regulates [Cl−]i in the steady state and previously showed that the blockage of KCC decreased [Cl−]i in breast cancer cells [11]. Although this mechanism should be verified in more detail in further studies, these findings suggest that the changes induced in [Cl−]i by KCC3 may be a critically important messenger that regulates cellular invasiveness in ESCC cells.
In summary, we found that KCC3 played a role in the cell migration and invasion of ESCC cells. An immunohistochemical analysis revealed that the expression of KCC3 in the invasive front of tumors was the strongest prognostic factor in patients with ESCC. A deeper understanding of the role of KCC3 may lead to its use as a crucial biomarker of tumor progression and/or a new therapeutic target for ESCC.
Acknowledgments
This work was supported by Grants-in-Aid for Young Scientists (B) (22791295, 24791440) and a Grant-in-Aid for Scientific Research (C) (22591464, 24591957, and 26461988) from the Japan Society for the Promotion of Science.
Conflict of Interests
None of the authors have any conflict of interests or financial ties to disclose.
Authors' Contribution
Atsushi Shiozaki and Kenichi Takemoto contributed equally to this work.
References
- 1.Lauf PK, Adragna NC. K-Cl cotransport: properties and molecular mechanism. Cellular Physiology and Biochemistry. 2000;10(5-6):341–354. doi: 10.1159/000016357. [DOI] [PubMed] [Google Scholar]
- 2.Dunham PB, Stewart GW, Ellory JC. Chloride-activated passive potassium transport in human erythrocytes. Proceedings of the National Academy of Sciences of the United States of America. 1980;77(3):1711–1715. doi: 10.1073/pnas.77.3.1711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chen YF, Chou CY, Ellory JC, Shen MR. The emerging role of KCl cotransport in tumor biology. The American Journal of Translational Research. 2010;2(4):345–355. [PMC free article] [PubMed] [Google Scholar]
- 4.Mount DB, Mercado A, Song L, et al. Cloning and characterization of KCC3 and KCC4, new members of the cation-chloride cotransporter gene family. The Journal of Biological Chemistry. 1999;274(23):16355–16362. doi: 10.1074/jbc.274.23.16355. [DOI] [PubMed] [Google Scholar]
- 5.Gillen CM, Brill S, Payne JA, Forbush B., III Molecular cloning and functional expression of the K-Cl cotransporter from rabbit, rat, and human: A new member of the cation-chloride cotransporter family. Journal of Biological Chemistry. 1996;271(27):16237–16244. doi: 10.1074/jbc.271.27.16237. [DOI] [PubMed] [Google Scholar]
- 6.Payne JA, Stevenson TJ, Donaldson LF. Molecular characterization of a putative K-Cl cotransporter in rat brain: a neuronal-specific isoform. Journal of Biological Chemistry. 1996;271(27):16245–16252. doi: 10.1074/jbc.271.27.16245. [DOI] [PubMed] [Google Scholar]
- 7.Race JE, Makhlouf FN, Logue PJ, Wilson FH, Dunham PB, Holtzman EJ. Molecular cloning and functional characterization of KCC3, a new K-Cl cotransporter. The American Journal of Physiology. 1999;277(6, part 1):C1210–C1219. doi: 10.1152/ajpcell.1999.277.6.C1210. [DOI] [PubMed] [Google Scholar]
- 8.Hsu YM, Chen YF, Chou CY, et al. KCl cotransporter-3 down-regulates E-cadherin/β-catenin complex to promote epithelial-mesenchymal transition. Cancer Research. 2007;67(22):11064–11073. doi: 10.1158/0008-5472.CAN-07-2443. [DOI] [PubMed] [Google Scholar]
- 9.Hsu YM, Chou CY, Chen HH, et al. IGF-1 upregulates electroneutral K-Cl cotransporter KCC3 and KCC4 which are differentially required for breast cancer cell proliferation and invasiveness. Journal of Cellular Physiology. 2007;210(3):626–636. doi: 10.1002/jcp.20859. [DOI] [PubMed] [Google Scholar]
- 10.Chen YF, Chou CY, Wilkins RJ, Ellory JC, Mount DB, Shen M. Motor protein-dependent membrane trafficking of KCl cotransporter-4 is important for cancer cell invasion. Cancer Research. 2009;69(22):8585–8593. doi: 10.1158/0008-5472.CAN-09-2284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kitagawa M, Niisato N, Shiozaki A, et al. A regulatory role of K+-Cl− cotransporter in the cell cycle progression of breast cancer MDA-MB-231 cells. Archives of Biochemistry and Biophysics. 2013;539(1):92–98. doi: 10.1016/j.abb.2013.06.014. [DOI] [PubMed] [Google Scholar]
- 12.Gagnon KB. High-grade glioma motility reduced by genetic knockdown of KCC3. Cellular Physiology and Biochemistry. 2012;30(2):466–476. doi: 10.1159/000339040. [DOI] [PubMed] [Google Scholar]
- 13.Shen MR, Chou CY, Hsu KF, et al. The KCI cotransporter isoform KCC3 can play an important role in cell growth regulation. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(25):14714–14719. doi: 10.1073/pnas.251388798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sobin L, Gospodarowicz M, Wittekind C, editors. TNM Classification of Malignant Tumors. 7th edition. Hoboken, NJ, USA: John Wiley & Sons; 2009. [Google Scholar]
- 15.Remmele W, Stegner HE. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe. 1987;8(3):138–140. [PubMed] [Google Scholar]
- 16.Nishihira T, Hashimoto Y, Katayama M, Mori S, Kuroki T. Molecular and cellular features of esophageal cancer cells. Journal of Cancer Research and Clinical Oncology. 1993;119(8):441–449. doi: 10.1007/BF01215923. [DOI] [PubMed] [Google Scholar]
- 17.Kunzelmann K. Ion channels and cancer. Journal of Membrane Biology. 2005;205(3):159–173. doi: 10.1007/s00232-005-0781-4. [DOI] [PubMed] [Google Scholar]
- 18.Schönherr R. Clinical relevance of ion channels for diagnosis and therapy of cancer. The Journal of Membrane Biology. 2005;205(3):175–184. doi: 10.1007/s00232-005-0782-3. [DOI] [PubMed] [Google Scholar]
- 19.Kim CJ, Cho YG, Jeong SW, et al. Altered expression of KCNK9 in colorectal cancers. APMIS. 2004;112(9):588–594. doi: 10.1111/j.1600-0463.2004.apm1120905.x. [DOI] [PubMed] [Google Scholar]
- 20.Shao XD, Wu KC, Hao ZM, Hong L, Zhang J, Fan D. The potent inhibitory effects of cisapride, a specific blocker for human ether-a-go-go-related gene (HERG) channel, on gastric cancer cells. Cancer Biology and Therapy. 2005;4(3):295–301. doi: 10.4161/cbt.4.3.1500. [DOI] [PubMed] [Google Scholar]
- 21.Wang X, Nagaba Y, Cross HS, Wrba F, Zhang L, Guggino SE. The mRNA of L-type calcium channel elevated in colon cancer: protein distribution in normal and cancerous colon. The American Journal of Pathology. 2000;157(5):1549–1562. doi: 10.1016/S0002-9440(10)64792-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cai R, Ding X, Zhou K, et al. Blockade of TRPC6 channels induced G2/M phase arrest and suppressed growth in human gastric cancer cells. International Journal of Cancer. 2009;125(10):2281–2287. doi: 10.1002/ijc.24551. [DOI] [PubMed] [Google Scholar]
- 23.Wu J, Zhang Y-C, Suo W-H, et al. Induction of anion exchanger-1 translation and its opposite roles in the carcinogenesis of gastric cancer cells and differentiation of K562 cells. Oncogene. 2010;29(13):1987–1996. doi: 10.1038/onc.2009.481. [DOI] [PubMed] [Google Scholar]
- 24.Nagata H, Che X, Miyazawa K, et al. Rapid decrease of intracellular pH associated with inhibition of Na +/H+ exchanger precedes apoptotic events in the MNK45 and MNK74 gastric cancer cell lines treated with 2-aminophenoxazine-3-one. Oncology Reports. 2011;25(2):341–346. doi: 10.3892/or.2010.1082. [DOI] [PubMed] [Google Scholar]
- 25.Chen J, Röcken C, Hoffmann J, et al. Expression of carbonic anhydrase 9 at the invasion front of gastric cancers. Gut. 2005;54(7):920–927. doi: 10.1136/gut.2004.047340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chen CD, Wang CS, Huang YH, et al. Overexpression of CLIC1 in human gastric carcinoma and its clinicopathological significance. Proteomics. 2007;7(1):155–167. doi: 10.1002/pmic.200600663. [DOI] [PubMed] [Google Scholar]
- 27.Shiozaki A, Otsuji E, Marunaka Y. Intracellular chloride regulates the G1/S cell cycle progression in gastric cancer cells. World Journal of Gastrointestinal Oncology. 2011;3(8):119–122. doi: 10.4251/wjgo.v3.i8.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Miyazaki H, Shiozaki A, Niisato N, et al. Chloride ions control the G1/S cell-cycle checkpoint by regulating the expression of p21 through a p53-independent pathway in human gastric cancer cells. Biochemical and Biophysical Research Communications. 2008;366(2):506–512. doi: 10.1016/j.bbrc.2007.11.144. [DOI] [PubMed] [Google Scholar]
- 29.Ohsawa R, Miyazaki H, Niisato N, et al. Intracellular chloride regulates cell proliferation through the activation of stress-activated protein kinases in MKN28 human gastric cancer cells. Journal of Cellular Physiology. 2010;223(3):764–770. doi: 10.1002/jcp.22088. [DOI] [PubMed] [Google Scholar]
- 30.Sasagawa H, Shiozaki A, Iitaka D, et al. Ki-67 labeling index as an independent prognostic factor in human esophageal squamous cell carcinoma. Esophagus. 2012;9(4):195–202. [Google Scholar]
- 31.Nakashima S, Shiozaki A, Ichikawa D, et al. Anti-phosphohistone H3 as an independent prognostic factor in human esophageal squamous cell carcinoma. Anticancer Research. 2013;33(2):461–467. [PubMed] [Google Scholar]
- 32.Shiozaki A, Nakashima S, Ichikawa D, et al. Prognostic significance of p21 expression in patients with esophageal squamous cell carcinoma. Anticancer Research. 2013;33(10):4329–4335. [PubMed] [Google Scholar]
- 33.Ishimoto T, Shiozaki A, Ichikawa D, et al. E2F5 as an independent prognostic factor in esophageal squamous cell carcinoma. Anticancer Research. 2013;33(12):5415–5420. [PubMed] [Google Scholar]
- 34.Jiang B, Hattori N, Liu B, et al. Expression and roles of Cl-channel ClC-5 in cell cycles of myeloid cells. Biochemical and Biophysical Research Communications. 2004;317(1):192–197. doi: 10.1016/j.bbrc.2004.03.036. [DOI] [PubMed] [Google Scholar]
- 35.Menegazzi R, Busetto S, Dri P, Cramer R, Patriarca P. Chloride ion efflux regulates adherence, spreading, and respiratory burst of neutrophils stimulated by tumor necrosis factor-α (TNF) on biologic surfaces. Journal of Cell Biology. 1996;135(2):511–522. doi: 10.1083/jcb.135.2.511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Niisato N, Eaton DC, Marunaka Y. Involvement of cytosolic Cl− in osmoregulation of α-ENaC gene expression. The American Journal of Physiology: Renal Physiology. 2004;287(5):F932–F939. doi: 10.1152/ajprenal.00131.2004. [DOI] [PubMed] [Google Scholar]
- 37.Shiozaki A, Miyazaki H, Niisato N, et al. Furosemide, a blocker of Na+/K+/2Cl− cotransporter, diminishes proliferation of poorly differentiated human gastric cancer cells by affecting G0/G1 state. Journal of Physiological Sciences. 2006;56(6):401–406. doi: 10.2170/physiolsci.RP010806. [DOI] [PubMed] [Google Scholar]
- 38.Hiraoka K, Miyazaki H, Niisato N, et al. Chloride ion modulates cell proliferation of human androgen-independent prostatic cancer cell. Cellular Physiology and Biochemistry. 2010;25(4-5):379–388. doi: 10.1159/000303042. [DOI] [PubMed] [Google Scholar]