

Abstract
Salmonella enterica subspecies enterica serovar 4,[5],12:i:- (S. 4,[5]12:i:-) is believed to be a monophasic variant of S. enterica serovar Typhimurium (S. Typhimurium). This study was conducted to corroborate this hypothesis and to identify the molecular and phenotypic characteristics of the S. 4,[5]12:i:- isolates in Japan. A total of 51 S. 4,[5]12:i:- isolates derived from humans, cattle, swine, chickens, birds, meat (pork), and river water in 15 prefectures in Japan between 2000 and 2010 were analyzed. All the S. 4,[5],12:i:- isolates were identified as S. Typhimurium by two different polymerase chain reactions (PCR) for identification of S. Typhimurium. Of the 51 S. 4,[5],12:i:- isolates, 39 (76.5%) harbored a 94-kb virulence plasmid, which is known to be specific for S. Typhimurium. These data suggest that the S. 4,[5],12:i:- isolates are monophasic variants of S. Typhimurium. The flagellar phase variation is induced by three adjacent genes (fljA, fljB, and hin) in the chromosome. The results of PCR mapping of this region and comparative genomic hybridization analysis suggested that the deletion of the fljAB operon and its flanking region was the major genetic basis of the monophasic phenotype of S. 4,[5],12:i:-. The fljAB operon and hin gene were detectable in eight of the S. 4,[5],12:i:- isolates with common amino acid substitutions of A46T in FljA and R140L in Hin. The introduction of these mutations into S. Typhimurium isolates led to the loss of selectability of isolates expressing the phase 2 H antigen. These data suggested that a point mutation was the genetic basis, at least in part, of the S. 4,[5],12:i:- isolates. The results of phenotypic analysis suggested that the S. 4,[5],12:i:- isolates in Japan consist of multiple distinct clones. This is the first detailed characterization of the S. 4,[5],12:i:- isolates derived from various sources across Japan.
Introduction
Nontyphoidal salmonellae are one of the most common cause of bacterial gastroenteritis in humans as well as salmonellosis in domestic and wild animals worldwide [1]. Serotyping is widely used as an epidemiological typing method to subdivide Salmonella species [2]. Each serovar is identified by the combination of lipopolysaccharide moieties on the cell surface (O antigens) and one or two different flagellar proteins (H antigens). Many serovars have the ability to express two different flagellin proteins, although individual cells can express one of the two flagellins [3]. According to the White-Kaufmann-Le Minor scheme, more than 2500 serovars to date have been recognized in the genus Salmonella [4].
The incidence of human salmonellosis caused by Salmonella enterica subspecies enterica serovar 4,[5],12:i:- (S. 4,[5],12:i:-) has been increasing in Europe, North and South America, and Asia since the mid-1990s [5]–[9]. This serovar is currently among the 10 most common serovars responsible for human infections in a variety of countries, including the second and sixth most prevalent serovar in Germany [10] and the United States [11], respectively. S. 4,[5],12:i:- was also reported as the fourth most common serovar in slaughtered pigs in the EU [10]. In Japan, the rate of distribution of this serovar was more than 2% (ninth most prevalent) in 2009 for the first time, and then remained relatively high to date [12]. Larger outbreaks caused by this serovar have been reported in the United States and Luxemburg [13], [14].
S. 4,[5],12:i:- does not appear in the White-Kaufmann-Le Minor scheme [4] and appears to be a monophasic variant of other biphasic serovars, which have lost phase 2 flagellin or the necessary switching mechanism of phase variation. Seven serovars of S. enterica subsp. enterica with same O and phase 1 H antigens are possible ancestors of this serovar, including Typhimurium (S. Typhimurium), Lagos, Agama, Farsta, Tsevie, Gloucester, and Tumodi [4]. Among these, S. Typhimurium is commonly isolated from humans, animals, and the environment, whereas the others are rarely isolated.
S. Typhimurium is believed to be an ancestor of S. 4,[5],12:i:- based on the following evidence. S. Typhimurium-specific sequences have been detected in S. 4,[5],12:i:- [15]. Some of the S. 4,[5],12:i:- isolates showed the same lysogenic patterns as those of S. Typhimurium by phage typing and displayed pulsed-field gel electrophoresis patterns identical or similar to those of S. Typhimurium isolates [15]–[18]. Different deletions and mutations can be responsible for the lack of phase 2 flagellin expression among the S. 4,[5],12:i:- isolates. Specifically, some isolates from Spain lack 16 genes, including the fljAB operon and flanking genes, which encode phase 2 flagellin expression-related proteins [19]. Some of the S. 4,[5],12:i:- isolates from the United States appeared to have smaller deletions or point mutations in the fljAB operon and/or flanking genes not identified by deoxyribonucleic acid (DNA) probes [18]. To date, no specific point mutations affecting the phase 2 flagellin expression have been identified.
In the present study, we characterized the S. 4,[5],12:i:- isolates derived from various sources in Japan for the following purpose: (i) to corroborate the hypothesis that the S. 4,[5],12:i:- isolates are monophasic variants of S. Typhimurium, (ii) to elucidate the genetic basis of the monophasic phenotype of the S. 4,[5],12:i:- isolates, (iii) to identify the molecular and phenotypic characteristics of the S. 4,[5],12:i:- isolates in Japan because only limited information is currently available.
Materials and Methods
Bacterial isolation, identification, and typing
The S. 4,[5],12:i:- isolates used in this study are listed in Table 1. A total of 51 isolates were derived from humans, cattle, swine, chickens, birds, meat (pork), and river water from 15 prefectures in Japan between 2000 and 2010. The isolates from humans and cattle were obtained from fecal samples of patients or affected animals with different sporadic infections. The swine isolates were obtained from fecal samples of healthy or affected animals. The isolates from chickens and crows were obtained from fecal samples or organs of healthy birds. The isolates from a penguin and a parrot were obtained from organs of diseased birds. The isolates from pork and river water were obtained from a previous monitoring study. S. Typhimurium strain LT2 [20] was used as a positive control for polymerase chain reaction (PCR) analysis or as a reference for comparative genomic hybridization analysis, and strains L-3900 and L-3287 were used to introduce point mutations identified among the S. 4,[5],12:i:- isolates by gene replacement. In Japan, L-3900 was isolated from cattle in 2010, whereas L-3287 was isolated from chicken in 2001. Both these strains were susceptible to kanamycin. The isolation of S. enterica was performed by the staff of the local Institute of Public Health or local Animal Hygiene Service Centers for diagnostic or monitoring purposes. Patient information was anonymized and de-identified prior to analysis. The approval from the Institutional Animal Care and Use Committee is not required in case of isolation for diagnostic purpose. Isolates H1–5 and C1–10 were the same as H1–5 and C1–10, respectively, as described in a previous report [7]. Salmonella spp. were identified based on colony morphology on selective media and biochemical properties, as previously described [21]. Serovar identification was performed by microtiter and slide agglutination methods according to the latest version of the White-Kaufmann-Le Minor scheme [4] using antiserum (Denka Seiken Co., Ltd., Tokyo, Japan). Phage typing was performed using S. Typhimurium typing phages according to the methods and schemes previously described by Anderson et al. [22]. All isolates were maintained at −80°C in Luria–Bertani (LB) broth (Becton, Dickenson and Company, Sparks, MD, USA) containing 25% (v/v) glycerol.
Table 1. Salmonella enterica serovar 4,[5],12:i:- isolates used in this study.
| PCR resultsa | ||||||||||||||
| Isolates | Source | Year | m-PCR | IS200 | up-fljA | fljA-fljB | fljB-hin | hin-down | fin | spvB | 94 kb plasmidb | Phage typec | LDCd | Resistance profilee |
| H1∼4 | Human | 2006 | + | + | - | - | - | + | - | + | + | 193 | + | - |
| H5 | Human | 2007 | + | + | - | - | - | - | - | - | - | 193 | + | ASSu |
| H6 | Human | 2008 | + | + | - | - | - | + | - | + | + | RDNC-a | + | - |
| H7 | Human | 2003 | + | + | - | - | - | + | - | + | + | 193 | + | - |
| H8 | Human | 2007 | + | + | + | + | + | + | - | + | + | 26 | + | - |
| H9∼11 | Human | 2007 | + | + | - | - | - | + | - | + | + | RDNC-a | + | - |
| H12 | Human | 2004 | + | + | - | - | - | + | - | - | - | RDNC-c | + | - |
| H13 | Human | 2007 | + | + | - | - | - | - | - | - | - | 193 | + | SSuT |
| H14 | Human | 2002 | + | + | - | - | - | - | - | + | + | UT | + | ASuT |
| C1 | Cattle | 2003 | + | + | - | - | - | + | - | + | + | RDNC-a | - | - |
| C2 | Cattle | 2005 | + | + | - | - | - | + | - | + | + | RDNC-a | - | - |
| C3∼4 | Cattle | 2007 | + | + | - | - | - | + | - | + | + | RDNC-a | - | - |
| C5∼8 | Cattle | 2008 | + | + | - | - | - | + | - | + | + | RDNC-a | - | - |
| C9∼10 | Cattle | 2008 | + | + | - | - | - | - | - | + | + | RDNC-a | + | A |
| C11 | Cattle | 2004 | + | + | - | - | - | + | - | + | + | RDNC-a | + | - |
| C12 | Cattle | 2005 | + | + | - | - | - | + | - | + | + | 120 | + | - |
| C13 | Cattle | 2005 | + | + | + | + | + | + | - | + | + | RDNC-b | + | - |
| C14 | Cattle | 2008 | + | + | - | - | - | - | - | - | - | UT | + | ASSuT |
| C15 | Cattle | 2007 | + | + | - | - | - | + | - | + | + | RDNC | + | - |
| C16 | Cattle | 2010 | + | + | - | - | - | + | - | + | + | RDNC-a | + | - |
| C17 | Cattle | 2010 | + | + | - | - | - | + | - | + | + | RDNC-b | + | A |
| S1 | Swine | 2008 | + | + | - | - | - | - | - | - | - | UT | + | ASSuT |
| S2 | Swine | 2009 | + | + | - | - | - | - | - | - | - | UT | + | ASSu |
| S3 | Swine | 2002 | + | + | - | - | - | - | - | - | - | RDNC-d | + | Ssu |
| S4 | Swine | 2003 | + | + | - | - | - | - | - | - | - | RDNC-d | + | SSuT |
| S5 | Swine | 2008 | + | + | - | - | - | - | - | - | - | 193 | + | SSuT |
| S6 | Swine | 2009 | + | + | - | - | - | - | + | + | + | 27 | + | ASSuT |
| K1 | Chicken | 2001 | + | + | - | - | - | + | - | + | + | RDNC-b | + | - |
| K2 | Chicken | 2004 | + | + | - | - | - | + | - | - | - | RDNC | + | - |
| K3 | Chicken | 2005 | + | + | - | - | - | + | - | - | - | RDNC-c | + | - |
| K4 | Chicken | 2006 | + | + | - | - | - | + | - | - | - | RDNC-c | + | - |
| K5 | Chicken | 2010 | + | + | - | - | - | + | - | + | + | RDNC | + | ASuT |
| B1 | Penguin | 2009 | + | + | + | + | + | + | - | + | + | RDNC | + | - |
| B2∼3 | Crow | 2000 | + | + | + | + | + | + | - | + | + | RDNC-e | + | - |
| B4 | Parrot | 2005 | + | + | + | + | + | + | - | + | + | RDNC-e | + | - |
| M1 | Pork | 2005 | + | + | - | - | - | + | - | + | + | RDNC-a | + | - |
| M2 | Pork | 2007 | + | + | - | - | - | + | - | + | + | RDNC-a | + | - |
| R1 | River water | 2007 | + | + | + | + | + | + | - | + | + | 26 | + | - |
| R2 | River water | 2007 | + | + | - | - | - | + | - | + | + | RDNC-a | + | ASu |
| R3 | River water | 2007 | + | + | + | + | + | + | - | + | + | 26 | + | - |
m-PCR, multiplex PCR to identify S. Typhimurium [25]; IS200, PCR to identify S. Typhimurium [15]; up-fljA, boundary region of fljA and its upstream intergenic region; fljA–fljB, boundary region of fljA and fljB; fljB–hin, boundary region of fljB and hin; hin-down, boundary region of hin and its downstream intergenic region; +, positive; -,negative.
+, presence; -, absence.
RDNC, reacted but did not conform; RDNC-a–e, same letter indicates the same lysogenic patterns among RDNC isolates.
LDC, lysine decarboxylase; +, positive; -, negative.
A, ampicillin; S, streptomycin; Su, sulfamethizole; T, tetracycline; -, pansusceptible.
Antimicrobial susceptibility testing
The Kirby–Bauer disc diffusion test was performed using Mueller–Hinton agar plates (Becton, Dickenson and Company) according to Clinical and Laboratory Standard Institute standards [23] using the following antimicrobials: ampicillin (10 µg), cefazolin (30 µg), kanamycin (30 µg), streptomycin (10 µg), tetracycline (30 µg), chloramphenicol (30 µg), fosfomycin (50 µg), colistin (10 µg), sulfamethizole (250 µg), and nalidixic acid (30 µg) (Becton, Dickinson and Company).
Plasmid isolation
Plasmid DNA was isolated by the method described by Kado and Liu [24] followed by phenol–chloroform extraction. The Bac-Tracker Supercoiled DNA ladder (Epicentre Biotechnologies, Madison, WI, USA) and a 94-kb plasmid from S. Typhimurium LT2 were used as size markers.
PCR and sequencing
All primers used in this study were purchased from Hokkaido System Science Co., Ltd. (Hokkaido, Japan) and are listed in Table S1. A single colony of each bacterial isolate was suspended in 50 µL of 25 mM NaOH and boiled for 5 min. After addition of 4 µL of 1 M Tris-HCl (pH 8.0), the suspension was centrifuged and the supernatant was used as a template DNA. Amplification was performed using an iCycler apparatus (Bio-Rad Laboratories, Hercules, CA, USA). Takara Ex Taq (Takara Bio Inc., Shiga, Japan) was used as DNA polymerase for each monoplex PCR. The Salmonella serovar Typhimurium Identification Kit (Takara Bio Inc.) was used to detect S. Typhimurium-related genes, including STM0292, STM2235, and STM4493, by multiplex PCR (m-PCR) as previously described [25]. Some of the PCR products were purified using the illustra ExoStar Kit for Enzymatic PCR and Sequencing Clean-up (GE Healthcare UK Ltd., Buckinghamshire, UK). Nucleotide sequences were determined on both strands using an Applied Biosystems 3130 xl genetic analyzer with the BigDye Terminator cycle sequencing kit (version 3.1; Applied Biosystems, Foster City, CA, USA). The sequences were assembled with Sequencher version 4 (Hitachi Solutions, Kanagawa, Japan) and the DNA alignments and deduced amino acid sequences were examined using the Basic Local Alignment Search Tool (http://blast.ncbi.nlm.nih.gov/Blast.cgi).
Comparative genomic hybridization (CGH) analysis
Copy number analysis of the selected S. 4,[5],12:i:- isolates was performed using the whole genomic CGH array (Roche NimbleGen, Inc., Madison, WI, USA) at their facility in Iceland according to previously published methods with some modifications [26]. In brief, a tiling array was designed with a mean probe density of 1 probe/10 bp, 50–75-mer length using the S. Typhimurium strain LT2 sequences of the chromosome (AE006468) and pSLT plasmid (AE006471). Labeling was performed using the NimbleGen Dual Color Labeling Kit according to the manufacturer's protocols. In brief, each DNA sample (1 µg) was denatured at 98°C in the presence of one optical density of 5′-Cy3- or 5′-Cy5-labeled random nonamer. The denatured sample was chilled on ice and then incubated with 100 U of (exo-) Klenow fragment and dNTP mix for 2 h at 37°C. Reactions were terminated by addition of 0.5 M ethylenediaminetetraacetic acid (pH 8.0) and the end products were precipitated with isopropanol and resuspended in water. The Cy-labeled test and reference samples (Cy3 and Cy5, respectively) were combined (31 µg each) and dried down by vacuum centrifugation. Each sample was rehydrated in 5.6 µL of PCR grade water included in the NimbleGen Sample Tracking Control Kit and added to the hybridization buffers included in the NimbleGen Hybridization Buffer Kit, denatured at 95°C for 5 min, and then cooled to 42°C. Hybridizations were conducted for 40–72 h at 42°C using the NimbleGen Hybridization System. The arrays were washed using the NimbleGen Wash Buffer Kit and immediately dried down by centrifugation. Arrays were scanned at a resolution of 5 µm using the GenePix4000B scanner (Axon Instruments, Molecular Devices Corp., Sunnyvale, CA, USA). Data were extracted from scanned images using NimbleScan software (Roche NimbleGen, Inc.), which allows for automated grid alignment, extraction, and generation of data files.
Gene replacement in S. Typhimurium
The primer pair GR1F and GR1R was used to amplify a region containing a point mutation in fljA, and the primer pair GR2F and GR2R was used to amplify a region containing a point mutation in hin from the S. 4,[5],12:i:- isolates by PCR (Fig. 1, Table S1). After digestion with the XbaI and HindIII restriction enzymes, the resulting fragment was cloned to the temperature-sensitive vector pTH18ks1 [27] and used as a vector for gene replacement. S. Typhimurium strains L-3900 and L-3287 were transformed with one of each vector by electroporation. The cells were spread on LB agar (Becton, Dickinson and Co.) supplemented with kanamycin and incubated at 28°C for 18 h. The colonies were then streaked on the same agar plates pre-warmed to 42°C and incubated at 42°C for 18 h. The single crossover strains were purified under the same conditions and then passaged at 28°C several times. The double crossover strains were screened by allele-specific PCR using the primer pair SNP1 and GR1R for detection of a point mutation in fljA and the primer pair SNP2 and GR2R for detection of a point mutation in hin (Fig. 1, Table S1). The introduced point mutation was verified by PCR and sequencing using appropriate primers. S. Typhimurium derivatives with the point mutation in hin were transformed with the vector for the fljA gene replacement. Double crossover strains were selected using the abovementioned procedure to obtain double mutants.
Figure 1. Schematic view of genetic organization of the chromosomal region related to flagellar phase variation of S. Typhimurium.
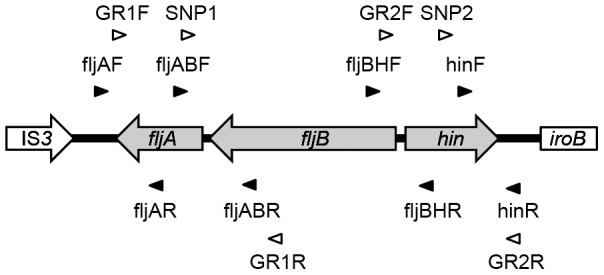
Gray arrows indicate gene related to phase variation. Closed triangles indicate the primer locations for polymerase chain reaction mapping. Open triangles indicate the primer locations for mutant construction.
Estimation of phase variation frequency
Phase variation frequency was estimated by the method described by Stocker [28] with minor modifications. In brief, the tested strains expressing the H-i antigen (phase 1) were serially passaged in LB broth media until the estimated number of generations reached 110. The culture was diluted to yield approximately 100 colonies on LB agar in 90-mm petri dishes and spread with a plastic spreader. The plates were incubated at 30°C for 18 h until the colonies grew to 1 mm in diameter. The plates were cooled in a refrigerator, and then 7 mL of semi-solid agar containing 0.35% Bacto Agar (Becton, Dickinson and Company) was pipetted on top. This semi-solid agar contained 0.7% (v/v) anti-H-i serum (Denka Seiken Co.). Once solidified, the plates were incubated at 37°C for 1–2 h. Colonies expressing the H-1,2 (phase 2) antigen were found to be surrounded by a wide zone of opacity with an indefinite edge, indicating that the organisms were swarming out into the semi-solid agar. However, colonies expressing the phase 1 antigen were surrounded by a narrow dense zone of opacity with a clear-cut edge. The frequencies of swarming colonies among ca. 7000 colonies were calculated, and then the frequency was divided by the number of generations to determine the frequency of phase variation per generation. Each experiment was performed thrice. Differences in the results were tested using the two-tailed unpaired Student's t test.
Results and Discussion
S. 4,[5],12:i:- is very likely a monophasic variant of S. Typhimurium
To prove the hypothesis that S. 4,[5],12:i:- is a monophasic variant of S. Typhimurium, several molecular characteristics of the S. 4,[5],12:i:- isolates were investigated. In the m-PCR to identify S. Typhimurium, three serovar-related genomic regions were successfully amplified from all the S. 4,[5],12:i:- isolates tested in this study (Table 1). No false positives were observed using 117 Salmonella serovars, with the exception of S. 4,[5],12:i:- [25], which strongly suggested that S. 4,[5],12:i:- originated from S. Typhimurium. The results of PCR analysis to detect the fliA–fliB intergenic region also support this statement. The location of IS200 between the genes fliA and fliB can be used as a specific marker for S. Typhimurium [29]. The amplicon sizes from the fliA–fliB intergenic regions from S. Typhimurium and other serovars were expected to be 1000 and 250 bp, respectively [15]. A 1000-bp amplicon was successfully detected in all the S. 4,[5],12:i:- isolates. These data suggest that S. 4,[5],12:i:- is a monophasic variant of S. Typhimurium. In other words, the S. Typhimurium-specific m-PCR, and PCR might be useful to verify that the tested S. 4,[5],12:i:- isolate is a monophasic variant of S. Typhimurium.
As shown in Table 1, 39 (76.5%) of the 51 S. 4,[5],12:i:- isolates harbored a 94-kb plasmid with or without other plasmids. The spvB gene, which is a marker of the S. enterica virulence plasmid [30], was detected by PCR in all of the isolates with the 94-kb plasmid, thereby supporting the possibility that these isolates originated from S. Typhimurium. However, the lack of this plasmid does not contradict the possibility that the isolate is S. Typhimurium. The prevalence of the virulence plasmid in the S. Typhimurium isolates obtained from swine with systemic infections was 92%, whereas less than 20% in isolates from diarrhea samples and animals without any symptoms [31].
Deletion is a major basis of the monophasic phenotype of the S. 4,[5],12:i:- isolates
Most S. enterica serovars possess two different flagellin proteins, including FliC (phase 1) and FljB (phase 2), which are encoded by the genes fliC and fljB, respectively. Flagellar phase variation is induced by inversion of the genetic region called the H segment, which contains the hin gene encoding for DNA invertase and the promoter for the fljB gene. The fljB constitutes an operon with the fljA gene, which encodes a negative regulator of fliC expression. FljA binds to the operator region of FliC mRNA and inhibits its translation, leading to the rapid degradation of FliC mRNA. When the H segment is in the “on” state, both fljB and fljA are transcribed, resulting in synthesis of phase 2 flagellin and inhibition of phase 1 flagellin. However, when the H segment is switched to the “off” state, neither fljB nor fljA are transcribed, resulting in the synthesis of phase 1 flagellin only (Fig. S1) [32], [33].
To determine whether the S. 4,[5],12:i:- isolates maintained the genetic structure of the fljAB–hin region, PCR mapping [7] to detect the boundary region of each gene was performed using the S. Typhimurium LT2 DNA sequence as a reference. The amplification targets were as follows; up-fljA, the boundary region of fljA and its upstream intergenic region; fljA–fljB, the boundary region of fljA and fljB; fljB–hin, the boundary region of fljB and hin; and hin-down, the boundary region of hin and its downstream intergenic region (Fig. 1). As shown in Table 1, three amplification patterns were observed: positive for only hin-down (31 isolates), all negative (12 isolates), and all positive (eight isolates). In eight of the positive isolates, the whole fljAB–hin structure was detectable with two common amino-acid substitutions: A46T in FljA and R140L in Hin. These amino-acid substitutions were not observed in six S. Typhimurium wild-type strains isolated in Japan including L-3900 and L-3287 (data not shown).
To compare the whole genome sequences of the S. 4,[5],12:i:- isolates with that of S. Typhimurium strain LT2, one of the each representative isolate from the three amplification patterns determined by PCR mapping was analyzed by CGH. As shown in Table 2 and Fig. S2, whole sequences of prophages Fels-1 and Fels-2 were not detectable among the three isolates. In strains C1 and C9, an additional 36-kb sequence downstream of Fels-2 containing the fljAB operon was not detectable (Fig. 2). In addition, the whole sequence of the Gifsy-1 prophage was not detectable in strains C1 and C9, whereas part of the Gifsy-1 prophage sequence was not detected in strain C13 (Table 2 and Fig. S2). Among these, a broad scale deletion event stretching from the Fels-2 prophage to the fljAB–hin region was determined as the genetic basis of the monophasic phenotype of the S. 4,[5],12:i:- isolates C1 and C9.
Table 2. Chromosomal genes that lacks in the CGH tested isolates.
| Presence/Absencea | ||||||
| NC_00319 tag number | Position (start-end) | Size (bp) | Description | C1 | C9 | C13 |
| STM0276–STM0279 | 316895–319135 | 2241 | putative cytoplasmic/periplasmic | + | + | - |
| proteins | ||||||
| STM0893–STM0929 | 962638–1005280 | 42643 | Fels-1 prophage | - | - | - |
| STM1011–STM1019 | 1104868–1109917 | 5050 | part of Gifsy-2 prophage | + | + | - |
| STM1555–STM1557 | 1632448–1635078 | 2631 | putative Na+/H+ antiporter | + | + | - |
| and others | ||||||
| STM2585–STM2636 | 2730851–2776671 | 45821 | Gifsy-1 prophage | - | - | -b |
| STM2694–STM2739 | 2844326–2877883 | 33558 | Fels-2 prophage | - | - | - |
| STM2740–STM2771 | 2877884–2914231 | 36348 | fljAB and upstream genes | - | - | + |
| STM2951 | 3094339–3096696 | 2358 | ygcF | + | + | - |
| STM3113 | 3271613–3272493 | 881 | nupG | - | - | + |
| STM3255–STM3260 | 3425101–3430141 | 5041 | putative phosphotransferase system | - | - | + |
| and others | ||||||
Chromosomal genes with log2 ratios <−0.5 were identified as absent genes.
Five genes (7782 bp) of the Gifsy-1 prophage were absent.
Figure 2. Partial quantitative data from the comparative genomic hybridization of the S. 4,[5],12:i:- isolates.
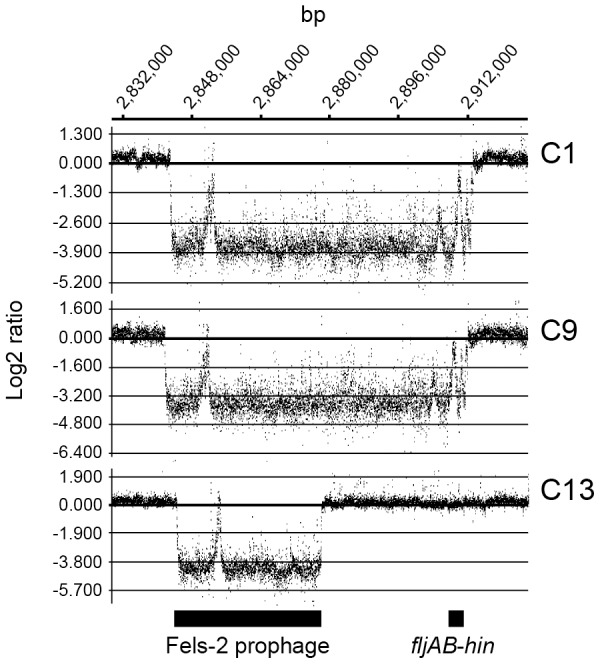
The ruler indicates the nucleotide number of S. Typhimurium LT2 chromosome (AE006468). The vertical scale indicates the log2 ratio of the signal intensities. C1, C9, and C13 indicate the name of the isolates listed in Table 1. The underlying bold lines indicate the location of specific genetic structures in the chromosome.
S. Typhimurium strain LT2 contains the DNA invertase gene fin, which contributes to the phase induction of H antigens other than hin. The fin gene is located in the Fels-2 prophage of S. Typhimurium LT2 [34]. All the S. 4,[5],12:i:- isolates were PCR-negative for fin, except for one isolate obtained from a swine, suggesting that the Fels-2 prophage was not distributed among most of the S. 4,[5],12:i:- isolates, as indicated by the CGH analysis of representative isolates.
In total, the presence/absence patterns of isolates C1 and C9 were identical, whereas strain C13 was different from other strains. Approximately 80% or more of the total length of the absence region corresponded to the Fels-1, Fels-2, Gifsy-1, and Gifsy-2 prophages (Table 2). Garaizar et al. [19] reported the deletion of most of the Fels-1 and Fels-2 sequences and a partial sequence of Gifsy-1 among the S. 4,[5],12:i:- isolates in Spain. Prophage sequences may be selectively neutral for this serovar.
Point mutations reduce the phase variation frequency of S. Typhimurium
To manifest the effect of the amino acid substitutions, A46T in FljA and R140L in Hin observed in the eight S. 4,[5],12:i:- isolates by phase variation frequency analysis, these point mutations were independently or simultaneously introduced to S. Typhimurium strains L-3900 and L-3287. As shown in Table 3, phase variation was successfully observed in both parental strains and the frequency from phase 1 to phase 2 of L-3900 was 1.84×10−4, whereas that of L-3287 was less than that of the detection limit (10−6). The phase variation frequency of the L-3900 fljA mutant was significantly lower (p = 0.04) than that of the parental strain. An isolate expressing the phase 2 H antigen was successfully selected from this mutant, but not from any other mutants. These data suggest that the point mutations reduced the phase variation frequency and may be the genetic basis of the monophasic phenotype of all fljAB–hin detectable isolates.
Table 3. Expression of phase 2 antigen and phase variation frequency.
| Genotypea | Selectability | Phase variation | ||
| Parental strain | fljA | hin | of phase 2b | frequencyc |
| L-3900 | WT | WT | + | 1.84×10−4 |
| A46T | WT | + | 9.11×10−5 * | |
| WT | R140L | - | <10−6 | |
| A46T | R140L | - | <10−6 | |
| L-3287 | WT | WT | + | <10−6 |
| A46T | WT | - | ND | |
| WT | R140L | - | ND | |
| A46T | R140L | - | ND | |
WT, wild type; A46T and R140L, amino acid substitutions.
+, selectable; -, not detected.
ND, not done;
*, significantly lower than parental strain (p = 0.04).
Hin invertase catalyzes DNA inversion of the H segment. This site-specific recombination event controls the alternate expression of two flagellin genes by reversing the orientation of the fljB promoter [32], [33]. The interaction between Hin invertase and the target DNA has been fully elucidated. The 52 carboxyl-terminal residues is the DNA-binding domain of the Hin invertase. Particularly, the sequence G139-R140-P141-R142 is essential to maintain DNA binding ability. The deletion of residues G139 and R140 abolished the sequence-specific binding to DNA [35]. The fin gene located in the Fels-2 prophage region encodes an invertase that can support inversion of the H segment without Hin invertase. As both the strains L-3900 and L-3287 were found to be negative for the fin gene by PCR, R140L in Hin may diminish the DNA-binding ability; thus, resulting in a reduction in the phase variation frequency. No information is available regarding the effect of the A46T substitution in FljA on the phase variation frequency to date.
S. 4,5,12:i:- isolates in Japan consist of multiple distinct clones
As shown in Table 1, phage typing of the 51 S. 4,[5],12:i:- isolates examined using the S. Typhimurium typing phages identified four phage types: DT193 (eight isolates), DT26 (three isolates), DT27 (one isolate), and DT120 (one isolate). The remaining 34 isolates were RDNC (reacts with phages but does not confirm to a recognized pattern) and four isolates were UT (untypable). Among the 34 RDNC isolates, five lysogenic patterns were observed in more than one isolate, which were named RDNC-a–e. All eight isolates of lysine decarboxylase-negative S. 4,[5],12:i:- belonged to RDNC-a. In Germany and Switzerland, DT193 was the most prevalent definitive phage type among the S. 4,[5],12:i:- isolates [6], [17]. Most of the S. 4,[5],12:i:- isolates from Spain belonged to the definitive phage-type U302 [16]. The S. 4,[5],12:i:- isolates from Japan appeared to consist of multiple clones with greater variation than those from European countries.
Antimicrobial susceptibility testing using nine antimicrobials showed that 14 (27.5%) of the 51 S. 4,[5],12:i:- isolates were resistant to one or more antimicrobials among ampicillin, streptomycin, sulfamethizole, and tetracycline. The remaining 37 isolates were pan-susceptible (Table 1). According to the data published to date, most of the S. 4,[5],12:i:- isolates from Europe (i.e., Spain, Germany, and Switzerland) appeared to have a multidrug resistance phenotype, whereas most of the S. 4,[5],12:i:- isolates from North and South American countries (i.e., USA and Brazil) appeared to be pan-susceptible or resistant to only a few antimicrobials [6], [9], [16], [17]. In Japan, pan-susceptible isolates were dominant, although multidrug resistance isolates with resistance to up to four antimicrobials were detected. All of the six isolates obtained from swines exhibited resistance to multiple antimicrobials. This may reflect the extensive use of antibiotics as feed additives in the pig industry in Japan [36].
Conclusions
The results of the molecular characterization of the 51 S. 4,[5],12:i:- isolates derived from various sources in Japan suggested that these isolates were very likely monophasic variants of S. Typhimurium. Deletion and point mutations were the bases of the monophasic phenotype of the S. 4,[5],12:i:- isolates. The results of phenotypic characterization suggested these isolates consisted of multiple distinct clones.
Supporting Information
A model for the molecular mechanism of phase variation in Salmonella cited from Yamamoto and Kutsukake [33] with slight modifications. This system consists of two major parts: (i) the switching mechanisms of fljB promoter orientation by inversion of H segments and (ii) the FljA-mediated translational repression of fliC mRNA, leading to the rapid degradation of the mRNA. IR, inverted repeat; fljBp, fljB promoter; fliCp, fliC promoter; OP, operator region.
(TIF)
Quantitative data from the comparative genomic hybridization of the S . 4,[5],12:i:- isolates. The ruler indicates the nucleotide number of S. Typhimurium LT2 chromosome (AE006468). The vertical scale indicates the log2 ratio of the signal intensities. C1, C9, and C13 indicate the name of the isolates listed in Table 1. The underlying bold lines indicate the locations of prophages in the chromosome.
(TIF)
Primers used in this study.
(DOCX)
Funding Statement
This work was supported by grants-in-aid from the Ministry of Agriculture, Forestry and Fisheries of Japan (Research project for ensuring food safety from farm to table 24-7304), and the Ministry of Health, Labour, and Welfare of Japan (H24-Shokuhin-Ippan-008). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, et al. (2010) The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis 50: 882–889. [DOI] [PubMed] [Google Scholar]
- 2.Grimont PAD, Grimont F, Bouvet P (2000) Taxonomy of the Genus Salmonella In: Wray C, Wray, A., etidor. Salmonella in Domestic Animals. Wallingford: CABI Publishing. pp. 1–17. [Google Scholar]
- 3. Iino T (1969) Genetics and chemistry of bacterial flagella. Bacteriol Rev 33: 454–475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Grimont P, Weil F (2007) Antigenic formulae of the Salmonella serovars, 9th ed. World Health Organization Centre for Reference and Research on Salmonella. Paris, France: Pasteur Institute. [Google Scholar]
- 5. Dionisi AM, Graziani C, Lucarelli C, Filetici E, Villa L, et al. (2009) Molecular characterization of multidrug-resistant strains of Salmonella enterica serotype Typhimurium and Monophasic variant (S. 4,[5],12:i:-) isolated from human infections in Italy. Foodborne Pathog Dis 6: 711–717. [DOI] [PubMed] [Google Scholar]
- 6. Gallati C, Stephan R, Hachler H, Malorny B, Schroeter A, et al. (2013) Characterization of Salmonella enterica subsp. enterica serovar 4,[5],12:i:- clones isolated from human and other sources in Switzerland between 2007 and 2011. Foodborne Pathog Dis 10: 549–554. [DOI] [PubMed] [Google Scholar]
- 7. Ido N, Kudo T, Sasaki K, Motokawa M, Iwabuchi K, et al. (2011) Molecular and phenotypic characteristics of Salmonella enterica serovar 4,5,12:i:- isolated from cattle and humans in Iwate Prefecture, Japan. J Vet Med Sci 73: 241–244. [DOI] [PubMed] [Google Scholar]
- 8. Majtan V, Majtanova L, Majtan J (2011) Phenotypic and molecular characterization of human Salmonella enterica serovar 4,[5],12:i:- isolates in Slovakia. Curr Microbiol 63: 491–495. [DOI] [PubMed] [Google Scholar]
- 9. Switt AI, Soyer Y, Warnick LD, Wiedmann M (2009) Emergence, distribution, and molecular and phenotypic characteristics of Salmonella enterica serotype 4,5,12:i. Foodborne Pathog Dis 6: 407–415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. European Food Safety Authority (2008) Report of the Task Force on Zoonoses Data Collection on the analysis of the baseline survey on the prevalence of Salmonella in slaughter pigs, in the EU, 2006–2007. EFSA Journal 135: 1–111. [Google Scholar]
- 11.Centers for Disease Control and Prevention (2008) Salmonella surveillance: annual summary, 2006.
- 12.National Institute of Infectious Diseases (2014) Infectious Agents Surveillance Report. Available: http://idsc.nih.go.jp/iasr/index.html. Accessed 2014 Jun 23.
- 13.Centers for Disease Control and Prevention (2007) Investigation of outbreaks of human infections caused by Salmonella serotype I 4,[5],12:i:-.
- 14. Mossong J, Marques P, Ragimbeau C, Huberty-Krau P, Losch S, et al. (2007) Outbreaks of monophasic Salmonella enterica serovar 4,[5],12:i:- in Luxembourg, 2006. Euro Surveill 12: E11–12. [DOI] [PubMed] [Google Scholar]
- 15. Echeita MA, Herrera S, Usera MA (2001) Atypical, fljB-negative Salmonella enterica subsp. enterica strain of serovar 4,5,12:i:- appears to be a monophasic variant of serovar Typhimurium. J Clin Microbiol 39: 2981–2983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. de la Torre E, Zapata D, Tello M, Mejia W, Frias N, et al. (2003) Several Salmonella enterica subsp. enterica serotype 4,5,12:i:- phage types isolated from swine samples originate from serotype Typhimurium DT U302. J Clin Microbiol 41: 2395–2400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Hauser E, Tietze E, Helmuth R, Junker E, Blank K, et al. (2010) Pork contaminated with Salmonella enterica serovar 4,[5],12:i:-, an emerging health risk for humans. Appl Environ Microbiol 76: 4601–4610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Zamperini K, Soni V, Waltman D, Sanchez S, Theriault EC, et al. (2007) Molecular characterization reveals Salmonella enterica serovar 4,[5],12:i:- from poultry is a variant Typhimurium serovar. Avian Dis 51: 958–964. [DOI] [PubMed] [Google Scholar]
- 19. Garaizar J, Porwollik S, Echeita A, Rementeria A, Herrera S, et al. (2002) DNA microarray-based typing of an atypical monophasic Salmonella enterica serovar. J Clin Microbiol 40: 2074–2078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. McClelland M, Sanderson KE, Spieth J, Clifton SW, Latreille P, et al. (2001) Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 413: 852–856. [DOI] [PubMed] [Google Scholar]
- 21.Edwards PR, Ewing WH (1986). Edwards and Ewing's identification of Enterobacteriaceae, 4th ed. New York, NY: Elsevier Science Publishing Co., Inc. [Google Scholar]
- 22. Anderson ES, Ward LR, Saxe MJ, de Sa JD (1977) Bacteriophage-typing designations of Salmonella typhimurium . J Hyg (Lond) 78: 297–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Clinical and Laboratory Standards Institute (2008) Performance Standards for Antimicrobial Disk and Dilution Susceptibility Test for Bacteria Isolated from Animals-Third Edition: Approved Standatrd M31-A3. Wayne, PA. Wayne, PA.
- 24. Kado CI, Liu ST (1981) Rapid procedure for detection and isolation of large and small plasmids. J Bacteriol 145: 1365–1373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Akiba M, Kusumoto M, Iwata T (2011) Rapid identification of Salmonella enterica serovars, Typhimurium, Choleraesuis, Infantis, Hadar, Enteritidis, Dublin and Gallinarum, by multiplex PCR. J Microbiol Methods 85: 9–15. [DOI] [PubMed] [Google Scholar]
- 26. Selzer RR, Richmond TA, Pofahl NJ, Green RD, Eis PS, et al. (2005) Analysis of chromosome breakpoints in neuroblastoma at sub-kilobase resolution using fine-tiling oligonucleotide array CGH. Genes Chromosomes Cancer 44: 305–319. [DOI] [PubMed] [Google Scholar]
- 27. Hashimoto-Gotoh T, Yamaguchi M, Yasojima K, Tsujimura A, Wakabayashi Y, et al. (2000) A set of temperature sensitive-replication/-segregation and temperature resistant plasmid vectors with different copy numbers and in an isogenic background (chloramphenicol, kanamycin, lacZ, repA, par, polA). Gene 241: 185–191. [DOI] [PubMed] [Google Scholar]
- 28. Stocker BA (1949) Measurements of rate of mutation of flagellar antigenic phase in Salmonella typhi-murium . J Hyg (Lond) 47: 398–413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Burnens AP, Stanley J, Sack R, Hunziker P, Brodard I, et al. (1997) The flagellin N-methylase gene fliB and an adjacent serovar-specific IS200 element in Salmonella Typhimurium. Microbiology 143: 1539–1547. [DOI] [PubMed] [Google Scholar]
- 30. Gulig PA, Danbara H, Guiney DG, Lax AJ, Norel F, et al. (1993) Molecular analysis of spv virulence genes of the Salmonella virulence plasmids. Mol Microbiol 7: 825–830. [DOI] [PubMed] [Google Scholar]
- 31. Namimatsu T, Asai T, Osumi T, Imai Y, Sato S (2006) Prevalence of the virulence plasmid in Salmonella Typhimurium isolates from pigs. J Vet Med Sci 68: 187–188. [DOI] [PubMed] [Google Scholar]
- 32. Simon M, Zieg J, Silverman M, Mandel G, Doolittle R (1980) Phase variation: evolution of a controlling element. Science 209: 1370–1374. [DOI] [PubMed] [Google Scholar]
- 33. Yamamoto S, Kutsukake K (2006) FljA-mediated posttranscriptional control of phase 1 flagellin expression in flagellar phase variation of Salmonella enterica serovar Typhimurium. J Bacteriol 188: 958–967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Kutsukake K, Nakashima H, Tominaga A, Abo T (2006) Two DNA invertases contribute to flagellar phase variation in Salmonella enterica serovar Typhimurium strain LT2. J Bacteriol 188: 950–957. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Feng JA, Johnson RC, Dickerson RE (1994) Hin recombinase bound to DNA: the origin of specificity in major and minor groove interactions. Science 263: 348–355. [DOI] [PubMed] [Google Scholar]
- 36. Takahashi T, Asai T, Kojima A, Harada K, Ishihara K, et al. (2006) Present situation of national surveillance of antimicrobial resistance in bacteria isolated from farm animals in Japan and correspondence to the issue. J Jap Assoc Infect Dis 80: 185–195 (in Japanese with English summary). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A model for the molecular mechanism of phase variation in Salmonella cited from Yamamoto and Kutsukake [33] with slight modifications. This system consists of two major parts: (i) the switching mechanisms of fljB promoter orientation by inversion of H segments and (ii) the FljA-mediated translational repression of fliC mRNA, leading to the rapid degradation of the mRNA. IR, inverted repeat; fljBp, fljB promoter; fliCp, fliC promoter; OP, operator region.
(TIF)
Quantitative data from the comparative genomic hybridization of the S . 4,[5],12:i:- isolates. The ruler indicates the nucleotide number of S. Typhimurium LT2 chromosome (AE006468). The vertical scale indicates the log2 ratio of the signal intensities. C1, C9, and C13 indicate the name of the isolates listed in Table 1. The underlying bold lines indicate the locations of prophages in the chromosome.
(TIF)
Primers used in this study.
(DOCX)