

Abstract
Growing evidence suggests epigenetic alteration is involved during the development and progression of prostate cancer. Previously, we found Nrf2, a key regulator of cellular antioxidant defense systems, was silenced through epigenetic mechanism during tumorigenesis in vivo TRAMP mice and in vitro TRAMP C1 cells. Sulforaphane (SFN) in cruciferous vegetable has been demonstrated to be a potent cancer prevention agent for years. The aim of this study is to investigate the potential of SFN to restore Nrf2 expression in TRAMP C1 cells through epigenetic modifications. Bisulfite genomic sequencing results indicated that SFN treatment led to demethylation of the first 5 CpGs in the promoter region of the Nrf2 gene in TRAMP C1 cells. Using methylation DNA immunoprecipitation (MeDIP) assay, SFN significantly reduced the ratio of anti-mecyt antibody binding to the Nrf2 promoter containing the first 5 CpGs. SFN increased mRNA and protein expressions of Nrf2 and Nrf2 downstream target gene NQO-1. In addition, SFN decreased the protein levels of DNMT1 and DNMT3a. SFN treatment also attenuated the protein expression levels of HDACs 1, 4, 5, and 7 while increased the level of active chromatin marker acetyl-Histone 3 (Ac-H3). SFN treatments also increased chromatin-immunoprecipitated DNA of Nrf2 gene promoter using anti-Ac-H3 antibody. Taken together, our current study shows that SFN regulates Nrf2's CpGs demethylation and reactivation in TRAMP C1 cells, suggesting SFN may exert its chemopreventive effect in part via epigenetic modifications of Nrf2 gene with subsequent induction of its downstream anti-oxidative stress pathway.
Keywords: epigenetics, DNA methylation, sulforaphane, Nrf2, prostate cancer
1. Introduction
Prostate cancer is one of the most deadly cancers among men in the United States [1]. It has been suggested that the incidence of prostate cancer is associated with increased intracellular oxidative stress [2-4], which is characterized by excess generation of reactive oxygen species (ROS) and/or reduction of antioxidant capacities. Chronic oxidative stress and associated pathological conditions such as inflammation had been demonstrated to drive genomic instability, genetic mutation and neoplastic transformation [5-8], thus it could serve as an essential factor in the development and progression of prostate cancer. Nuclear factor erythroid-2 related factor-2 (Nrf2 or NFE2L2), a helix-loop-helix basic leucine zipper transcription factor, is a key regulator in the cellular defense system against oxidative stress [9-13]. Through its binding to antioxidant response element (ARE) in the promoter region [14, 15], activation of Nrf2 results in the induction of a series of anti-oxidative stress/detoxifying enzymes and proteins, such as heme oxygenase-1 (HO-1), NAD(P)H:quinone oxidoreductase-1 (NQO-1), UDP-glucuronosyltransferases (UGT), and glutathione-S-transferases (GST) [16, 17] among others. It has been reported that Nrf2-deficient mice has an increased susceptibility towards carcinogen induced tumorigenesis [18, 19]. Interestingly, our previous work found that the expression of Nrf2 and its downstream genes such as NQO1, UGT1A1, and GSTM1 are gradually down-regulated during the development of prostate tumor in TRAMP mice [20, 21]. In addition to TRAMP mice, it was also reported that Nrf2 and members of GST mu family were distinctly decreased in human prostate cancer samples, which leads to extensive oxidative stress and DNA damage [22]. Recently, we reported the attenuated expression of Nrf2 may be caused by epigenetic mechanism in prostate cancer of TRAMP mice and tumorigenic TRAMP-C1 cells [23].
Epigenetics is defined as changes in gene expression without alteration of DNA nucleotide sequences [24]. Epigenetic regulation, particularly DNA methylation and histone modification, would change the interaction between gene promoters and transcription factors, resulting in either transcription promotion or repression. It has been reported that cancer progression is usually accompanied with epigenetic silencing of critical tumor suppressor genes through CpG island hypermethylation in their promoter region [25, 26]. For instance in prostate cancer, coordinated hypermethylation of APC and GSTP1 can serve as a specific diagnostic marker in early stages of prostate cancer development [27, 28]. In this context, a series of genes silenced through DNA methylation has been found in TRAMP prostate cancer and its derived cell lines [29, 30]. Drugs which target on those enzymes responsible for epigenetic silencing, such as DNA methyltransferases (DNMTs) and histone deacetylases (HDACs), could be useful in cancer prevention and therapeutic strategy. Treatment of 5-azadeoxycytidine (5-aza, a DNMT inhibitor) and trichostatin A (TSA, an HDAC inhibitor) has been shown to inhibit cell proliferation [31, 32]. It has been reported 5-aza shows inhibitory effect towards prostate tumorigenesis in TRAMP mice in vivo [33]. However, the side effects, such as toxicity and nonspecific gene modulation limited their use as cancer chemopreventive/therapeutic agents [34].
Many compounds from our daily consumption of fruits and vegetables have been shown to exhibit cancer preventive effects through epigenetic mechanisms [35-37]. It has been reported that epigallocatechin-3-gallate (EGCG) from green tea inhibits DNMTs [38] and histone acetyltransferases (HATs) [39]. We have found that curcumin, a bioactive dietary component from turmeric, inhibits DNMTs activity potentially resulting in re-expression of Nrf2 in TRAMP C1 cell line [40]. Sulforaphane (SFN), one of the most widely investigated isothiocyanates found in crucifers, has been shown to be a very potent cancer chemopreventive agent in numerous animal models of different cancers [41]. First identified as a potent phase 2 detoxifying enzyme inducer, SFN has been shown to be a multi-targeted chemopreventive agent including inhibition of histone deacetylase (HDAC) activity [42], suggesting epigenetic alteration may be involved in the mechanism of chemoprevention by SFN. The aim of this study is to investigate the potential of SFN to reactivate the expression of Nrf2 through epigenetic regulation.
2. Materials and Methods
2.1 Cell culture and treatment
TRAMP C1 cells (generously provided by Dr. Barbara Foster, Department of Pharmacology and Therapeutics, Roswell Park Cancer Institute, Buffalo, NY) were maintained in DMEM with 10% fetal bovine serum at 37°C in a humidified 5% CO2 atmosphere. Sulforaphane (SFN) was purchased from LKT Laboratories, Inc. (St. Paul, MN, USA). Other chemicals, 5-azadeoxycytidine (5-aza) and trichostain A (TSA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Cells were seeded in 10 cm plates for 24 h and then treated with either 0.1% DMSO, 500 nM 5-aza or different concentrations of SFN in DMEM medium containing 1% FBS. The medium was changed every 2 days. On day 4, for the 5-aza and TSA combination treatment, 100 nM TSA was added to the 5-aza containing medium. Cells were harvested on day 5 for DNA, protein or total RNA extraction.
2.2 DNA extraction and bisulfite genomic sequencing
Genomic DNA was isolated from DMSO, SFN, or 5-aza/TSA treated TRAMP C1 cells using the QIAamp® DNA mini Kit (Qiagen, Valencia, CA). Then 500 ng genomic DNA was subjected to bisulfite conversion using EZ DNA Methylation Gold Kits (Zymo Research Corp., Orange, CA) following the manufacturer's instructions. The converted DNA was amplified by PCR using Platinum Taq DNA polymerase (Invitrogen, Grand Island, NY) using primers that amplify the first 5 CpGs located between -1085 and -1226 of murine Nrf2 gene with the translation start site defined as +1. PCR products were cloned into pCR4 TOPO vector using a TOPO™ TA Cloning Kit (Invitrogen, Carlsbad, CA). Plasmids from at least ten colonies of each treatment group were prepared using QIAprep Spin Miniprep Kit (Qiagen, Valencia, CA) and sequenced (Genwiz, Piscataway, NJ)
2.3 Methylation DNA immunoprecipitation (MeDIP) analysis
The MeDIP analysis was carried out using MagMeDIP Kit (Diagenode, Denville, NJ) according to the manufacturer's instruction with some modifications. Briefly, 5 μg DNA extracted from treated cells was adjusted to 50 μL in TE buffer and then sonicated in ice-cold water using a Bioruptor sonicator (Diagenode Inc., Sparta, NJ) to shear the DNA to an average size of 300-500 base pairs (bp). The fragmented DNA was denaturated at 95°C for 3 min and followed by immunoprecipitation with anti-methylcytosine antibody at 4°C overnight. After incubation, the pulled-down DNA on magnetic beads were washed and digested with proteinase K and then isolated from beads. The primer set, sense 5′-TGA GAT ATT TTG CAC ATC CGA TA-3′ and anti-sense 5′-ACT CTC AGG GTT CCT TTA CAC G-3′, which covers the DNA sequence of the first 5 CpGs of murine Nrf2 was used for regular PCR and qPCR assays. For regular PCR, 1 μL of each MeDIP and input DNA was used as template for 35 cycles PCR amplification using Platinum PCR SuperMix kit (Invitrogen, Carlsbad, CA). The PCR products were then analyzed by agarose gel electrophoresis and visualized by ethidium bromide staining using a Gel Documentation 2000 system (Bio-Rad, Hercules, CA). For qPCR, the enrichment of MeDIP DNA was calculated according to the calibration of a series of dilution of input DNA, and the relative methylated DNA ratios were then calculated based on the control as 100% of methylated DNA.
2.4 RNA isolation and quantative real-time polymerase chain reaction (qPCR)
Total RNA was extracted from the treated cells using RNeasy Mini Kit (QIAGEN, Valencia, CA). Then first-strand cDNA was synthesized from 1 μg total RNA using SuperScript III First-Strand Synthesis System for RT-PCR (Invitrogen, Grand Island, NY) according to the manufacturer's instructions. To determine the RNA expression of specific genes, the cDNA was used as the template for real time PCR using Power SYBR Green PCR Master Mix (Applied Biosystem, Carlsbads, CA), while β-actin was used as an internal loading control. The sequence of the primers used for real time PCR were as follows: Nrf2: sense 5′-AGC AGG ACT GGA GAA GTT-3′ and anti-sense 5′-TTC TTT TTC CAG CGA GGA GA-3′; NQO1: sense 5′-AGC CCA GAT ATT GTG GCC G-3′ and anti-sense 5′-CCT TTC AGA ATG GCT GGC AC-3′; β-actin: sense 5′-CGT TCA ATA CCC CAG CCA TG-3′ and anti-sense 5′-GAC CCC GTC ACC AGA GTC C-3′.
2.5 Protein lyses preparation and Western blotting
Treated cells from above were harvested using RIPA buffer supplemented with protein inhibitor cocktail (Sigma, St. Louis, MO). Protein concentrations of the cleared lysates were determined using the bicinchoninic acid (BCA) method (Pierce, Rockford, IL). Total proteins (20 μg) from each sample were separated by 4-15% SDS-polyacrylamide gel electrophoresis (Bio-Rad, Hercules, CA). Then the proteins were transferred to a polyvinylidene difluoride (PVDF) membrane (Millipore, Bedford, MA) followed by blocking with 5% BSA in Tris-buffered saline-0.1% Tween 20 (TBST) buffer. Then the membrane was sequentially incubated with specific primary antibodies and HRP-conjugated secondary antibodies. The blots were visualized by SuperSignal enhanced chemiluminiscence (ECL) detection system and recorded using a Gel Documentation 2000 system (Bio-Rad, Hercules, CA). Primary antibodies were purchased from different resources: anti-Nrf2, anti-NQO1 and anti-β-actin from Santa Cruz Biotechnology (Santa Cruz, CA); anti-DNMT1 and anti-DNMT3a from IMGENEX (San Diego, CA); anti-HDAC1, anti-HDAC2, anti-HDAC5, and anti-HDAC7 from Cell Signaling (Boston, MA).
2.6 Chromatin immunoprecipitation (ChIP) assay
Chromatin immunoprecipitation (ChIP) assay was performed using MAGnify Chromatin Immunoprecipitation System (Invitrogen, Grand Island, NY) according to the manufacturer's protocol. Briefly, formaldehyde at a final concentration of 1% was added to fix the cells (∼5.0×106 cells in 10-cm dish). After incubation at room temperature for 10 min, excess formaldehyde was quenched by 5 M glycine. After washing twice with PBS, cells were scraped, pelleted and then resuspended in Cell Lysis Buffer containing protease inhibitor cocktail. The samples were sonicated in ice cold water using a Bioruptor sonicator (Diagenode Inc., Sparta, NJ) to shear the cross-linked DNA to an average length of 200-500 bp and centrifuged at 12,000 rpm to remove insoluble material. The chromatin solutions were then diluted using dilution buffer, and 10 μL of each was reserved as input control. Diluted chromatin solutions were then incubated overnight at 4°C with protein A magnetic beads and specific antibody against anti-acetyl-Histone 3 (Ac-H3) (Millipore, Lake Placid, NY) or nonspecific Rabbit IgG The immunoprecipitated complex-magnetic beads were collected using magnetic separator. After reverse cross-linking, DNA was purified according to manufacturer's instruction. Each purified DNA (1 μL) was used as template for regular PCR amplification using the same primers as those used for MeDIP analysis described above. The PCR products were then analyzed by agarose gel electrophoresis and visualized using EtBr staining. In parallel, qPCR assay was also performed to determine the amount of ChIP-purified DNA using Power SYBR Green PCR Master Mix (Applied Biosystem, Carlsbads, CA).
2.7 Statistical analysis
Data are represented as the mean ± SD from three independent experiments at least. Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by Student's t-test to determine statistically significant difference among the means (P < 0.05).
3. Results
3.1 SFN decreases methylated CpG ratio in the promoter region of Nrf2 gene in TRAMP C1 cells
Our previous study showed that Nrf2 transscription was significantly suppressed when the first five CpGs in the promoter of Nrf2 gene were hypermethylated [23]. Therefore, bisulfite sequencing was carried out to investigate if SFN treatment would demethylate the five CpGs of Nrf2. Fig. 1 shows that the 5 CpGs were hyperrmethylated in the control group (methylation ratio 89.3%). Treatment with SFN (1.0 and 2.5 μM) and the combination of 5-aza (500 nM)/TSA (100 nM) after 5 days incubation reduced the methylation level to 56.0%, 38.7% and 46.7%, respectively. It suggested that SFN has demethylation potential on the promoter of Nef2 gene.
Fig. 1.
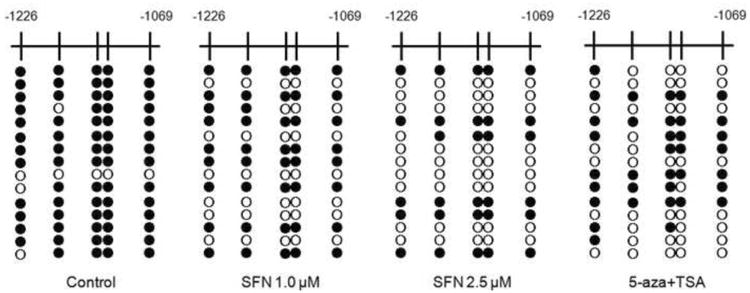
Effect of SFN on methylation status of the Nrf2 promoter regions in TRAMP C1 cells. Cells were treated with 1.0 or 2.5 μM SFN for 5 days, and then genomic DNA was extracted from the treated cells. A combination treatment of 5-aza (500 nM)/TSA (100 nM) was used as positive control, while TSA was added 24 h before cell collection. The methylation pattern of the first five CpGs located at -1226 to -1085 in Nrf2 promoter region, where the translational starting site is considered as +1 was determined. Bisulfite genomic sequencing was performed as described in Materials and Methods. Black dots indicated methylated CpGs while open circles indicate unmethylated CpGs. At least ten clones were picked randomly and sequenced from each of the three independent experiments, and fifteen clones were selected based on the relative frequency of methylation from each treatment.
3.2 SFN decreases the binding of anti-methyl cytosine antibody to the promoter region of Nrf2 gene in TRAMP C1 cells
MeDIP analysis was performed using anti-methyl cytosine antibody by immunoprecipitation and the enriched methylated DNA was then used as template for PCR amplification of the Nrf2 promoter region containing the first 5 CpGs (Fig. 2). Both regular PCR (Fig. 2A) and qPCR (Fig. 2B) results show that SFN and the combination of 5-aza/TSA treatment significantly reduced the binding of the enriched Nrf2 promoter containing the first 5 CpGs to anti-methyl cytosine antibody (Fig. 2).
Fig. 2.
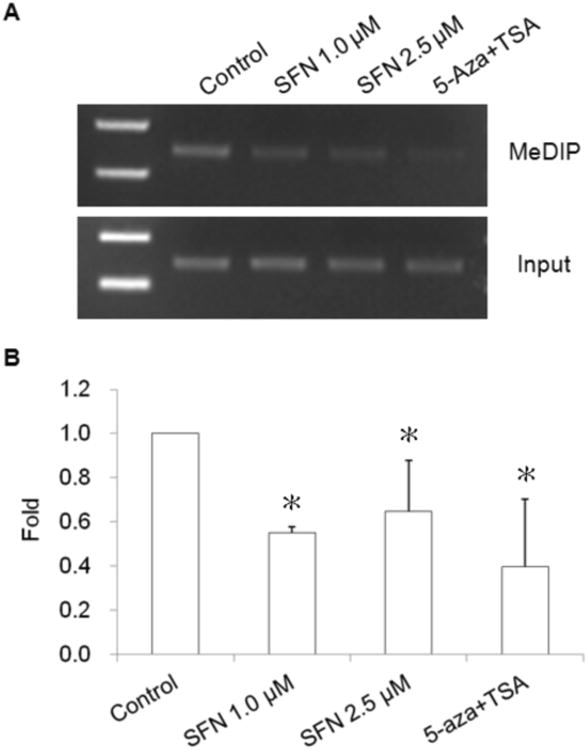
Effect of SFN on methylation of Nrf2 promoter region in TRAMP C1 cells using methylation DNA immunopredipitation (MeDIP) assay. Genomic DNA (10 μg) extracted from SFN or 5-aza/TSA treated TRAMP C1 cells were used for MeDIP analysis. Genomic DNA were sonicated, denatured and subjected to DNA immuniprecipitation (IP) with anti-methyl cytosine antibody. Then regular PCR (A) and qPCR (B) were performed to analyze the enrichment of methylated fragments using the primers covering the DNA sequence containing the first five CpGs in promoter region of the Nrf2 gene. (A) Regular PCR was performed to compare the immunoprecipitated DNA with their inputs, a representative result is shown from three independent experiments. (B) The enrichment of the MeDIP DNA was determined by qPCR on the basis of the standard curve from a serial dilution of the inputs. Relative methylated ratio was calculated by comparing with control group as 100% methylated DNA. Data are expressed as mean ± SD from four independent experiments. * Different from control, P < 0.05.
3.3 SFN induces expression of Nrf2 and its downstream gene
Previously the expression of Nrf2 and its downstream genes has been shown to be dramatically decreased in TRAMP prostate cancer and tumorigeneric TRAMP C1 cell line [20, 23]. To examine if demethylation of the promoter region results in the transcription activation of Nrf2, the mRNA and protein expression levels of Nrf2 and its targeted gene NQO1 were measured. TRAMP C1 cells were treated with SFN for 5 days, and then qPCR was performed to evaluate the mRNA expression of Nrf2 and NQO1. The results indicated that both 1.0 and 2.5 μM SFN treatments increased Nrf2 and NQO1 mRNA expressions (Fig. 3A). Consistently, protein expression of Nrf2 and NOQ1 was also significantly induced upon SFN treatment in TRAMP C1 cells (Fig. 3B).
Fig. 3.
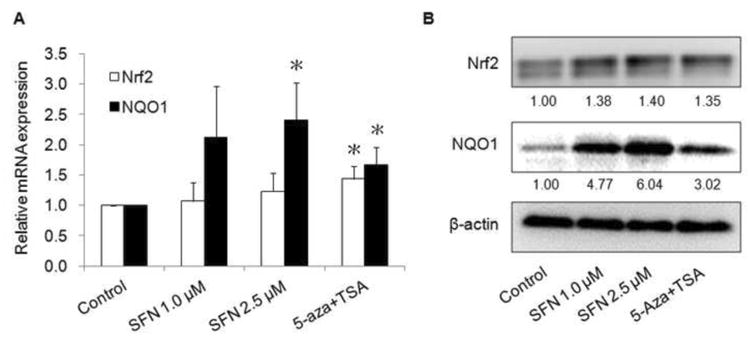
Effect of SFN on the mRNA (A) and protein (B) expressions of Nrf2 and NQO-1. After 5 days of treatment, mRNA and protein were isolated from TRAMP C1 cells as described above. The mRNA and protein levels of Nrf2 and NQO-1 were determined using qPCR and Western blotting, respectively. (A) The mRNA level was determined using ABI7900HT qPCR system. Data are expressed as mean ± SD from three independent experiments. * Different from control, P < 0.05. (B) For protein analysis, the bands which were bound with specific Nrf2 and NQO1 antibodies were visualized using a Gel Documentation 2000 system (Bio-Rad, Hercules, CA) and quantified using Quantity One software. β-Actin was used as an equal loading control.
3.4 SFN alters expressions of epigenetics modifying enzymes
To further study the epigenetics mechanism underlying the demethylation effect of SFN on the Nrf2 promoter region, we next examined if SFN can alter the expression of those epigenetics modifying enzymes. DNA methylation is mediated by a family of DNA methyltransferases (DNMTs) which include a maintenance methyltransferase DNMT1 and two de novo methyltransferases DNMT3a and DNMT3b. As shown in Fig. 4A, SFN decreased the protein levels of DNMT1 and DNMT3a in a dose-dependent manner in TRAMP C1 cells. However, no significant alteration was observed with the expression of DNMT3b (data not shown).
Fig. 4.
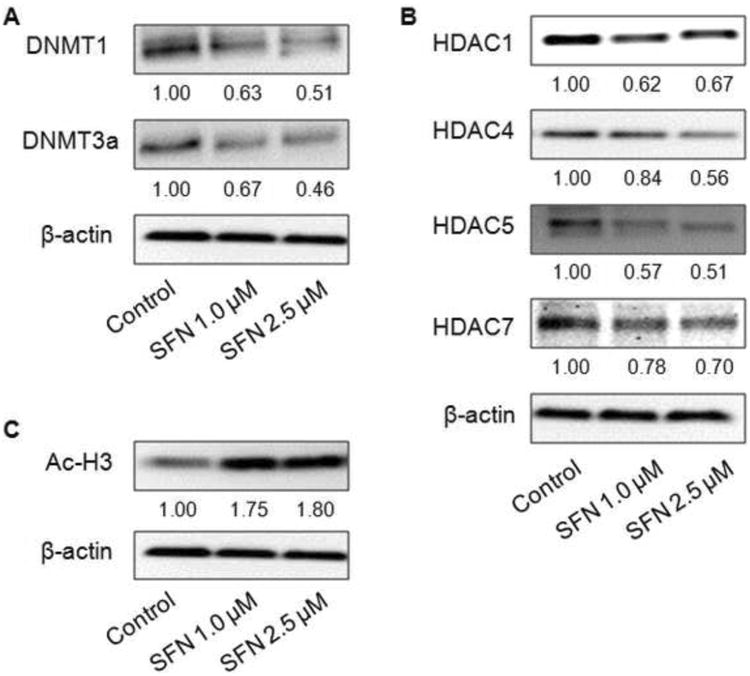
Effect of SFN on the protein expressions of DNMTs (A), HDACs (B), and acetylated histone 3 (Ac-H3) (C) in TRAMP C1 cells. Cell lysates were prepared after SFN with the indicated doses for 5 days, followed by Western blotting to analyze protein expressions of DNMTs, HDACs, and Ac-H3 using specific antibodies. β-Actin was used as an equal loading control.
In addition, SFN has been reported to be an HDAC inhibitor. Correspondingly, we found that SFN significantly decreased the protein levels of HDAC1, HDAC4, HDAC5, and HDAC7 in a dose-dependent manner (Fig. 4B) while no considerable changes of HDAC2, HDAC3, and HDAC8 protein expressions (data not shown). The global level of acetylated histone 3 (Ac-H3) was also highly induced by SFN treatment (Fig. 4C). Next, chromatin immunoprecipitation (ChIP) assays were performed to further examine the association between Nrf2 gene promoter and Ac-H3 level after SFN treatments in TRAMP C1 cells. In agreement with the observation of Ac-H3 level, the amount of immunoprecipitated DNA of Nrf2 promoter containing the first 5 CpGs fragment in SFN treatments (1.0 and 2.5 μM) was higher as compared to control treatments as shown in Fig. 5.
Fig. 5.
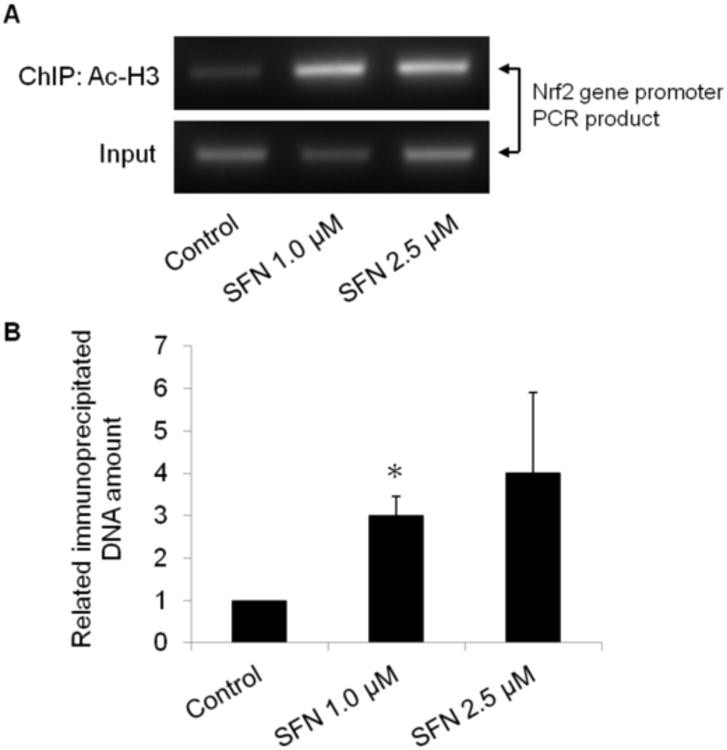
Effect of SFN on the binding of Nrf2 promoter region to acetylated histone 3 (Ac-H3). ChIP assay was performed as described in Material & Methods using antibody against Ac-H3. Regular PCR (A) and qPCR (B) were performed to analyze the enrichment of DNA binding to Ac-H3 using primers which amplify the DNA sequence containing the first five CpGs in the promoter region of Nrf2 gene. (A) Regular PCR was performed to compare the immunoprecipitated DNA versus their inputs of each treatment, and a representative result is shown from three independent experiments. (B) The enrichment of the Ac-H3 binding DNA was determined by qPCR based on the standard curve which was obtained from a serial dilution of the inputs. Relative ratio was calculated by the fold change compared to control group. Data are expressed as mean ± SD from three independent experiments. * Different from control, P < 0.05.
4. Discussion
The imbalance between ROS and cellular antioxidant capacity could result in oxidative stress in organisms. Oxidative stress causes biochemical alterations of proteins, nucleic acids, and lipids in cells, which may further lead to inflammation or carcinogenesis [43]. Nrf2, a transcription factor, plays an important role in regulating phase II antioxidant and detoxification enzymes which eliminate carcinogens and reactive intermediates and hence protect cells from oxidative stress induced damages [44-47]. In recent studies of prostate cancer, low expressions of Nrf2 and its downstream target genes such as GST family are found in human [22] and in TRAMP prostate tumors [20]. Accumulating evidence also shows that dietary phytochemicals can enhance the expressions of phase II antioxidant and detoxification enzymes mediated by the Nrf2 pathway [48]. For example, ! -rich tocopherols can inhibit prostate carcinogenesis in TRAMP mice while it increases the expressions of Nrf2 and its target genes including UGT, GST, HO-1, and glutathione peroxidases (GPx) [20, 21]. In addition ! -rich tocopherols could cause CpG demethylation in the Nrf2 promoter and higher Nrf2 expression in the prostate of TRAMP mice and in TRAMP C1 cells, potentially contributing to the prevention of prostate tumorigenesis [49]. In addition, curcumin has been demonstrated to restore the epigenetically silenced Nrf2 gene through DNA demethylation in TRAMP C1 cells [40]. Likewise, it has also been shown to possess cancer chemopreventive activity potentially through enhancing the expressions of Nrf2 and its downstream target genes [50, 51]. Furthermore, it has been demonstrated that broccoli sprouts containing abundant SFN can inhibit prostate tumorigenesis in TRAMP mice, which may be through the activation of Nrf2 pathway [52]. Activation of Nrf2 pathway by SFN plays a role in chemoprevention in several animal models such as benzo[a]pyrene-induced forestomach tumors using wild-type and Nrf2-deficient ICR mice [53] and DMBA/TPA-induced skin carcinogenesis with wild-type and Nrf2-deficient C57BL/6 Mice [54]. In our current study, SFN restores the expression of Nrf2 and its target gene NQO1 (Fig. 3), via demethylating the CpGs of the promoter of Nrf2 gene in TRAMP C1 cells, may play a role in chemoprevention in TRAMP mouse model.
Epigenetic regulation plays a critical process in cancer development, which includes DNA methylation and histone modification. Cellular DNA methyltransferases (DNMTs) and histone deacetylases (HDACs) which cause epigenetic silencing are the major targets of cancer prevention and therapeutic strategy. However, some anti-cancer drugs including 5-azacitdine and 5-aza-2′-deoxycytidine (DNMTs inhibitors) and vorinostat, belinostat, romidepsin, and panobinostat (HDAC inhibitors) may have adverse side effects [55-57]. Many studies have demonstrated that some dietary phytochemicals, such as epigallocatechin-3-gallate (EGCG) [39], curcumin [40], and genistein [58], with cancer chemopreventive and/or anti-cancer activities have epigenetic regulation potentials. The epigenetic effect of SFN on gene transcription has also been recently reported. SFN may cause site-specific CpG demethylation of the first exon of hTERT gene through the suppression of DNMT1 and DNMT3a in a dose- and time-dependent manner in human breast cancer cells [59]. In our present study, we also found that SFN treatment exhibited hypomethylation effect on the first 5 CpGs in the Nrf2 promoter in TRAMP C1 cells (Fig. 1). This hypomethylation effect is supported by MeDIP assay showing that SFN significantly decreased the amount of the Nrf2 promoter containing the first 5 CpGs using anti-mecyt antibody (Fig. 2). These results might be associated with the suppression of the protein expression of DNMT1 and DNMT3a (Fig. 4A). With respect to HDACs, it has been reported that SFN as well as its two major metabolites, SFN-cysteine and SFN-N-acetylcysteine, can suppress HDAC activity in vitro [60]. SFN also shows a similar HDAC inhibitory effect in BPH-1, LnCaP and PC-3 prostate epithelial cells [61]. It has been reported that the levels of the active chromatin markers (acetyl-H3, acetyl-H3K9 and acetyl-H4) are increased, but the levels of the inactive chromatin markers (trimethyl-H3K9 and trimethyl-H3K27) are decreased in MCF-7 and MDA-MB-231 cells after SFN treatment [59]. In in vivo study, the suppressive effect of SFN against the growth of PC-3 xenografts might be mediated by inhibiting HDAC activity [62]. SFN can also retard intestinal carcinogenesis in Apc-minus mice through histone modification accompanied with inhibition of HDAC activity in the colonic mucosa and polyps [63]. In addition, HDAC activity in peripheral blood mononuclear cells (PBMC) was inhibited in the human subjects after 3 and 6 hrs following consumption of SFN-rich broccoli sprouts [62]. Our present results show that the protein expression levels of HDAC1, HDAC4, HDAC5, and HDAC7 were significantly decreased in a dose-dependent manner while the Ac-H3 protein level was highly induced by SFN (Fig. 4B and C). Furthermore, we also demonstrated that SFN treatment increased the binding of Ac-H3, one of the transcriptionally active chromatin markers, to the promoter of Nrf2 gene (Fig. 5). It has also been showed that dietary bioactive compounds such as SFN may interfere with the formation of a transcriptional regulator complex which consists of methyl-CpG binding proteins, DNMT and HDAC [64, 65]. The release of co-repressor complex further results in the concomitant enrichment of acetyl histones such as Ac-H3 [65-67]. Therefore, our current findings are in agreement with previous investigations and support the evidence of epigenetic effect of SFN in restoring Nrf2 expression, including DNA demethylation and chromatin remodeling process.
SFN has been shown to inhibit prostate carcinogenesis in TRAMP mice [68]. Gradual down-regulation of Nrf2 and Nrf2 downstream target genes during the progression of prostate tumorigenesis in TRAMP mice has been reported [20], and SFN-rich broccoli sprouts increases the expression levels of Nrf2 and its downstream target genes and inhibits tumorigenesis [52]. In our current study, SFN is an epigenetic regulator and restores the silenced Nrf2 gene in TRAMP C1 cells through DNA demethylation and histone modifications. These finding suggest that SFN epigenetically restore the expression of Nrf2 and Nrf2 downstream antioxidant and detoxification related genes during the initiation, progression and development of prostate cancer in TRAMP mice may play a role in SFN's chemoprevention effects.
Acknowledgments
We thank all the members in Dr. Ah-Ng Tony Kong's lab for their helpful discussion and preparation of this manuscript.
Footnotes
Conflict of interest No potential conflicts of interest were disclosed.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Siegel R, Ward E, Brawley O, Jemal A. Cancer statistics, 2011: the impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA: a cancer journal for clinicians. 2011;61:212–36. doi: 10.3322/caac.20121. [DOI] [PubMed] [Google Scholar]
- 2.Kumar B, Koul S, Khandrika L, Meacham RB, Koul HK. Oxidative stress is inherent in prostate cancer cells and is required for aggressive phenotype. Cancer research. 2008;68:1777–85. doi: 10.1158/0008-5472.CAN-07-5259. [DOI] [PubMed] [Google Scholar]
- 3.Minelli A, Bellezza I, Conte C, Culig Z. Oxidative stress-related aging: A role for prostate cancer? Biochimica et biophysica acta. 2009;1795:83–91. doi: 10.1016/j.bbcan.2008.11.001. [DOI] [PubMed] [Google Scholar]
- 4.De Marzo AM, Meeker AK, Zha S, Luo J, Nakayama M, Platz EA, et al. Human prostate cancer precursors and pathobiology. Urology. 2003;62:55–62. doi: 10.1016/j.urology.2003.09.053. [DOI] [PubMed] [Google Scholar]
- 5.Yu BP. Cellular defenses against damage from reactive oxygen species. Physiological reviews. 1994;74:139–62. doi: 10.1152/physrev.1994.74.1.139. [DOI] [PubMed] [Google Scholar]
- 6.Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet. 2001;357:539–45. doi: 10.1016/S0140-6736(00)04046-0. [DOI] [PubMed] [Google Scholar]
- 7.De Marzo AM, Platz EA, Sutcliffe S, Xu J, Gronberg H, Drake CG, et al. Inflammation in prostate carcinogenesis. Nature reviews Cancer. 2007;7:256–69. doi: 10.1038/nrc2090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Federico A, Morgillo F, Tuccillo C, Ciardiello F, Loguercio C. Chronic inflammation and oxidative stress in human carcinogenesis. International journal of cancer Journal international du cancer. 2007;121:2381–6. doi: 10.1002/ijc.23192. [DOI] [PubMed] [Google Scholar]
- 9.Nguyen T, Sherratt PJ, Pickett CB. Regulatory mechanisms controlling gene expression mediated by the antioxidant response element. Annual review of pharmacology and toxicology. 2003;43:233–60. doi: 10.1146/annurev.pharmtox.43.100901.140229. [DOI] [PubMed] [Google Scholar]
- 10.Motohashi H, Yamamoto M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends in molecular medicine. 2004;10:549–57. doi: 10.1016/j.molmed.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 11.Kobayashi A, Ohta T, Yamamoto M. Unique function of the Nrf2-Keap1 pathway in the inducible expression of antioxidant and detoxifying enzymes. Methods in enzymology. 2004;378:273–86. doi: 10.1016/S0076-6879(04)78021-0. [DOI] [PubMed] [Google Scholar]
- 12.Chen C, Kong AN. Dietary chemopreventive compounds and ARE/EpRE signaling. Free radical biology & medicine. 2004;36:1505–16. doi: 10.1016/j.freeradbiomed.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 13.Kwak MK, Wakabayashi N, Kensler TW. Chemoprevention through the Keap1-Nrf2 signaling pathway by phase 2 enzyme inducers. Mutation research. 2004;555:133–48. doi: 10.1016/j.mrfmmm.2004.06.041. [DOI] [PubMed] [Google Scholar]
- 14.Kang KW, Lee SJ, Kim SG. Molecular mechanism of nrf2 activation by oxidative stress. Antioxidants & redox signaling. 2005;7:1664–73. doi: 10.1089/ars.2005.7.1664. [DOI] [PubMed] [Google Scholar]
- 15.Nguyen T, Nioi P, Pickett CB. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. The Journal of biological chemistry. 2009;284:13291–5. doi: 10.1074/jbc.R900010200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Li Y, Jaiswal AK. Regulation of human NAD(P)H:quinone oxidoreductase gene. Role of AP1 binding site contained within human antioxidant response element. The Journal of biological chemistry. 1992;267:15097–104. [PubMed] [Google Scholar]
- 17.Friling RS, Bergelson S, Daniel V. Two adjacent AP-1-like binding sites form the electrophile-responsive element of the murine glutathione S-transferase Ya subunit gene. Proceedings of the National Academy of Sciences of the United States of America. 1992;89:668–72. doi: 10.1073/pnas.89.2.668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ramos-Gomez M, Kwak MK, Dolan PM, Itoh K, Yamamoto M, Talalay P, et al. Sensitivity to carcinogenesis is increased and chemoprotective efficacy of enzyme inducers is lost in nrf2 transcription factor-deficient mice. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:3410–5. doi: 10.1073/pnas.051618798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Khor TO, Huang MT, Prawan A, Liu Y, Hao X, Yu S, et al. Increased susceptibility of Nrf2 knockout mice to colitis-associated colorectal cancer. Cancer Prev Res (Phila) 2008;1:187–91. doi: 10.1158/1940-6207.CAPR-08-0028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Barve A, Khor TO, Nair S, Reuhl K, Suh N, Reddy B, et al. Gamma-tocopherol-enriched mixed tocopherol diet inhibits prostate carcinogenesis in TRAMP mice. International journal of cancer Journal international du cancer. 2009;124:1693–9. doi: 10.1002/ijc.24106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Barve A, Khor TO, Reuhl K, Reddy B, Newmark H, Kong AN. Mixed tocotrienols inhibit prostate carcinogenesis in TRAMP mice. Nutrition and cancer. 2010;62:789–94. doi: 10.1080/01635581003605896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Frohlich DA, McCabe MT, Arnold RS, Day ML. The role of Nrf2 in increased reactive oxygen species and DNA damage in prostate tumorigenesis. Oncogene. 2008;27:4353–62. doi: 10.1038/onc.2008.79. [DOI] [PubMed] [Google Scholar]
- 23.Yu S, Khor TO, Cheung KL, Li W, Wu TY, Huang Y, et al. Nrf2 expression is regulated by epigenetic mechanisms in prostate cancer of TRAMP mice. PloS one. 2010;5:e8579. doi: 10.1371/journal.pone.0008579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Devinoy E, Rijnkels M. Epigenetics in mammary gland biology and cancer. Journal of mammary gland biology and neoplasia. 2010;15:1–4. doi: 10.1007/s10911-010-9171-3. [DOI] [PubMed] [Google Scholar]
- 25.Baylin SB. DNA methylation and gene silencing in cancer. Nature clinical practice Oncology. 2005;2(Suppl 1):S4–11. doi: 10.1038/ncponc0354. [DOI] [PubMed] [Google Scholar]
- 26.Lopez J, Percharde M, Coley HM, Webb A, Crook T. The context and potential of epigenetics in oncology. British journal of cancer. 2009;100:571–7. doi: 10.1038/sj.bjc.6604930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nakayama M, Gonzalgo ML, Yegnasubramanian S, Lin X, De Marzo AM, Nelson WG. GSTP1 CpG island hypermethylation as a molecular biomarker for prostate cancer. Journal of cellular biochemistry. 2004;91:540–52. doi: 10.1002/jcb.10740. [DOI] [PubMed] [Google Scholar]
- 28.Jeronimo C, Henrique R, Hoque MO, Mambo E, Ribeiro FR, Varzim G, et al. A quantitative promoter methylation profile of prostate cancer. Clinical cancer research : an official journal of the American Association for Cancer Research. 2004;10:8472–8. doi: 10.1158/1078-0432.CCR-04-0894. [DOI] [PubMed] [Google Scholar]
- 29.Morey SR, Smiraglia DJ, James SR, Yu J, Moser MT, Foster BA, et al. DNA methylation pathway alterations in an autochthonous murine model of prostate cancer. Cancer research. 2006;66:11659–67. doi: 10.1158/0008-5472.CAN-06-1937. [DOI] [PubMed] [Google Scholar]
- 30.Pulukuri SM, Rao JS. CpG island promoter methylation and silencing of 14-3-3sigma gene expression in LNCaP and Tramp-C1 prostate cancer cell lines is associated with methyl-CpG-binding protein MBD2. Oncogene. 2006;25:4559–72. doi: 10.1038/sj.onc.1209462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Walton TJ, Li G, Seth R, McArdle SE, Bishop MC, Rees RC. DNA demethylation and histone deacetylation inhibition co-operate to re-express estrogen receptor beta and induce apoptosis in prostate cancer cell-lines. The Prostate. 2008;68:210–22. doi: 10.1002/pros.20673. [DOI] [PubMed] [Google Scholar]
- 32.Kim TY, Bang YJ, Robertson KD. Histone deacetylase inhibitors for cancer therapy. Epigenetics : official journal of the DNA Methylation Society. 2006;1:14–23. doi: 10.4161/epi.1.1.2644. [DOI] [PubMed] [Google Scholar]
- 33.McCabe MT, Low JA, Daignault S, Imperiale MJ, Wojno KJ, Day ML. Inhibition of DNA methyltransferase activity prevents tumorigenesis in a mouse model of prostate cancer. Cancer research. 2006;66:385–92. doi: 10.1158/0008-5472.CAN-05-2020. [DOI] [PubMed] [Google Scholar]
- 34.Miyamoto K, Ushijima T. Diagnostic and therapeutic applications of epigenetics. Japanese journal of clinical oncology. 2005;35:293–301. doi: 10.1093/jjco/hyi088. [DOI] [PubMed] [Google Scholar]
- 35.Suzuki T, Miyata N. Epigenetic control using natural products and synthetic molecules. Current medicinal chemistry. 2006;13:935–58. doi: 10.2174/092986706776361067. [DOI] [PubMed] [Google Scholar]
- 36.Hauser AT, Jung M. Targeting epigenetic mechanisms: potential of natural products in cancer chemoprevention. Planta medica. 2008;74:1593–601. doi: 10.1055/s-2008-1081347. [DOI] [PubMed] [Google Scholar]
- 37.Meeran SM, Ahmed A, Tollefsbol TO. Epigenetic targets of bioactive dietary components for cancer prevention and therapy. Clinical epigenetics. 2010;1:101–16. doi: 10.1007/s13148-010-0011-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fang MZ, Wang Y, Ai N, Hou Z, Sun Y, Lu H, et al. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits DNA methyltransferase and reactivates methylation-silenced genes in cancer cell lines. Cancer research. 2003;63:7563–70. [PubMed] [Google Scholar]
- 39.Pandey M, Shukla S, Gupta S. Promoter demethylation and chromatin remodeling by green tea polyphenols leads to re-expression of GSTP1 in human prostate cancer cells. International journal of cancer Journal international du cancer. 2010;126:2520–33. doi: 10.1002/ijc.24988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Khor TO, Huang Y, Wu TY, Shu L, Lee J, Kong AN. Pharmacodynamics of curcumin as DNA hypomethylation agent in restoring the expression of Nrf2 via promoter CpGs demethylation. Biochemical pharmacology. 2011;82:1073–8. doi: 10.1016/j.bcp.2011.07.065. [DOI] [PubMed] [Google Scholar]
- 41.Cheung KL, Kong AN. Molecular targets of dietary phenethyl isothiocyanate and sulforaphane for cancer chemoprevention. The AAPS journal. 2010;12:87–97. doi: 10.1208/s12248-009-9162-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ho E, Clarke JD, Dashwood RH. Dietary sulforaphane, a histone deacetylase inhibitor for cancer prevention. The Journal of nutrition. 2009;139:2393–6. doi: 10.3945/jn.109.113332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Valko M, Leibfritz D, Moncol J, Cronin MT, Mazur M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;39:44–84. doi: 10.1016/j.biocel.2006.07.001. [DOI] [PubMed] [Google Scholar]
- 44.Chen C, Kong AN. Dietary chemopreventive compounds and ARE/EpRE signaling. Free Radic Biol Med. 2004;36:1505–16. doi: 10.1016/j.freeradbiomed.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 45.Kundu JK, Surh YJ. Nrf2-Keap1 signaling as a potential target for chemoprevention of inflammation-associated carcinogenesis. Pharm Res. 2010;27:999–1013. doi: 10.1007/s11095-010-0096-8. [DOI] [PubMed] [Google Scholar]
- 46.Li W, Kong AN. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol Carcinog. 2009;48:91–104. doi: 10.1002/mc.20465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang DD. Mechanistic studies of the Nrf2-Keap1 signaling pathway. Drug metabolism reviews. 2006;38:769–89. doi: 10.1080/03602530600971974. [DOI] [PubMed] [Google Scholar]
- 48.Su ZY, Shu L, Khor TO, Lee JH, Fuentes F, Kong AN. A Perspective on Dietary Phytochemicals and Cancer Chemoprevention: Oxidative Stress, Nrf2, and Epigenomics. Topics in current chemistry. 2012 doi: 10.1007/128_2012_340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Huang Y, Khor TO, Shu L, Saw CL, Wu TY, Suh N, et al. A gamma-tocopherol-rich mixture of tocopherols maintains Nrf2 expression in prostate tumors of TRAMP mice via epigenetic inhibition of CpG methylation. The Journal of nutrition. 2012;142:818–23. doi: 10.3945/jn.111.153114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Shen G, Xu C, Hu R, Jain MR, Gopalkrishnan A, Nair S, et al. Modulation of nuclear factor E2-related factor 2-mediated gene expression in mice liver and small intestine by cancer chemopreventive agent curcumin. Molecular cancer therapeutics. 2006;5:39–51. doi: 10.1158/1535-7163.MCT-05-0293. [DOI] [PubMed] [Google Scholar]
- 51.Garg R, Gupta S, Maru GB. Dietary curcumin modulates transcriptional regulators of phase I and phase II enzymes in benzo[a]pyrene-treated mice: mechanism of its anti-initiating action. Carcinogenesis. 2008;29:1022–32. doi: 10.1093/carcin/bgn064. [DOI] [PubMed] [Google Scholar]
- 52.Keum YS, Khor TO, Lin W, Shen G, Kwon KH, Barve A, et al. Pharmacokinetics and pharmacodynamics of broccoli sprouts on the suppression of prostate cancer in transgenic adenocarcinoma of mouse prostate (TRAMP) mice: implication of induction of Nrf2, HO-1 and apoptosis and the suppression of Akt-dependent kinase pathway. Pharmaceutical research. 2009;26:2324–31. doi: 10.1007/s11095-009-9948-5. [DOI] [PubMed] [Google Scholar]
- 53.Fahey JW, Haristoy X, Dolan PM, Kensler TW, Scholtus I, Stephenson KK, et al. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc Natl Acad Sci U S A. 2002;99:7610–5. doi: 10.1073/pnas.112203099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Xu C, Huang MT, Shen G, Yuan X, Lin W, Khor TO, et al. Inhibition of 7,12-dimethylbenz(a)anthracene-induced skin tumorigenesis in C57BL/6 mice by sulforaphane is mediated by nuclear factor E2-related factor 2. Cancer Res. 2006;66:8293–6. doi: 10.1158/0008-5472.CAN-06-0300. [DOI] [PubMed] [Google Scholar]
- 55.Garcia-Manero G, Assouline S, Cortes J, Estrov Z, Kantarjian H, Yang H, et al. Phase 1 study of the oral isotype specific histone deacetylase inhibitor MGCD0103 in leukemia. Blood. 2008;112:981–9. doi: 10.1182/blood-2007-10-115873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rasheed WK, Johnstone RW, Prince HM. Histone deacetylase inhibitors in cancer therapy. Expert opinion on investigational drugs. 2007;16:659–78. doi: 10.1517/13543784.16.5.659. [DOI] [PubMed] [Google Scholar]
- 57.Griffiths EA, Gore SD. DNA methyltransferase and histone deacetylase inhibitors in the treatment of myelodysplastic syndromes. Seminars in hematology. 2008;45:23–30. doi: 10.1053/j.seminhematol.2007.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Majid S, Dar AA, Ahmad AE, Hirata H, Kawakami K, Shahryari V, et al. BTG3 tumor suppressor gene promoter demethylation, histone modification and cell cycle arrest by genistein in renal cancer. Carcinogenesis. 2009;30:662–70. doi: 10.1093/carcin/bgp042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Meeran SM, Patel SN, Tollefsbol TO. Sulforaphane causes epigenetic repression of hTERT expression in human breast cancer cell lines. PLoS One. 2010;5:e11457. doi: 10.1371/journal.pone.0011457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Myzak MC, Karplus PA, Chung FL, Dashwood RH. A novel mechanism of chemoprotection by sulforaphane: inhibition of histone deacetylase. Cancer Res. 2004;64:5767–74. doi: 10.1158/0008-5472.CAN-04-1326. [DOI] [PubMed] [Google Scholar]
- 61.Myzak MC, Hardin K, Wang R, Dashwood RH, Ho E. Sulforaphane inhibits histone deacetylase activity in BPH-1, LnCaP and PC-3 prostate epithelial cells. Carcinogenesis. 2006;27:811–9. doi: 10.1093/carcin/bgi265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Myzak MC, Tong P, Dashwood WM, Dashwood RH, Ho E. Sulforaphane retards the growth of human PC-3 xenografts and inhibits HDAC activity in human subjects. Exp Biol Med (Maywood) 2007;232:227–34. [PMC free article] [PubMed] [Google Scholar]
- 63.Myzak MC, Dashwood WM, Orner GA, Ho E, Dashwood RH. Sulforaphane inhibits histone deacetylase in vivo and suppresses tumorigenesis in Apc-minus mice. FASEB J. 2006;20:506–8. doi: 10.1096/fj.05-4785fje. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Saxena NK, Sharma D. Epigenetic Reactivation of Estrogen Receptor: Promising Tools for Restoring Response to Endocrine Therapy. Molecular and cellular pharmacology. 2010;2:191–202. [PMC free article] [PubMed] [Google Scholar]
- 65.Meeran SM, Patel SN, Li Y, Shukla S, Tollefsbol TO. Bioactive dietary supplements reactivate ER expression in ER-negative breast cancer cells by active chromatin modifications. PloS one. 2012;7:e37748. doi: 10.1371/journal.pone.0037748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Sharma D, Blum J, Yang X, Beaulieu N, Macleod AR, Davidson NE. Release of methyl CpG binding proteins and histone deacetylase 1 from the Estrogen receptor alpha (ER) promoter upon reactivation in ER-negative human breast cancer cells. Molecular endocrinology. 2005;19:1740–51. doi: 10.1210/me.2004-0011. [DOI] [PubMed] [Google Scholar]
- 67.Macaluso M, Cinti C, Russo G, Russo A, Giordano A. pRb2/p130-E2F4/5-HDAC1-SUV39H1-p300 and pRb2/p130-E2F4/5-HDAC1-SUV39H1-DNMT1 multimolecular complexes mediate the transcription of estrogen receptor-alpha in breast cancer. Oncogene. 2003;22:3511–7. doi: 10.1038/sj.onc.1206578. [DOI] [PubMed] [Google Scholar]
- 68.Singh SV, Warin R, Xiao D, Powolny AA, Stan SD, Arlotti JA, et al. Sulforaphane inhibits prostate carcinogenesis and pulmonary metastasis in TRAMP mice in association with increased cytotoxicity of natural killer cells. Cancer Res. 2009;69:2117–25. doi: 10.1158/0008-5472.CAN-08-3502. [DOI] [PMC free article] [PubMed] [Google Scholar]