

Abstract
Sphingosine-1-phosphate (S1P) is a pleiotropic lipid mediator that regulates cell survival, migration, the recruitment of immune cells, angiogenesis, and lymphangiogenesis, all of which are involved in cancer progression. S1P is generated inside cancer cells by sphingosine kinases then exported outside of the cell into the tumor microenvironment where it binds to any of five G protein coupled receptors and proceeds to regulate a variety of functions. We have recently reported on the mechanisms underlying the “inside-out” signaling of S1P, its export through the plasma membrane, and its interaction with cell surface receptors. Membrane lipids, including S1P, do not spontaneously exchange through lipid bilayers since the polar head groups do not readily go through the hydrophobic interior of the plasma membrane. Instead, specific transporter proteins exist on the membrane to exchange these lipids. This review summarizes what is known regarding S1P transport through the cell membrane via ATP-binding cassette transporters and the spinster 2 transporter and discusses the roles for these transporters in cancer and in the tumor microenvironment. Based on our research and the emerging understanding of the role of S1P signaling in cancer and in the tumor microenvironment, S1P transporters and S1P signaling hold promise as new therapeutic targets for cancer drug development.
1. Introduction
It is well recognized that the tumor microenvironment (TME) plays a key role in cancer progression and metastasis [1–3]. Tumors influence the surrounding microenvironment through the release of extracellular signals, such as cytokines, chemokines, and lipid mediators [4–7]. These bioactive molecules secreted from cancer cells and noncancer components in the TME, such as blood vessels, lymphatic vessels, and inflammatory cells, are considered to be potential therapeutic targets.
Sphingosine-1-phosphate (S1P) has emerged as a new player in the TME and cancer progression during the last decade [4]. S1P generated inside cells is exported outside of cells into the TME where it stimulates specific S1P receptors on the cell surface (Figure 1). This “inside-out” signaling of S1P is considered to play a fundamental role in cancer progression [8]. To date, several S1P transporters have been identified [9–13]. Here, we review the “inside-out” signaling of S1P with a focus on S1P transporters. We will discuss the importance of S1P signaling in cancer and the possibility of targeting S1P transporters for cancer treatment.
Figure 1.
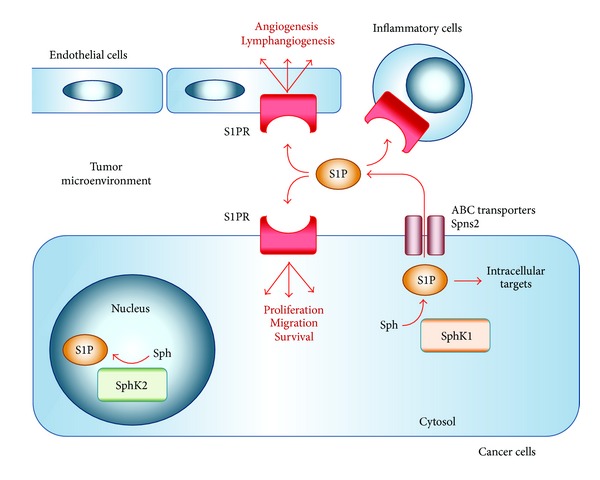
“Inside-out” signaling of sphingosine-1-phosphate (S1P). S1P is generated from Sph (sphingosine) by SphK1 (sphingosine kinase 1) in the cytosol of cancer cells and exported via ABC (ATP-binding cassette) transporters or Spns2 (spinster 2) outside of cells (tumor microenvironment). S1P stimulates specific S1P receptors (S1PR1-5) to promote numerous cellular functions, such as cell proliferation, migration, angiogenesis, and lymphangiogenesis.
2. “Inside-Out” Signaling of S1P
S1P is a pleiotropic lipid mediator that regulates cell survival, proliferation, migration, angiogenesis, lymphangiogenesis, and the recruitment of immune cells (Figure 1), making it involved in many physiological and pathological conditions including immune function, inflammation, and cancer [14–19]. S1P is generated from sphingosine inside the cells by two sphingosine kinases (SphK1 and SphK2) [20–23]. The balance between the levels of S1P and its metabolic precursors of ceramide and sphingosine has been regarded as a rheostat that could determine whether a cell proliferates or dies [24]. S1P generated within cells is exported out from cells via S1P transporters on the cell membrane, which will be described in detail later in this review. S1P outside of the cells can stimulate any of five specific G protein coupled receptors (S1PR1-5) [10, 12], with each S1P receptor coupled to different G proteins, which regulate activation or inhibition of the downstream intracellular signaling involved in various cellular functions (Figure 2).
Figure 2.
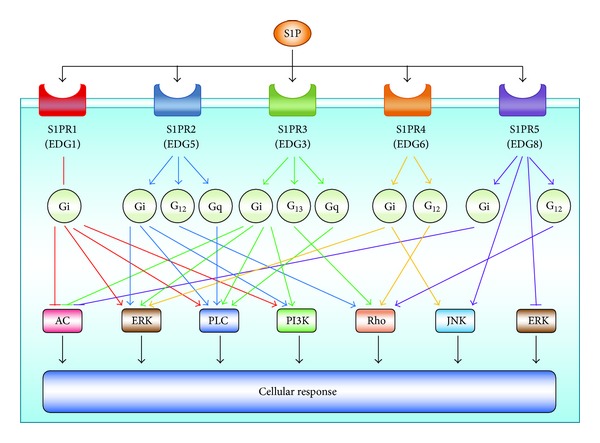
Downstream signaling pathways of sphingosine-1-phosphate (S1P) receptors. S1P is a ligand for the five different specific G protein coupled receptors: S1PR1/EDG1; S1PR2/EDG5; S1PR3/EDG3; S1PR4/EDG6; and S1PR5/EDG8. Each S1P receptor is coupled to different G proteins, which regulate activation or inhibition of the indicated downstream signaling pathways, such as adenylyl cyclase-cyclic AMP, AC; extracellular signal-regulated kinase, ERK; phospholipase C, PLC; phosphatidylinositol 3-kinase, PI3K; the small GTPases of the Rho family; and Jun amino terminal kinase, JNK. Only a few examples of these pathways are illustrated.
Cancer cells and each type of cell in the TME, such as inflammatory cells and endothelial cells, express different combinations of S1P receptors, which contribute to each cellular function regulated by S1P. For example, S1PR1 is important for B and T lymphocyte egression from secondary lymphatic organs, such as lymph nodes [25]. In endothelial cells, S1PR1 and S1PR2 are known to play an important role in vascular development [26–29]. Stimulation of S1PR1 and/or S1PR3 often promotes cell proliferation and migration in normal and cancer cells, while S1PR2 may inhibit the signaling that promotes cell proliferation and migration [30–32]. Altogether, this “inside-out” signaling of S1P plays a pivotal role in cancer cells and in the TME by stimulating the S1P receptors on each type of cell [33, 34].
In addition to its “inside-out” signaling, S1P is also known to have a variety of intracellular functions. Intracellular S1P produced by SphK1 can bind to TRAF2 (tumor necrosis factor receptor-associated factor 2) and function as a cofactor required for its E3 ubiquitin ligase activity and consequently, Lys-63-linked polyubiquitination of RIP1 (receptor-interacting protein 1) and NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) activation [35]. Similarly, it has been shown that S1P enhances cIAP2 (cellular inhibitor of apoptosis 2) mediated K63-linked polyubiquitination of IRF-1 (interferon regulatory factor-1), which is essential for IL-1-induced production of chemokines CXCL10 and CCL5 [36]. Furthermore, S1P produced by SphK2 in the nucleus acts as an endogenous inhibitor of specific histone deacetylases (HDAC1 and HDAC2), thereby regulating gene transcription, including that of the cyclin dependent kinase inhibitor p21 [37].
The relative levels of S1P within body fluids and tissues are important to a variety of physiologic processes [25, 38]. Levels of S1P are maintained by S1P synthesis and degradation, which create an S1P gradient within the tissue [39]. S1P is dephosphorylated to regenerate sphingosine by S1P phosphatases (SPPs) and/or lipid phosphate phosphatases (LPPs). S1P is also irreversibly degraded to hexadecenal and ethanolamine phosphate by S1P lyase (SPL). The current consensus is that trafficking of immune cells is controlled by this S1P gradient. For example, in the blood and lymph, S1P levels are relatively high, but in secondary lymphatic tissue, such as lymph nodes and the thymus, S1P is maintained at very low levels [40]. Importantly, any inhibition of S1P signaling, and therefore altering of this S1P gradient, results in alteration of immune cell trafficking. S1P also regulates vascular integrity. In the plasma, a decrease in the amount of S1P causes increased vascular permeability, likely due to loss of signaling through S1PR1 on endothelial cells [41–43]. S1P through its tissue gradients, intracellular functions, and “inside-out” signaling is important in both physiologic and pathologic processes.
3. Export of S1P via ATP-Binding Cassette (ABC) Transporters and Spinster 2 (Spns2) Transporter
Until recently, the process through which S1P produced inside cells by the two SphKs reaches its receptors on the cell surface remained obscure. Membrane lipids, including S1P, do not spontaneously exchange through the lipid bilayers of the plasma membrane since the polar head groups do not readily go through the hydrophobic interior. Though there are many transporter proteins on the membrane for lipid exchange [9], sphingosine is known to spontaneously translocate without the aid of a transporter across intracellular membranes when added to cells or produced intracellularly [9].
Studies from several laboratories, including ours, have suggested the involvement of ABC transporters in the export of S1P from various types of cells in vitro [10]. S1P has been shown to be exported from mast cells via ABCC1 (also known as multidrug resistant protein 1; MRP1) [9], from astrocytes via ABCA1 [44], from endothelial cells via ABCA1 and ABCC1 [45], and from thyroid carcinoma cells via ABCC1 [46]. Using pharmacological and molecular approaches, we demonstrated that ABCC1 and ABCG2 (also known as breast cancer resistance protein; BCRP) are involved in estradiol-mediated transport of S1P and dihydro-S1P out of MCF-7 human breast cancer cells [12]. S1P is exported from erythrocytes and platelets by other transporters in the ABC transporter family [47, 48]. In erythrocytes, S1P is exported by an ATP-dependent and vanadate- and glyburide-sensitive transporter [47], while in platelets, S1P export requires an extracellular stimulus such as thrombin and is exported through two independent transporters, a Ca2+-dependent transporter and an ATP-dependent glyburide-sensitive transporter [48, 49]. Collectively, these studies suggest that members of the large family of ABC transporters are responsible for export of S1P in various types of cells; however, in studies using mice with ABC transporter deficiencies, including animals with knockout of ABCA1, ABCA7, and ABCC1, S1P levels and related functions have been found to be unaltered [45], indicating the existence of compensatory mechanisms with other transporters.
Spns2, a member of the MFS (major facilitator superfamily) that does not have a typical ATP binding motif, has been recently discovered to export S1P from cells [11, 50–52]. Spns2 was identified independently by two groups, both of whom revealed that it transports S1P through observations in zebrafish. They showed that a mutation in Spns2 caused abnormal development resulting in cardia bifida (two hearts) [53] and that the phenotype of the Spns2 mutation is rescued by providing exogenous S1P [11]. The cardia bifida phenotype in the Spns2 knockout zebrafish was the clue that linked Spns2 to S1P, since the same phenotype was seen in S1PR2 knockout zebrafish [11, 53]. We have also shown that Spns2 can export endogenous S1P and dihydro-S1P from cells [13]. In addition, human Spns2 can transport several S1P analogues, including phosphorylated FTY720 [54]. Importantly, Spns2 was the first S1P transporter discovered to be physiologically functional in vivo, in contrast to the ABC transporters [54].
It has been suggested that Spns2 is important for vascular development [29]. We have observed that lymph nodes from Spns2 knockout mice have aberrant lymphatic sinuses that appear collapsed, with reduced numbers of lymphocytes [13]. Our data suggest that Spns2 is an S1P transporter in vivo that plays a role in regulation of levels of S1P not only in the blood, but also in the lymph nodes and lymphatic fluid, influencing lymphocyte trafficking and lymphatic vessel network organization [13]. The recent finding that blood endothelial cells purified from the aorta of Spns2-deficient mice are unable to release S1P [52] seems to support a strong role for Spns2 in S1P regulation. The levels of S1P in the plasma of Spns2 knockout mice have been observed to be decreased to 60% of that of wild type mice with endothelial cells contributing 40% of the total plasma S1P [54]. There was no difference between Spns2 knockout mice and wild type in S1P release activity in both erythrocytes and platelets, showing that a disruption of Spns2 does not affect the S1P release from erythrocytes or platelets [52]. Furthermore, bone marrow reconstitution studies revealed that Spns2 was not involved in S1P release from blood cells and suggested a role for Spns2 in other cells [51]. Consistent with these data, specific deletion of Spns2 on endothelial cells has been shown to result in a lack of lymphocyte egress, mimicking observations in global Spns2-knockout mice. These data suggest that Spns2 functions in endothelial cells, not blood cells, to establish the S1P gradient required for T and B cells to egress from their respective lymphoid organs [51].
4. Targeting S1P Transporters for Treatment of Cancer Patients
There have been an increasing number of studies, implicating roles for S1P in different stages of cancer progression in subtypes of both adult and pediatric malignancies. SphKs and S1P signaling have been suggested to also have a role in acquisition of drug resistance [55, 56]. There is a growing body of literature, with several clinical and pathological reports revealing the importance of SphK1 on cancer metastasis and prognosis [57–61]. Previous clinical studies have shown that SphK1 is overexpressed in human breast cancer and its expression correlates with poor patient outcomes [58, 62].
Studies linking the molecular interactions of S1P signaling with other oncogenic pathways, such as Ras, STAT3 (signal transducer and activator of transcription 3), NF-κB, and estrogen signaling, have been published. K-Ras mutations are known to increase the production of S1P in a SphK1-dependent manner and expression of the K-Ras oncogene leads to plasma membrane localization of SphK1. The RAS/RAF (rapidly accelerated fibrosarcoma)/MEK (mitogen-activated protein kinases)/ERK (extracellular signal-regulated kinases) pathway likely mediates this process, as constitutively active B-Raf or MEK are capable of activating SphK1 [63]. S1P produced by upregulated SphK1 in tumor cells activates S1PR1, which has been shown to lead to activation of STAT3 [34]. S1P is also involved in the activation of NF-κB, thereby regulating the transcription of the proinflammatory cytokines TNF-α (tumor necrosis factor-alpha) and IL-6 (interleukin-6) [34]. Estradiol is known to also stimulate SphK1 activation and the release of S1P, through which estradiol is capable of activating the S1PR3, resulting in EGFR transactivation in a matrix metalloprotease-dependent manner [31]. Altogether, these molecular interactions of S1P signaling with other oncogenic pathways suggest the importance of S1P signaling in cancer.
Communication among tumor cells, the host microenvironment, and inflammatory cells via systemic S1P regulates metastasis. S1P generated in cancer cells is secreted into tissue interstitial fluid in the body and affects the TME by altering immune cells, evoking inflammation, and inducing angiogenesis and lymphangiogenesis thereby promoting cancer metastasis [34, 64]. We have recently measured S1P levels within tumor interstitial fluid and found that significantly higher levels of S1P compared with the interstitial fluid of normal tissue and that inhibiting the “inside-out” signaling of S1P by FTY720 (known as fingolimod) significantly decreased S1P levels in the tumor interstitial fluid (unpublished data). Altogether, the “inside-out” signaling of S1P with overexpressed SphK1 and S1P transporters plays an important role in cancer progression through its effects on the TME.
It has recently been reported that a specific pharmacological inhibitor of SphK1 had no effect on cell proliferation in vitro [65]. This result generated some arguments that S1P, the product of SphK1, may not be an ideal anticancer target. On the other hand, targeting S1P signaling leads to a significant suppression of cancer progression in vivo [64], especially under conditions in which cancer is associated with inflammation [34]. Considering the importance of “inside-out” signaling of S1P within the TME through promotion of angiogenesis and lymphangiogenesis [4, 66, 67], the effects of targeting S1P signaling may only be adequately assessed in vivo.
ABC transporters were originally described as multidrug resistance genes and have been shown to be overexpressed in various solid and hematological cancers [68]. Expression of ABC transporters has been correlated with resistance to chemotherapies and poor prognosis for patients with certain types of cancer [68]. ABCB1, also known as multidrug resistant gene 1 (MDR1), has been targeted in a number of clinical trials that failed to demonstrate significant benefit [69]. Since it has been suggested that several ABC transporters are involved in secretion of S1P from stromal, endothelial, and cancer cells, it is possible that these transporters may play an important role in the pathological processes regulated by S1P and may worsen the biology of cancer cells. Considering that ABCB1 does not have a role for S1P transport, targeting ABCC1 and/or G2 may be a more promising treatment option for cancer patients. In contrast to the ABC transporters, with some of them originally described as multidrug resistant genes, the roles of Spns2 in cancer remain unknown.
Spns2, a 549 amino acids protein, belongs to the MFS transporter family as determined based on its predicted amino acid sequence [49]. The crystal structure of Spns2 and the precise mechanism of S1P transport via this transporter are still under investigation. S1P import by this transporter also remains to be described. To elucidate the role of Spns2 in cancer, we are currently in the midst of a process of developing gene targeting techniques utilizing nanoparticles to downregulate spns2 expression in cancer cells. How effective this may be in light of other transporters that contribute to S1P export, particularly in vivo setting, is yet to be determined. Further investigation is needed to clarify the function and role of Spns2 in normal physiological conditions and in the pathological condition of cancer.
The ABC transporter family and Spns2 are found in various types of cells and have different roles in each type of cell, so that it is possible that targeting a specific transporter may present some difficulties for cancer therapy for patients. In addition to targeting S1P transporters, a possible strategy for cancer treatment involves targeting S1P itself using a monoclonal antibody. In an animal model, neutralization of systemic S1P using Sphingomab, an anti-S1P monoclonal antibody, was shown to suppress lung metastasis [70]. The humanized version of Sphingomab, Sonepcizumab/ASONEP [71], has finished phase I trials and has recently entered phase II efficacy and safety studies for the treatment of renal cell carcinoma and age-related macular degeneration [72]. Targeting S1P itself may be one of the promising strategies for controlling cancer progression and metastasis.
5. Conclusion
S1P promotes a variety of intracellular and extracellular biological functions and is transported from inside cells through several members of the ABC transporter family and the enigmatic Spns2 transporter. Once through the plasma membrane, S1P exerts its “inside-out” signaling in the context of the TME and promotes cancer progression. Targeting S1P transporters and S1P signaling in cancer progression is a promising direction for development of the next generation of cancer therapeutics.
Acknowledgment
This work was supported by NIH Grants (R01CA160688 to Kazuaki Takabe and T32CA085159-10 to Krista P. Terracina).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Hanahan D, Coussens LM. Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell. 2012;21(3):309–322. doi: 10.1016/j.ccr.2012.02.022. [DOI] [PubMed] [Google Scholar]
- 2.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 3.Fang H, DeClerck YA. Targeting the tumor microenvironment: from understanding pathways to effective clinical trials. Cancer Research. 2013;73(16):4965–4977. doi: 10.1158/0008-5472.CAN-13-0661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Takabe K, Spiegel S. Export of sphingosine-1-phosphate and cancer progression. The Journal of Lipid Research. 2014 doi: 10.1194/jlr.R046656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wiig H, Tenstad O, Iversen PO, Kalluri R, Bjerkvig R. Interstitial fluid: the overlooked component of the tumor microenvironment? Fibrogenesis & Tissue Repair. 2010;3:p. 12. doi: 10.1186/1755-1536-3-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Haslene-Hox H, Oveland E, Berg KC, et al. A new method for isolation of interstitial fluid from human solid tumors applied to proteomic analysis of ovarian carcinoma tissue. PLoS ONE. 2011;6(4) doi: 10.1371/journal.pone.0019217.e19217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Wiig H, Swartz MA. Interstitial fluid and lymph formation and transport: physiological regulation and roles in inflammation and cancer. Physiological Reviews. 2012;92(3):1005–1060. doi: 10.1152/physrev.00037.2011. [DOI] [PubMed] [Google Scholar]
- 8.Kunkel GT, MacEyka M, Milstien S, Spiegel S. Targeting the sphingosine-1-phosphate axis in cancer, inflammation and beyond. Nature Reviews Drug Discovery. 2013;12(9):688–702. doi: 10.1038/nrd4099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mitra P, Oskeritzian CA, Payne SG, Beaven MA, Milstien S, Spiegel S. Role of ABCC1 in export of sphingosine-1-phosphate from mast cells. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(44):16394–16399. doi: 10.1073/pnas.0603734103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kim RH, Takabe K, Milstien S, Spiegel S. Export and functions of sphingosine-1-phosphate. Biochimica et Biophysica Acta. 2009;1791(7):692–696. doi: 10.1016/j.bbalip.2009.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kawahara A, Nishi T, Hisano Y, Fukui H, Yamaguchi A, Mochizuki N. The sphingolipid transporter Spns2 functions in migration of zebrafish myocardial precursors. Science. 2009;323(5913):524–527. doi: 10.1126/science.1167449. [DOI] [PubMed] [Google Scholar]
- 12.Takabe K, Kim RH, Allegood JC, et al. Estradiol induces export of sphingosine 1-phosphate from breast cancer cells via ABCC1 and ABCG2. The Journal of Biological Chemistry. 2010;285(14):10477–10486. doi: 10.1074/jbc.M109.064162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Nagahashi M, Kim EY, Yamada A, et al. Spns2, a transporter of phosphorylated sphingoid bases, regulates their blood and lymph levels, and the lymphatic network. The FASEB Journal. 2013;27(3):1001–1011. doi: 10.1096/fj.12-219618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Olivera A, Spiegel S. Sphingosine-1-phosphate as second messenger in cell proliferation induced by PDGF and FCS mitogens. Nature. 1993;365(6446):557–560. doi: 10.1038/365557a0. [DOI] [PubMed] [Google Scholar]
- 15.Takabe K, Paugh SW, Milstien S, Spiegel S. Inside-out signaling of sphingosine-1-phosphate: therapeutic targets. Pharmacological Reviews. 2008;60(2):181–195. doi: 10.1124/pr.107.07113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Shida D, Takabe K, Kapitonov D, Milstien S, Spiegel S. Targeting SphK1 as a new strategy against cancer. Current Drug Targets. 2008;9(8):662–673. doi: 10.2174/138945008785132402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pyne NJ, Pyne S. Sphingosine 1-phosphate and cancer. Nature Reviews Cancer. 2010;10(7):489–503. doi: 10.1038/nrc2875. [DOI] [PubMed] [Google Scholar]
- 18.Spiegel S, Milstien S. The outs and the ins of sphingosine-1-phosphate in immunity. Nature Reviews Immunology. 2011;11(6):403–415. doi: 10.1038/nri2974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Huang WC, Nagahashi M, Terracina KP, Takabe K. Emerging role of sphingosine-1-phosphate in inflammation, cancer, and lymphangiogenesis. Biomolecules. 2013;3(3):408–434. doi: 10.3390/biom3030408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Liu H, Chakravarty D, Maceyka M, Milstien S, Spiegel S. Sphingosine kinases: a novel family of lipid kinases. Progress in Nucleic Acid Research and Molecular Biology. 2002;71:493–511. doi: 10.1016/s0079-6603(02)71049-0. [DOI] [PubMed] [Google Scholar]
- 21.Maceyka M, Payne SG, Milstien S, Spiegel S. Sphingosine kinase, sphingosine-1-phosphate, and apoptosis. Biochimica et Biophysica Acta. 2002;1585(2-3):193–201. doi: 10.1016/s1388-1981(02)00341-4. [DOI] [PubMed] [Google Scholar]
- 22.Nava VE, Hobson JP, Murthy S, Milstien S, Spiegel S. Sphingosine kinase type 1 promotes estrogen-dependent tumorigenesis of breast cancer MCF-7 cells. Experimental Cell Research. 2002;281(1):115–127. doi: 10.1006/excr.2002.5658. [DOI] [PubMed] [Google Scholar]
- 23.Liu H, Toman RE, Goparaju SK, et al. Sphingosine kinase type 2 is a putative BH3-only protein that induces apoptosis. Journal of Biological Chemistry. 2003;278(41):40330–40336. doi: 10.1074/jbc.M304455200. [DOI] [PubMed] [Google Scholar]
- 24.Cuvillier O, Pirianov G, Kleuser B, et al. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nature. 1996;381(6585):800–803. doi: 10.1038/381800a0. [DOI] [PubMed] [Google Scholar]
- 25.Cyster JG, Schwab SR. Sphingosine-1-phosphate and lymphocyte egress from lymphoid organs. Annual Review of Immunology. 2012;30:69–94. doi: 10.1146/annurev-immunol-020711-075011. [DOI] [PubMed] [Google Scholar]
- 26.Shoham AB, Malkinson G, Krief S, et al. S1P1 inhibits sprouting angiogenesis during vascular development. Development. 2012;139(20):3859–3869. doi: 10.1242/dev.078550. [DOI] [PubMed] [Google Scholar]
- 27.Gaengel K, Niaudet C, Hagikura K, et al. The sphingosine-1-phosphate receptor s1pr1 restricts sprouting angiogenesis by regulating the interplay between VE-cadherin and VEGFR2. Developmental Cell. 2012;23(3):587–599. doi: 10.1016/j.devcel.2012.08.005. [DOI] [PubMed] [Google Scholar]
- 28.Jung B, Obinata H, Galvani S, et al. Flow-regulated endothelial S1P receptor-1 signaling sustains vascular development. Developmental Cell. 2012;23(3):600–610. doi: 10.1016/j.devcel.2012.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mendelson K, Zygmunt T, Torres-Vázquez J, Evans T, Hla T. Sphingosine 1-phosphate receptor signaling regulates proper embryonic vascular patterning. The Journal of Biological Chemistry. 2013;288(4):2143–2156. doi: 10.1074/jbc.M112.427344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rosenfeldt HM, Hobson JP, Maceyka M, et al. EDG-1 links the PDGF receptor to Src and focal adhesion kinase activation leading to lamellipodia formation and cell migration. The FASEB Journal. 2001;15(14):2649–2659. doi: 10.1096/fj.01-0523com. [DOI] [PubMed] [Google Scholar]
- 31.Sukocheva O, Wadham C, Holmes A, et al. Estrogen transactivates EGFR via the sphingosine 1-phosphate receptor Edg-3: the role of sphingosine kinase-1. Journal of Cell Biology. 2006;173(2):301–310. doi: 10.1083/jcb.200506033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Goparaju SK, Jolly PS, Watterson KR, et al. The S1P2 receptor negatively regulates platelet-derived growth factor-induced motility and proliferation. Molecular & Cellular Biology. 2005;25(10):4237–4249. doi: 10.1128/MCB.25.10.4237-4249.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nagahashi M, Ramachandran S, Rashid OM, Takabe K. Lymphangiogenesis: a new player in cancer progression. World Journal of Gastroenterology. 2010;16(32):4003–4012. doi: 10.3748/wjg.v16.i32.4003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Liang J, Nagahashi M, Kim EY, et al. Sphingosine-1-phosphate links persistent STAT3 activation, chronic intestinal inflammation, and development of colitis-associated cancer. Cancer Cell. 2013;23(1):107–120. doi: 10.1016/j.ccr.2012.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Alvarez SE, Harikumar KB, Hait NC, et al. Sphingosine-1-phosphate is a missing cofactor for the E3 ubiquitin ligase TRAF2. Nature. 2010;465(7301):1084–1088. doi: 10.1038/nature09128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Harikumar KB, Yester JW, Surace MJ. K63-linked polyubiquitination of transcription factor IRF1 is essential for IL-1-induced production of chemokines CXCL10 and CCL5. Nature Immunology. 2014;15(3):231–238. doi: 10.1038/ni.2810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hait NC, Allegood J, Maceyka M, et al. Regulation of histone acetylation in the nucleus by sphingosine-1-phosphate. Science. 2009;325(5945):1254–1257. doi: 10.1126/science.1176709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Schwab SR, Cyster JG. Finding a way out: Lymphocyte egress from lymphoid organs. Nature Immunology. 2007;8(12):1295–1301. doi: 10.1038/ni1545. [DOI] [PubMed] [Google Scholar]
- 39.Serra M, Saba JD. Sphingosine 1-phosphate lyase, a key regulator of sphingosine 1-phosphate signaling and function. Advances in Enzyme Regulation. 2010;50(1):349–362. doi: 10.1016/j.advenzreg.2009.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Schwab SR, Pereira JP, Matloubian M, Xu Y, Huang Y, Cyster JG. Immunology: lymphocyte sequestration through S1P lyase inhibition and disruption of S1P gradients. Science. 2005;309(5741):1735–1739. doi: 10.1126/science.1113640. [DOI] [PubMed] [Google Scholar]
- 41.Camerer E, Regard JB, Cornelissen I, et al. Sphingosine-1-phosphate in the plasma compartment regulates basal and inflammation-induced vascular leak in mice. Journal of Clinical Investigation. 2009;119(7):1871–1879. doi: 10.1172/JCI38575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lee MJ, Thangada S, Claffey KP, et al. Vascular endothelial cell adherens junction assembly and morphogenesis induced by sphingosine-1-phosphate. Cell. 1999;99(3):301–312. doi: 10.1016/s0092-8674(00)81661-x. [DOI] [PubMed] [Google Scholar]
- 43.Mendoza A, Bréart B, Ramos-Perez WD, et al. The transporter Spns2 is required for secretion of lymph but not plasma sphingosine-1-phosphate. Cell Reports. 2012;2(5):1104–1110. doi: 10.1016/j.celrep.2012.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sato K, Malchinkhuu E, Horiuchi Y, et al. Critical role of ABCA1 transporter in sphingosine 1-phosphate release from astrocytes. Journal of Neurochemistry. 2007;103(6):2610–2619. doi: 10.1111/j.1471-4159.2007.04958.x. [DOI] [PubMed] [Google Scholar]
- 45.Lee YM, Venkataraman K, Hwang SI, Han DK, Hla T. A novel method to quantify sphingosine 1-phosphate by immobilized metal affinity chromatography (IMAC) Prostaglandins and Other Lipid Mediators. 2007;84(3-4):154–162. doi: 10.1016/j.prostaglandins.2007.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bergelin N, Blom T, Heikkilä J, et al. Sphingosine kinase as an oncogene: autocrine sphingosine 1-phoshate modulates ML-1 thyroid carcinoma cell migration by a mechanism dependent on protein kinase C-α and ERK1/2. Endocrinology. 2009;150(5):2055–2063. doi: 10.1210/en.2008-0625. [DOI] [PubMed] [Google Scholar]
- 47.Kobayashi N, Yamaguchi A, Nishi T. Characterization of the ATP-dependent sphingosine 1-phosphate transporter in rat erythrocytes. The Journal of Biological Chemistry. 2009;284(32):21192–21200. doi: 10.1074/jbc.M109.006163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kobayashi N, Nishi T, Hirata T, et al. Sphingosine 1-phosphate is released from the cytosol of rat platelets in a carrier-mediated manner. Journal of Lipid Research. 2006;47(3):614–621. doi: 10.1194/jlr.M500468-JLR200. [DOI] [PubMed] [Google Scholar]
- 49.Nishi T, Kobayashi N, Hisano Y, Kawahara A, Yamaguchi A. Molecular and physiological functions of sphingosine 1-phosphate transporters. Biochimica et Biophysica Acta: Molecular and Cell Biology of Lipids. 2013;1841(5):759–765. doi: 10.1016/j.bbalip.2013.07.012. [DOI] [PubMed] [Google Scholar]
- 50.Hisano Y, Kobayashi N, Kawahara A, Yamaguchi A, Nishi T. The sphingosine 1-phosphate transporter, SPNS2, functions as a transporter of the phosphorylated form of the immunomodulating agent FTY720. The Journal of Biological Chemistry. 2011;286(3):1758–1766. doi: 10.1074/jbc.M110.171116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Fukuhara S, Simmons S, Kawamura S, et al. The sphingosine-1-phosphate transporter Spns2 expressed on endothelial cells regulates lymphocyte trafficking in mice. Journal of Clinical Investigation. 2012;122(4):1416–1426. doi: 10.1172/JCI60746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hisano Y, Kobayashi N, Yamaguchi A, Nishi T. Mouse SPNS2 functions as a sphingosine-1-phosphate transporter in vascular endothelial cells. PLoS ONE. 2012;7(6) doi: 10.1371/journal.pone.0038941.e38941 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Osborne N, Brand-Arzamendi K, Ober EA, et al. The spinster homolog, two of hearts, is required for sphingosine 1-phosphate signaling in zebrafish. Current Biology. 2008;18(23):1882–1888. doi: 10.1016/j.cub.2008.10.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hisano Y, Nishi T, Kawahara A. The functional roles of S1P in immunity. Journal of Biochemistry. 2012;152(4):305–311. doi: 10.1093/jb/mvs090. [DOI] [PubMed] [Google Scholar]
- 55.Salas A, Ponnusamy S, Senkal CE, et al. Sphingosine kinase-1 and sphingosine 1-phosphate receptor 2 mediate Bcr-Abl1 stability and drug resistance by modulation of protein phosphatase 2A. Blood. 2011;117(22):5941–5952. doi: 10.1182/blood-2010-08-300772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cuvillier O, Ader I, Bouquerel P, Brizuela L, Gstalder C, Malavaud B. Hypoxia, therapeutic resistance, and sphingosine 1-phosphate. Advances in Cancer Research. 2013;117:117–141. doi: 10.1016/B978-0-12-394274-6.00005-4. [DOI] [PubMed] [Google Scholar]
- 57.van Brooklyn JR, Jackson CA, Pearl DK, Kotur MS, Snyder PJ, Prior TW. Sphingosine kinase-1 expression correlates with poor survival of patients with glioblastoma multiforme: roles of sphingosine kinase isoforms in growth of glioblastoma cell lines. Journal of Neuropathology and Experimental Neurology. 2005;64(8):695–705. doi: 10.1097/01.jnen.0000175329.59092.2c. [DOI] [PubMed] [Google Scholar]
- 58.Ruckhäberle E, Rody A, Engels K, et al. Microarray analysis of altered sphingolipid metabolism reveals prognostic significance of sphingosine kinase 1 in breast cancer. Breast Cancer Research and Treatment. 2008;112(1):41–52. doi: 10.1007/s10549-007-9836-9. [DOI] [PubMed] [Google Scholar]
- 59.Li W, Yu CP, Xia JT, et al. Sphingosine kinase 11s associated with gastric cancer progression and poor survival of patients. Clinical Cancer Research. 2009;15(4):1393–1399. doi: 10.1158/1078-0432.CCR-08-1158. [DOI] [PubMed] [Google Scholar]
- 60.Liu SQ, Su YJ, Qin MB, Mao YB, Huang JA, Tang GD. Sphingosine kinase 1 promotes tumor progression and confers malignancy phenotypes of colon cancer by regulating the focal adhesion kinase pathway and adhesion molecules. International Journal of Oncology. 2013;42(2):617–626. doi: 10.3892/ijo.2012.1733. [DOI] [PubMed] [Google Scholar]
- 61.Pyne S, Edwards J, Ohotski J, Pyne NJ. Sphingosine 1-phosphate receptors and sphingosine kinase 1: novel biomarkers for clinical prognosis in breast, prostate, and hematological cancers. Frontiers in Oncology. 2012;2(article 168) doi: 10.3389/fonc.2012.00168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zhang Y, Wang Y, Wan Z, Liu S, Cao Y, Zeng Z. Sphingosine kinase 1 and cancer: a systematic review and meta-analysis. PLoS ONE. 2014;9(2) doi: 10.1371/journal.pone.0090362.e90362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gault CR, Eblen ST, Neumann CA, Hannun YA, Obeid LM. Oncogenic K-Ras regulates bioactive sphingolipids in a sphingosine kinase 1-dependent manner. The Journal of Biological Chemistry. 2012;287(38):31794–31803. doi: 10.1074/jbc.M112.385765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Nagahashi M, Ramachandran S, Kim EY, et al. Sphingosine-1-phosphate produced by sphingosine kinase 1 promotes breast cancer progression by stimulating angiogenesis and lymphangiogenesis. Cancer Research. 2012;72(3):726–735. doi: 10.1158/0008-5472.CAN-11-2167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Schnute ME, McReynolds MD, Kasten T, et al. Modulation of cellular S1P levels with a novel, potent and specific inhibitor of sphingosine kinase-1. Biochemical Journal. 2012;444(1):79–88. doi: 10.1042/BJ20111929. [DOI] [PubMed] [Google Scholar]
- 66.Takabe K, Yamada A, Rashid OM. Twofer anti-vascular therapy targeting sphingosine-1-phosphate for breast cancer. Gland Surgery. 2012;1(2):80–83. doi: 10.3978/j.issn.2227-684X.2012.07.01. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Aoyagi T, Nagahashi M, Yamada A, Takabe K. The role of sphingosine-1-phosphate in breast cancer tumor-induced lymphangiogenesis. Lymphatic Research and Biology. 2012;10(3):97–106. doi: 10.1089/lrb.2012.0010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kruh GD, Belinsky MG. The MRP family of drug efflux pumps. Oncogene. 2003;22(47):7537–7552. doi: 10.1038/sj.onc.1206953. [DOI] [PubMed] [Google Scholar]
- 69.Amiri-Kordestani L, Basseville A, Kurdziel K, Fojo AT, Bates SE. Targeting MDR in breast and lung cancer: discriminating its potential importance from the failure of drug resistance reversal studies. Drug Resistance Updates. 2012;15(1-2):50–61. doi: 10.1016/j.drup.2012.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ponnusamy S, Selvam SP, Mehrotra S, et al. Communication between host organism and cancer cells is transduced by systemic sphingosine kinase 1/sphingosine 1-phosphate signalling to regulate tumour metastasis. EMBO Molecular Medicine. 2012;4(8):761–775. doi: 10.1002/emmm.201200244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sabbadini RA. Sphingosine-1-phosphate antibodies as potential agents in the treatment of cancer and age-related macular degeneration. British Journal of Pharmacology. 2011;162(6):1225–1238. doi: 10.1111/j.1476-5381.2010.01118.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Schwalm S, Pfeilschifter J, Huwiler A. Targeting the sphingosine kinase/sphingosine 1-phosphate pathway to treat chronic inflammatory kidney diseases. Basic and Clinical Pharmacology and Toxicology. 2014;114(1):44–49. doi: 10.1111/bcpt.12103. [DOI] [PubMed] [Google Scholar]