

Abstract
99mTc is the standard radionuclide used for nuclear medicine imaging. In addition to gamma irradiation, 99mTc emits low-energy Auger and conversion electrons that deposit their energy within nanometers of the decay site. To study the potential for DNA damage, direct DNA binding is required. Plasmid DNA enables the investigation of the unprotected interactions between molecules and DNA that result in single-strand breaks (SSBs) or double-strand breaks (DSBs); the resulting DNA fragments can be separated by gel electrophoresis and quantified by fluorescent staining. This study aimed to compare the plasmid DNA damage potential of a 99mTc-labeled HYNIC-DAPI compound with that of 99mTc pertechnetate (99mTcO4 −). pUC19 plasmid DNA was irradiated for 2 or 24 hours. Direct and radical-induced DNA damage were evaluated in the presence or absence of the radical scavenger DMSO. For both compounds, an increase in applied activity enhanced plasmid DNA damage, which was evidenced by an increase in the open circular and linear DNA fractions and a reduction in the supercoiled DNA fraction. The number of SSBs elicited by 99mTc-HYNIC-DAPI (1.03) was twice that caused by 99mTcO4 − (0.51), and the number of DSBs increased fivefold in the 99mTc-HYNIC-DAPI-treated sample compared with the 99mTcO4 − treated sample (0.02 to 0.10). In the presence of DMSO, the numbers of SSBs and DSBs decreased to 0.03 and 0.00, respectively, in the 99mTcO4 – treated samples, whereas the numbers of SSBs and DSBs were slightly reduced to 0.95 and 0.06, respectively, in the 99mTc-HYNIC-DAPI-treated samples. These results indicated that 99mTc-HYNIC-DAPI induced SSBs and DSBs via a direct interaction of the 99mTc-labeled compound with DNA. In contrast to these results, 99mTcO4 − induced SSBs via radical formation, and DSBs were formed by two nearby SSBs. The biological effectiveness of 99mTc-HYNIC-DAPI increased by approximately 4-fold in terms of inducing SSBs and by approximately 10-fold in terms of inducing DSBs.
Introduction
The study of cellular DNA damage by various chemical and physical events is hampered by the existence of effective repair systems. Moreover, cellular and nuclear membranes are barriers that are difficult to overcome. Plasmid DNA is a model that enables the investigation of unprotected interactions between molecules and DNA that result in single-strand breaks (SSBs) or double-strand breaks (DSB); the resulting fragments can be separated by gel electrophoresis and quantified by fluorescent staining. Auger electron-emitting radiotracers can be implanted directly into DNA via the incorporation of radiolabeled thymidine analogues or by indirect DNA labeling with radioactive dyes to study the influence of geometry [1], [2] or to characterize different isotopes of the same element [3].
Most data have been obtained using 125I-labeled radiotracers. Recently, 99mTc has become an interesting candidate for imaging and radiotherapy. Haefliger et al. demonstrated the induction of DSBs by Auger electrons from 99mTc complexes with DNA-binding ligands using gel electrophoresis; however, the damage was not quantified under consideration of the absorbed dose [4]. Santos-Cuevas et al. demonstrated by microscopy and with proliferation assays that 99mTc-labeled bombesin effectively trafficked to the nucleus in prostate and breast cancer cells [5]. Schipper et al. reported that 99mTc and 131I accumulated in NIS-positive neuroendocrine tumor cells in vitro and in a preclinical model, suggesting the potential for both imaging and therapy [6]. Kriehuber et al. investigated cell survival, the induction of apoptosis and micronucleus formation in SCL-II cells after exposure to the Auger electron emitter 99mTc [7], but cellular uptake was not sufficient to ascertain the radiobiological effectiveness. Kotzerke et al. compared the uptake of 99mTc versus 211At in NIS-positive rat thyroid cells [8], Wendisch et al. investigated the clonogenic survival after intracellular uptake of 99mTc vs. 131I [9], whereas Freudenberg et al. compared 99mTc to 188Re. [10]. Because nuclear uptake was negligible, DNA damage by low-energy Auger electrons could not be detected. Wunderlich et al. increased the intracellular accumulation of 99mTc by preincubation with Sn-complexes [11]. Recently, the importance of low-energy Auger and conversion electrons was reported by Cambien et al., who investigated 99mTcO4 − mediated thyroid stunning in vivo [12].
One possibility for transporting radioisotopes into the nucleus is to use radiolabeled fluorescent dyes [13], [14]. DAPI (4′,6-diamidino-2-phenylindole) is a DNA-specific fluorescent probe that interacts with DNA by inserting into the minor grooves of AT-rich DNA sequences [15]. In our study, DAPI was labeled with 99mTc via a HYNIC (6-hydrazinonicotinic) linker. We characterized the damage to plasmid DNA caused by the Auger electron emitter 99mTc using quantitative gel electrophoresis in the presence or absence of the radical scavenger DMSO and compared the radiobiological effectiveness with that of unbound 99mTc-pertechnetate.
Methods
Preparation of 99mTc-HYNIC-DAPI
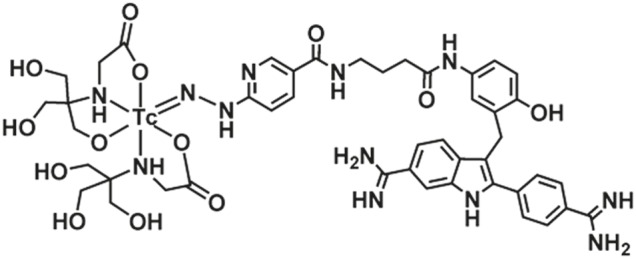
Chemicals were purchased from Sigma-Aldrich Chemical Company (Taufkirchen, Germany) unless otherwise stated. The following procedure was used to radiolabel 6-hydrazinonicotinamide (HYNIC)-conjugated DAPI (N-(4-(3-((6-carbamimidoyl-2-(4-carbamimidoylphenyl)-1H-indol-3-yl)methyl)-4-hydroxyphenylamino)-4-oxobutyl)-6-(2-(2,2,2-trifluoracetoxy)hydrazinyl)nicotinamide) (HYNIC-DAPI; ABX, Radeberg, Germany) with 99mTc (Figure 1). Briefly, 0.2 mL of an argon-purged N-(2-hydroxy-1,1-bis(hydroxymethyl)ethyl)glycine (tricine) coligand solution (200 mg tricine (0.11 mmol)/10 mL deionized water plus 1.5 mg tin(II)chloride dihydrate (0.53 mmol)) was added to 5 µg (7.00*10−9 mol) of HYNIC-DAPI. After 5 min, 1 mL of 99mTc generator eluate (1.5–2 GBq, Mallinckrodt Pharmaceuticals, Neustadt, Germany) was added, and the mixture was lightly shaken for 30–60 min at room temperature. The radiochemical purity of [99mTc]Tc(Tricin)nHYNIC-DAPI (99mTc-HYNIC-DAPI) was determined by analytical HPLC (Chromolith Performance RP-18e 100-4.6 mm, Merck, Darmstadt, Germany) using a linear solvent gradient of 4–15% B over 20 min (A = H2O with 0.05% TFA; B = acetonitrile with 0.05% TFA) and a flow rate of 1.4 mL/min. The same HPLC conditions were used to determine the stability of the radioligand solution after storing it at room temperature.
Figure 1. Supposed structure of [99mTc]Tc(Tricin)nHYNIC-DAPI (99mTc-HYNIC-DAPI) with HYNIC as monodentate ligand.
Plasmid DNA
The pUC19 plasmid (2686 base pairs; MW 1.75*106 Daltons) originating from E. coli ER2272 was purchased from New England Biolabs, Ipswich, UK.
DNA stock solutions were diluted in TE buffer (10 mM Tris-HCl and 1 mM EDTA, pH 7.5) to a final concentration of 0.1 µg/µL and were stored at −20°C. Only samples containing > 95% closed circular DNA were used. pUC19 was digested with BamHI (Invitrogen, Karlsruhe, Germany) to obtain linear DNA.
Irradiation Procedure
The necessary samples, each containing 200 ng plasmid DNA at 0.1 µg/µL and various amounts of the radioactive solutions, were placed in microtubes (Eppendorf, Hamburg, Germany). One radiotracer-free microtube served as the control.
99mTc-HYNIC-DAPI was added to the pUC19 plasmid solutions (0.1–25 MBq/20 µL = 5.0–1250 MBq/mL) in a total sample volume of 20 µL.
99mTc-pertechnetate solutions were adjusted to the same radioactivity as the 99mTc-HYNIC-DAPI solutions and were also added to the plasmid samples. In an additional test series, 0.2 M DMSO (final concentration) was added to the radiotracer solutions. Each sample was separated into two 10 µL aliquots to enable the accumulation of radiation dose for 2 h at 4°C or for 24 h at −20°C.
After irradiation, 10 µL of each irradiated DNA solution was mixed with 1.25 µL of loading buffer. The mixture was pipetted into the wells of a 1.4% agarose gel in TAE buffer. The samples were run at 4 V/cm for 120 min at 6°C. After electrophoresis, the closed circular (supercoiled), open circular and linear plasmid DNA fractions were identified based on mobility differences in the gel. The gel was stained with ethidium bromide (0.5 µg/mL) for 30 min, and then the different forms of DNA were visualized using an UV transilluminator (Diana III Digital Imaging System, Raytest, Straubenhardt, Germany). The gel was imaged using a charge-coupled device (CCD) camera. The relative amount of DNA in each conformation was quantified by integrating the corresponding intensity using the open-source platform software Fiji [16]. To quantitate the DNA-bound 99mTc-HYNIC-DAPI activity, the DNA bands corresponding to the supercoiled, linear and open circular forms were excised, and the gel pieces were analyzed using a gamma counter (Cobra II Auto-Gamma, Perkin Elmer life sciences).
Dosimetry
The dose calculations were performed using the Geant4 Monte Carlo toolkit. Although the real shape of the liquid volume in the microtube is similar to an inverse cone with a spherical apex, the target volume was considered to be a sphere with a 10-µL volume composed of water. The decay sites were chosen to be randomly distributed inside the sphere, corresponding to a homogeneous activity distribution. The emission spectrum of 99mTc was taken from Howell [17] (Table 1). An S value for self-irradiation was obtained by simulating 1 million trajectories and scoring the energy deposition inside the target volume. The total dose was calculated as the product of S and the time-integrated activity. It must be noted that no plasmid binding of radioactivity was considered, because that would require nanodosimetric calculations for the plasmid DNA.
Table 1. Emission Spectrum of 99mTc (Data from Howell [17]).
| Nomenclature | Average energy (keV) | Yield per decay | Range (µm) |
| gamma2 | 141 | 88.9% | |
| IC 1 M,N | 1.82 | 99.1% | 0.165 |
| IC 2 K | 119 | 8.43% | 193 |
| IC 2 L | 137 | 1.36% | 244 |
| IC 2 M,N | 140 | 0.37% | 251 |
| IC 3 K | 122 | 0.59% | 199 |
| IC 3 L | 140 | 0.25% | 250 |
| Auger KLL | 15.3 | 1.26% | 5.57 |
| Auger KLX | 17.8 | 0.47% | 7.25 |
| CK LLX | 0.043 | 1.93% | 0.003 |
| Auger LMM | 2.05 | 8.68% | 0.199 |
| Auger LMX | 2.32 | 1.37% | 0.241 |
| Auger LXY | 2.66 | 0.12% | 0.3 |
| CK MMX | 0.116 | 74.7% | 0.006 |
| Auger MXY | 0.226 | 110% | 0.011 |
| CK NNX | 0.033 | 198% | 0.002 |
| X Kalpha1 | 18.4 | 3.89% | |
| X Kalpha2 | 18.3 | 2.17% | |
| X Kbeta1 | 20.6 | 0.76% | |
| X Kbeta2 | 21.0 | 0.15% | |
| X Kbeta3 | 20.6 | 0.27% | |
| X L | 2.45 | 0.49% | |
| X M | 0.236 | 0.12% |
Calculation of SSBs and DSBs
The mean number of single-strand breaks (NSSB) and double-strand breaks (NDSB) per plasmid were calculated based on the theoretical considerations published by Cowan et al. [18]. NDSB was estimated based on the relative proportion L of linear plasmids:
NSSB was calculated using the fraction SC of supercoiled plasmids:
Results
Preparation of 99mTc-HYNIC-DAPI
The yield and stability of 99mTc-HYNIC-DAPI in the reaction solution (saline) were determined using reverse-phase HPLC (Figure 2). The reaction yield of 99mTc-HYNIC-DAPI increased to >95% in one hour. 99mTc-HYNIC-DAPI exhibited low stability in the kit solution (50% at 6 h, <1% after 12 h) at room temperature. Therefore, the peak at a shorter retention time (1.33 min) in the HPLC trace, representative of molecules such as pertechnetate, increased from <5% at 1 h to approximately 100% at 12 h.
Figure 2. RP-HPLC Chromatogram of 99mTc-HYNIC-DAPI.
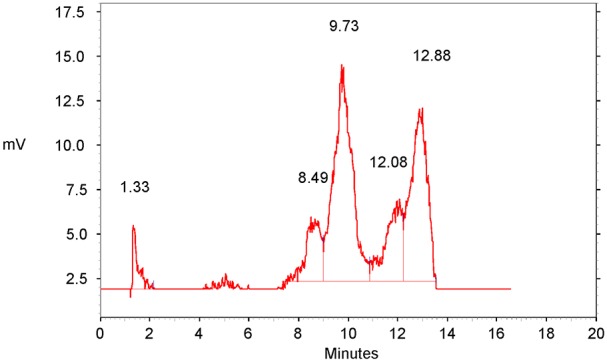
HPLC trace of the radioactivity 60(3.6%). The signals from 8.49 to 12.88 min represent different 99mTc-HYNIC-DAPI derivatives (96.4%). After 12 h, the peak at 1.33 min increased to 100%.
Dose calculations
The S value for the dose calculations was 2.53*10−10 Gy/(Bq s) inside a sphere with a 10-µL volume and a homogeneous activity distribution. Therefore, incubating plasmids with 1000 MBq/mL of 99mTc in 10 µL for 2 h yields a dose of 16.3 Gy. Irradiation for 24 h produces a total dose of 74 Gy.
Strand breaks in pUC19 plasmid DNA
The fractions of the different conformations of plasmid DNA (open circular, linear and supercoiled) that were observed after 2 or 24 h of irradiation with 99mTc-pertechnetate or 99mTc-HYNIC-DAPI are presented in Figure 3. For both compounds, an increase in applied activity enhanced the plasmid damage, resulting in an increase in the open circular and linear fractions and a decrease in the supercoiled fraction. The DNA damage was more extensive after 24 h of irradiation than after 2 h of irradiation. Compared with 99mTc-pertechnetate, the radiotoxicity of 99mTc-HYNIC-DAPI was obviously higher. For example, after a 24 h irradiation with 1000 MBq/mL of 99mTc-pertechnetate or 99mTc-HYNIC-DAPI, the open circular DNA fractions were 39.3% and 58.2%, respectively, and the linear DNA fractions were 1.9% and 9.3%, respectively.
Figure 3. Dose-Response of 99mTc-pertechnetate and 99mTc-HYNIC-DAPI.

Dose-dependent decrease in supercoiled DNA exposed to 99mTc-pertechnetate or 99mTc-HYNIC-DAPI for 2 (A) or 24 hours (B). The dose-dependent increase in open circular DNA was similar for both radiotracers, but the fraction of linear DNA was greater in the presence of 99mTc-HYNIC-DAPI compared with 99mTc-pertechnetate.
Based on the measured fluorescence intensities, the numbers of SSBs and DSBs were calculated (Table 2). In the example above, the number of SSBs created by 99mTc-HYNIC-DAPI (1.03) was twice that elicited by 99mTc-pertechnetate (0.51). The number of DSBs increased fivefold (from 0.02 to 0.10) in the 99mTc-HYNIC-DAPI-treated samples compared with the 99mTc-pertechnetate-treated samples. The ratio of the number of SSBs to the number of DSBs decreased from 25 for 99mTc-pertechnetate to 10 for 99mTc-HYNIC-DAPI.
Table 2. Mean numbers of DSBs and SSBs induced by 99mTc pertechnetate and 99mTc-HYNIC-DAPI in the absence or presence of DMSO.
| Activity(MBq/mL) | − DMSO | + DMSO | |||||||
| 99mTc-pertechnetate | 99mTc-HYNIC-DAPI | 99mTc-pertechnetate | 99mTc-HYNIC-DAPI | ||||||
| SSBs | DSBs | SSBs | DSBs | SSBs | DSBs | SSBs | DSBs | ||
| 2 h | 0 | 0.02 | 0.00 | 0.02 | 0.00 | ||||
| 2.5 | 0.02 | 0.00 | 0.02 | 0.00 | |||||
| 5 | 0.01 | 0.00 | 0.02 | 0.00 | |||||
| 25 | 0.02 | 0.00 | 0.04 | 0.00 | |||||
| 50 | 0.02 | 0.00 | 0.06 | 0.01 | 0.05 | 0.00 | |||
| 125 | 0.04 | 0.00 | 0.12 | 0.02 | 0.07 | 0.01 | |||
| 250 | 0.07 | 0.00 | 0.24 | 0.03 | 0.16 | 0.02 | |||
| 500 | 0.21 | 0.01 | 0.33 | 0.04 | 0.24 | 0.02 | |||
| 1000 | 0.38 | 0.01 | 0.60 | 0.06 | 0.02 | 0.00 | 0.46 | 0.02 | |
| 1250 | 0.47 | 0.01 | 0.59 | 0.06 | 0.02 | 0.00 | 0.55 | 0.03 | |
| 24 h | 0 | 0.03 | 0.00 | 0.03 | 0.00 | ||||
| 2.5 | 0.04 | 0.00 | 0.03 | 0.00 | |||||
| 5 | 0.02 | 0.00 | 0.05 | 0.00 | |||||
| 25 | 0.05 | 0.00 | 0.11 | 0.02 | |||||
| 50 | 0.10 | 0.00 | 0.15 | 0.02 | 0.11 | 0.01 | |||
| 125 | 0.15 | 0.00 | 0.32 | 0.04 | 0.18 | 0.02 | |||
| 250 | 0.16 | 0.01 | 0.49 | 0.06 | 0.47 | 0.04 | |||
| 500 | 0.31 | 0.02 | 0.70 | 0.08 | 0.52 | 0.04 | |||
| 1000 | 0.51 | 0.02 | 1.03 | 0.10 | 0.03 | 0.00 | 0.95 | 0.06 | |
| 1250 | 0.61 | 0.02 | 1.09 | 0.09 | 0.03 | 0.00 | 1.37 | 0.08 | |
To assess the chemotoxicity of unlabeled HYNIC-DAPI, we performed experiments with an identical setup using the same ligand concentrations as those used for 99mTc-HYNIC-DAPI. The results revealed that supercoiled plasmid DNA remained completely intact throughout all the incubations in the presence or absence of DMSO. Thus, no chemotoxic effects were observed (data not shown).
To verify the contribution of reactive oxygen species to the formation of SSBs and DSBs, additional experiments were performed in the absence or presence of 0.2 M DMSO. Irradiation of plasmid DNA with 99mTc-HYNIC-DAPI produced similar effects in the absence or presence of DMSO (Figures 4 and 5). For example, the open circular and linear fractions did not noticeably change. After 24 h, 1000 MBq/mL of 99mTc-HYNIC-DAPI resulted in (56.0 ±1.1)% open circular DNA in the absence of DMSO and (57.8±6.4)% open circular DNA in the presence of DMSO. The corresponding linear fractions were (7.2±1.6)% and (6.0±1.7)%, respectively. These results demonstrated that SSBs and DSBs were not markedly reduced by DMSO, suggesting that direct interactions between the 99mTc-labeled compound and DNA predominated. In contrast to these results, the number of SSBs induced by 1000 MBq/mL 99mTc-pertechnetate in the absence of DMSO was significantly reduced from 40% open circular DNA to only 3.0%. Furthermore, the formation of DSBs was completely prevented when the 99mTc-pertechnetate irradiation occurred in the presence of DMSO, as indicated by the absence of a linear DNA fraction (Figure 5). The dependence of SSBs and DSBs on the total number of disintegrations in the presence or absence of DMSO is illustrated in Figure 6.
Figure 4. Example images of agarose gel indicating the effect of DMSO.
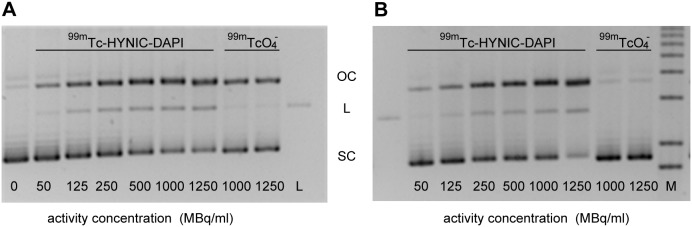
Representative images of agarose gels in the absence (A) or presence (B) of DMSO. The plasmid DNA samples were irradiated for 24 h with 50–1250 MBq/mL 99mTc-HYNIC-DAPI. The influence of DMSO on DNA damage (OC: open circular, L: linear, SC: super coiled plasmid DNA fraction) caused by 99mTc-HYNIC-DAPI was compared with that elicited by 99mTc-pertechnetate (99mTcO4 −). Notably, the formation of open circular and linear DNA in response to 99mTc-pertechnetate could be prevented by DMSO. The first lane is a non-irradiated plasmid sample, L is a non-irradiated linear plasmid, and M is the marker. The DNA bands at the bottom represent supercoiled DNA.
Figure 5. Effect of DMSO on plasmid damage.

The influence of the radical scavenger DMSO on the DNA conformational changes caused by 99mTc-pertechnetate and 99mTc-HYNIC-DAPI after 2 (A) or 24 h (B) of irradiation. The formation of open circle and linear DNA in response to 99mTc-pertechnetate was nearly completely suppressed in the presence of DMSO. These data indicated that the plasmid damage by 99mTc-pertechnetate was primarily caused by radicals. There was no noticeable effect of DMSO in the 99mTc-HYNIC-DAPI-treated samples, indicating that the plasmid damage was caused by direct interactions between the radioactive compound and the DNA.
Figure 6. Number of DSBs and SSBs per DNA molecule.

Analysis of the mean number of DSBs (A) and SSBs (B) per plasmid molecule as a function of 99mTc-HYNIC-DAPI decays during a 2 h irradiation. The numbers of DSBs and SSBs were calculated based on the fluorescence intensity of the linear and supercoiled DNA fractions in the presence (□) or absence (▪) of DMSO.
Estimation of plasmid-bound activity
After quantifying the DNA damage by ethidium bromide staining and image processing, the DNA bands representing each plasmid conformation (supercoiled, open circular and linear) were excised, and the DNA-bound activity in the 99mTc-pertechnetate and 99mTc-HYNIC-DAPI samples was measured using a gamma counter. The bound activity was quantitated using decay-corrected values. 99mTc-pertechnetate-irradiated plasmid DNA exhibited a low amount of bound activity (<0.005 MBq) that was independent of the administered activity. In contrast, 99mTc-HYNIC-DAPI demonstrated a correlation between applied activity and DNA-bound activity, reaching a maximum value of 0.72 MBq in the sample dosed with 1000 MBq/mL (Figure 7). When plotting the bound activity against the fractions of open circular DNA or linear DNA (Figure 8), linear correlations were apparent. These correlations indicated that increased plasmid damage resulted from increased 99mTc-HYNIC-DAPI accumulation.
Figure 7. Measurement of the Plasmid DNA-bound radioactivity.
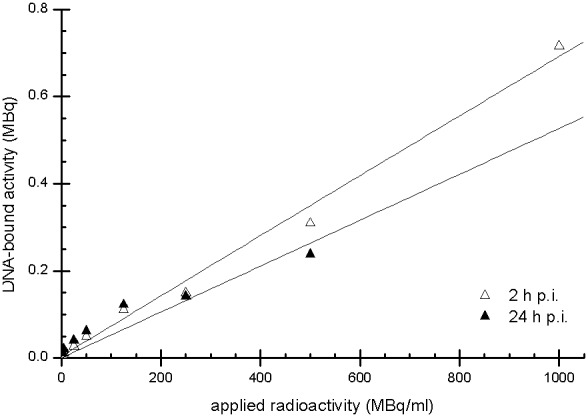
The measurements were performed using a gamma counter, and the activity was decay-corrected based on the application time. The data were obtained from a single experiment. A good linear correlation was observed between the applied activity and the bound activity.
Figure 8. Fluorescence intensity with respect to DNA-bound activity.
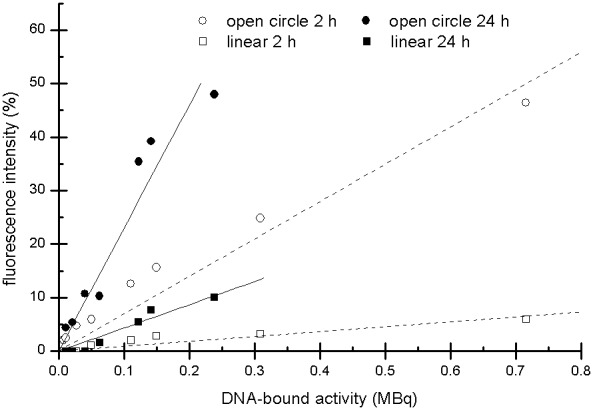
Dependence of the relative fluorescent intensity of open circle and linear plasmid DNA on the DNA-bound activity. A good correlation was observed between the activity and the radiation damage. The higher fluorescent intensity after the 24 h incubation time was due to the increased number of radioactive decays.
Discussion
DAPI is a DNA-specific probe that forms a fluorescent complex by inserting into the minor groove of AT-rich DNA sequences [15]. The HYNIC linker is widely used for labeling bioactive substances with 99mTc without a loss of avidity [19]. We found that unlabeled HYNIC-DAPI was not chemotoxic at the tested low concentrations (data not shown).
The plasmids were incubated with 99mTc-HYNIC-DAPI in excess of unlabeled DAPI, which can be explained by stoichiometric calculations. Radiochemistry labeling was performed using 1500 MBq 99mTc and 5 µg HYNIC-DAPI. HYNIC-DAPI has a molar mass of 943 g/mol, which corresponds to a ratio of 4.7*1013 atoms of 99mTc to 3.2*1015 molecules of HYNIC-DAPI. Assuming 100% turnover, the reaction would yield 1 labeled 99mTc-HYNIC-DAPI molecule per 67 non-radioactive HYNIC-DAPI molecules. Here, the term non-radioactive refers to HYNIC-DAPI that was unlabeled or that was labeled with 99Tc, which is considered non-radioactive because of its long half-life (2.1*105 years). Two hundred nanograms of pUC19 plasmid DNA contain 6.9*1010 plasmid molecules. When incubating 1 MBq 99mTc-HYNIC-DAPI with 200 ng of pUC19 plasmid DNA, 0.45 99mTc-HYNIC-DAPI molecules would bind to each plasmid molecule on average.
99mTc-pertechnetate bound to plasmid DNA to a much lesser extent (by approximately 100-fold less, data not shown), suggesting that the binding of 99mTc-HYNIC-DAPI to DNA was a specific event. DAPI molecules bind to double-stranded DNA in the minor groove. The distance between two minor grooves is 10 base pairs; therefore, we assumed that there were 270 binding sites per plasmid for 99mTc-HYNIC-DAPI. Considering the competition by non-radioactive HYNIC-DAPI, a maximum of 4 99mTc-HYNIC-DAPI molecules could bind to each pUC19 plasmid DNA on average. For our labeling procedure, 4 99mTc-HYNIC-DAPI molecules per plasmid corresponded to the application of 10 MBq (500 MBq/mL) of 99mTc-HYNIC-DAPI. However, our data did not demonstrate the saturation of plasmid damage at higher activities (Figure 3), indicating that not every available HYNIC-DAPI molecule bound to plasmid DNA.
The DNA damage caused by unbound 99mTc-HYNIC-DAPI was expected to be comparable to that caused by 99mTc-pertechnetate. In total, 1000 MBq/mL of unbound, but homogeneously distributed, 99mTc in a volume of 10 µL leads to an absorbed dose of 74 Gy within 24 h. According to Table 2, this dose produced 0.5 SSBs and 0.02 DSBs on average in the absence of DMSO (24 h, 1000 MBq/mL, 99mTc-pertechnetate). In the presence of DMSO, the formation of SSBs and DSBs was completely prevented (0.03 SSBs, no DSBs). These data suggested that the DNA damage was radical mediated and an indirect effect of unbound 99mTc.
After 2 h of irradiation, 1000 MBq/mL 99mTc-HYNIC-DAPI produced 60% more (from 0.38 to 0.60) SSBs and 300% more (from 0.013 to 0.056) DSBs compared with unbound 99mTc-pertechnetate. These increases resulted from approximately 0.7 MBq DNA-bound radioactivity (Figure 7).
Within 24 h, 500 MBq/mL of unbound 99mTc (Figure 3) caused 26% of the DNA to adopt an open circular configuration. The same percentage was induced by 125 MBq/mL of 99mTc-HYNIC-DAPI, which corresponds to a 4-fold enhancement. The potential for plasmid linearization was approximately ten times higher for 99mTc-HYNIC-DAPI because 50 MBq/mL yielded 2% linear plasmid, whereas 500 MBq/ml of 99mTc-pertechnetate was required to elicit the same effect. A higher linear plasmid fraction could not be induced by 99mTc-pertechnetate because of limited specific activity.
We did not perform X-ray irradiation; hence, we could not determine the relative biological effectiveness of 99mTc-HYNIC-DAPI and 99mTcO4 −. Nevertheless, we compared the biological effectiveness of each compound, which we defined as the ratio of the activities of 99mTc-HYNIC-DAPI versus 99mTcO4 − that caused the same effect. Based on this definition, the biological effectiveness increased by approximately 4-fold in inducing SSBs and by approximately 10-fold in inducing DSBs, which is in the range reported for Auger electrons emitted in close proximity to DNA [20].
The SSBs created by unbound 99mTc-pertechnetate could be prevented by DMSO. In contrast, DSBs and SSBs caused by 99mTc-HYNIC-DAPI could not be prevented by DMSO. This indicated that the impact of the Auger events was direct or indirect with an extremely high local energy transfer that could not be antagonized by DMSO. SSBs are usually assumed to be radical-mediated; however, we demonstrated that the induction of SSBs by DNA-bound 99mTc-HYNIC-DAPI might occur via another mechanism. It is possible that SSBs were also caused by direct interactions between emitted low-energy electrons and plasmid-bound 99mTc-HYNIC-DAPI that merely damaged a single strand. On average, 4.9 electrons are emitted by the decay of 99mTc via internal conversion and gamma emission. This event creates a dramatic charge transfer in the electron shell, which can cause a Coulomb explosion of the molecule, as discussed by Pomplun and Sutmann [21].
The absorbed dose was calculated based on the absorbed energy inside an incubation volume of 10 µL. Thus, no regional heterogeneities, such as DNA-bound activity, were considered. Therefore, the same amount of 99mTc yielded the same absorbed dose regardless of whether 99mTc-pertechnetate or 99mTc-HYNIC-DAPI was used. We expected that the dose to the plasmids was higher with 99mTc-HYNIC-DAPI due to the emission of low-energy electrons. 99mTc emits an average of 4.9 electrons with less than 2.7 keV of energy and a maximum range of 300 nm [17]. However, the term ‘absorbed dose’ is a macroscopic quantity that does not account for stochastic effects. Hence, we did not calculate the absorbed dose on plasmid level because 99mTc-HYNIC-DAPI binding is a random process, as are radioactive decay and the direction of radiation emission.
The ratio of SSBs to DSBs induced by irradiation is approximately 40∶1 per gray of gamma radiation [22]; this ratio decreases with increasing dose because SSBs are converted to DSBs. High LET irradiation induces more DSBs because the high density of radicals increases the possibility of DSBs being generated by chance. We calculated the SSB and DSB yields based on the conversion of supercoiled plasmid DNA into linear DNA. The numbers of SSBs and DSBs per plasmid molecule based on the number of decays over 2 h are plotted in Figure 6. As expected, there was a linear correlation between the number of decays and the number of SSBs and DSBs per plasmid molecule. DNA-bound 99mTc-HYNIC-DAPI produced DNA damage with a SSB:DSB ratio of 10∶1 in the absence of DMSO. Furthermore, 0.64*1013 decays/mL induced 0.06 DSBs and 0.68 SSBs per molecule, which corresponded with 1 DSB per 10.2*1013 decays/mL. Balagurumoorthy et al. reported values between 5.57*1013 and 29.6*1013 decays/mL for various 125I compounds in the absence of DMSO [2]. The yield of DSBs per decay of 99mTc was calculated as the product of the plasmid concentration and the number of DSBs per decay/mL. Although Balagurumoorthy obtained DSB yields ranging from 0.10 to 0.52 per decay of 125I, our data demonstrated a DSB yield of 0.03 per decay of 99mTc. Therefore, our observed number of decays/mL corresponded with the data published by Balagurumoorthy, although the DSB yield for 99mTc appeared to be a tenth of that for 125I because of the lower plasmid amount. We irradiated 200 ng of pUC19 plasmid DNA in a volume of 20 µL, which corresponds to 3.44*1012 plasmid molecules/mL, whereas Balagurumoorthy used 3.06*1013 molecules/mL.
A more reliable calculation of the DSB yield per plasmid-bound decay requires knowledge of the amount of plasmid-bound radioactivity. Furthermore, the radiochemical stability of 99mTc-HYNIC-DAPI will introduce variability in the number of decays per plasmid. The plasmid-bound applied activity varied between 2% and 10% in different experiments. Assuming 5% plasmid-bound applied activity, we calculated 0.5 to 1.5 DSBs per plasmid-bound decay. Humm and Charlton calculated the probability of DSB formation due to DNA-bound Auger electron emission for 125I, 123I and 99mTc to be 1.1, 0.73 and 0.43, respectively [23].
Several studies have characterized the influence of the distance between the decaying isotope and DNA on DSB yield. Balagurumoorthy et al. synthesized a series of 125I-labeled Hoechst derivatives with a minor groove-binding motif and an increasing distance between the radioactive molecule and the binding position (ranging from 10.5 Å to 13.9 Å) [2]. The DSB yield decreased with distance from 0.2 DSBs/decay to 0.025 DSBs/decay, demonstrating the crucial influence of relatively small distances. Although DMSO could not prevent the formation of DSBs at the closest distance, it reduced DSBs by 10-fold at greater distances. Our data did not indicate an effect of DMSO on DNA-bound 99mTc-HYNIC-DAPI. Therefore, we assumed that the radioactive molecule was below the critical distance from the DNA helix, even if stereoisomers could exist.
Walicka et al. investigated the effect of different DMSO concentrations (0.26 to 3 M) on the radiotoxicity of the Auger electron emitter 125I [24]. The authors concluded, that the most radiotoxic effects from DNA-incorporated 125I were due to indirect mechanisms. In contrast to our experiments Walicka et al. performed experiments using V79 cells, whereas we used plasmids as biological model without any repair or radioprotection by other cellular structures. Additionally, the authors irradiated their cells at −135°C what could be cryotoxic to cells. Balagurumoorthy et al. found that in the absence of DMSO, 125IEH-induced DSBs in nicked or linear DNA were caused by both direct and indirect mechanisms, whereas in its presence, DSBs occurred predominantly by direct ionization of DNA [25].
Conclusion
The direct binding of 99mTc-HYNIC-DAPI to plasmid DNA was demonstrated and quantified. Plasmid-bound 99mTc-HYNIC-DAPI induced SSBs and DSBs with high efficiency. DNA damage could not be prevented by the radical scavenger DMSO, suggesting that the damage was a direct effect. This statement is true for both DSBs and SSBs. The biological effectiveness increased by approximately 4-fold for inducing SSBs and approximately 10-fold for inducing DSBs, which was within the range reported for Auger electrons emitted in close proximity to DNA.
Acknowledgments
The authors would like to thank Prof. L. E. Feinendegen for encouraging our studies and for helpful discussion.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
The research (JK, GW) was partly supported by the Deutsche Forschungsgemeinschaft (DFG, http://www.dfg.de/, Grant No. KO 1695/4-1). Sandra Ferl (SF) was supported by the Europäischer Sozialfonds (ESF, http://www.esf.de/portal/generator/8/startseite.html, grant code 080951832). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Adelstein SJ, Kassis AI (1996) Strand breaks in plasmid DNA following positional changes of Auger-electron-emitting radionuclides. Acta Oncol 35: 797–801. [DOI] [PubMed] [Google Scholar]
- 2. Balagurumoorthy P, Xu X, Wang K, Adelstein SJ, Kassis AI (2012) Effect of distance between decaying (125)I and DNA on Auger-electron induced double-strand break yield. Int J Radiat Biol 88: 998–1008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Pomplun E, Terrissol M, Hille R (2002) Ratio of complex double strand break damage induced by 125IUdR and 123IUdR correlates with experimental in vitro cell killing effectiveness. Radiat Prot Dosimetry 99: 81–82. [DOI] [PubMed] [Google Scholar]
- 4. Haefliger P, Agorastos N, Spingler B, Georgiev O, Viola G, et al. (2005) Induction of DNA-double-strand breaks by auger electrons from 99mTc complexes with DNA-binding ligands. Chembiochem 6: 414–421. [DOI] [PubMed] [Google Scholar]
- 5. Santos-Cuevas CL, Ferro-Flores G, Rojas-Calderon EL, Garcia-Becerra R, Ordaz-Rosado D, et al. (2011) 99mTc-N2S2-Tat (49–57)-bombesin internalized in nuclei of prostate and breast cancer cells: kinetics, dosimetry and effect on cellular proliferation. Nucl Med Commun 32: 303–313. [DOI] [PubMed] [Google Scholar]
- 6. Schipper ML, Riese CG, Seitz S, Weber A, Behe M, et al. (2007) Efficacy of 99mTc pertechnetate and 131I radioisotope therapy in sodium/iodide symporter (NIS)-expressing neuroendocrine tumors in vivo. Eur J Nucl Med Mol Imaging 34: 638–650. [DOI] [PubMed] [Google Scholar]
- 7. Kriehuber R, Kadenbach K, Schultz F, Weiss DG (2004) Study on cell survival, induction of apoptosis and micronucleus formation in SCL-II cells after exposure to the auger electron emitter (99m)Tc. Int J Radiat Biol 80: 875–880. [DOI] [PubMed] [Google Scholar]
- 8. Kotzerke J, Wendisch M, Freudenberg R, Runge R, Oehme L, et al. (2012) Sodium-iodide symporter positive cells after intracellular uptake of (99m)Tc versus alpha-emitter 211At. Reduction of clonogenic survival and characterization of DNA damage. Nuklearmedizin 51: 170–178. [DOI] [PubMed] [Google Scholar]
- 9. Wendisch M, Freudenberg R, Drechsel J, Runge R, Wunderlich G, et al. (2010) [99mTc reduces clonogenic survival after intracellular uptake in NIS-positive cells in vitro more than 131I]. Nuklearmedizin 49: 154–160. [DOI] [PubMed] [Google Scholar]
- 10. Freudenberg R, Wendisch M, Runge R, Wunderlich G, Kotzerke J (2012) Reduction in clonogenic survival of sodium-iodide symporter (NIS)-positive cells following intracellular uptake of (99m)Tc versus (188)Re. Int J Radiat Biol 88: 991–997. [DOI] [PubMed] [Google Scholar]
- 11. Wunderlich G, Wendisch M, Aurich D, Runge R, Freudenberg R, et al. (2012) Preincubation with Sn-complexes causes intensive intracellular retention of (99m)Tc in thyroid cells in vitro. Nuklearmedizin 51: 179–185. [DOI] [PubMed] [Google Scholar]
- 12. Cambien B, Franken PR, Lamit A, Mauxion T, Richard-Fiardo P, et al. (2014) 99mTcO4–, Auger-Mediated Thyroid Stunning: Dosimetric Requirements and Associated Molecular Events. PLoS One 9: e92729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Balagurumoorthy P, Wang K, Adelstein SJ, Kassis AI (2008) DNA double-strand breaks induced by decay of (123)I-labeled Hoechst 33342: role of DNA topology. Int J Radiat Biol 84: 976–983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Yasui LS, Chen K, Wang K, Jones TP, Caldwell J, et al. (2007) Using Hoechst 33342 to target radioactivity to the cell nucleus. Radiat Res 167: 167–175. [DOI] [PubMed] [Google Scholar]
- 15. Tanious FA, Veal JM, Buczak H, Ratmeyer LS, Wilson WD (1992) DAPI (4′,6-diamidino-2-phenylindole) binds differently to DNA and RNA: minor-groove binding at AT sites and intercalation at AU sites. Biochemistry 31: 3103–3112. [DOI] [PubMed] [Google Scholar]
- 16. Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, et al. (2012) Fiji: an open-source platform for biological-image analysis. Nat Methods 9: 676–682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Howell RW (1992) Radiation spectra for Auger-electron emitting radionuclides: report No. 2 of AAPM Nuclear Medicine Task Group No. 6. Med Phys 19: 1371–1383. [DOI] [PubMed] [Google Scholar]
- 18. Cowan R, Collis CM, Grigg GW (1987) Breakage of double-stranded DNA due to single-stranded nicking. J Theor Biol 127: 229–245. [DOI] [PubMed] [Google Scholar]
- 19. Babich JW, Solomon H, Pike MC, Kroon D, Graham W, et al. (1993) Technetium-99m-labeled hydrazino nicotinamide derivatized chemotactic peptide analogs for imaging focal sites of bacterial infection. J Nucl Med 34: 1964–1974. [PubMed] [Google Scholar]
- 20. Kassis AI, Fayad F, Kinsey BM, Sastry KS, Adelstein SJ (1989) Radiotoxicity of an 125I-labeled DNA intercalator in mammalian cells. Radiat Res 118: 283–294. [PubMed] [Google Scholar]
- 21. Pomplun E, Sutmann G (2004) Is coulomb explosion a damaging mechanism for (125)IUdR? Int J Radiat Biol 80: 855–860. [DOI] [PubMed] [Google Scholar]
- 22. Kassis AI, Harapanhalli RS, Adelstein SJ (1999) Strand breaks in plasmid DNA after positional changes of Auger electron-emitting iodine-125: direct compared to indirect effects. Radiat Res 152: 530–538. [PubMed] [Google Scholar]
- 23. Humm JL, Charlton DE (1989) A new calculational method to assess the therapeutic potential of Auger electron emission. Int J Radiat Oncol Biol Phys 17: 351–360. [DOI] [PubMed] [Google Scholar]
- 24. Walicka MA, Ding Y, Adelstein SJ, Kassiss AI (2000) Toxicity of DNA-incorporated iodine-125: quantifying the direct and indirect effects. Radiat Res 154: 326–330. [DOI] [PubMed] [Google Scholar]
- 25. Balagurumoorthy P, Chen K, Adelstein SJ, Kassis AI (2008) Auger electron-induced double-strand breaks depend on DNA topology. Radiat Res 170: 70–82. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.