

Abstract
AIM: To investigate whether activin regulates the cell proliferation of human gastric cancer cell line SNU-16 through the mRNA changes in activin receptors, Smads and p21CIP1/WAF1.
METHODS: The human gastric cancer cell lines were cultured, RNAs were purified, and RT-PCRs were carried out with specifically designed primer for each gene. Among them, the two cell lines SNU-5 and SNU-16 were cultured with activin A for 24, 48 and 72 h. The cell proliferation was measured by MTT assay. For SNU-16, changes in ActRIA, ActRIB, ActRIIA, ActRIIB, Smad2, Smad4, Smad7, and p21CIP1/WAF1 mRNAs were detected with RT-PCR after the cells were cultured with activin A for 24, 48 and 72 h.
RESULTS: The proliferation of SNU-16 cells was down regulated by activin A whereas other cells showed no change. Basal level of inhibin/activin subunits, activin receptors, Smads, and p21CIP1/WAF1 except for activin βB mRNAs was observed to have differential expression patterns in the human gastric cancer cell lines, AGS, KATO III, SNU-1, SNU-5, SNU-16, SNU-484, SNU-601, SNU-638, SNU-668, and SNU-719. Interestingly, significantly higher expressions of ActR IIA and IIB mRNAs were observed in SNU-16 cells when compared to other cells. After activin treatment, ActR IA, IB, and IIA mRNA levels were decreased whereas ActR IIB mRNA level increased in SNU-16 cells. Smad4 mRNA increased for up to 48 h whereas Smad7 mRNA increased sharply at 24 h and returned to the initial level at 48 h in SNU-16 cells. In addition, expression of the p21CIP1/WAF1, the mitotic inhibitor, peaked at 72 h after activin treatment in SNU-16 cells.
CONCLUSION: Our results suggest that inhibition of cell growth by activin is regulated by the negative feedback effect of Smad7 on the activin signaling pathway, and is mediated through p21CIP1/WAF1 activation in SNU-16 cells.
Keywords: Human gastric cancer cell lines, Activin A, Cell proliferation, Activin receptors, Smads, p21CIP1/WAF1
INTRODUCTION
Inhibins and activins, members of the transforming growth factor-β (TGF-β) superfamily, are polypeptides that were originally isolated from ovarian fluid, based on their effect on pituitary follicle-stimulating hormone (FSH) production. Inhibins are heterodimers that are composed of a common α subunit and one of the two homologous β subunits (βA and βB). Activins are either heterodimers or homodimers of inhibin β subunits (βAβA, βBβB, and βAβB)[1-3].
Activin plays an important role in the proliferation, differentiation, and apoptosis of target cells. The biological activity of activin is mediated by receptor complexes consisting of two different types of receptors, the type I (ActR I) and type II (ActR II) activin serine/threoine kinase receptors[4]. ActR II binds to activin independently of ActR I, but is unable to signal without ActR I. The formation of heteromeric complexes of ActR I and ActR II is required for the mediation of the cellular signal[5]. ActR I and ActR II mRNAs have been widely detected in several human tissues, including prostate cancer, breast cancer, and epithelial ovarian cancer[6-8].
Smad proteins have been known as intracellular signaling mediators of the activin[9,10]. Smad1, Smad2, Smad3, and Smad5 proteins are phosphorylated by activated specific type I serine/threonine kinase receptors, and thus act in a pathway-restricted fashion. Smad4 protein forms hetero-oligomeric complexes with pathway-restricted Smad proteins, which translocate into the nuclei and activate transcriptional responses. Smad6 and Smad7 function as inhibitors of TGF-β family signaling, including activin[11-13]. Smad7 selectively interferes with different activin signaling pathways and inhibits erythroid leukemia cells and liver Hep3B cells by blocking intracellular activin signaling[14,15]. In addition, the mitotic inhibitor p21CIP1/WAF1 thas suppresses the growth of malignant cells in vitro and in vivo can be up-regulated by activin[16-18].
This study was to investigate whether activin can affect cell proliferation in human gastric cancer cell line SNU-16 through the changes in activin receptors, Smads and p21CIP1/WAF1 mRNA levels.
MATERIALS AND METHODS
Cell culture and reagents
Human gastric cancer cell lines AGS and KATO III were purchased from the American Tissue Culture Collection (ATCC, Rockville, MD). SNU-1, SNU-5, SNU-16, SNU-484, SNU-601, SNU-638, SNU-668, and SNU-719 cells were supplied from the Korean Cell Line Bank. Cells were cultured in RPMI 1640 medium (GIBCO, Grand Island, NY) containing 10% fetal bovine serum (FBS, GIBCO), 100 U/mL penicillin and 100 μg/mL streptomycin at 37 °C in a humidified 50 mL/L CO2 and 95% air. The appropriate amount of recombinant human activin A (activin A, R & D systems, Minneapolis, MN) was added to the culture media.
MTT assay
Cell proliferation was measured with CellTiter 96 Aqueous One Solution (Promega, Madison, WI). Cells were seeded at 5 × 103 cells/well in 6-well plates and incubated with different concentrations of activin A (0, 10, 50, and 100 ng/mL) at 37 °C for 24, 48, and 72 h respectively. Cell viability was determined by a colorimetric assay with PMS/MTS (20 μL/well). The absorbance was determined at 492 nm with background subtraction at 650 nm.
Cell number assay
For the cell viability assay, cells were seeded at 5 × 104 cells/mL in 6-well plates. The cells were cultured with 100 ng/mL of activin A for 24, 48, and 72 h respectively. Then cells were washed with phosphate buffered saline (PBS) and resuspended in PBS. Viable cells were counted by the trypan blue exclusion method at each time using a hemocytometer.
RNA extraction and RT-PCR procedures
Total RNA was extracted from cultured cells using the RNAzol B solution kit following the manufacturer's protocol (Tel-test, Friendswood, Texas). First-strand cDNA synthesis was performed using a cDNA synthesis kit (Roche, Mannheim, Germany). cDNA synthesis was performed by reverse transcription in a total volume of 20 μL reaction mixture containing 1 μg RNA, 1× reaction buffer, 1 mmol/L dNTP, 5 μmol/L oligodT, 20 units of RNase inhibitor, and 20 units of AMV reverse transcriptase. The reaction mixture was incubated at 42 °C for 1 h, terminated by heating at 95 °C for 5 min. Polymerase chain reaction (PCR) was performed with 5 μL (α, βA, βB subunits, and p21CIP1/WAF1) or 1 μL (activin receptors, Smads and β-actin) cDNA in a 50 μL reaction mixture of 1× PCR buffer, 0.2 mmol/L of each dNTP, 20 pmoL primer, and 1 unit of Taq DNA polymerase (Roche). Primer sequences are shown in Table 1. The conditions for amplification were as follows: first denaturation at 95 °C for 4 min, then denaturation at 95 °C for 30 s, annealing at 50 °C to 62 °C for 30 s, and extension at 72 °C for 1 min for 30 cycles (25 cycles for β-actin) and a final extension at 72 °C for 7 min. Ten μL of PCR products was separated by electrophoresis on a 2% agarose gel containing ethidium bromide and visualized by image analysis (Gel Doc 1000 gel document system, Bio-Rad, Hercules, CA). The intensities of PCR bands were measured by densitometry (Model GS-700, Bio-Rad) and expressed relative to β-actin.
Table 1.
Oligonucleotide sequences of PCR primers
| Oligo | Sequence | Annealing temperature (°C) | Size (bp) |
| Inhibin α | 5'-AGGAAGAGGAGGATGTCTCC-3' | 50 | 823 |
| 5'-GAGTAACCTCCATCCGAGGT-3' | |||
| Activin βA | 5'-CTGAACGCGATCAGAAAGCT-3' | 54 | 1014 |
| 5'-TCCTCCACGATCATGTTCTG-3' | |||
| Activin βB | 5'-TTCGCCGAGACAGATGGCCT-3' | 59 | 765 |
| 5'-AATCATGTTGGGCACGTCCC-3' | |||
| ActR IA | 5'-GCCCAAGGTCAACCCCAAACTCT-3' | 60 | 265 |
| 5'-GGATTTTCCTTTAGTGGGCAGCT-3' | |||
| ActR IB | 5'-ACCAGCTGCCTCCAGGCCAACTA-3' | 62 | 245 |
| 5'-GTGCTCAGGCTCCTTGAGGTGAC-3' | |||
| ActR IIA | 5'-ACCAGTGTTGATGTGGATCTT-3' | 60 | 456 |
| 5'-TACAGGTCCATCTGCAGCAGT-3' | |||
| ActR IIB | 5'-CATGGAACGAACTGTGTCAT-3' | 62 | 730 |
| 5'-AGATCCACTGAGTCTGGAGA-3' | |||
| Smad2 | 5'-TAGGTGGGGAAGTTTTTGCT-3' | 60 | 410 |
| 5'-TTTGCATGGGACTTGATTGG-3' | |||
| Smad4 | 5'-AAGGTGAAGGTGATGTTTG-3' | 60 | 509 |
| 5'-TAAGGGCCCCAACGGTAAA-3' | |||
| Smad7 | 5'-GTGGGGAGGCTCTACTGTGTC-3' | 58 | 477 |
| 5'-GGCTACCGGCTGTTGAAGATG-3' | |||
| p21CIP1/WAF1 | 5’-CCTCTTCGGCCCAGTGGAC-3’ | 60 | 369 |
| 5'-CCGTTTTCGACCCTGAGAG-3' | |||
| β-actin | 5'-TACCTCATGAAGATCCTCA-3 | 50 | 267 |
| 5'-TTCGTGGATGCCACAGGAC-3' |
ActR: activin receptor. ActR: activin receptor.
Statistical analysis
Values were expressed as mean ± SD. Student’s t test was used to evaluate differences between control and activin A-treated samples. aP < 0.05 and bP < 0.01 were considered statistically significant.
RESULTS
Effects of activin A on cell proliferation
Ten human gastric cancer cell lines were treated with various concentrations of activin A (10-100 ng/mL) for 24, 48, and 72 h respectively. The effect of activin on cell proliferation was assessed by MTT assay (Table 2). Activin induced a significant decrease in SNU-16 cell proliferation in a dose- and time-dependent manner, whereas other cells showed no detectable response. To evaluate the growth inhibition of activin in SNU-5 and SNU-16 cells, the cells were cultured with 100 ng/mL of activin A for 24, 48, and 72 h respectively. The cell number was counted by a hemocytometer. Activin A treatment resulted in a significant decrease in the number of SNU-16 cells in a time-dependent manner, whereas no effect was observed in the number of SNU-5 cells (Figure 1).
Table 2.
Effects of activin A on cell proliferation in human gastric cancer cell lines (mean ± SD, ng/mL)
| Times |
24 h |
48 h |
72 h |
|||||||||
| Activin A | 0 | 10 | 50 | 100 ng/mL | 0 | 10 | 50 | 100 | 0 | 10 | 50 | 100 |
| AGS | 100 | 92.7 ± 5.1 | 93.6 ± 4.3 | 93.6 ± 4.2 | 100 | 93.6 ± 2.9 | 94.1 ± 3.1 | 92.9 ± 3.9 | 100 | 94.0 ± 3.9 | 94.2 ± 3.7 | 91.7 ± 3.8 |
| KATO III | 100 | 94.2 ± 3.3 | 92.3 ± 4.6 | 92.1 ± 4.8 | 100 | 93.1 ± 3.6 | 89.8 ± 4.2 | 89.9 ± 3.7 | 100 | 91.1 ± 4.0 | 85.6 ± 4.2 | 84.1 ± 6.8 |
| SNU-1 | 100 | 94.2 ± 2.6 | 94.0 ± 2.8 | 94.5 ± 2.9 | 100 | 94.6 ± 2.0 | 93.4 ± 5.7 | 94.7 ± 3.3 | 100 | 94.0 ± 3.5 | 94.0 ± 2.8 | 93.9 ± 4.1 |
| SNU-5 | 100 | 93.9 ± 0.6 | 94.7 ± 2.4 | 94.1 ± 1.8 | 100 | 94.1 ± 3.3 | 94.2 ± 5.6 | 94.8 ± 4.0 | 100 | 94.7 ± 2.6 | 94.0 ± 4.3 | 94.8 ± 4.0 |
| SNU-16 | 100 | 87.3 ± 2.3a | 81.7 ± 3.3a | 77.2 ± 4.5a | 100 | 87.8 ± 2.6a | 74.3 ± 3.4b | 61.5 ± 2.9b | 100 | 81.2 ± 4.6a | 62.2 ± 4.5b | 45.2 ± 4.0b |
| SNU-484 | 100 | 94.3 ± 3.9 | 94.0 ± 3.0 | 92.1 ± 2.9 | 100 | 94.9 ± 1.5 | 92.6 ± 3.6 | 94.8 ± 1.8 | 100 | 93.4 ± 4.8 | 92.5 ± 5.5 | 91.8 ± 6.8 |
| SNU-601 | 100 | 94.2 ± 2.0 | 94.2 ± 1.8 | 91.6 ± 2.3 | 100 | 94.6 ± 3.4 | 94.5 ± 2.1 | 94.0 ± 2.9 | 100 | 94.9 ± 4.7 | 91.9 ± 6.1 | 91.3 ± 6.7 |
| SNU-638 | 100 | 91.1 ± 2.6 | 92.5 ± 1.1 | 91.5 ± 2.5 | 100 | 93.3 ± 3.7 | 92.6 ± 4.2 | 91.8 ± 4.2 | 100 | 93.9 ± 3.3 | 92.0 ± 3.8 | 91.3 ± 4.1 |
| SNU-668 | 100 | 94.2 ± 2.8 | 94.3 ± 2.3 | 94.4 ± 3.1 | 100 | 91.7 ± 1.4 | 91.8 ± 1.6 | 92.3 ± 4.8 | 100 | 93.2 ± 1.5 | 94.0 ± 2.2 | 94.9 ± 2.3 |
| SNU-719 | 100 | 92.8 ± 2.8 | 91.6 ± 4.0 | 91.4 ± 1.2 | 100 | 91.7 ± 2.2 | 92.4 ± 5.2 | 91.7 ± 3.4 | 100 | 91.8 ± 2.0 | 88.7 ± 2.8 | 89.2 ± 3.1 |
Values are the percentage for three individual experiments, each with triplicate samples.
P < 0.05 and
P < 0.01 vs the control values.
Figure 1.
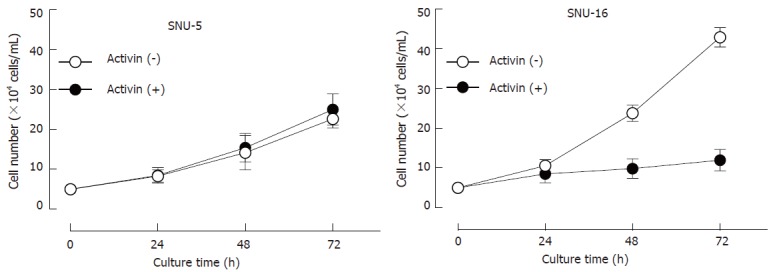
Effects of activin A on cell viability in SNU-5 and SNU-16 cells. Cells (5 × 104 cells/mL) were cultured with activin A (100 ng/mL) in a time-dependent manner and viable cells were counted with a hemocytometer. Values are the mean ± SD of three individual experiments. bP < 0.01 vs the control values.
Expression of inhibin/activin subunit, activin receptor, Smads, and p21CIP1/WAF1 mRNAs
Basal expressions of inhibin/activin subunits, activin receptors, Smads, and p21CIP1/WAF1 mRNA in human gastric cancer cell lines were investigated by RT-PCR (Figure 2). The expression of inhibin α mRNA was detectable only in SNU-638 cells whereas the activin βA mRNA was expressed in the majority of cells tested, but not in SNU-5 or SNU-719 cells. The activin βB was not detected in all cells. The activin receptor IA and IB mRNAs were expressed in all the cell lines tested. The primers for ActR IIA and IIB amplification were designed to locate within intracellular kinase domains of ActR IIA and IIB. The ActR IIA and IIB mRNAs were expressed in all the cell lines tested. Interestingly, significantly higher expressions of ActR IIA and IIB mRNAs were observed in SNU-16 cells when compared to other cells. The Smad2, 4, and 7 mRNAs were expressed in all cells and p21CIP1/WAF1 mRNAs were not expressed in SNU-1, SNU-484, and SNU-668 cells.
Figure 2.
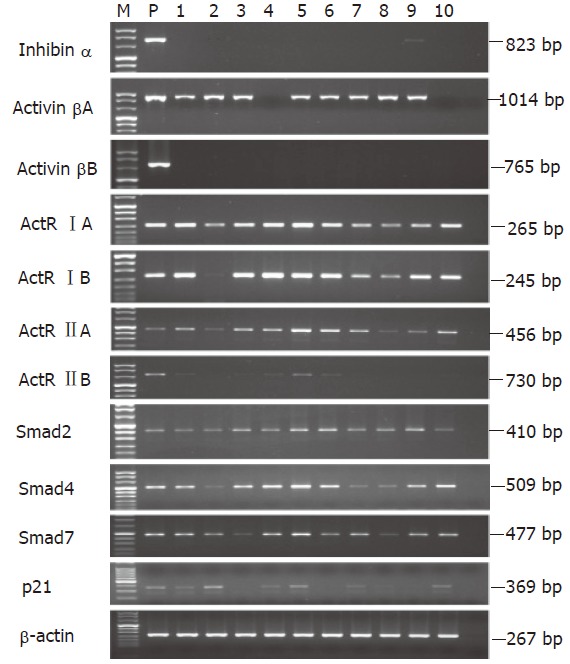
mRNA expressions of inhibin/activin subunits, activin receptors, Smads, and p21CIP1/WAF1 in human gastric cancer cell lines. The mRNA levels were analyzed by RT-PCR. M: DNA size marker; P: positive control; lane 1: AGS; lane 2: KATO III; lane 3: SNU-1; lane 4: SNU-5; lane 5: SNU-16; lane 6: SNU-484; lane 7: SNU-601; lane 8: SNU-638; lane 9: SNU-668; lane 10: SNU-719. Positive control was used for inhibin/activin subunits (mouse ovary), activin receptor (K562), Smad and p21CIP1/WAF1 (human keratinocytes) respectively.
Effects of activin A on expression of activin receptors, Smad and p21CIP1/WAF1 genes
To detect the effects of activin on ActR I and ActR II mRNA expression in SNU-16 cells, RT-PCR analysis was performed using the cells treated with 100 ng/mL of activin A for various durations (Figure 3A). Expression of ActR IA, IB, and IIA mRNA was decreased after activin A treatment, whereas ActR IIB mRNA level was increased significantly for up to 48 h in SNU-16 cells. To detect the effects of activin-signaling on Smad mRNA expression in SNU-16 cells, RT-PCR analysis was investigated using the cells treated with 100 ng/mL of activin A for various durations (Figure 3B). Although no detectable change was observed in Smad2 mRNA of SNU-16 cells, the expression of Smad4 mRNA was increased at 48 h after activin A treatment. Expression of Smad7 mRNA was increased vigorously at 24 h after activin A treatment in SNU-16 cells and decreased significantly to below initial basal level at 48 h when compared to control cells. In addition, to determine the regulation of p21CIP1/WAF1 mRNA expression by activin in SNU-16 cells, the cells were treated with 100 ng/mL of activin A for various durations and investigated by RT-PCR (Figure 3C). Expression of p21CIP1/WAF1 mRNA was highly increased at 72 h after activin A stimulation in SNU-16 cells.
Figure 3.
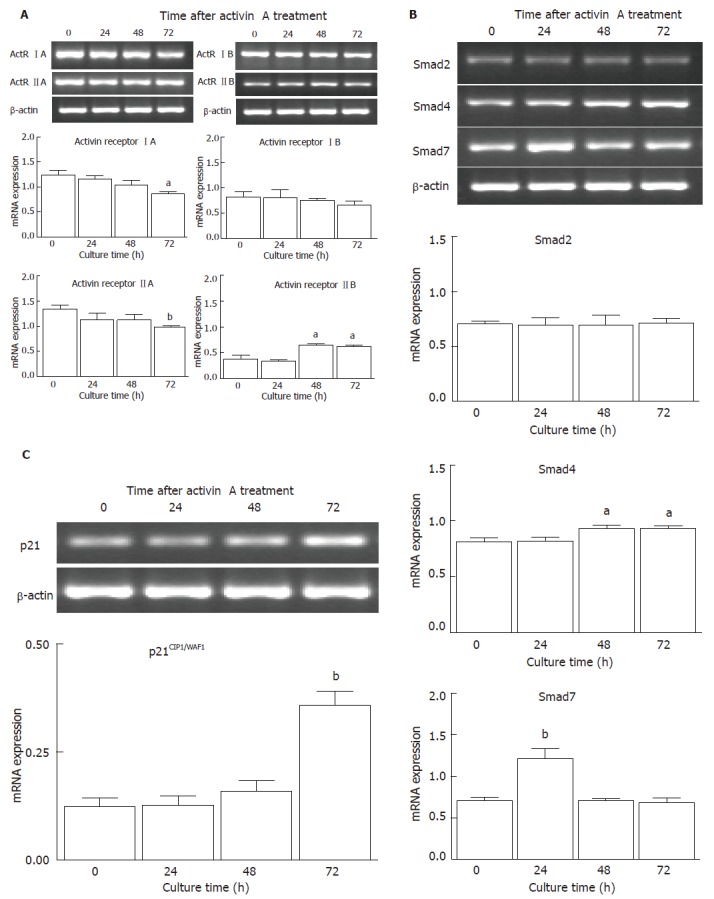
Effects of activin A on activin receptor mRNA expression (A), smad mRNA expression (B), and p21CIP1/WAF1 mRNA expression (C) in SNU-16 cells. Cells (5 × 104 cells/mL) were cultured with activin A (100 ng/mL) in a time-dependent manner and mRNA levels were measured by RT-PCR. Values are mean ± SD of three individual experiments and reported as the ratio of activin receptors to β-actin signals, the ratio of Smad to β-actin signals, and the ratio of p21CIP1/WAF1 to β-actin signals, respectively. aP < 0.05 and bP < 0.01 vs the control values.
DISCUSSION
Activin has several biological functions, including regulation of cell proliferation and inhibition of tumor cells. Activin inhibits cell growth in human prostate cancer LNCaP cells[6], human HepG2 hepatoma cells [19], and mouse B-cell hybridoma cells[20]. In this study, treatment with activin A (10-100 ng/mL) induced a significant decrease in the proliferation of SNU-16 cells in a dose-and time-dependent manner. In contrast, some human gastric cancer cell lines were relatively resistant to the growth inhibition induced by exogenous activin treatment. Interestingly, activin A (100 ng/mL) decreased the number of SNU-16 cells in a time-dependent manner. The mechanism of growth inhibition by activin in SNU-16 cells remains uncertain, but activin may act as a growth inhibitor in SNU-16 cells, and the subsequent loss of this autocrine growth inhibitory pathway may lead to the development of cancer.
Basal expressions of inhibin/activin subunits, activin receptors, Smads, and p21CIP1/WAF1 mRNAs in the human gastric cancer cell lines were demonstrated to have differential expression patterns for inhibin α, activin βA subunits, activin receptors, and Smads, as well as the presence of p21CIP1/WAF1 and the absence of activin βB subunits. Interestingly, significantly higher expressions of ActR IIA and IIB mRNAs were observed in SNU-16 cells when compared to other cells.
To determine the time-course of activin’s effects on mRNA levels of ActRs, Smads, and p21CIP1/WAF1, RT-PCR was performed using SNU-16 cells treated with activin A (100 ng/mL) for different time durations. Activin A decreased ActR IA, IB, and IIA mRNA levels whereas increased ActR IIB mRNA level in SNU-16 cells. Treatment with activin A (10-100 ng/mL) could not affect ActR IIA and IIB mRNA expression for 24 h in OVCAR-3 cells[21]. Activins act primarily through Smad2, possibly in partnership with Smad4, which forms heteromeric complexes with ligand-specific Smads after activation. In OVCAR-3 cells, no detectable change has been induced in Smad4 mRNA expression within 72 h of activin treatment, but the Smad2 mRNA level gradually increases and is significantly higher than that in the control at 72 h[22]. Activin–induced HepG2 liver cell apoptosis involves ActRs and Smad proteins. Overexpression of ActR IB and IIB or Smad2 and Smad4 stimulates apoptosis, whereas the dominant negative mutant forms of ActR IIB or Smad2 block activin-stimulated apoptosis. Signal transduction from the cell surface to the nuclei through Smad proteins is required for activin-induced cell death in liver cells[23]. In SNU-16 cells, no difference was observed in Smad2 mRNA levels but the Smad4 mRNA level increased for up to 48 h after activin treatment. Taken together, these results suggest that enhanced levels of ActR IIB and Smad4 mRNAs likely contribute to activin signal transduction. A novel activity of Smad7 is the inhibition of erythroid differentiation by blocking the intracellular activin signaling[14]. Smad7 expression can be induced by activin, and the overexpression of Smad7 suppresses activin-induced apoptosis in mouse B hybridomas[24]. In the present study, expression of Smad7 mRNA increased vigorously after 24 h of activin stimulation in SNU-16 cells, and then decreased at 48 h to a level significantly below the initial basal level. These results indicate that Smad7 acts as a selective blocker of activin and provides feedback regulation in the activin- signaling pathway. In human pituitary tumors with activin-inhibited proliferation, p21CIP1/WAF1 gene expression is up-regulated in a dose-dependent manner after 4 h[25]. The ectopic expression of mouse Smad7 in HS-72 cells suppresses the activin A-induced cell cycle arrest in G1 phase by abolishing the activin A-induced expression of p21CIP1/WAF1 and the hypophosphorylation of retinoblastoma protein[25]. Compared with control cells, expression of p21CIP1/WAF1 mRNA at 72 h after activin stimulation was higher in SNU-16 cells in this study, suggesting that the activin-induced inhibition of cell growth in SNU-16 cells is mediated through p21CIP1/WAF1 activation.
In conclusion, inhibin/activin subunits, activin receptors, Smads, and p21CIP1/WAF1 are expressed in human gastric cancer cell lines. Moreover, the inhibition of cell growth by activin is regulated by the negative feedback effect of Smad7 on the activin signaling pathway, and is mediated through p21CIP1/WAF1 activation in SNU-16 cells. Further investigation is needed to determine the molecular mechanisms of activin’s inhibitory effects on cell growth, apoptosis, and cancer progression.
Footnotes
Supported by the Research Fund 2003 from the Catholic University of Korea
S- Editor Wang J L- Editor Wang XL E- Editor Zhang Y
References
- 1.Ying SY. Inhibins and activins: chemical properties and biological activity. Proc Soc Exp Biol Med. 1987;186:253–264. doi: 10.3181/00379727-186-42611a. [DOI] [PubMed] [Google Scholar]
- 2.Ying SY. Inhibins, activins, and follistatins: gonadal proteins modulating the secretion of follicle-stimulating hormone. Endocr Rev. 1988;9:267–293. doi: 10.1210/edrv-9-2-267. [DOI] [PubMed] [Google Scholar]
- 3.Vale W, Hsueh A, Rivier C, Yu J. The inhibin/activin family of hormones and growth factors. In: Sporn MB, Roberts AB, eds , editors. Peptide Growth Factors and Their Receptors. New York: Springer-Verlag; 1990. pp. 211–248. [Google Scholar]
- 4.Mathews LS. Activin receptors and cellular signaling by the receptor serine kinase family. Endocr Rev. 1994;15:310–325. doi: 10.1210/edrv-15-3-310. [DOI] [PubMed] [Google Scholar]
- 5.Attisano L, Wrana JL, Montalvo E, Massagué J. Activation of signalling by the activin receptor complex. Mol Cell Biol. 1996;16:1066–1073. doi: 10.1128/mcb.16.3.1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dalkin AC, Gilrain JT, Bradshaw D, Myers CE. Activin inhibition of prostate cancer cell growth: selective actions on androgen-responsive LNCaP cells. Endocrinology. 1996;137:5230–5235. doi: 10.1210/endo.137.12.8940339. [DOI] [PubMed] [Google Scholar]
- 7.Ying S, Zhang Z, Huang G, Hsieh F. Inhibin activin subunits and activin receptors in human breast cell-lines and lactating rat mammary-glands. Int J Oncol. 1995;7:481–485. doi: 10.3892/ijo.7.3.481. [DOI] [PubMed] [Google Scholar]
- 8.Liu QY, Niranjan B, Gomes P, Gomm JJ, Davies D, Coombes RC, Buluwela L. Inhibitory effects of activin on the growth and morpholgenesis of primary and transformed mammary epithelial cells. Cancer Res. 1996;56:1155–1163. [PubMed] [Google Scholar]
- 9.Yingling JM, Wang XF, Bassing CH. Signaling by the transforming growth factor-beta receptors. Biochim Biophys Acta. 1995;1242:115–136. doi: 10.1016/0304-419x(95)00007-2. [DOI] [PubMed] [Google Scholar]
- 10.Heldin CH, Miyazono K, ten Dijke P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature. 1997;390:465–471. doi: 10.1038/37284. [DOI] [PubMed] [Google Scholar]
- 11.Hayashi H, Abdollah S, Qiu Y, Cai J, Xu YY, Grinnell BW, Richardson MA, Topper JN, Gimbrone MA Jr, Wrana JL, et al. The MAD-related protein Smad7 associates with the TGFbeta receptor and functions as an antagonist of TGFbeta signaling. Cell. 1997;89:1165–1173. doi: 10.1016/s0092-8674(00)80303-7. [DOI] [PubMed] [Google Scholar]
- 12.Imamura T, Takase M, Nishihara A, Oeda E, Hanai J, Kawabata M, Miyazono K. Smad6 inhibits signalling by the TGF-beta superfamily. Nature. 1997;389:622–626. doi: 10.1038/39355. [DOI] [PubMed] [Google Scholar]
- 13.Nakao A, Afrakhte M, Morén A, Nakayama T, Christian JL, Heuchel R, Itoh S, Kawabata M, Heldin NE, Heldin CH, et al. Identification of Smad7, a TGFbeta-inducible antagonist of TGF-beta signalling. Nature. 1997;389:631–635. doi: 10.1038/39369. [DOI] [PubMed] [Google Scholar]
- 14.Kitamura K, Aota Si, Sakamoto R, Yoshikawa SI, Okazaki K. Smad7 selectively interferes with different pathways of activin signaling and inhibits erythroid leukemia cell differentiation. Blood. 2000;95:3371–3379. [PubMed] [Google Scholar]
- 15.Kanamaru C, Yasuda H, Fujita T. Involvement of Smad proteins in TGF-beta and activin A-induced apoptosis and growth inhibition of liver cells. Hepatol Res. 2002;23:211–219. doi: 10.1016/s1386-6346(01)00175-9. [DOI] [PubMed] [Google Scholar]
- 16.el-Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, Lin D, Mercer WE, Kinzler KW, Vogelstein B. WAF1, a potential mediator of p53 tumor suppression. Cell. 1993;75:817–825. doi: 10.1016/0092-8674(93)90500-p. [DOI] [PubMed] [Google Scholar]
- 17.Skapek SX, Rhee J, Spicer DB, Lassar AB. Inhibition of myogenic differentiation in proliferating myoblasts by cyclin D1-dependent kinase. Science. 1995;267:1022–1024. doi: 10.1126/science.7863328. [DOI] [PubMed] [Google Scholar]
- 18.Yang ZY, Perkins ND, Ohno T, Nabel EG, Nabel GJ. The p21 cyclin-dependent kinase inhibitor suppresses tumorigenicity in vivo. Nat Med. 1995;1:1052–1056. doi: 10.1038/nm1095-1052. [DOI] [PubMed] [Google Scholar]
- 19.Zauberman A, Oren M, Zipori D. Involvement of p21(WAF1/Cip1), CDK4 and Rb in activin A mediated signaling leading to hepatoma cell growth inhibition. Oncogene. 1997;15:1705–1711. doi: 10.1038/sj.onc.1201348. [DOI] [PubMed] [Google Scholar]
- 20.Hashimoto O, Yamato K, Koseki T, Ohguchi M, Ishisaki A, Shoji H, Nakamura T, Hayashi Y, Sugino H, Nishihara T. The role of activin type I receptors in activin A-induced growth arrest and apoptosis in mouse B-cell hybridoma cells. Cell Signal. 1998;10:743–749. doi: 10.1016/s0898-6568(98)00021-7. [DOI] [PubMed] [Google Scholar]
- 21.Choi KC, Kang SK, Nathwani PS, Cheng KW, Auersperg N, Leung PC. Differential expression of activin/inhibin subunit and activin receptor mRNAs in normal and neoplastic ovarian surface epithelium (OSE) Mol Cell Endocrinol. 2001;174:99–110. doi: 10.1016/s0303-7207(00)00447-0. [DOI] [PubMed] [Google Scholar]
- 22.Ito I, Minegishi T, Fukuda J, Shinozaki H, Auersperg N, Leung PC. Presence of activin signal transduction in normal ovarian cells and epithelial ovarian carcinoma. Br J Cancer. 2000;82:1415–1420. doi: 10.1054/bjoc.1999.1127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chen W, Woodruff TK, Mayo KE. Activin A-induced HepG2 liver cell apoptosis: involvement of activin receptors and smad proteins. Endocrinology. 2000;141:1263–1272. doi: 10.1210/endo.141.3.7361. [DOI] [PubMed] [Google Scholar]
- 24.Ishisaki A, Yamato K, Nakao A, Nonaka K, Ohguchi M, ten Dijke P, Nishihara T. Smad7 is an activin-inducible inhibitor of activin-induced growth arrest and apoptosis in mouse B cells. J Biol Chem. 1998;273:24293–24296. doi: 10.1074/jbc.273.38.24293. [DOI] [PubMed] [Google Scholar]
- 25.Danila DC, Inder WJ, Zhang X, Alexander JM, Swearingen B, Hedley-Whyte ET, Klibanski A. Activin effects on neoplastic proliferation of human pituitary tumors. J Clin Endocrinol Metab. 2000;85:1009–1015. doi: 10.1210/jcem.85.3.6473. [DOI] [PubMed] [Google Scholar]